
Identification of an Entomopathogenic Fungus, Pseudozyma flocculosa (Traquair, Shaw & Jarvis), and Its Efficacy against Tetranychus urticae Koch




Abstract
:1. Introduction
2. Materials and Methods
2.1. Tetranychus urticae Mites
2.2. Preparation of Eggs, Larvae, and Protonymphs of T. urticae
2.3. Fungus and Fungal Suspensions
2.4. Morphological and Molecular Identification
2.5. Laboratory Bioassays
2.6. Field Trials
2.7. Statistical Analysis
3. Results
3.1. Conidia Isolation and Identification
3.2. Pathogenicity of Multiple Conidium Concentrations on Adult T. urticae
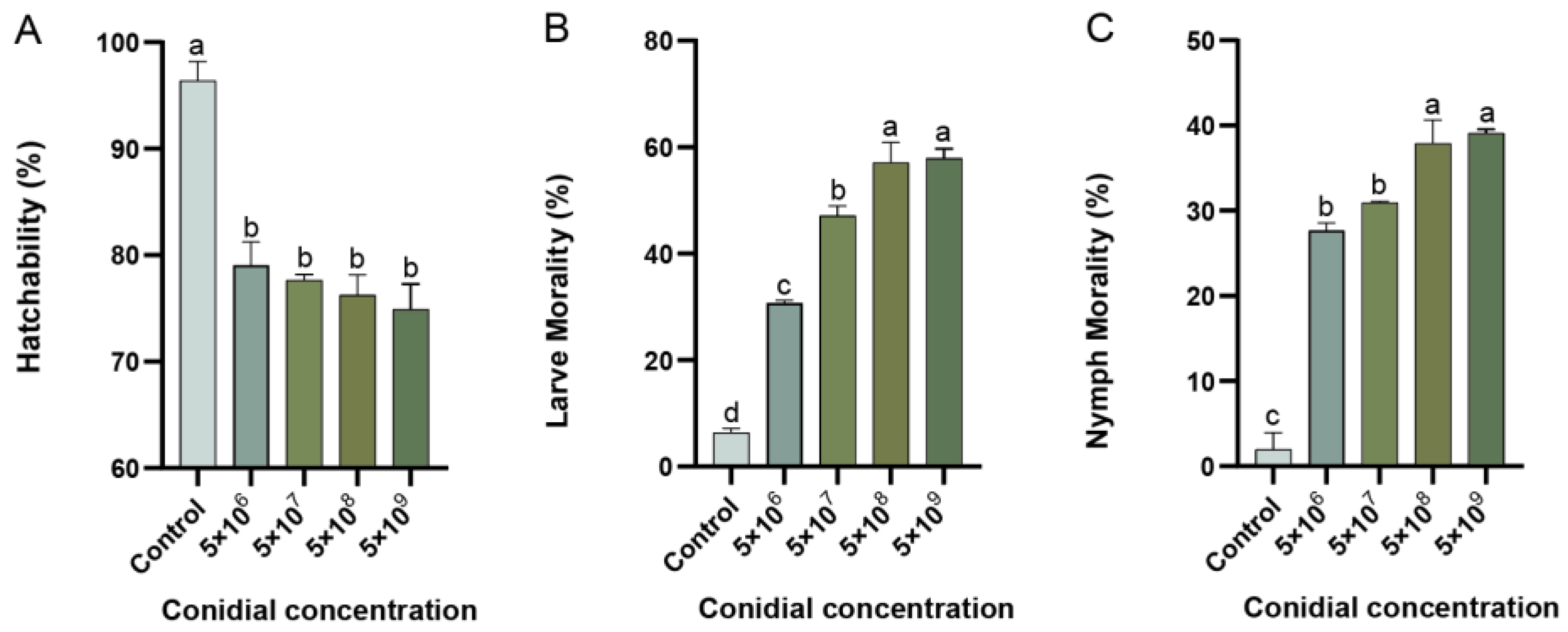
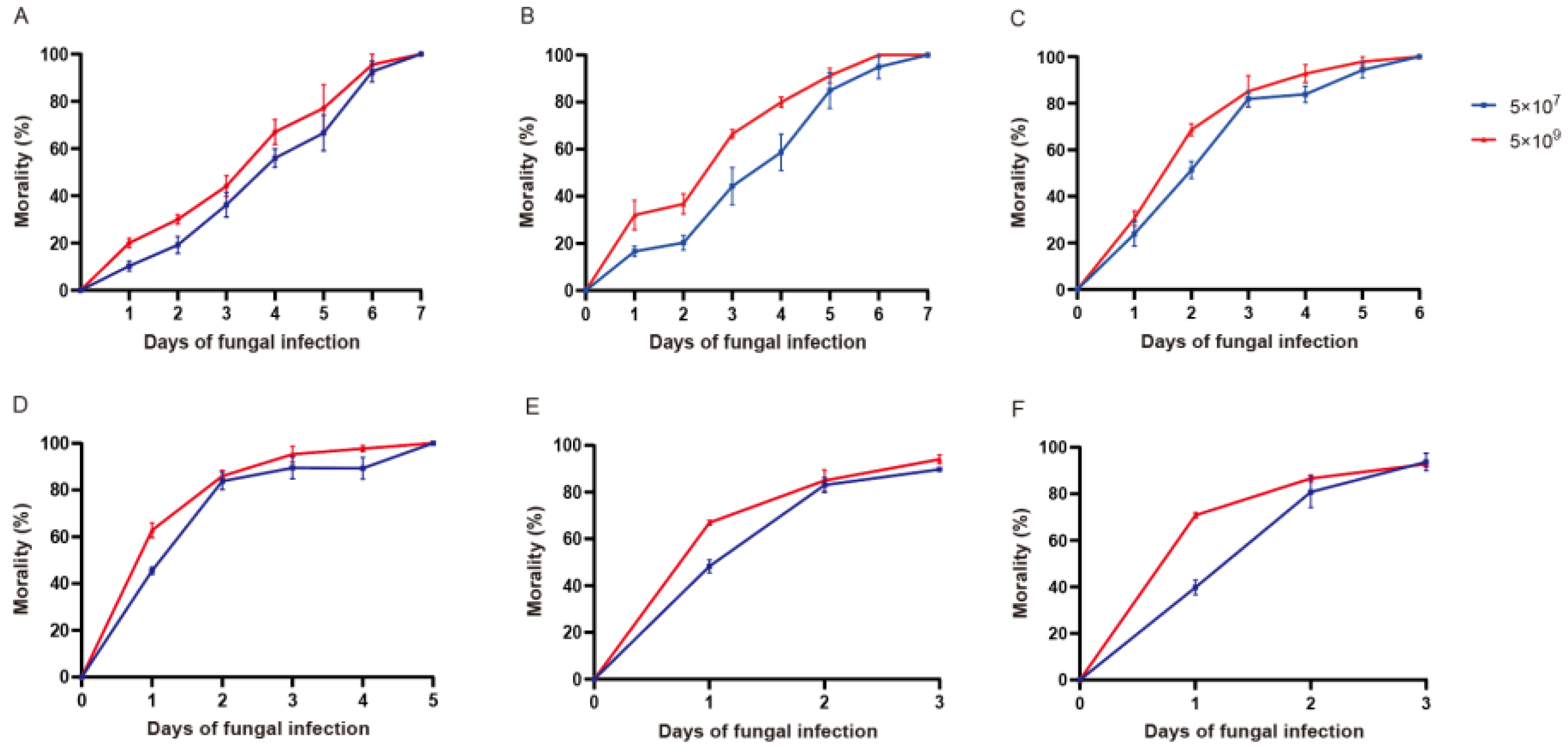
3.3. Mortality at Different Stages of T. urticae after Exposure to P. flocculosa Concentrations
3.4. Differential Pathogenicity of P. flocculosa against T. urticae at Different Temperatures
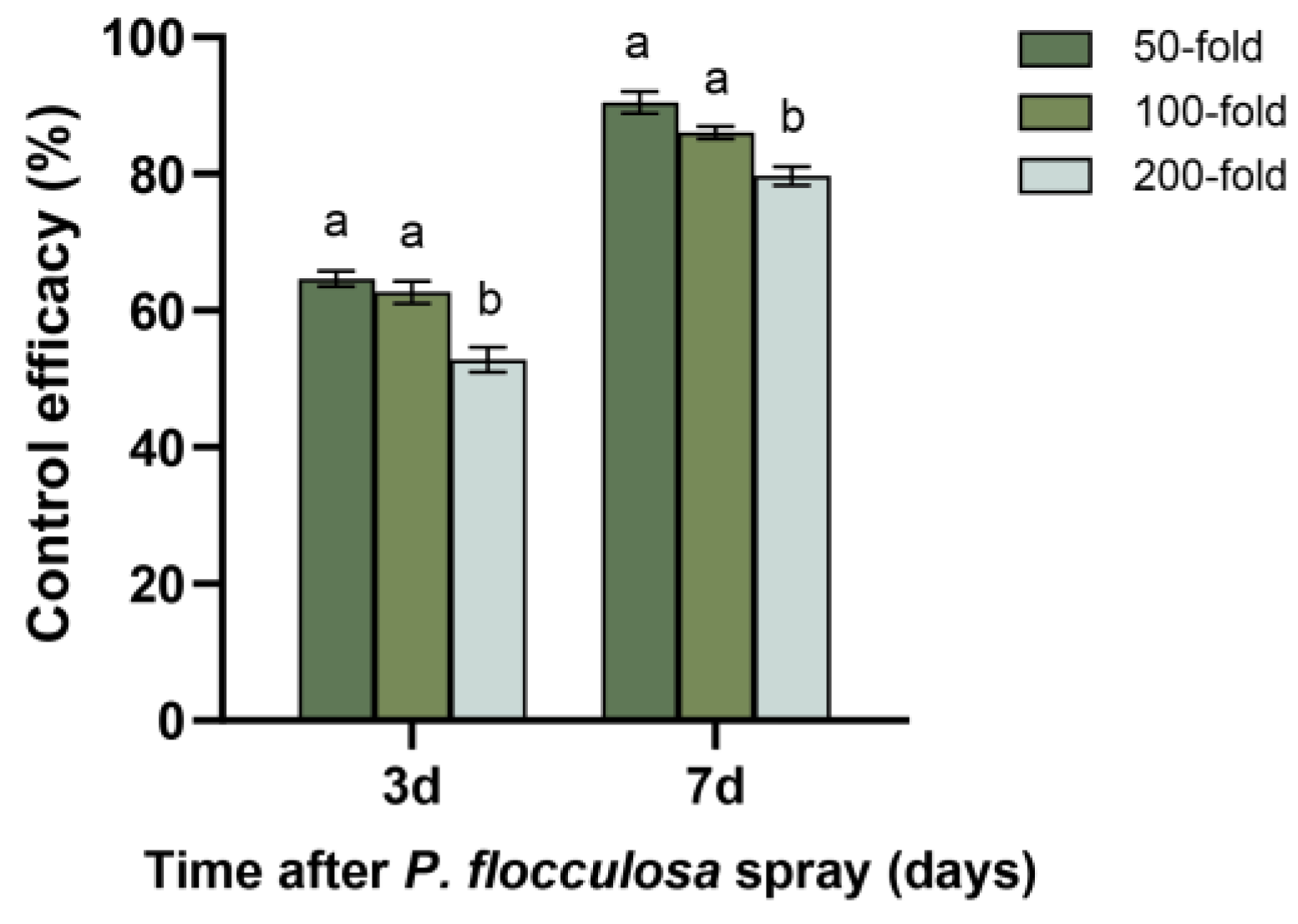
3.5. Field Efficacy of P. flocculosa
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Van Leeuwen, T.; Vontas, J.; Tsagkarakou, A.; Dermauw, W.; Tirry, L. Acaricide resistance mechanisms in the two-spotted spider mite Tetranychus urticae and other important Acari: A review. Insect Biochem. Mol. Biol. 2010, 40, 563–572. [Google Scholar] [CrossRef] [PubMed]
- Tian, T.; Wu, M.M.; Zhang, Y.; Xu, D.D.; Wu, M.Y.; Xie, W.; Su, Q.; Wang, S.L. Pesticide Resistance and Related Mutation Frequencies of Tetranychus urticae in Hainan, China. Horticulturae 2022, 8, 590. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, D.D.; Zhang, Y.J.; Wu, Q.J.; Xie, W.; Guo, Z.J.; Wang, S.L. Frequencies and mechanisms of pesticide resistance in Tetranychus urticae field populations in China. Insect Sci. 2022, 29, 827–839. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Tian, T.; Zhang, K.; Zhang, Y.J.; Wu, Q.J.; Xie, W.; Guo, Z.J.; Wang, S.L. Lack of fitness cost and inheritance of resistance to abamectin based on the establishment of a near-isogenic strain of Tetranychus urticae. J. Integr. Agric. 2023, 22, 1809–1819. [Google Scholar] [CrossRef]
- Liu, Z.G.; Tobi Fadiji, T.; Yang, J.; Li, Z.G.; Tchuenbou-Magaia, F. Impact of mechanical stimulation on the life cycle of horticultural plant. Hortic Plant J. 2023, 9, 381–394. [Google Scholar] [CrossRef]
- Canassa, F.; Esteca, F.C.N.; Moral, R.A.; Meyling, N.V.; Klingen, I.; Delalibera, I. Root inoculation of strawberry with the entomopathogenic fungi Metarhizium robertsii and Beauveria bassiana reduces incidence of the two spotted spider mite and selected insect pests and plant diseases in the field. J. Pest Sci. 2020, 93, 261–274. [Google Scholar] [CrossRef]
- Olaniyi, O.G.; Rhodes, E.M.; Chase, C.A.; Liburd, O.E. The Effect of Summer Cover Crops and strawberry Cultivars on the Twospotted Spider Mite, Tetranychus urticae (Acari: Tetranychidae) and the Predatory Mite, Neoseiulus californicus (Acari: Phytoseidae) in Organic strawberry Production Systems in Florida. J. Econ. Entomol. 2021, 114, 2135–2146. [Google Scholar] [CrossRef] [PubMed]
- Kheradmand, K.; Heidari, M.; Sedaratian-Jahromi, A.; Talaei-Hassanloui, R.; Havasi, M. Biological responses of Tetranychus urticae (Acari: Tetranychidae) to sub-lethal concentrations of the entomopathogenic fungus Beauveria bassiana. Bull. Entomol. Res. 2022, 112, 70–77. [Google Scholar] [CrossRef]
- Maniania, N.K.; Bugeme, D.M.; Wekesa, V.W.; Delalibera, I.; Knapp, M. Role of entomopathogenic fungi in the control of Tetranychus evansi and Tetranychus urticae (Acari: Tetranychidae), pests of horticultural crops. Exp. Appl. Acarol. 2008, 46, 259–274. [Google Scholar] [CrossRef]
- Mongkolsamrit, S.; Khonsanit, A.; Thanakitpipattana, D.; Tasanathai, K.; Noisripoom, W.; Lamlertthon, S.; Himaman, W.; Houbraken, J.; Samson, R.A.; Luangsa-Ard, J. Revisiting Metarhizium and the description of new species from Thailand. Stud. Mycol. 2020, 95, 171–251. [Google Scholar] [CrossRef]
- Chen, W.; Xie, W.; Cai, W.; Thaochan, N.; Hu, Q.B. Entomopathogenic fungi biodiversity in the soil of three provinces located in Southwest China and first approach to evaluate their biocontrol potential. J. Fungi 2021, 7, 984. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.K.; Sridhar, J.; Murali-Baskaran, R.K.; Senthil-Nathan, S.; Kaushal, P.; Dara, S.K.; Arthurs, S. Microbial biopesticides for insect pest management in India: Current status and future prospects. J. Invertebr. Pathol. 2019, 165, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Ullah, M.S.; Lim, U.T. Laboratory bioassay of Beauveria bassiana against Tetranychus urticae (Acari: Tetranychidae) on leaf discs and potted bean plants. Exp. Appl. Acarol. 2015, 65, 307–318. [Google Scholar] [CrossRef] [PubMed]
- Aak, A.; Hage, M.; Rukke, B.A. Biological control of Cimex lectularius with Beauveria bassiana: Effects of substrate, dosage, application strategy, and bed bug physiology. Pest Manag. Sci. 2023, 79, 4599–4606. [Google Scholar] [CrossRef] [PubMed]
- Al Khoury, C.; Guillot, J.; Nemer, N. Lethal activity of beauvericin, a Beauveria bassiana mycotoxin, against the two-spotted spider mites, Tetranychus urticae Koch. J. Appl. Entomol. 2019, 143, 974–983. [Google Scholar] [CrossRef]
- Rasool, S.; Markou, A.; Hannula, S.E.; Biere, A. Effects of tomato inoculation with the entomopathogenic fungus Metarhizium brunneum on spider mite resistance and the rhizosphere microbial community. Front. Microbiol. 2023, 14, 1197770. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Shi, W.B.; Feng, M.G. Histopathological and molecular insights into the ovicidal activities of two entomopathogenic fungi against two-spotted spider mite. J. Invertebr. Pathol. 2014, 117, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Parker, B.L.; Skinner, M.; Costa, S.D.; Gouli, S.; Reid, W.; El Bouhssini, M. Entomopathogenic fungi of Eurygaster integriceps Puton (Hemiptera: Scutelleridae): Collection and characterization for development. Biol. Control 2003, 27, 260–272. [Google Scholar] [CrossRef]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W. Fungal Barcoding Consortium, Fungal Barcoding Consortium Author List. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA bar-code marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [PubMed]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Hlina, B.L. ecotox: A Package for Analysis of Ecotoxicology. R Package Version 1.4.2. 2020. Available online: https://cran.r-project.org/package=ecotox (accessed on 14 September 2023).
- Hammami, W.; Chain, F.; Michaud, D.; Bélanger, R.R. Proteomic analysis of the metabolic adaptation of the biocontrol agent Pseudozyma flocculosa leading to glycolipid production. Proteome Sci. 2010, 8, 7. [Google Scholar] [CrossRef] [PubMed]
- Traquair, J.A.; Shaw, L.A.; Jarvis, W.R. New species of Stephanoascus with Sporothrix anamorphs. Can. J. Bot. 1988, 66, 926–933. [Google Scholar] [CrossRef]
- Bélanger, R.R.; Labbé, C.; Lefebvre, F.; Teichmann, B. Mode of action of biocontrol agents: All that glitters is not gold. Can. J. Plant Pathol. 2012, 34, 469–478. [Google Scholar] [CrossRef]
- Laur, J.; Ramakrishnan, G.B.; Labbé, C.; Lefebvre, F.; Spanu, P.D.; Bélanger, R.R. Effectors involved in fungal-fungal interaction lead to a rare phenomenon of hyperbiotrophy in the tritrophic system biocontrol agent-powdery mildew-plant. New Phytol. 2018, 217, 713–725. [Google Scholar] [CrossRef] [PubMed]
- Santhanam, P.; Madina, M.H.; Albuini, F.M.; Labbé, C.; Fietto, L.G.; Bélanger, R.R. A unique effector secreted by Pseudozyma flocculosa mediates its biocontrol activity. BMC Biol. 2023, 21, 118. [Google Scholar] [CrossRef] [PubMed]
- Dogan, Y.O.; Hazir, S.; Yildiz, A.; Butt, T.M.; Cakmak, I. Evaluation of entomopathogenic fungi for the control of Tetranychus urticae (Acari: Tetranychidae) and the effect of Metarhizium brunneum on the predatory mites (Acari: Phytoseiidae). Biol. Control 2017, 111, 6–12. [Google Scholar] [CrossRef]
- Tamai, M.A.; Alves, S.B.; Neves, P.J. Beauveria Bassiana PDE. Patogenicidade de beauveria bassiana (bals.) Vuill. Ao ácaro Tetranychus urticae koch. Sci. Agric. 1999, 56, 285–288. [Google Scholar] [CrossRef]
- de Souza, D.S.; Barth, A.I.; Berté, A.L.W.; Bizarro, G.L.; Heidrich, D.; da Silva, G.L.; Johann, L.; Maciel, M.J. Evaluation of the activity of filamentous fungi isolated from soils of the Pampa biome applied in the biological control of Tetranychus urticae (Acari: Tetranychidae) and Polyphagotarsonemus latus (Acari: Tarsonemidae). Exp. Appl. Acarol. 2021, 85, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.L.; Zhang, Y.; Xu, D.D.; Zhang, Y.J.; Wu, Q.J.; Xie, W.; Guo, Z.J. Inventors: Institute of Vegetables and Flowers, Chinese Academy of Agricultural Sciences, Assignee. A Pathogenic Fungus with Insecticidal Activity and Its Application. ZL 20211 1198819.X, 7 April 2023. [Google Scholar]
- Sáenz-de-Cabezón Irigaray, F.J.; Marco-Mancebón, V.; Pérez-Moreno, I. The entomopathogenic fungus Beauveria bassiana and its compatibility with triflumuron: Effects on the twospotted spider mite Tetranychus urticae. Biol. Control 2003, 26, 168–173. [Google Scholar] [CrossRef]
- Wekesa, V.W.; Knapp, M.; Maniania, N.K.; Boga, H.I. Effects of Beauveria bassiana and Metarhizium anisopliae on mortality, fecundity and egg fertility of Tetranychus evansi. J. Appl. Entomol. 2006, 130, 155–159. [Google Scholar] [CrossRef]
- Zhang, X.; Jin, D.C.; Zou, X.; Guo, J. Laboratory and field evaluation of an entomopathogenic fungus, Isaria cateniannulata strain 08XS-1, against Tetranychus urticae (Koch). Pest Manag. Sci. 2016, 72, 1059–1066. [Google Scholar] [CrossRef] [PubMed]
- Bugeme, D.M.; Knapp, M.; Boga, H.I.; Ekesi, S.; Maniania, N.K. Susceptibility of developmental stages of Tetranychus urticae (Acari: Tetranychidae) to infection by Beauveria bassiana and Metarhizium anisopliae (Hypocreales: Clavicipitaceae). Int. J. Trop. Insect Sci. 2014, 34, 190–196. [Google Scholar]
- Vey, A.; Fargues, J. Histological and ultrastructural studies of Beauveria bassiana infection in Leptinotarsa decemlineta larvae during ecdysis. J Invertebr Pathol 1977, 30, 207–215. [Google Scholar] [CrossRef]
- Catalán, T.P.; Wozniak, A.; Niemeyer, H.M.; Kalergis, A.M.; Bozinovic, F. Interplay between thermal and immune ecology: Effect of environmental temperature on insect immune response and energetic costs after an immune challenge. J. Insect Physiol. 2012, 58, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Nie, P.C.; Yang, R.L.; Zhou, J.J.; Dewer, Y.; Shang, S.Q. Elucidating the Effect of temperature Stress on the Protein Content, Total Antioxidant Capacity, and Antioxidant Enzyme Activities in Tetranychus urticae (Acari: Tetranychidae). Insects 2023, 14, 429. [Google Scholar] [CrossRef]
- Kutch, I.C.; Sevgili, H.; Wittman, T.; Fedorka, K.M. Thermoregulatory strategy may shape immune investment in Drosophila melanogaster. J. Exp. Biol. 2014, 217, 3664–3669. [Google Scholar] [CrossRef]
- Zaki, O.; Weekers, F.; Compere, P.; Jacques, P.; Thonart, P.; Sabri, A. Morphological differences between aerial and submerged sporidia of bio-fongicide Pseudozyma flocculosa CBS 16788. PLoS ONE 2018, 13, e0201677. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentration (Conidia/mL) | N 1 | Corrected Mortality Rate | LT50 (95% FL 2) 3 | Slope (±SE) 4 | Chi-Square (df) | Heterogeneity | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1d | 2d | 3d | 4d | 5d | 6d | 7d | 8d | 9d | ||||||
| 8 × 106 | 102 | 7.97% | 6.04% | 30.61% | 68.34% | 75.13% | 78.27% | 85.94% | 95.20% | 100.00% | 3.15 (2.47–3.77) ab | 3.73 ± 0.22 | 39.54 (7) | 5.65 |
| 8 × 107 | 92 | 3.57% | 13.38% | 51.48% | 74.58% | 78.69% | 88.59% | 92.21% | 97.34% | 100.00% | 3.05 (2.81–3.28) b | 3.43 ± 0.21 | 6.62 (7) | 1.00 |
| 8 × 108 | 100 | 14.57% | 19.23% | 62.57% | 77.38% | 91.89% | 91.80% | 92.53% | 96.17% | 100.00% | 2.39 (1.99–2.77) a | 3.78 ± 0.22 | 19.32 (7) | 2.76 |
| 8 × 109 | 94 | 16.60% | 18.66% | 64.09% | 82.23% | 88.98% | 92.55% | 93.64% | 98.70% | 100.00% | 2.26 (1.79–2.69) a | 3.59 ± 0.22 | 24.05 (7) | 3.44 |
| Temperature | 5 × 107 | 5 × 109 | ||||
|---|---|---|---|---|---|---|
| N 1 | LT50 (95% FL 2) 3 | Slope (±SE) 4 | N 1 | LT50 (95% FL 2) 3 | Slope (±SE) 4 | |
| 24 °C | 78 | 3.12 (2.14–4.07) c | 3.75 ± 0.29 | 80 | 2.52 (1.60–3.35) c | 3.22 ± 0.25 |
| 26 °C | 91 | 2.69 (1.68–3.60) bc | 3.62 ± 0.26 | 91 | 1.87 (1.11–2.51) bc | 3.18 ± 0.24 |
| 28 °C | 71 | 1.78 (1.57–1.98) b | 3.51 ± 0.31 | 106 | 1.44 (1.28–1.58) b | 3.61 ± 0.28 |
| 30 °C | 69 | 1.06 (0.32–1.56) a | 2.87 ± 0.36 | 96 | 0.81 (0.61–0.97) a | 3.03 ± 0.37 |
| 34 °C | 80 | 1.00 (0.75–1.19) a | 2.90 ± 0.48 | 103 | 0.65 (0.36–0.87) a | 2.28 ± 0.45 |
| 36 °C | 78 | 1.17 (0.99–1.33) a | 3.74 ± 0.51 | 85 | 0.53 (0.17–0.80) a | 1.95 ± 0.49 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Dong, R.; Hu, S.; Guo, Z.; Wang, S. Identification of an Entomopathogenic Fungus, Pseudozyma flocculosa (Traquair, Shaw & Jarvis), and Its Efficacy against Tetranychus urticae Koch. Horticulturae 2024, 10, 221. https://doi.org/10.3390/horticulturae10030221
Zhang Y, Dong R, Hu S, Guo Z, Wang S. Identification of an Entomopathogenic Fungus, Pseudozyma flocculosa (Traquair, Shaw & Jarvis), and Its Efficacy against Tetranychus urticae Koch. Horticulturae. 2024; 10(3):221. https://doi.org/10.3390/horticulturae10030221
Chicago/Turabian StyleZhang, Yan, Rui Dong, Shouyin Hu, Zhaojiang Guo, and Shaoli Wang. 2024. "Identification of an Entomopathogenic Fungus, Pseudozyma flocculosa (Traquair, Shaw & Jarvis), and Its Efficacy against Tetranychus urticae Koch" Horticulturae 10, no. 3: 221. https://doi.org/10.3390/horticulturae10030221