Abstract
Much is unknown about the positive effect of plant growth regulators, such as brassinosteroids, on high-temperature stress tolerance in pepper plants. This study aimed to reveal the effect of exogenous brassinosteroids (BRs) on high-temperature-stressed bell peppers by foliar application. BR treatments (1 µM), in both bulk (Br) and nano-encapsulated (N-Br) forms, were applied to pepper plants subjected to high-temperature stress (35 °C). The results indicated an increase in plant biomass, number of fruits, and relative water content and a significantly lower flower abscission in response to Br and N-Br under high temperatures. Br and N-Br applications at high temperatures resulted in the lowest leaf water potential. Br and N-Br, especially N-Br, were more effective than Br in the upregulation of the antioxidant enzyme activity, such as catalase (CAT), superoxide dismutase (SOD), peroxidase (POD), and the overall antioxidant capacity of heat-stressed plants. MDA and electrolyte leakage significantly declined as a result of BR application and the proline content of the leaves was significantly higher in Br and N-Br treatments at high temperatures. Further analysis of the data pointed to SOD, DPPH, proline, RWC, and leaf water potential in pepper leaves as the most affected traits in response to brassinosteroid application under high temperature. Glucose and fructose levels also increased under high temperature, and only N-Br administration showed a significant effect on reducing the sugar levels. At high temperatures, the ratio of saturated to unsaturated fatty acids was greatest when neither Br nor N-Br was present. N-Br could reduce this ratio effectively. Conclusively, the overall performance of bell pepper improved in response to both types of BR application with no significant discrimination being found to prioritize the encapsulated form of BR application.
Keywords:
antioxidant; catalase; fatty acid; fructose; glucose; peroxidase; superoxide dismutase; thermal stress 1. Introduction
Because of all the vitamins and antioxidants in its fruit, Capsicum annum L. is one of the most common and well-known vegetables of the Solanaceae plant family. Capsicum peppers are cultivated in a relatively warm climate, but their growth and development are sensitive to rising stresses including heat, salinity, and drought [1]. Plant tissues undergo molecular, cellular, and physiological changes in response to abiotic stressors. In recent years, high-temperature stress has become more frequent as one of climate change’s most serious non-biological impacts [2]. The potential damages associated with high temperatures include leaf burning, organ shedding, ageing, diminished plant development, and fruit damage, with a subsequent reduction in yield and productivity [3]. Numerous studies have been dedicated to the observation of the influence of high-temperature stress on fruit growth and development. During flowering and fruiting, high temperatures inhibit the growth and productivity of fruits, particularly in the case of green peppers [4]. It was earlier reported that bell peppers continued to generate flowers at high temperatures (38 °C) and that the blossoms did not fall off until they opened. However, post pollination, high-temperature stress inhibited the fertilization and fruit set [5]. High temperatures decrease the cell’s relative humidity, hence reducing cell size and plant growth. A reduced production of assimilates as a result of decreased photosynthesis and higher respiration contributes to the relative decrease in the growth rate of plants under high-temperature stress [6]. The plant resists the damaging effects of stress at the cellular level by enhancing metabolism and managing osmotic potential via the accumulation of organic and mineral substances in cells to regulate the turgor pressure [7,8]. Diverse abiotic stressors induce the overproduction of reactive oxygen species in plants, damaging proteins, lipids, carbohydrates, and deoxyribonucleic acid (DNA). Lipid peroxidation results in the formation of stress marker metabolites such as malondialdehyde (MDA) and other unsaturated aldehydes. To overcome the impacts of stress, plants employ specific mechanisms, like activating enzymatic and non-enzymatic pathways [6]. A range of biotic and abiotic stressors are induced by climate change with potential disturbing effects on the normal physiological and molecular processes of plants [9,10,11]. In this setting, phytohormones and other metabolites play an important role in the regulation of a wide range of physiological and/or biochemical responses to enhance plant stress tolerance [12,13,14,15].
Brassinosteroids (BRs) are unique phytohormones that are evolving due to their various roles in plant development and growth [16,17]. Brassinosteroids are considered the sixth family of plant hormones with pleiotropic properties [18]. The use of exogenous brassinosteroids increases plant resistance to a variety of stressors [19]. The interplay of BRs with other plant hormones was detected in several physiological and growth processes in plants [20].
The exogenous application of BRs can alter the activity of enzymatic and the synthesis of non-enzymatic antioxidants, thus decreasing lipid peroxidation and ROS formation that can contribute to plant resistance to environmental challenges [17,21].
BR application on salinity-stressed peppers enhanced plant performance by increasing antioxidant enzyme activity, proline content, total anthocyanin, and mineral concentrations that are believed to be orchestrated in complex physiological and molecular networks upon salt stress in plants [22,23]. An elevated antioxidant enzyme activity, with a potential contributary role in preventing lipid peroxidation, and the improved chlorophyll content and photosynthesis, which resulted in a better growth and productivity of heat-stressed tomatoes and soybeans, were claimed to be aided by BR application [24,25]. Brassinosteroids also play a role in overcoming low-temperature stress in Arabidopsis [26], peppers [27], and grapes [28]. Utilizing encapsulated or nanofertilizers or phytohormones can be a useful strategy toward sustainable agriculture [29,30]. The encapsulated formulations regulate the duration of release of elements and the rate based on the plant’s nutritional requirements, hence contributing to the maximum absorption of nutrients and leaching prevention, resulting in higher yield and productivity [31,32].
Due to increased excitability, solubility, and reactivity, nano-encapsulated substances have a greater impact on plant biomass than non-encapsulated substances [33]. The tiny size of nano-capsules may facilitate the absorption of these particles and the delivery of the active ingredients more efficiently. Nano-encapsulated fertilizers can also permeate plant leaves more effectively and extensively [34]. They can be absorbed through the cuticle and pores and penetrate the vascular bundles, mesophilic cells, and apoplasts in the shortest time and with a higher bioavailability rate [35]. There are several examples of nano-encapsulated biostimulants and fertilizers with positive outcomes upon application in crop plants. For example, basil plants produced more phenol, flavonoids, and antioxidant compounds following the foliar application of nano-zinc and copper fertilizers when compared to their conventional application [36].
This investigation seeks to comprehend the morphological and physiological reactions of bell pepper plants to high temperatures, and to find out the potential applicability of bulk and encapsulated brassinosteroids for enhancing heat stress endurance.
2. Materials and Methods
2.1. Experimental Design and Plantation
Aristotle’s cultivar of Capsicum annum was grown at a research greenhouse in Isfahan University of Technology, Isfahan, Iran. This research was developed as a factorial experiment with 3 biological replicates in a completely randomized design (RCD). Pepper seedlings (in the three-leaves stage) were obtained from Ghaem Isfahan Seedling Production Company and transplanted in 5 L pots filled with cocopeat/perlite (1:1) to grow under a temperature regime of 25 ± 3 °C (day—16 h) to 18 ± 3 °C (night—8 h) and a relative air humidity of 65–68% until the appearance of the 6th true leaves. The plants received half of the concentration of nutritional solution comprising N = 116, P = 21, K = 82, S = 28, Mg = 21, Ca = 125, Fe = 6.8, Mn = 1.97, B = 0.70, Zn = 0.25, Cu = 0.07, B = 0.70, and Mo = 0.05 mg L−1 of tab water for the first 2 weeks and the full dosage of the same solution until the end of the experiment [37]. Right after the emergence of the initial flower buds, the bell pepper plants were sprayed weekly with 1 µM bulk brassinosteroids (Br), 1 µM nano-encapsulated brassinosteroids of (N-Br), and water serving as the control. The applied concentration (1 µM) was selected after an intensive literature review to find out the most commonly applied quantity of BRs in studies related to high-temperature stress and plants. The high-temperature stress treatment condition was achieved by stopping and/or lowering the wet-pad ventilator in the treatment section of the greenhouse, when a constant 35 °C temperature for 6 h was provided for the heat treatment groups every day. The vegetative stage is identified after the seedling has fully established itself and before the first flower bud emerges, whereas the reproductive stage is determined when the first flower appears on 50% of the plants. Plant samples were collected on the 56th day of treatments and subjected to different measurements or stored properly for analysis of the required indices.
2.2. Brassinosteroid Production in Nano-Capsules and Their Physicochemical Properties
Nano-encapsulated brassinosteroids (N-Brs) were created by following the technique of Mozafari et al. [38] in the department of food science (University of Isfahan, Iran). Epibrassinolide (Sigma-Aldrich, St. Louis, MO, USA) and soy lecithin (L-a-Lecithin, Sigma-Aldrich, St. Louis, MO, USA) (4% concentration) were utilized for nano-encapsulation. The manufacturing of nano-capsules was based on ultrasound treatment in a single step without the use of organic solvents. The lecithin solution was rotary-mixed with BRs for 3 h at 30 °C in a nitrogen environment at 600 rpm on a magnetic stirrer. The obtained solution was homogenized for six minutes under ultrasonic instrument (HUP-180) at 180 Watt to unify the size of the carriers. The resulting encapsulation solution was then put at ambient temperature for one hour and then refrigerated at 4 °C to stabilize [39].
A scanning electron microscope (FEI Quanta FEG 450, Hillsborough, OR, USA) was utilized to examine the microstructure of the nano-encapsulated BRs, the surface topography, the size, and the shape of the generated lecithin nanoparticles [40]. A zeta-size analyzer with a laser beam was used to assess the particles’ size and zeta potential (as an indication of particle surface charge). The average particle size was calculated using a dynamic light scattering particle analyzer instrument (HORIBA-SZ100, Kyoto, Japan) at room temperature.
A representative microscopic image of the lecithin-coated nano-encapsulated brassinosteroid particles after freeze-drying is presented in Supplementary Figure S1. Nano-capsules were irregular with uneven surfaces projecting a teeth-like structure and cavities after freeze-drying. The majority of nano-capsules created by this process were smaller than 100 nanometers (Figure S2) and the sample’s zeta potential ranged from −82.2 mV to −77.1 mV, with a mean of −79.1 mV and a standard deviation of ±2.71 mV (Figure S3).
2.3. Growth and Physiological Parameters
The fresh weight (FW) and dry weight (DW) of shoot and root tissues were determined at harvest and after 48 h of oven-drying at 70 °C, respectively. In addition, the number of leaves, flowers, fruit, and aborted flowers was also recorded for statistical analysis.
The leaf water potential (LWP) was measured in four leaves of different biological replicates of each treatment at midday (11:30 am–12:30 pm) using a pressure chamber (3115, Soil Moisture Equipment Corp, Santa Barbara, CA, USA) [41].
Between 8:00 and 9:00 am, the chlorophyll fluorescence level was determined with a fluorometer (RS232, Handy PEA, King’s Lynn, UK). The leaves were incubated in the dark for 30 min before quantifying the maximal photochemical quenching capacity of photosystem II [42].
The relative water content (RWC) of the pepper plant leaves was calculated following the method of Dhopte and Manuel [43] and Equation (1). In brief, leaf discs of 7 mm in diameter were sampled from different biological replicates of different treatments and their initial weight (FW) was determined before being hydrated in distilled water for 48 h in darkness (TW) to saturate and reach a constant weight. The labelled leaf disc samples were then dried at 105 °C to obtain a constant dry weight (DW) and to calculate the RWC using Equation (1):
RWC% = (FW − DW)/(TW − DW) × 100
The electrolyte leakage (EL) rate of the leaf samples was determined by applying the method of Lutts [44]. The initial electrical conductivity (EC1) was assessed in three discs of 1 cm in diameter from fully expanded leaves, which were washed with and immersed in deionized water after 24 h of incubation at room temperature, by a conductivity metre apparatus (Hanna, HI98130, Kehl am Rhein, Germany). The final electrical conductivity (EC2) of the same samples was determined after boiling the solutions in a water bath at 120 °C for 20 min and subsequent cooling to 25 °C. Electrolyte leakage (EL) was then calculated using the following equation (Equation (2)).
where EL is electrolyte leakage, EC1 is initial electrical conductivity, and EC2 is final electrical conductivity.
EL = EC1/EC2 × 100
2.4. Determination of Proline, Antioxidants, and Lipid Peroxidation
The proline content in leaf samples was assessed based on the Bates assay [45]. Leaves of different biological samples (0.1 g) were homogenized in 10 mL of aqueous 3% sulfosalicylic acid and centrifuged to obtain a clear supernatant. This supernatant (2 mL) was then mixed with ninhydrin and glacial acetic acid which was then incubated at 100 °C for 60 min. The reaction solution was then cooled in an ice bath and gently reacted to 4 mL of toluene for 15–20 s. The absorbance was measured at 520 nm, and the proline content is expressed in µmol L-proline equivalent g−1 FW.
The antioxidant activity of methanolic extract of pepper leaves (3 g leaf samples in 5 mL of CH₃OH) against DPPH was determined according to Koleva et al. [46]. The clear supernatant (1.4 mL) was reacted with 0.6 mL of 130 μM 2,2-diphenyl-1-picrylhydrazyl (DPPH) ethanolic solution. The absorbance values were recorded at 515 nm in a spectrophotometer (UV 160A-Shimadzu, Kyoto, Japan). After 30 min, against the same ratio of ethanol and methanol as a blank, the results are presented as a percentage % of DPPH scavenging.
Total soluble protein was obtained from leaf samples after homogenization in a 1% polyvinylpyrrolidone (PVP)- and EDTA (4 mM)-containing potassium phosphate buffer (pH 6.8, 100 mM) to assay the antioxidant enzyme activity. The catalase (CAT) enzyme activity was measured spectrophotometrically at 240 nm as described by Tabatabaei and Ehsanzadeh [47]. The superoxide dismutase (SOD) enzyme activity was determined following the method reported by Khoshbakht et al. [48] and using the recorded absorbance values at 560 nm. Ascorbate peroxidase (APX) enzymatic activity was established based on the absorbance variation at 290 nm and is expressed as U mg−1 protein [49].
Lipid peroxidation rate was evaluated by measuring the quantity of MDA in reaction with thiobarbituric acid as outlined by Jiang and Zhang [50]. The absorbance of the supernatant from the reaction solution was monitored at 532 and 600 nm with the spectrophotometer and the MDA content was estimated by its calculated molar extinction coefficient of 155 Mm–1 cm–1 and is shown as nmol g–1 (FW).
2.5. Glucose and Fructose Content
Leaf samples were homogenized in ethanol (80%) by grinding and ultra-sonication (10 min) and then centrifuged at 4 °C (3000 rpm) for 10 min. The ultra-membrane-filtered supernatant was analyzed in an HPLC system coupled with a refractive index detector (Shimadzu, Kyoto, Japan). The applied HPLC column was an SCR-101 N (30 cm and 9.7 mm) with an SCR (N)-fitted guard column (5 cm 4 mm). The mobile phase was deionized water at a 0.7 mL min−1 flow rate at 60 °C [51].
2.6. Total Protein and Fatty Acids Content
The Bradford assay was used to measure the total protein concentration [52]. Fresh leaf samples (1 g) were homogenized in sodium phosphate buffer (4 mL, pH = 7.2) and cold (4 °C)-centrifuged for 10 min at 1000 rpm. The absorbance of the reaction solution was recorded at 595 nm by a spectrophotometer. The total protein content was then calculated using a calibration curve created by different concentrations of bovine serum albumin (BSA) as standard.
The fatty acid content and composition of the pepper leaves were determined according to the method of Goli et al. [53] using a gas chromatography system (Agilent 6890N, Santa Clara, CA, USA) coupled with a flame ionization detector (FID) and an HP-88 capillary column (100 m, 250 µm). Nitrogen was used as the carrier gas with a flow rate of 1.1 mL min−1. The oven temperature was set to rise by 5 °C per min starting from 150 °C, a hold of 1 min, an increase up to 190 °C, a hold of 2 min, and finally an increase up to 240 °C with a final hold of 8 min. The temperatures of the injector and detector were 150 °C and 250 °C, respectively.
2.7. Statistical Analysis
All collected data were subjected to statistical analysis using Statistix 8 (Tallahassee, FL, USA) by a two-way analysis of variance, and the mean values of each treatment were compared using the least significant difference (LSD) test at the 95% (p ≤ 0.05) level of probability, with the results presented as the means ± standard deviation of at least three biological replicates. The principal component analysis (PCA) chart was created using Statgraphics Centurion, Version XVI.
3. Results
3.1. Growth and Yield Parameters
The ANOVA and the main effect of heat stress and different BR applications on all measured parameters are presented in Tables S1–S8 of the Supplementary Materials.
The shoot fresh weight (FW) and dry weight (DW) of the pepper plants significantly (p < 0.05) decreased under the heat stress (Figure 1A,B). The application of Br and N-Br significantly improved the shoot dry weight under the high temperature applied. The heat stress slightly affected the root fresh and dry weight but not significantly (p < 0.05). N-Br application decreased the fresh and dry weight of the root under both control and high-temperature conditions (Figure 1C,D).
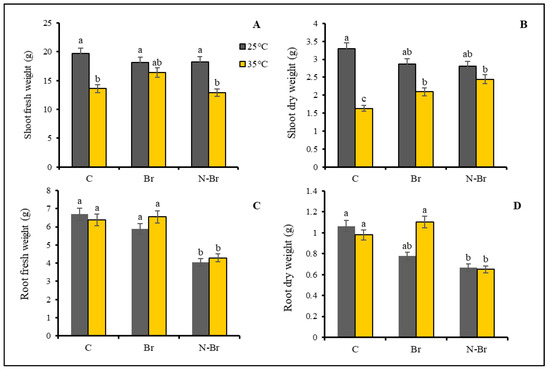
Figure 1.
The interaction effects of different sources of Br application and high temperature (35 °C) on growth parameters of bell pepper. Significant differences are noted by different letters on each column at (p < 0.05) according to the least significant difference (LSD) test. C: No-brassinosteroid control, Br: bulk brassinosteroid (1 µM), N-Br: nano-encapsulated brassinosteroid (1 µM).
The applied heat stress did not have a significant effect on the number of leaves and flowers (Figure 2A,B). The brassinosteroid treatments did not have any significant effect on the number of leaves under optimum temperature but the highest number of leaves per plant was recorded in heat-stressed plants after the bulk Br treatment (Figure 2A). The application of both brassinosteroid and nano-encapsulated brassinosteroid significantly increased the number of flowers in pepper plants grown under optimum temperature, while no significant changes were observed in the number of flowers in heat-stressed peppers (Figure 2B). The applied high temperature significantly increased the rate of flower abscission and both Br and N-Br treatments could prevent the flowers falling off (Figure 2C).
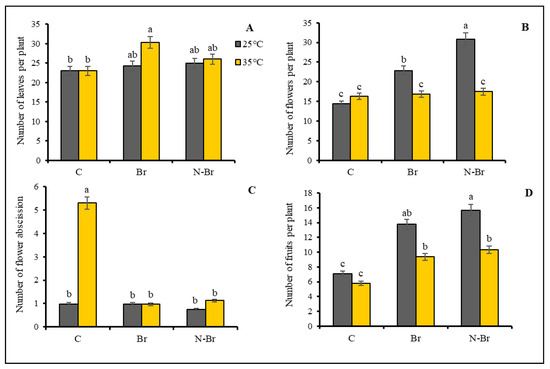
Figure 2.
The interaction effects of different sources of Br application and high temperature (35 °C) on flower abscission and number of flowers, fruit, and leaves of bell peppers. Significant differences are noted by different letters on each column at (p < 0.05) according to the least significant difference (LSD) test. C: No-brassinosteroid control, Br: bulk brassinosteroid (1 µM), N-Br: nano-encapsulated brassinosteroid (1 µM).
The number of mature fruits per plant significantly increased (p < 0.05) when Br and N-Br were administered under both optimum and high-temperature stress conditions (Figure 2D). However, the brassinosteroid type (bulk or encapsulated) did not show any significant differences in their effect on the number of fruits under the high-temperature stress (Figure 2D).
3.2. Physiological Parameters
The leaf water potential (LWP) increased (smaller minus) in response to Br and N-Br but decreased (larger minus) in response to Br and high temperatures. N-Br application produced the greatest LWP, but Br and N-Br application at high temperatures resulted in the lowest LWP (Figure 3A).
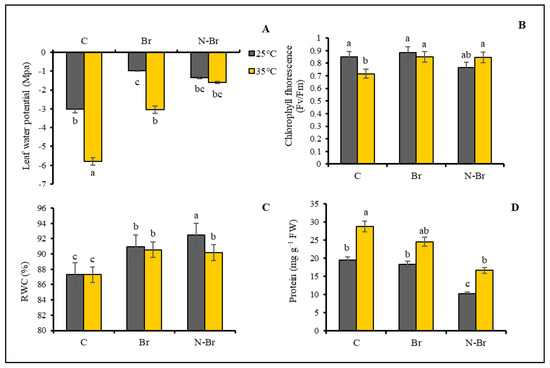
Figure 3.
The interaction effects of different sources of Br application and high temperature (35 °C) on physiological parameters of bell peppers. Significant differences are noted by different letters on each column at (p < 0.05) according to the least significant difference (LSD) test. C: No-brassinosteroid control, Br: bulk brassinosteroid (1 µM), N-Br: nano-encapsulated brassinosteroid (1 µM), RWC: relative water content.
A significantly lower level of leaf chlorophyll fluorescence after high-temperature treatment was compensated by the Br and N-Br treatments (Figure 3B). As presented in Figure 3C, the Br and N-Br application increased the relative water content (RWC) of pepper leaves significantly under both normal and high temperatures (Figure 3C).
The total protein content of the pepper’s leaves was significantly increased under the high-temperature stress and significantly decreased only when brassinosteroid was applied in its nano-encapsulated form (Figure 3D).
The catalase enzyme (CAT) activity was significantly increased (p < 0.05) in pepper plants under heat stress (Figure 4A). The application of Br and N-Br also resulted in a remarkable upregulation of CAT activity under both optimum and high-temperature conditions compared with their non-Br-treated counterparts. On the other hand, the POD and SOD activities were not significantly different between the control and heat-stressed plants (Figure 4B,C). The same upregulation was observed in the case of POD and SOD activity when the plants were treated with 1 µM brassinosteroids and the highest POD activities were recorded in N-Br-treated plants, while the highest SOD activities were found in both Br- and N-Br-treated plants under the high-temperature condition (Figure 4B,C). In the absence of brassinosteroid treatment, the antioxidant capacity (percentage of DPPH inhibition) of pepper plants was found to be slightly but not significantly higher after heat stress (Figure 4D). Moreover, the Br and N-Br treatment reduced the overall non-enzymatic antioxidant capacity of the plants under the optimum temperature. The highest antioxidant capacity was recorded for the heat-stressed plants after N-Br treatment (Figure 4D).
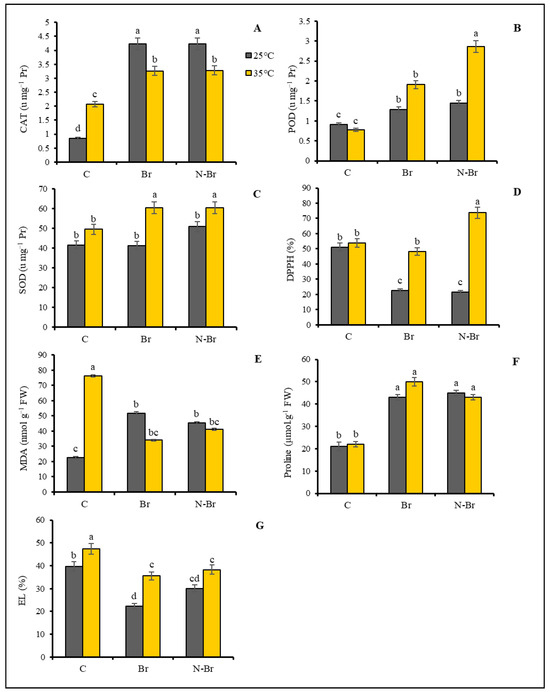
Figure 4.
The interaction effects of different sources of Br application and high temperature (35 °C) on some stress indices of bell peppers. Significant differences are noted by different letters on each column at (p < 0.05) according to the least significant difference (LSD) test. C: No-brassinosteroid control, Br: bulk brassinosteroid (1 µM), N-Br: nano-encapsulated brassinosteroid (1 µM), CAT: catalase enzyme activity, POD: peroxidase enzymes activity, SOD: superoxide dismutase enzymes activity, DPPH: 2,2-diphenyl-1-picrylhydrazyl, MDA: malondialdehyde, EL: electrolyte leakage.
The maximum MDA level was quantified in the heat-stressed plants without the Br or N-Br treatments, while the application of both Br and N-Br significantly reduced the MDA level under the high-temperature stress (Figure 4E). The opposite trend was observed for the MDA level after Br or N-Br treatments of the control plants. The high-temperature stress resulted in a slightly higher (non-significantly) proline content in pepper leaves (Figure 4F). Both Br- and N-Br-treated plants showed increased proline levels under optimum and high-temperature conditions (Figure 4F). Heat stress significantly increased the EL rate in the leaves of pepper plants and the application of both Br and N-Br significantly reduced the electrolyte leakage (%) of both stressed and non-stressed plants when compared to the non-BR treatments (Figure 4G).
3.3. Sugars and Fatty Acids
The applied high-temperature stress significantly increased (p < 0.05) the glucose and fructose content of the bell pepper leaves (Figure 5A,B). The brassinosteroid application had no effect on the glucose content of the leaves under optimum temperature, but the nano-encapsulated Br (N-Br) significantly reduced the glucose content of the heat-stressed peppers (Figure 5A).
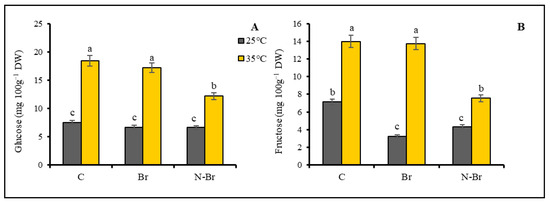
Figure 5.
The interaction effects of different sources of Br application and high temperature (35 °C) on leaf glucose (A) and fructose (B) content of bell peppers. Significant differences are noted by different letters on each column at (p < 0.05) according to the least significant difference (LSD) test. Br: bulk brassinosteroid (1 µM), N-Br: nano-encapsulated brassinosteroid (1 µM).
Both forms of BRs (Br and N-Br) significantly reduced the fructose content of the leaves under the control temperature, but under the high-temperature treatment, only the N-Br application resulted in a decreased fructose content of the leaves (Figure 5B).
Bell peppers grown under high temperature (35 °C) and no Br treatment showed a higher content of both saturated and unsaturated fatty acids, as well as a higher ratio of saturated to unsaturated fatty acids (Table 1). N-Br could effectively reduce this ratio. Palmitic acid was the most abundant of fatty acids, followed by linoleic acid. Oleic acid was lowest in all treatments (Table 1).

Table 1.
Interaction effects of brassinosteroid treatment and high temperature on the content and composition of saturated and unsaturated fatty acids (mg 100 g–1 DW) in bell pepper leaves.
The principal component analysis (PCA) revealed that Br and N-Br applications resulted in the highest relative water content (RWC), fatty acid content, and antioxidant enzyme activity, when high-temperature stress was applied (Figure 6A). The lowest RWC and fatty acid content (treatment number 21) was associated with the non-brassinosteroid control plants under high-temperature treatments. N-Br-treated plants appeared to have a higher RWC, fatty acid content, and antioxidant enzyme activity and a lower flower abscission and high water potential (more minus) under high-temperature stress (Figure 6A). The spider diagram revealed that when high temperatures were applied, SOD, DPPH, proline, RWC, and leaf water potential were the parameters that changed most. These are the most crucial parameters in pepper plants that are affected due to the applied high temperatures, as indicated by yellow lines (C-35 °C) in Figure 6B.
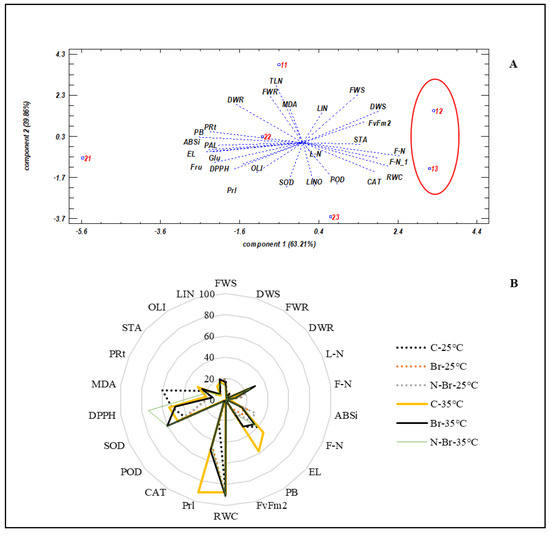
Figure 6.
The principal component analysis (A) and spider diagram (B) of the interaction between the various sources of Br application and high temperature on bell pepper plants for all of the assessed parameters. 11: C-25 °C, 12: Br-25 °C, 13: N-Br-25 °C, 21: C-35 °C, 22: Br-35 °C, 23: N-Br-35 °C. FWS: shoot fresh weight, DWS: shoot dry weight, FWR: root fresh weight, DWR: root dry weight, L-N: number of leaves per plant, F-N: number of flowers per plant, ABSi: flower abscission per plant, F-N: number of fruits per plant, EL: electrolyte leakage, PB: leaf water potential, FvFm: chlorophyll fluorescence, RWC: relative water content, Prl: proline, CAT: catalase, POD: peroxidase, PRt: protein, MDA: malondialdehyde, DPPH: DPPH, SOD: superoxide dismutase, PAL: palmitic acid, LIN: linolelaidic acid, STA: stearic acid, OLI: oleic acid, LINO: linoleic acid, TLN: α-linoleic acid, Glu: glucose, Fru: fructose.
4. Discussion
4.1. The Interacting Effect of Br and N-Br on Morphological Characteristics of Bell Pepper
Heat stress impairs the function of the photosynthetic apparatus and prevents photosynthesis in plant cells [24,54]. The exogenous application of brassinosteroids (BRs) has been proven to significantly enhance the photosynthetic activities of several plant species under suboptimum temperatures and abiotic stresses [55,56,57], resulting in increased growth and yield-associated indices, as also demonstrated in our current study on pepper plants.
Our results confirm that both forms of bulk and nano-encapsulated brassinosteroids (Br and N-Br) at a 1 µM concentration promote the growth of pepper plants under heat stress as manifested in the shoot and root biomass and the number of leaves, flowers, and fruits (Figure 1 and Figure 2). Wu et al. observed similar results in kiwi plants. The exogenous Br application improved kiwifruit root elongation, lateral root development that resulted in a more efficient water and nutrient utilization, fruit yield, and quality, suggesting a correlation between the higher root biomass and the overall better growth [58]. The induced cell elongation and root growth are believed to be associated with differential gene expression in gibberellic acid metabolism in rice and in response to Br application [59]. The correlation between high temperature and flower drop has been reported earlier [60,61]. We also found a significantly higher rate of flower abscission in our pepper plants under the high-temperature stress (Figure 2).
The regulation of flower and fruitlet abscission has been linked to the role of several plant growth regulators including brassinosteroids in the ethylene-induced shedding of plant organs [62]. According to Ma et al. [63], the exogenous application of brassinosteroid resulted in a reduced fruitlet abscission of litchi that was correlated with the lower ethylene content which itself was associated with the downregulation of the ethylene biosynthetic genes. The authors could not conclude the concentration-dependent effect of ethylene on the abscission rate, but another study on Arabidopsis revealed that the ethylene production was increased when high concentrations of BRs (>0.5 μM) were applied [64]. According to our results, the application of 1 μM brassinosteroids in both bulk and encapsulated forms prevented the flowers’ abscission under high temperature, which could be due to the lower ethylene production that needs to be proved by ethylene quantification in future studies. Similar to our findings, the brassinosteroid (BB16) application under optimum conditions resulted in higher fruit numbers and fruit weights of strawberry plants [65].
4.2. The Interaction Effect of Br and N-Br on Physiological Characteristics of Bell Pepper
Chlorophyll fluorescence is reported to be altered in plants upon high-temperature stress, and higher temperatures usually result in decreased chlorophyll fluorescence levels [66]. The potentially positive role of BRs on photosynthetic efficiency in several plants has been reported earlier [67]. We also observed a significantly higher chlorophyll fluorescence level in the leaves of our heat-stressed pepper plants after both Br and N-Br treatments (Figure 3).
According to earlier reports, BRs cooperate with other external stimuli to regulate the stomatal aperture, a crucial factor under dehydration stress [68,69], resulting in a more efficiently controlled RWC of the plants, which was also evident in our findings.
BRs were found to be associated with upregulation of the expression of numerous stress-related genes, as revealed by a transcriptomic analysis [70].
The foliar application of BR (1 μM) on Lycopersicon lycopersicum L. resulted in a significantly enhanced productivity of salinity-stressed tomato plants after a decline in EL, H2O2, superoxide, and lipid peroxidation, and a subsequent increase in photosynthetic activity (total chlorophyll, and carotenoids) and the proline level [71]. In line with our findings, other studies also revealed the positive effects of BRs on plants under heat stress [72,73,74].
4.3. The Interacting Effect of Br and N-Br on Antioxidant Enzyme Activities, MDA, and Proline Content of Bell Pepper
Numerous studies have established that Br application is enhancing photosynthetic efficiency, decreasing transpiration rate, altering nutrient metabolism, and modulating Rubisco activity in the context of abiotic stresses [75]. Our study demonstrated that heat stress led to a higher lipid peroxidation in pepper. However, membrane lipid peroxidation was significantly reduced in heat-stressed plants supplemented with Br, as compared to their heat-treated counterparts without Br treatment.
Moreover, heat stress increased the pepper plant’s ROS production and lipid peroxidation similar to earlier reports on tomatoes [24,76]. According to Ogweno et al. [24], the higher content of small-molecule antioxidants (as expressed in a DPPH assay) in heat-stressed leaves correlated with better growth. We also found a higher antioxidant capacity in our N-Br-treated peppers (Figure 4D). Additionally, BRs were reported to enhance the activity of antioxidant and Calvin cycle enzymes, thereby protecting the photosynthetic apparatus from suboptimum-temperature stress and consequently decreasing ROS accumulation and enhancing the protection of photosynthetic machinery [77]. It is also reported that the foliar application of 200 nM brassinosteroids to Brassica rapa L. for five days increased antioxidant activity and reduced the heat-stress-associated physiological damage [78].
According to Ahmed et al. [3], BR-treated plants exhibited a higher quantum yield of the PSII and rubisco activities, and the overexpression of their genes resulted in enhanced photosystem I and II functions, as well as an optimal Fv/Fm ratio in pepper. We also noted that the Fv/Fm ratio significantly decreased during heat stress in control (non-BR) plants, indicating damage to photosystem II (PSII). This finding aligns with the results of Arief et al. [79], reporting a similar decline in the Fv/Fm ratio due to applied abiotic stresses such as heat and draught, attributing it to PSII damage. Furthermore, Bhattarai et al. [80] found that under heat stress, chlorophyll fluorescence decreased as plant injury increased, suggesting that plants exhibiting lower heat injury indices maintained higher chlorophyll fluorescence levels. In contrast, our results showed that the Br- and N-Br-treated plants exhibited higher Fv/Fm values compared to the control, indicating that Br effectively mitigated the adverse effects of heat stress on PSII. This is consistent with the findings of Thussagunpanit et al. [81], who demonstrated that EBR applications reduced PSII overexcitation under heat stress conditions in rice. Thus, the protective role of Br appears to enhance the resilience of chlorophyll fluorescence in stressed plants, which could explain the differential responses observed between the control and treated groups.
4.4. The Interacting Effect of Br and N-Br on Glucose and Fructose Content of Bell Pepper
Several reports have been published on the effect of temperature on vegetable fruits and shoot dry matter and glucose levels [82,83]. The heat exposure of grapevines increased the glucose and fructose concentration in berries by 17–21% [84]. It is speculated that soluble carbohydrates play a dual role in relation to reactive oxygen species. Soluble sugars may be involved in or associated with metabolic processes that generate reactive oxygen species (ROS). Correlatively, low glucose can decrease mitochondrial ROS production [85]. Therefore, the lower levels of glucose and fructose in our N-Br-treated peppers can correlate with lower MDA and EL and the higher photosynthetic performance of the plants when subjected to heat stress.
BR application on rice could reduce the negative effects of heat stress on the dark reactions of photosynthesis [81,86]. Under high-temperature conditions, the BR-pretreated plants showed higher values of photosynthesis when compared to the non-BR-treated and heat-stressed plants.
4.5. The Interaction Effect of Br and N-Br on Saturated and Unsaturated Fatty Acids of Pepper
The saturated/unsaturated fatty acids ratio increased when Br and N-Br were utilized for heat-stressed plants, with N-Br exhibiting the greatest effect (Table 1). In all treatments, palmitic acid (C16:1) was the most abundant saturated fatty acid, and linoleic acid (C18:2) was the richest unsaturated fatty acid. The saturated/unsaturated fatty acids ratio increased at 35 °C and in the presence of Br and N-Br. Early studies demonstrated that fatty acid saturation can improve the heat stability of plant membranes [87]. Unsaturated fatty acids modify numerous membrane-bound protein functions in spinach [88]. Adaptation to moderately high temperatures (35 °C) induced changes in membrane fatty acid content and lipid saturation rate [89,90]. In contrast, the heat stress increased the amount of unsaturated fatty acids, such as palmitic acid and stearic acid. At moderate to high temperatures, membranes with a higher content of saturated fatty acids showed an improved permeability and integrity of membranes and enhanced membrane dynamics under heat stress. They observed a correlation between the degree of leaf membrane lipid saturation and the pepper plant’s ability to minimize heat stress damage [83].
In a study with mango fruits, BR treatment resulted in an elevated content of unsaturated fatty acids (linoleic acid and linolenic acid) when compared with the control. The same treatment reduced the content of palmitoleic acid, palmitic acid, stearic acid, and oleic acid [91]. In another study on safflower [92], 24-epibrassinolide treatment at a 0.1 μM concentration increased the oil and fatty acid content except the palmitic acid percentage, which also remained unchanged or declined slightly in our Br- and N-Br-treated pepper plants.
5. Conclusions
Heat stress at 35 °C resulted in a mild decline in growth parameters of bell peppers. But the effect was more pronounced when physiological parameters were investigated. Br and N-Br treatments appear to increase pepper’s tolerance to heat stress primarily via enhanced enzymatic antioxidant systems and secondly through osmotic adjustment by glucose, fructose, and proline content variation. By maintaining membrane integrity and permeability through fatty acid modification, Br and N-Br could enhance pepper tolerance to heat stress. Br and N-Br showed a significant effect on the reproductive phase of growth by increasing the number of flowers and fruits under optimum and high-temperature stress conditions, respectively. Decreased floral abscission (a concern for greenhouse farmers during the summer) after high-temperature stress was also confirmed in response to the BR application. This could be due to the increasing level of availability of essential assimilates for floral development and fertility. The reduced lipid peroxidation, as manifested in a lower MDA and electrolyte leakage due to the BR application, is suggestive of their protective role in the abiotic stress tolerance of bell peppers. The data analysis revealed that SOD activity, antioxidant capacity (DPPH), proline level, relative water content (RWC), and leaf water potential were the most affected parameters in response to brassinosteroid application under high-temperature stress. More research is necessary to determine the molecular mechanism of BRs and their effect on reproductive physiology in pepper plants under heat stress. Despite the overall positive and significant effects of BR application for pepper plants under high temperature, we have not observed a statistically significant difference between the bulk form and the encapsulated forms of BRs in the majority of studied traits in response to heat stress; therefore, future research is needed to confirm the justifiability of encapsulation for BR applications.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/horticulturae10101062/s1, Figure S1: SEM image of nano-encapsulated brassinosteroid with a magnification of 16000; Figure S2: Results of nano-capsule size distribution based on DLS; Figure S3: Zeta potential results of BR nano-capsules with DLS device; Table S1: Analysis of variance of heat stress and brassinosteroid source on growth parameters of pepper; Table S2: The main effect of heat stress and brassinosteroid source on growth parameters of pepper; Table S3: Analysis of variance of heat stress and brassinosteroid source on physiological and biochemical parameters of pepper; Table S4: The main effect of heat stress and brassinosteroid source on physiological and biochemical parameters of pepper; Table S5: Analysis of variance of heat stress and brassinosteroid source on antioxidant enzymes and glucose and fructose of pepper; Table S6: The main effect of heat stress and brassinosteroid source on antioxidant enzymes and glucose and fructose of pepper; Table S7: Analysis of variance of heat stress and brassinosteroid source on fatty acid composition of pepper; Table S8: The main effect of heat stress and brassinosteroid source on fatty acid composition of pepper.
Author Contributions
Conceptualization, M.H.; methodology, M.H.; software, B.H.; validation, G.P.K., I.M. and A.S.; formal analysis, M.H.; investigation, B.H.; resources, M.H.; data curation, B.H.; writing—original draft preparation, B.H.; writing—review and editing, M.H., G.P.K. and I.M.; visualization, B.H. and A.S.; supervision, M.H.; funding acquisition, M.H. and I.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
All data generated or analyzed during this study are included in this published article and additional information will be available upon request to the corresponding authors.
Acknowledgments
The authors of this work are grateful to the Isfahan University of Technology for providing the research fund and support. This research was also supported by the Research Excellence Programme of the Hungarian University of Agriculture and Life Sciences.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Fan, R.; Tao, X.Y.; Xia, Z.Q.; Sim, S.; Hu, L.S.; Wu, B.D.; Wang, Q.H.; Hao, C.Y. Comparative transcriptome and metabolome analysis of resistant and susceptible piper species upon infection by the oomycete phytophthora capsici. Front. Plant Sci. 2022, 13, 864927. [Google Scholar] [CrossRef] [PubMed]
- Nolan, R.H.; Collins, L.; Leigh, A.; Ooi, M.K.J.; Curran, T.J.; Fairman, T.A.; Resco de Dios, V.; Bradstock, R. Limits to post-fire vegetation recovery under climate change. Plant Cell Environ. 2021, 44, 3471–3489. [Google Scholar] [CrossRef]
- Ahmed, W.; Xia, Y.; Li, R.; Bai, G.; Siddique, K.H.M.; Guo, P. Non-coding RNAs: Functional roles in the regulation of stress response in Brassica crops. Genomics 2020, 112, 1419–1424. [Google Scholar] [CrossRef] [PubMed]
- Taheri, M.; Haghighi, M. Benzyl adenine is more effective than potassium silicate on decreasing the detrimental effects of heat stress in pepper (Capsicum annum cv. PS301). Iran Agric. Res. 2018, 37, 89–98. [Google Scholar] [CrossRef]
- Pagamas, P.; Nawata, E. Sensitive stages of fruit and seed development of chili pepper (Capsicum annuum L. var. Shishito) exposed to high-temperature stress. Scie Hortic. 2008, 117, 21–25. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Roychowdhury, R.; Fujita, M. Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants. Int. J. Mol. Sci. 2013, 14, 9643–9684. [Google Scholar] [CrossRef]
- Shabala, S.; Cuin, T.A. Potassium transport and plant salt tolerance. Physiol. Plant. 2008, 133, 651–669. [Google Scholar] [CrossRef] [PubMed]
- Jogawat, A. Osmolytes and their role in abiotic stress tolerance in plants. In Molecular Plant Abiotic Stress: Biology and Biotechnology; Wiley: London, UK, 2019; pp. 91–104. [Google Scholar] [CrossRef]
- Raza, A.; Razzaq, A.; Mehmood, S.S.; Hussain, M.A.; Wei, S.; He, H. Omics: The way forward to enhance abiotic stress tolerance in Brassica napus L. GM Crops Food 2021, 12, 251–281. [Google Scholar] [CrossRef]
- Saddiq, M.S.; Afzal, I.; Iqbal, S.; Hafeez, M.B.; Raza, A. Low sodium content in leaves improves grain yield and physiological performance of wheat genotypes in saline- sodic soil. Trop. Agric. Res. Pesqu. Agropec. Trop. 2021, 51, e67663. [Google Scholar] [CrossRef]
- dos Santos, T.B.; Ribas, A.F.; de Souza, S.G.H.; Budzinski, I.G.F.; Domingues, D.S. Physiological responses to drought, salinity, and heat stress in plants: A review. Stresses 2022, 2, 113–135. [Google Scholar] [CrossRef]
- Raza, A. Metabolomics: A systems biology approach for enhancing heat stress tolerance in plants. Plant Cell Rep. 2020, 41, 741–763. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Parvin, K.; Bardhan, K.; Nahar, K.; Anee, T.I.; Masud, A.A.C.; Fotopoulos, V. Biostimulants for the regulation of reactive oxygen species metabolism in plants under abiotic stress. Cells 2021, 10, 2537. [Google Scholar] [CrossRef] [PubMed]
- Napieraj, N.; Janicka, M.; Reda, M. Interactions of polyamines and phytohormones in plant response to abiotic stress. Plants 2023, 12, 1159. [Google Scholar] [CrossRef]
- Sarraf, M.; Janeeshma, E.; Arif, N.; QudratUllahFarooqi, M.; Kumar, V.; Ansari, N.A.; Ghani, M.I.; Ahanger, M.A.; Hasanuzzaman, M. Understanding the role of beneficial elements in developing plant stress resilience: Signalling and crosstalk with phytohormones and microbes. Plant Stress 2023, 10, 100224. [Google Scholar] [CrossRef]
- Manghwar, H.; Hussain, A.; Ali, Q.; Liu, F. Brassinosteroids (BRs) role in plant development and coping with different stresses. Int. J. Mol. Sci. 2022, 23, 1012. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.P.S.D.; Alencar, A.A.d.S.; Sudré, C.P.; Araújo, M.d.S.B.d.; Lobato, A.K.d.S. Brassinosteroids: Relevant evidence related to mitigation of abiotic and biotic stresses in plants. Agronomy 2024, 14, 840. [Google Scholar] [CrossRef]
- Peres, A.L.G.L.; Soares, J.S.; Tavares, R.G.; Righetto, G.; Zullo, M.A.T.; Mandava, N.B.; Menossi, M. Brassinosteroids, the sixth class of phytohormones: A molecular view from the discovery to hormonal interactions in plant development and stress adaptation. Int. J. Mol. Sci. 2019, 20, 331. [Google Scholar] [CrossRef]
- Miao, R.; Li, C.; Liu, Z.; Zhou, X.; Chen, S.; Zhang, D.; Luo, J.; Tang, W.; Wang, C.; Wu, J.; et al. The role of endogenous brassinosteroids in the mechanisms regulating plant reactions to various abiotic stresses. Agronomy 2024, 14, 356. [Google Scholar] [CrossRef]
- Basit, F.; Liu, J.; An, J.; Chen, M.; He, C.; Zhu, X.; Li, Z.; Hu, J.; Guan, Y. Brassinosteroids as a multidimensional regulator of plant physiological and molecular responses under various environmental stresses. Environ. Sci. Pollut. Res. 2021, 28, 44768–44779. [Google Scholar] [CrossRef]
- Wu, W.; Zhang, Q.; Ervin, E.H.; Yang, Z.; Zhang, X. Physiological mechanism of enhancing salt stress tolerance of perennial ryegrass by 24-epibrassinolide. Front. Plant Sci. 2017, 8, 1017–1026. [Google Scholar] [CrossRef]
- Abbas, S.; Latif, H.H.; Elsherbiny, E.A. Effect of 24-epibrassinolide on the physiological and genetic changes on two varieties of pepper under salt stress conditions. Pak. J. Bot. 2013, 45, 1273–1284. [Google Scholar]
- Zulfiqar, F.; Nafees, M.; Chen, J.; Darras, A.; Ferrante, A.; Hancock, J.T.; Ashraf, M.; Zaid, A.; Latif, N.; Corpas, F.J.; et al. Chemical priming enhances plant tolerance to salt stress. Front. Plant Sci. 2022, 13, 2883. [Google Scholar] [CrossRef] [PubMed]
- Ogweno, O.; Otieno, J.; Shun Song, X.; Shi, K.; Hu, W.H.; Mao, W.H.; Hong Zhou, Y.; Yu, J.Q.; Nogués, S. Brassinosteroids alleviate heat-induced inhibition of photosynthesis by increasing carboxylation efficiency and enhancing antioxidant systems in Lycopersicon esculentum. J. Plant Growth Regul. 2008, 27, 49–57. [Google Scholar] [CrossRef]
- Wang, W.; Xie, Y.; Liu, C.; Jiang, H. The exogenous application of brassinosteroids confers tolerance to heat stress by increasing antioxidant capacity in soybeans. Agriculture 2022, 12, 1095. [Google Scholar] [CrossRef]
- Eremina, M.; Unterholzner, S.J.; Rathnayake, A.I.; Castellanos, M.; Khan, M.; Kugler, K.G.; May, S.T.; Mayer, K.F.; Rozhon, W.; Poppenberger, B. Brassinosteroids participate in the control of basal and acquired freezing tolerance of plants. Proc. Natl. Acad. Sci. USA 2016, 113, E5982–E5991. [Google Scholar] [CrossRef]
- Yang, P.; Wang, Y.; Li, J.; Bian, Z. Effects of brassinosteroids on photosynthetic performance and nitrogen metabolism in pepper seedlings under chilling stress. Agronomy 2019, 9, 839. [Google Scholar] [CrossRef]
- Wang, Y.; Jiang, Q.; Wang, X.; Xi, Z. Brassinosteroid stimulates hydrogen peroxide biosynthesis and reduces the effect of cold stress. J. Plant Growth Regul. 2023, 42, 3757–3769. [Google Scholar] [CrossRef]
- Zulfiqar, F.; Ashraf, M. Nanoparticles potentially mediate salt stress tolerance in plants. Plant Physiol. Biochem. 2021, 160, 257–268. [Google Scholar] [CrossRef]
- Hamed, R.; Jodeh, S.; Alkowni, R. Nano bio fertilizer capsules for sustainable agriculture. Sci Rep. 2024, 14, 13646. [Google Scholar] [CrossRef]
- Zahra, Z.; Habib, Z.; Hyun, H.; Shahzad, H.M.A. Overview on recent developments in the design, application, and impacts of nanofertilizers in agriculture. Sustainability 2022, 14, 9397. [Google Scholar] [CrossRef]
- Zulfiqar, F.; Navarro, M.; Ashraf, M.; Akram, N.A.; Munne-Bosch, S. Nanofertilizer use for sustainable agriculture: Advantages and limitations. Plant Sci. 2019, 289, 110270. [Google Scholar] [CrossRef] [PubMed]
- Garg, D.; Sridhar, K.; Stephen Inbaraj, B.; Chawla, P.; Tripathi, M.; Sharma, M. Nano-biofertilizer formulations for agriculture: A systematic review on recent advances and prospective applications. Bioengineering 2023, 10, 1010. [Google Scholar] [CrossRef] [PubMed]
- Yadav, A.; Yadav, K.; Abd-Elsalam, K.A. Nanofertilizers: Types, delivery and advantages in agricultural sustainability. Agrochemicals 2023, 2, 296–336. [Google Scholar] [CrossRef]
- Hong, J.; Wang, C.; Wagner, D.C.; Gardea-Torresdey, J.L.; He, F.; Rico, C.M. Foliar application of nanoparticles: Mechanisms of absorption, transfer, and multiple impacts. Environ. Sci. Nano 2021, 8, 1196–1210. [Google Scholar] [CrossRef]
- Abbasifar, A.; Shahrabadi, F.; Valizadehkaji, B. Effects of green synthesized zinc and copper nano- fertilizers on the morphological and biochemical attributes of basil plant. J. Plant Nutr. 2020, 43, 1104–1118. [Google Scholar] [CrossRef]
- Jones, J.B. Hydroponics: A Practical Guide for the Soilless Grower; CRC Press: New York, NY, USA, 2016. [Google Scholar]
- Mozafari, R.; Johnson, M.; Hatziantoniou, C.; Demetzos, S.C. Nanoliposomes and their applications in food nanotechnology. J. Liposome Res. 2008, 18, 309–327. [Google Scholar] [CrossRef]
- Hallaji, B.; Haghighi, M.; Abolghasemi, R.; Mozafarian, M. Effect of foliar applications of aminolevulinic acid (bulk and nano-encapsulated) on bell pepper under heat stress. Plant Stress 2024, 12, 100477. [Google Scholar] [CrossRef]
- Seetapan, N.; Bejrapha, P.; Srinuanchai, W.; Ruktanonchai, U.R. Rheological and morphological characterizations on physical stability of gamma-oryzanol-loaded solid lipid nanoparticles (SLNs). Micron 2010, 41, 51–58. [Google Scholar] [CrossRef]
- Rodriguez-Dominguez, C.M.; Forner, A.; Martorell, S.; Choat, B.; Lopez, R.; Peters, J.M.R.; Pfautsch, S.; Mayr, S.; Carins-Murphy, M.R.; McAdam, S.A.M.; et al. Leaf water potential measurements using the pressure chamber: Synthetic testing of assumptions towards best practices for precision and accuracy. Plant Cell Environ. 2022, 45, 037–2061. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence a practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Dhopte, A.M.; Livera, M.M. Principles and Techniques for Plant Scientist; Agrobios: Chopasani, India, 2002. [Google Scholar]
- Lutts, S.; Kinet, J.; Bouharmont, J. NaCl-induced senescence in leaves of rice (Oryza sativa L.) cultivars differing in salinity tolerate. Ann. Bot. 1996, 78, 389–398. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Koleva, I.I.; Van Beek, T.A.; Linssen, J.P.; Groot, A.D.; Evstatieva, L.N. Screening of plant extracts for antioxidant activity: A comparative study on three testing methods. Phytochem Anal. 2002, 13, 8–17. [Google Scholar] [CrossRef]
- Tabatabaei, S.; Ehsanzadeh, P. Photosynthetic pigments, ionic and antioxidative behaviour of hulled tetraploid wheat in response to NaCl. Photosynthetica 2016, 54, 340–350. [Google Scholar] [CrossRef]
- Khoshbakht, D.; Asghari, M.R.; Haghighi, M. Effects of foliar applications of nitric oxide and spermidine on chlorophyll fluorescence, photosynthesis and antioxidant enzyme activities of citrus seedlings under salinity stress. Photosynthetica 2018, 56, 1313–1325. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Jiang, M.; Zhang, J. Effect of abscisic acid on active oxygen species, antioxidative defence system and oxidative damage in leaves of maize seedlings. Plant Cell Physiol. 2001, 42, 1265–1273. [Google Scholar] [CrossRef]
- Adams, S.H.; Stanhope, K.L.; Grant, R.W.; Cummings, B.P.; Havel, P.J. Metabolic and endocrine profiles in response to systemic infusion of fructose and glucose in rhesus macaques. Endocrinology 2008, 149, 3002–3008. [Google Scholar] [CrossRef][Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Goli, S.A.H.; Kadivar, M.; Keramat, J.; Fazilati, M. Conjugated linoleic acid (ClA) production and lipase-catalyzed interesterification of purified ClA with canola oil. Eur. J. Lipid Sci. Technol. 2008, 110, 400–404. [Google Scholar] [CrossRef]
- Abdelmageed, A.H.A.; Gruda, N. Influence of high temperatures on gas exchange rate and growth of eight tomato cultivars under controlled heat stress conditions. Eur. J. Hortic. Sci. 2009, 74, 152–162. [Google Scholar]
- Niu, J.H.; Anjum, A.S.; Wang, R.; Li, J.H.; Liu, M.R.; Song, J.X.; Zohaib, A.; Lv, J.; Wang, S.G.; Zong, X.F. Exogenous application of brassinolide can alter morphological and physiological traits of Leymus chinensis (Trin) Tzvelev under room and high temperatures. Chil. J. Agric. Res. 2016, 76, 27–33. [Google Scholar] [CrossRef]
- Li, J.; Yang, P.; Kang, J.; Gan, Y.; Yu, J.; Calderon-Urrea, A.; Jian, L.; Zhang, G.; Feng, Z.; Xie, J. Transcriptome analysis of pepper revealed a role of 24-epibrassinolide in response to chilling. Front. Plant Sci. 2016, 7, 1281. [Google Scholar] [CrossRef] [PubMed]
- Shu, S.; Tang, Y.; Yuan, Y.; Sun, J.; Zhong, M.; Guo, S. The role of 24-epibrassinolide in the regulation of photosynthetic characteristics and nitrogen metabolism of tomato seedlings under a combined low temperature and weak light stress. Plant Physiol. Biochem. 2016, 7, 344–353. [Google Scholar] [CrossRef]
- Wu, Z.; Gu, S.; Gu, H.; Cheng, D.; Li, L.; Guo, X.; Wang, M.; He, S.; Li, M.; Chen, J. Physiological and transcriptomic analyses of brassinosteroid function in kiwifruit root Zhiyong. Environ. Exp. Bot. 2022, 194, 104685. [Google Scholar] [CrossRef]
- Tong, H.; Xiao, Y.; Liu, D.; Gao, S.; Liu, L.; Yin, Y.; Jin, Y.; Qian, Q.; Chu, C. Brassinosteroid regulates cell elongation by modulating gibberellin metabolism in rice. Plant Cell 2014, 26, 4376–4393. [Google Scholar] [CrossRef]
- Upreti, K.K.; Srinivasa Rao, N.K.; Jayaram, H.L. Floral abscission in capsicum under high temperature: Role of endogenous hormones and polyamines. Indian J. Plant Physiol. 2012, 17, 207–214. [Google Scholar]
- Dutta, S.K.; Gurung, G.; Yadav, A.; Laha, R.; Mishra, V.K. Factors associated with citrus fruit abscission and management strategies developed so far: A review. New Zealand J. Crop. Hortic. Sci. 2022, 51, 467–488. [Google Scholar] [CrossRef]
- Shi, Y.; Song, B.; Liang, Q.; Su, D.; Lu, W.; Liu, Y.; Li, Z. Molecular regulatory events of flower and fruit abscission in horticultural plants. Hortic. Plant J. 2023, 9, 867–883. [Google Scholar] [CrossRef]
- Ma, X.; Yuan, Y.; Li, C.; Wu, Q.; He, Z.; Li, J.; Zhao, M. Brassinosteroids suppress ethylene-induced fruitlet abscission through LcBZR1/2-mediated transcriptional repression of LcACS1/4 and LcACO2/3 in litchi. Hortic. Res. 2021, 8, 105. [Google Scholar] [CrossRef]
- Lv, B.; Tian, H.; Zhang, F.; Liu, J.; Lu, S.; Bai, M.; Li, C.; Ding, Z. Brassinosteroids regulate root growth by controlling reactive oxygen species homeostasis and dual effect on ethylene synthesis in Arabidopsis. PLoS Genet. 2018, 14, e1007144. [Google Scholar] [CrossRef] [PubMed]
- Furio, R.; Salazar, S.; Mariotti-Martínez, J.; Martínez-Zamora, G.; Coll, Y.; Díaz-Ricci, J. Brassinosteroid applications enhance the tolerance to abiotic stresses, production and quality of strawberry fruits. Horticulturae 2022, 8, 572. [Google Scholar] [CrossRef]
- Ji, W.; Luo, H.; Song, Y.; Hong, E.; Li, Z.; Lin, B.; Fan, C.; Wang, H.; Song, X.; Jin, S.; et al. Changes in photosynthetic characteristics of paeonia suffruticosa under high temperature stress. Agronomy 2022, 12, 1203. [Google Scholar] [CrossRef]
- Sadura, I.; Janeczko, A. Brassinosteroids and the tolerance of cereals to low and high temperature stress: Photosynthesis and the physicochemical properties of cell membranes. Int. J. Mol. Sci. 2022, 23, 342. [Google Scholar] [CrossRef]
- Gudesblat, G.E.; Schneider-Pizon, J.; Betti, C.; Mayerhofer, J.; Vanhoutte, I.; Dongen, W. Speechless integrates brassinosteroid and stomata signalling pathways. Nat. Cell Biol. 2012, 14, 548–554. [Google Scholar] [CrossRef] [PubMed]
- Daszkowska-Golec, A.; Szarejko, I. Open or close the gate—Stomata action under the control of phytohormones in drought stress conditions. Front. Plant Sci. 2013, 4, 138. [Google Scholar] [CrossRef]
- Sharma, I.; Ching, E.; Saini, S.; Bhardwaj, R.; Pati, P.K. Exogenous application of brassinosteroid offers tolerance to salinity by altering stress responses in rice variety Pusa Basmati-1. Plant Physiol. Biochem. 2013, 69, 17–26. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Mir, R.A.; Alyemeni, M.N.; Ahmad, P. Combined effects of brassinosteroid and kinetin mitigates salinity stress in tomato through the modulation of antioxidant and osmolyte metabolism. Plant Physiol. Biochem. 2020, 147, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Neha; Twinkle; Mohapatra, S.; Sirhindi, G.; Dogra, V. Seed priming with brassinolides improves growth and reinforces antioxidative defenses under normal and heat stress conditions in seedlings of Brassica juncea. Physiol. Plant. 2022, 174, e13814. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, Y.; Chen, H.; Xiang, J.; Zhang, Y.; Wang, Z.; Zhu, D.; Zhang, Y. Brassinosteroids mediate endogenous phytohormone metabolism to alleviate high temperature injury at panicle initiation stage in rice. Rice Sci. 2023, 30, 70–86. [Google Scholar] [CrossRef]
- Nazir, F.; Jahan, B.; Kumari, S.; Iqbal, N.; Albaqami, M.; Sofo, A.; Khan, M.I.R. Brassinosteroid modulates ethylene synthesis and antioxidant metabolism to protect rice (Oryza sativa) against heat stress-induced inhibition of source—Sink capacity and photosynthetic and growth attributes. J. Plant Physiol. 2023, 289, 154096. [Google Scholar] [CrossRef] [PubMed]
- Hafeez, M.B.; Zahra, N.; Zahra, K.; Raza, A.; Khan, A.; Shaukat, K.; Khan, S. Brassinosteroids: Molecular and physiological responses in plant growth and abiotic stresses. Plant Stress 2021, 2, 100029. [Google Scholar] [CrossRef]
- Camejo, D.; Rodríguez, P.; Morales, M.A.; Dell’Amico, J.M.; Torrecillas, A.; Alarcón, J.J. High temperature effects on photosynthetic activity of two tomato cultivars with different heat susceptibility. J. Plant Physiol. 2005, 162, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.P.; Huang, L.F.; Cheng, F.; Zhou, Y.H.; Xia, X.J.; Mao, W.H.; Shi, K.; Yu, J. Brassinosteroids accelerate recovery of photosynthetic apparatus from cold stress bybalancing the electron partitioning, carboxylation and redox homeostasis in cucumber. Physiol. Plant. 2013, 148, 133–145. [Google Scholar] [CrossRef]
- Lee, H.J.; Lee, J.H.; Lee, S.G.; An, S.; Lee, H.S.; Choi, C.K.; Kim, S.K. Foliar application of biostimulants affects physiological responses and improves heat stress tolerance in Kimchi cabbage. Hortic. Environ. Biotech. 2020, 61, 207. [Google Scholar] [CrossRef]
- Arief, M.A.A.; Kim, H.; Kurniawan, H.; Nugroho, A.P.; Kim, T.; Cho, B.-K. Chlorophyll fluorescence imaging for early detection of drought and heat stress in strawberry plants. Plants 2023, 12, 1387. [Google Scholar] [CrossRef]
- Bhattarai, S.; Harvey, J.T.; Djidonou, D.; Leskovar, D.I. Exploring morpho-physiological variation for heat stress tolerance in tomato. Plants 2021, 10, 347. [Google Scholar] [CrossRef]
- Thussagunpanit, J.; Jutamanee, K.; Sonjaroon, W.; Kaveeta, L.; Chai-Arree, W.; Pankean, P.; Suksamrarn, A. Effects of brassinosteroid and brassinosteroid mimic on photosynthetic efficiency and rice yield under heat stress. Photosynthetica 2015, 53, 312–320. [Google Scholar] [CrossRef]
- Adams, S.R.; Cockshull, K.E.; Cave, C.R.J. Effect of temperature on the growth and development of tomato fruits. Ann. Bot. 2001, 88, 869–877. [Google Scholar] [CrossRef]
- Motamedi, M.; Haghighi, M.; Goli, A. Physiological changes of sweet and hot peppers in vegetative and reproductive growth stages treated by Ca and H2O2 under unforeseen heat stresses. Sci. Hortic. 2019, 249, 306–313. [Google Scholar] [CrossRef]
- Sepuveda, G.; Kliewer, W.M. Effect of high temperature on grapevines (Vitis vinifera L.). II. Distribution of soluble sugars. Am. J. Enol. Vitic. 1986, 37, 20–25. [Google Scholar] [CrossRef]
- Barros, M.H.; Bandy, B.; Tahara, E.B.; Kowaltowski, A.J. Higher respiratory activity decreases mitochondrial reactive oxygen release and increases life span in Saccharomyces cerevisiae. J. Biol. Chem. 2004, 279, 49883–49888. [Google Scholar] [CrossRef] [PubMed]
- Thussagunpanit, J.; Jutamanee, K.; Kaveeta, L.; Chai-arree, W.; Pankean, P.; Homvisasevongsa, S.; Suksamrarn, A. Comparative effects of brassinosteroid and brassinosteroid mimic on improving photosynthesis, lipid peroxidation and rice seed set under heat stress. J. Plant Growth Regul. 2015, 34, 320–331. [Google Scholar] [CrossRef]
- Raison, J.K.; Roberts, J.K.; Berry, J.A. Correlations between the thermal stability of chloroplast (thylakoid) membranes and the composition and fluidity of their polar lipids upon acclimation of the higher plant, Nerium oleander, to growth temperature. Biochim. Biophys. Acta. 1982, 688, 218–228. [Google Scholar] [CrossRef]
- Combos, Z.; Wada, H.; Hideg, E.; Murata, N. The unsaturation of membrane lipids stabilizes photosynthesis against heat stress. Plant. Physiol. 1994, 104, 563–567. [Google Scholar] [CrossRef]
- Quinn, P.J.; Joo, F.; Vigh, L. The role of unsaturated lipids in membrane structure and stability. Prog. Biophys. Mol. 1989, 53, 71–103. [Google Scholar] [CrossRef] [PubMed]
- Saidi, Y.; Peter, M.; Finka, A.; Cicekli, C.; Vigh, L.; Goloubinoff, P. Membrane lipid composition affects plant heat sensing and modulates Ca2+-dependent heat shock response. Plant Signal. Behav. 2010, 5, 1530–1533. [Google Scholar] [CrossRef]
- Li, B.; Zhang, C.; Cao, B.; Qin, G.; Wang, W.; Tian, S. Brassinolide enhances cold stress tolerance of fruit by regulating plasma membrane proteins and lipids. Amino Acids 2012, 43, 2469–2480. [Google Scholar] [CrossRef]
- Zafari, M.; Ebadi, A.; Sedghi, M.; Jahanbakhsh, S.; Miransari, M.J. Alleviating effect of 24-epibrassinolide on seed oil content and fatty acid composition under drought stress in safflower. J. Food Compost. Anal. 2020, 92, 103544. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).