Abstract
Bacillus strain PE1, which was isolated from potatoes harvested in the Yaqui Valley, Mexico, was evaluated as a potential biological control agent against Fusarium languescens. The draft genome sequence was obtained through Illumina NovaSeq sequencing, revealing a genomic size of 4,071,293 bp, with a G + C content of 44.13%, an N50 value of 357,305 bp, and 27 contigs. The taxonomic affiliation was confirmed by analyzing the 16S rRNA gene and overall genome relatedness indices (OGRIs) and constructing a phylogenomic tree based on the whole genome, which showed a close relationship to Bacillus cabrialesii subsp. cabrialesii. Genomic annotation using RAST and Prokka identified 4261 coding DNA sequences (CDSs) distributed across 331 subsystems, highlighting genes associated with biocontrol, stress response, and iron acquisition. AntiSMASH 7.1 was used for genome mining, revealing seven biosynthetic gene clusters that potentially produce biocontrol-related metabolites. In vitro assays confirmed the antagonistic activity of strain PE1 against Fusarium languescens CE2, demonstrating its potential to inhibit mycelial growth. The study provides a genomic basis for investigating B. cabrialesii subsp. cabrialesii PE1 as a potential biological control agent in potato production.
1. Introduction
The potato (Solanum tuberosum L.) is a widely cultivated crop and is one of the top five most produced crops worldwide [1]. However, unfavorable conditions such as diseases can make potato production unsustainable, negatively impacting productivity and yield [2]. Fusarium is one of the most severe plant pathogens that attack potatoes, following late blight. Fusarium-related diseases are serious soil-borne diseases that cause economic losses worldwide. They can affect potatoes at any growth stage by inducing Fusarium wilt on plants and Fusarium dry rot on tubers [3]. Several Fusarium species have been associated with potatoes, mainly Fusarium sambucinum, F. solani, F. graminearum, F. oxysporum, F. verticillioides, and F. oxysporum f. sp. tuberosi [4,5]. Fusarium species cause different symptoms in potatoes. Fusarium oxysporum f. sp. tuberosi causes vascular wilt, and F. solani and F. sambucinum lead to Fusarium dry rot in tubers [3,4,5,6]. Fusarium diseases are caused by individual Fusarium or co-occurring species [7]. The relative importance of the Fusarium species varies depending on factors such as local climate, agricultural practices, and host susceptibility [8]. For instance, according to Montoya-Martinez and Cota-Barreras [9,10], Fusarium languescens is a significant regional phytopathogen in Sonora, Mexico, that could impact potato production.
Currently, new alternative control methods to synthetic fungicides are being considered for sustainable agricultural systems, due to the development of fungicide resistance by Fusarium strains [11,12,13], as well as the negative impact of these synthetic compounds on the environment and health [14]. Thus, a promising approach that has gained attention is the use of biological control agents (BCAs) [15,16]. Reports suggest that bacterial antagonists can be used to combat phytopathogenic Fusarium species, for example, the inoculation of Pseudomonas fluorescens to potato tuber seeds [5]. Similarly, postharvest studies using P. fluorescens and Bacillus subtilis have shown antagonistic properties against dry rot caused by F. sambucinum and F. solani, respectively [17].
Bacillus is a predominant bacterial genus, and numerous Bacillus species have been reported as biocontrol agents [18,19]. Bacillus uses various direct and indirect mechanisms to promote plant growth and control pathogen proliferation. These mechanisms include solubilization and mineralization of nutrients such as phosphorus and potassium, nitrogen fixation, production of 1-aminocyclopropne-1-carboxlyic acid (ACC), phytohormones, antimicrobial compounds, hydrolytic enzymes, and siderophores, as well as abiotic-stress tolerance [18,20]. However, identifying appropriate strains of potential biocontrol agents and characterizing associated metabolites remains a complex and arduous task [21,22,23]. To address this issue, bioinformatics tools such as genome mining for biosynthetic genes can be used to rapidly predict the secondary metabolites produced by a BCA strain [22,23,24]. Thus, the identification of genetic blueprints is essential for recognizing bioactive secondary metabolites and enzymes that mediate most biocontrol mechanisms [22].
In this context, there is a need to reduce the heavy reliance on the chemical control of Fusarium diseases and explore the use of BCAs as a sustainable alternative by effectively searching for beneficial strains [25]. This study aims to explore the ability of Bacillus cabrialesii subsp. cabrialesii strain PE1—isolated from potatoes—as a promising biological control agent against Fusarium languescens CE2, a regional phytopathogen. Both microorganisms were isolated from a commercial field in the Yaqui Valley, Mexico, the birthplace of the Green Revolution in the 1950s. This strategy was carried out by sequencing, annotation, and mining of the strain PE1 genome to identify biosynthetic gene clusters associated with its biocontrol ability; in addition, this genomic insight was supported by testing the antagonistic activity of extracellular metabolites produced by strain PE1 against F. languescens CE2.
2. Materials and Methods
2.1. Bacteria Isolation and Culture Conditions
The bacterial strain PE1 was isolated from potatoes harvested from a commercial field in the Yaqui Valley, Mexico (27°17′43.7″ N 109°51′44.1″ W). The site was selected because the plants exhibited symptoms of disease. Potatoes without symptoms were selected for sampling. The isolation process involved superficial disinfection of potatoes using commercial sodium hypochlorite at a concentration of 1.5% for 15 min. Then, disinfected potatoes were thrice washed with sterilized water before being cut and macerated. The serial dilution method (1:10) was subsequently employed up to 10−6. Thus, 1 mL of each dilution was evenly spread on nutrient agar (NA) Petri dishes and incubated at 28 °C for 2 days. After incubation, strain PE1 was isolated, purified, and characterized based on its morphological traits, such as cell and colony shape, color, elevation, and opacity. Following these assessments, the strain was cryopreserved at −80 °C, using a nutrient broth (NB) culture medium containing glycerol (30%), at the Colección de Microorganismos Edáficos y Endófitos Nativos (COLMENA, itson.edu.mx/micrositios/COLMENA, accessed on 18 February 2024).
2.2. Genomic Analysis
High-quality genomic DNA was extracted from a fresh culture of strain PE1 grown in nutrient broth (NB), under growth conditions set at 30 °C for 24 h and at 121 rpm, obtaining 1 × 106 Colony Forming Units (CFU/mL). Thus, 40 µL of the cell suspension was lysed with 120 μL of TE buffer containing lysozyme (final concentration 0.1 mg/mL) and RNase A (final concentration 0.1 mg/mL), incubated for 25 min at 37 °C. Proteinase K (final concentration 0.1 mg/mL) and SDS (final concentration 0.5% v/v) were added and incubated for 5 min at 65 °C. Genomic DNA was purified using an equal volume of SPRI beads and resuspended in EB buffer (10 mM Tris-HCl, pH 8.0). The extracted total DNA (DNA ≥ 1 μg, concentration ≥ 20 ng/μL) was then quantified with the Quant-iT dsDNA HS kit (ThermoFisher Scientific) assay in a plate reader and diluted as appropriate.
The Illumina NovaSeq platform (2 × 250 bp) was used for DNA sequencing, and library preparation was carried out using the Nextera XT Library Prep Kit, following the manufacturer’s protocol, but with the following modifications: input DNA was increased 2-fold, and PCR elongation time was increased to 45 s. DNA quantification and library preparation were carried out on a Hamilton Microlab STAR automated liquid handling system (Hamilton Bonaduz AG, Reno, Nevada, USA). Libraries were sequenced on an Illumina NovaSeq 6000 (Illumina, San Diego, CA, USA), using a 250 bp paired-end protocol. Genomic information analysis was performed following the workflow reported by Ortega-Urquieta, 2022 [26]. Trimmomatic version 0.30 was used to remove adapter sequences and eliminate low-quality bases. The SPAdes version 3.15.4 generated a de novo assembly, and the contigs were ordered regarding the genome of Bacillus cabrialesii subsp. cabrialesii TE3T (GenBank accession number GCA_004124315.2), using Mauve Contig Mover version 2.4.0. Plasmid detection was performed with PlasmidFinder 2.1. The genome sequence of strain PE1 was analyzed for contamination using CheckM version 1.0.18 [27].
To affiliate the strain PE1 at a species level, its genome was compared to its more closely related strains (Table 1; 16S rRNA similarity ≥ 99.6%) by using the overall genome relatedness indices (OGRIs): average nucleotide identity (ANI) by the OrthoANI algorithm [28] and the Genome to Genome Distance Calculator (GGDC) version 3.0 by BLAST [29]. A whole-genome-based phylogenetic tree was constructed using Type (strain) Genome Server (TYGA) (https://tygs.dsmz.de/, accessed on 10 January 2024) [29].

Table 1.
16S rRNA gene and OGRIs-based taxonomic affiliation of strain PE1.
2.3. Genome Annotation and Genome Mining
The genome annotation for strain PE1 was performed using the Rapid Annotation Using Subsystem Technology (RAST) server version 2.0 and the RASTtk pipeline based on the PathoSystems Resource Integration Center (PATRIC). A second annotation was conducted using Proksee, which incorporates the Rapid Prokaryotic Genome Annotation (Prokka), and this process generated the circular chromosome map of strain PE1, including coding sequences (CDSs), tRNAs, rRNAs, guanine–cytosine (GC), and skew content. To identify biosynthetic gene clusters associated with biocontrol, the genome of strain PE1 was submitted to the Antibiotics & Secondary Metabolite Analysis Shell (AntiSMASH) 7.1 web server (https://antismash.secondarymetabolites.org, accessed on 18 February 2024) [30], under the ‘relaxed’ parameter.
2.4. Evaluation of the Antagonistic Activity of Strain PE1 against Fusarium languescens CE2 through Extracellular Metabolites
Fusarium languescens CE2 was obtained from a national microbial culture collection, named COLMENA (Coleccion de Microorganismos Edaficos y Endofitos Nativos, [31]). This is a phytopathogenic strain previously isolated from the Yaqui Valley, Mexico [9]. In this study, the strain CE2 was used as a model for Fusarium. This assay was carried out following the methods described by Montoya-Martinez et al. (2023) [9] and Baard et al. (2023) [32], with modifications. Thus, a liquid bacterial culture was prepared by inoculating a 1 × 104 CFU of strain PE1 into 20 mL of NB. The culture was incubated at 30 °C, with constant shaking at 120 rpm, for 72 h. Afterward, the bacterial culture was centrifuged at 5000 rpm for 10 min, and the resulting supernatant was collected and filtered through a hydrophilic syringe filter (0.22 µm). The effectiveness of the cell-free supernatant (CF) against F. languescens CE2 was tested using two methods: a well-diffusion method and casting agar plates with 50% CF. To perform the well-diffusion method, a 0.6 cm diameter agar plug containing growing mycelia from F. languescens CE2 was placed at the center of a fresh Potato Dextrose Agar (PDA) plate. Then, three paper discs, each with a diameter of 0.6 cm, were placed equidistantly on the plate, and 25 μL of cell-free supernatant was added over each disc. The plates were sealed and incubated at 30 °C for three days. The antagonistic effect of the cell-free supernatant on F. languescens CE2 mycelial growth was measured in terms of area (mm2) using ImageJ 1.54g [33]. For the second method, agar plates with 50% cell-free supernatant were used; thus, Potato Dextrose Agar (PDA) was prepared at double concentration and sterilized, and then the cell-free supernatant was added to a 50% concentration v/v. Then, a 0.6 cm diameter agar plug containing growing mycelia from F. languescens CE2 was placed at the center of each plate. The inoculated plates were then incubated at 30 °C for six days, and the area (mm2) with mycelial growth was measured. For the control treatment, the cell-free supernatant was replaced with sterile distilled water.
The experiment was conducted with three replicates. Statistical analyses were performed using R Studio Version 4.3.0 [34]. Data were analyzed using a one-way ANOVA (p ≤ 0.05). The presented values represent the mean between replicates, and the bars represent the standard deviation (SD).
3. Results
3.1. Genomic Analysis
The bacterial DNA was subjected to sequencing, yielding a total of 1,017,168 paired-end reads (2 × 250 bp). Subsequent assembly of the obtained reads resulted in the draft genome of strain PE1, comprising 27 contigs (≥500 bp). The assembled genome encompassed a total length of 4,071,293 bp, with a G + C content of 44.13%, an N50 value of 357, 305 bp, and 5 L50. Importantly, plasmids were not detected within the genome of strain PE1, indicating its genomic stability and absence of extrachromosomal genetic elements.
Based on the 16S rRNA gene, strain PE1 was taxonomically affiliated with the genus Bacillus, showing 100% similarity to Bacillus cabrialesii subsp. cabrialesii TE3T, 100% to B. inaquosorum KCTC 13429 T, and 99.86% to B. tequilensis KCTC 13622 T (Table 1). Based on the OGRIs analysis, this strain was strongly affiliated with Bacillus cabrialesii subsp. cabrialesii due to these values being higher than those delimiting the species affiliation (ANI ≥ 95–96% and GGDC ≥ 70%) (Table 1). Finally, this taxonomic affiliation was confirmed by the construction of a whole-genome-based phylogenomic tree (Figure 1), showcasing the close evolutionary relationship of strain PE1. Thus, this strain belongs to Bacillus cabrialesii subsp. cabrialesii.
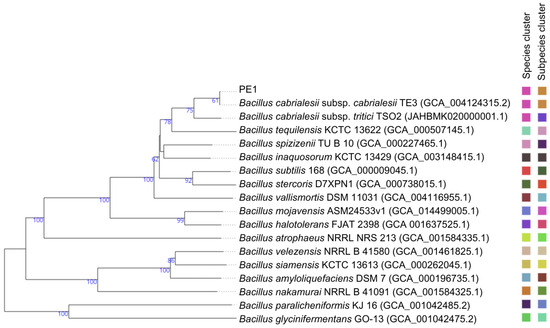
Figure 1.
Phylogenetic relationship between strain PE1 and closely related species based on genome sequences constructed by TYGS. Tree inferred with FastME 2.1.6.1 [35] from GBDP distances calculated from genome sequences. The branch lengths are scaled in terms of the GBDP distance formula d5. The numbers above the branches are GBDP pseudo-bootstrap support values > 60% from 100 replications, with an average branch support of 73.0%. The tree was rooted at the midpoint [36].
3.2. Genome Annotation
According to the RAST prediction, the genome of B. cabrialesii subsp. cabrialesii PE1 contains 94 RNA sequences and 4261 protein-coding DNA sequences (CDSs) distributed across 331 subsystems, as shown in Figure 2. Notable subsystems in the genome of strain PE1 include coding sequences related to biocontrol, such as virulence, disease, and defense (36 CDSs). These are further subdivided into resistance to antibiotics and toxic compounds, invasion and intracellular resistance, and bacteriocins and ribosomally synthesized antibacterial peptides. Additionally, the iron acquisition and metabolism (30 CDSs) subsystem includes genes related to siderophores, which function as iron chelators, decreasing the accessibility of iron to phytopathogenic microorganisms. The carbohydrates subsystem (272 CDSs) contains coding sequences related to the production of acetoin and butanediol. These volatile components are capable of inducing systemic resistance in plants [37]. The cell wall and capsule (86 CDSs) subsystem contains coding sequences for exopolysaccharide biosynthesis (EpsC and EpsD), which are genes related to colonization [22]. Additionally, subsystems related to bacterial resilience for the development of agricultural bioproducts were identified, including the stress response (43 CDSs), which covers osmotic and oxidative stress. Furthermore, complementing these results, the circular chromosome map based on Prokka predicted a total of 5395 CDSs, 130 tRNAs, and 1 tmRNA (Figure 3).
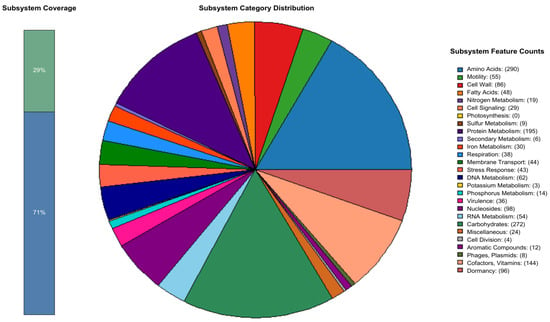
Figure 2.
Pie chart of the subsystem category distribution of CDSs from Bacillus cabrialesii subsp. cabrialesii PE1 constructed by RAST server version 2.0 and R studio version 4.3.0. CDSs, 4261; and subsystems, 331.
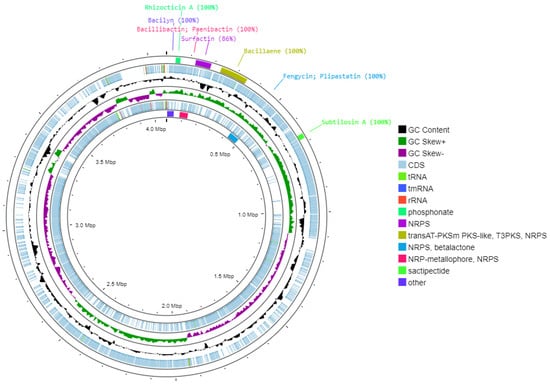
Figure 3.
Circular chromosome map of Bacillus cabrialesii subsp. cabrialesii PE1, including the distribution of CDS, tRNAs, rRNAs, and GC content skew generated through genome annotation from PROKKA. Additionally, the identified biosynthetic gene clusters associated with biocontrol by AntiSMASH [30] are also shown.
3.3. Genome Mining
The AntiSMASH 7.1 web server was used for genome mining on Bacillus cabrialesii subsp. cabrialesii PE1, revealing seven distinct genomic regions. Notable biosynthetic gene clusters were detected in regions 5, 6, 14, 20, and 21 (Table 2 and Figure 3).
Table 2.
Biosynthetic gene clusters found in B. cabrialesii subsp. cabrialesii PE1 obtained from genome mining by the antiSMASH web server.
3.4. Antagonistic Activity of Bacillus cabrialesii subsp. cabrialesii PE1 against Fusarium Languescens CE2, through Extracellular Metabolites
Extracellular metabolites produced by B. cabrialesii subsp. cabrialesii PE1 significantly inhibited the growth of F. languescens CE2, as demonstrated by the reduction of 41.6% in the mycelial growth area, from 969.60 ± 55.72 mm2 (control treatment) to 566.20 ± 29.54 mm2 (Figure 4a). On the other hand, 50% of the cell-free supernatant (CF) obtained from B. cabrialesii subsp. cabrialesii PE1 also showed a significant inhibitory effect on the mycelial growth of F. languescens CE2, from 1835.18 ± 125.09 mm2 (control treatment) to 83.31 ± 6.32 mm2 95.4% (Figure 4b).
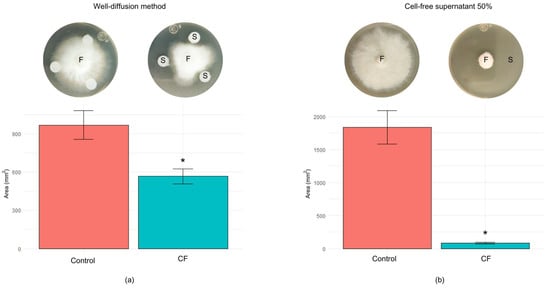
Figure 4.
Antagonistic activity of extracellular metabolites from B. cabrialesii subsp. cabrialesii PE1 against F. languescens CE2 in PDA: (a) well-diffusion method, 3 days of incubation; and (b) cell-free supernatant 50% v/v, 6 days of incubation. The antagonistic effect of the cell-free supernatant on F. languescens CE2 mycelial growth was measured in terms of area (mm2). * Statistically significant difference (p ≤ 0.05). F: F. languescens CE2. S: cell-free supernatant from B. cabrialesii subsp. cabrialesii PE1.
4. Discussion
Sustainable plant disease management faces the challenge of meeting the global demand for safe and diversified food. Thus, bioprospecting is a key tool to reach this goal, which involves isolating microorganisms from the habitat where they will be used and directly screening their potential against phytopathogens, under specific edaphoclimatic conditions [38,39]. Strain PE1 was isolated and genomically characterized under this objective. This work establishes the basis for studying and evaluating Bacillus cabrialesii subsp. cabrialesii PE1 as a biological control agent against Fusarium languescens in potato production in the Yaqui Valley, Mexico.
Biological control offers several opportunities for improved disease control, particularly where conventional approaches are limited or compromised; this is particularly relevant for Fusarium-related diseases, where chemical resistance and the potential harm to human and environmental health from fungicide use are major concerns [5,40]. In this sense, there are four main routes of action underlying biological control of plant diseases: (i) exploitation competition for resources, (ii) interference competition for space via antibiosis or toxic secondary metabolites, (iii) hyperparasitism, and (iv) induced resistance [39]. Bacillus cabrialesii subsp. cabrialesii PE1 demonstrated biocontrol potential against F. languescens CE2 by producing secondary metabolites, including potentially subtilosin A, bacillibactin, bacillaene, fengycin, surfactin, bacilyn, and rhizocticin A, as revealed by genome mining (Figure 2 and Figure 3; Table 2). The genus Bacillus is well-characterized and exhibits multiple beneficial properties in plant nutrition and antimicrobial activity against phytopathogens. Among diverse species of this genus, B. velezensis, B. subtilis, B. amyloliquefaciens, and B. cereus have been reported to be active against Fusarium [41,42]. Additionally, Bacillus cabrialesii subsp. Tritici TSO2T and Bacillus cabrialesii subsp. cabrialesii TE3T have been reported as biocontrol agents [9,43]. The biocontrol capacity of these biological control strains is largely attributed to their ability to produce extracellular enzymes (cellulase, glucanases, proteases, chitinases, beta-xylosidase, chitin deacetylases, catalase, peroxide dismutase, and peroxidase); and antimicrobial secondary metabolites such as organic compounds (1,2-benzenedicarboxylic acid, 6-undecylamine, 2-methyloctacosane, 9-octadecenoic acid and 1-tetradecanamine, and N,N-dimethyl), lipopeptides (mycosubtilin, fengycin B, iturin A, surfactin A, iturin, bacillomycin, bacilysin, and mersacidin) and siderophores [41,44].
In vitro assays of B. cabrialesii subsp. cabrialesii PE1 demonstrated a reduction in the growth of F. languescens CE2 by 41.6% and 95.4%, respectively, depending on the concentration strategy using CF (Figure 4). This validates the production and diversity of secondary metabolites produced by B. cabrialesii subsp. cabrialesii PE1 and highlights the importance of exploring the action modes to quantify their roles in the biocontrol efficacy [45]. Similar metabolites found in the genome information have been associated with Fusarium biocontrol. For instance, Bacillus subtilis SG6 has been reported to act against F. graminearum by producing fengycins and surfactins [42]. Additionally, under field conditions, Bacillus velezensis LM2303 inhibited F. graminearum, where the observed inhibition was attributed to the presence of three antifungal metabolites (fengycin B, iturin A, and surfactin A) and eight antibacterial metabolites, including bacillaene [44], which were also identified in the strain PE1 genome. Another report showed strong antifungal activity by disrupting the cell walls, membranes, and cytoskeleton of Fusarium oxysporum f. sp. cucumerinum hyphae due to plipastatin [46], which was found in region 14.2 of the strain PE1 genome. Thus, the production of lipopeptides such as fengycin/plipastatin and surfactin, as well as polyketides such as bacillaene by strain PE1, may be involved in the demonstrated biocontrol activity in the in vitro assay. However, further deeper studies, such as metabolomics, are necessary to complement these findings.
Additionally, the iron acquisition and metabolism subsystem in the genome of B. cabrialesii subsp. cabrialesii PE1 includes 15 CDSs related to siderophores, which are iron chelators that decrease the accessibility of iron to phytopathogenic microorganisms. Siderophores are classified into three functional groups based on their structure: hydroxamate, catecholate, and carboxylate [47]. Bacillus sp. produces catecholate as its main siderophore, including bacillibactin [48], which has also been identified through genome mining in strain PE1. Siderophores play a crucial role in the biological control of Fusarium wilt of pepper [49], and they could be applicable in the case of biocontrol of Fusarium affecting potato.
The genome annotation also suggests that strain PE1 may possess additional bio-control mechanisms. For example, the strain may produce acetoin and butanediol, which are volatile compounds that can induce systemic resistance in plants [37]. Furthermore, the strain may be associated with exopolysaccharide biosynthesis, which enhances colonization and biocontrol [22]. In this context, strain PE1 may have multiple biocontrol mechanisms that could be expressed based on the environment. Identifying the responsible mechanisms is a significant challenge due to the involvement of biotic and abiotic factors that can affect the performance of biocontrol agents and their mechanisms of action [50]. In many cases, complex interactions between plants, biocontrol agents, and pathogens involve various mechanisms [51,52]. Characterizing the genome of selected strains is a crucial step in developing biological control agents (BCAs), as it provides valuable information about their potential as biopesticides [22,52]. This process allows further analysis of strains based on their key attributes. Identifying and characterizing the genotypic traits of strains is essential in determining their potential as biocontrol agents.
5. Conclusions
This research showcases the biocontrol capabilities of Bacillus cabrialesii subsp. cabrialesii PE1 against the causal agent that affects potato cultivation, Fusarium languescens CE2. Genome mining identified seven biosynthetic gene clusters associated with biocontrol within this strain, providing insights into potential genomic determinants of its biocontrol ability. The in vitro assessments suggest that lipopeptides, such as fengycin and surfactin, as well as polyketides like bacillaene, may contribute to the observed biocontrol activity. However, further investigations are required, particularly in the field of metabolomics, to improve and refine these findings. Additionally, exploring the diverse functional activities and genes of strain PE1 is crucial to effectively use it as a biocontrol agent in the field.
Author Contributions
Conceptualization, S.d.l.S.-V. and B.V.-A.; methodology, formal analysis, all authors, and data curation, B.V.-A. and A.C.M.-M.; writing—original draft preparation, all authors; writing—review and editing, all authors; supervision, project administration, and funding acquisition, S.d.l.S.-V. All authors have read and agreed to the published version of the manuscript.
Funding
We acknowledge funding from GetGenome (a newly formed charitable organization that provides equitable access to genomics technology for early career researchers all over the world) and the Instituto Tecnológico de Sonora (PROFAPI_2024_0001). In addition, A.C.M.-M. was funded by Consejo Nacional de Humanidades, Ciencias y Tecnologías (CONAHCYT) for a postdoctoral fellowship (application number: 2306476).
Data Availability Statement
The complete genome sequence was deposited in DDBJ/ENA/GenBank and is openly available in NCBI under accession number JBAMMW000000000, under BioProject number PRJNA1080047, and BioSample number SAMN40099177.
Acknowledgments
The authors thank all members of the LBRM-COLMENA for their support in the development of this research project, mainly Abraham Ruíz Castrejón.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Calıskan, M.E.; Bakhsh, A.; Jabran, K. (Eds.) Potato Production Worldwide; Academic Press: Cambridge, MA, USA, 2022. [Google Scholar]
- Waheed, A.; Chuang, L.; Murad, M.; Mushtaq, A.; Khalid, A.K.; Hamed, A.; Ghramh Zhongwei, W.; Daoyuan, Z. Sustainable Potato Growth under Straw Mulching Practices. Sustainability 2023, 15, 10442. [Google Scholar] [CrossRef]
- Khedher, S.B.; Mejdoub-Trabelsi, B.; Tounsi, S. Biological potential of Bacillus subtilis V26 for the control of Fusarium wilt and tuber dry rot on potato caused by Fusarium species and the promotion of plant growth. Biol. Control. 2021, 152, 104444. [Google Scholar] [CrossRef]
- García-Ávila, C.J.; Valenzuela-Tirado, G.A.; Florencio Anastasio, J.G.; Ruiz-Galván, I.; Moreno-Velázquez, M.; Hernández-Macías, B.; López-Buenfil, J.A.; Bravo-Pérez, D.; Pineda-Ríos, J.M.; Quezada-Salinas, A.; et al. Organisms associated with damage to post-harvest potato tubers. Rev. Mex. Fitopatol. 2018, 36, 308–320. [Google Scholar]
- Lahlali, R.; Ezrari, S.; Radouane, N.; Kenfaoui, J.; Esmaeel, Q.; El Hamss, H.; Belabess, Z.; Barka, E.A. Biological Control of Plant Pathogens: A Global Perspective. Microorganisms 2022, 10, 596. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, R.K.; Kumar, R.; Sharma, S.; Sagar, V.; Aggarwal, R.; Naga, K.C.; Lal, M.C.; Chourasia, K.N.; Kumar, D.; Kumar, M. Potato dry rot disease: Current status, pathogenomics and management. 3 Biotech 2020, 10, 503. [Google Scholar] [CrossRef] [PubMed]
- Ferrigo, D.; Raiola, A.; Causin, R. Fusarium toxins in cereals: Occurrence, legislation, factors promoting the appearance and their management. Molecules 2016, 21, 627. [Google Scholar] [CrossRef] [PubMed]
- Pfordt, A.; Ramos Romero, L.; Schiwek, S.; Karlovsky, P.; von Tiedemann, A. Impact of environmental conditions and agronomic practices on the prevalence of Fusarium species associated with ear-and stalk rot in maize. Pathogens 2020, 9, 236. [Google Scholar] [CrossRef]
- Montoya-Martínez, A.C.; Figueroa-Brambila, K.M.; Escalante-Beltrán, A.; López-Montoya, N.D.; Valenzuela-Ruíz, V.; Parra-Cota, F.I.; Estrada Alvarado, M.I.; de los Santos-Villalobos, S. Biological Control Mechanisms of Bacillus cabrialesii subsp. tritici TSO2T against Fusarium languescens, the Causal Agent of Wilt in Jalapeño Peppers. Horticulturae 2023, 9, 964. [Google Scholar] [CrossRef]
- Cota-Barreras, C.I.; García-Estrada, R.S.; León-Félix, J.; Valenzuela-Herrera, V.; Mora-Romero, G.A.; Leyva-Madrigal, K.Y.; Tovar-Pedraza, J.M. Phylogeny, distribution, and pathogenicity of fusarioid fungi associated with chickpea wilt in Sinaloa and Sonora, Mexico. Available online: https://doi.org/10.21203/rs.3.rs-2960826/v1 (accessed on 7 July 2023).
- de Chaves, M.A.; Reginatto, P.; da Costa, B.S.; de Paschoal, R.I.; Teixeira, M.L.; Fuentefria, A.M. Fungicide resistance in Fusarium graminearum species complex. Curr. Microbiol. 2022, 79, 62. [Google Scholar] [CrossRef]
- Mao, Y.; Zhang, Z.; Shen, J.; Yin, X.; Wang, T.; Zheng, X.; Duan, Y. The intrinsic resistance of Fusarium solani to the Fusarium-specific fungicide phenamacril is attributed to the natural variation of both T218S and K376M in myosin5. Pestic. Biochem. Physiol. 2023, 196, 105595. [Google Scholar] [CrossRef]
- Zhang, N.; Xu, Y.; Zhang, Q.; Zhao, L.; Zhu, Y.; Wu, Y.; Li, Z.; Yang, W. Detection of fungicide resistance to fludioxonil and tebuconazole in Fusarium pseudograminearum, the causal agent of Fusarium crown rot in wheat. PeerJ 2023, 11, e14705. [Google Scholar] [CrossRef] [PubMed]
- Mir, S.A.; Padhiary, A.; Ekka, N.J.; Baitharu, I.; Nayak, B. Environmental impacts of synthetic and biofungicides. In Current Developments in Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2023; pp. 487–504. [Google Scholar] [CrossRef]
- Sulaiman, M.A.; Bello, S.K. Biological control of soil-borne pathogens in arid lands: A review. J. Plant Dis. Prot. 2023, 131, 293–313. [Google Scholar] [CrossRef]
- Ramlawi, S.; Chiu, J.O.; Cloutier, A.; Avis, T.J. Suppression of Fusarium dry rot of potato using beneficial bacterial treatments. J. Plant Pathol. 2021, 103, 269–281. [Google Scholar] [CrossRef]
- Mnif, I.; Hammami, I.; Triki, M.A.; Azabou, M.C.; Ellouze-Chaabouni, S.; Ghribi, D. Antifungal efficiency of a lipopeptide biosurfactant derived from Bacillus subtilis SPB1 versus the phytopathogenic fungus, Fusarium solani. Environ. Sci. Pollut. Res. Int. 2015, 22, 18137–18147. [Google Scholar] [CrossRef] [PubMed]
- Etesami, H.; Jeong, B.R.; Glick, B.R. Biocontrol of plant diseases by Bacillus spp. Physiol. Mol. Plant Pathol. 2023, 126, 102048. [Google Scholar] [CrossRef]
- Chen, K.; Tian, Z.; He, H.; Long, C.A.; Jiang, F. Bacillus species as potential biocontrol agents against citrus diseases. Biol. Control. 2020, 151, 104419. [Google Scholar] [CrossRef]
- Miljaković, D.; Marinković, J.; Balešević-Tubić, S. The significance of Bacillus spp. in disease suppression and growth promotion of field and vegetable crops. Microorganisms 2020, 8, 1037. [Google Scholar] [CrossRef] [PubMed]
- Shimbori, E.M.; Querino, R.B.; Costa, V.A.; Zucchi, R.A. Taxonomy and biological control: New challenges in an old relationship. Neotrop. Entomol. 2023, 52, 351–372. [Google Scholar] [CrossRef]
- Bertê, R.; Teixeira, G.M.; de Oliveira, J.P.; Nicoletto, M.L.A.; da Silva, D.V.; de Godoy, G.G.; Sanches, D.S.; de Resende, J.T.V.; Pereira, U.d.P.; da Rocha, U.N.; et al. Genome mining reveals high biosynthetic potential of biocontrol agent Bacillus velezensis B. BV10. Genes 2022, 13, 1984. [Google Scholar] [CrossRef]
- Paterson, J.; Jahanshah, G.; Li, Y.; Wang, Q.; Mehnaz, S.; Gross, H. The contribution of genome mining strategies to the understanding of active principles of PGPR strains. FEMS Microbiol. Ecol. 2017, 93, fiw249. [Google Scholar] [CrossRef]
- Montoya-Martínez, A.C.; Valenzuela-Ruíz, V.; Ortega-Urquieta, M.E.; Parra-Cota, F.I.; de los Santos-Villalobos, S. Genomic mining for the identification of promising mechanisms of bioactivity in biological control agents. In Biocontrol Agents for Improved Agriculture; Academic Press: Cambridge, MA, USA, 2024; pp. 143–163. [Google Scholar]
- Panth, M.; Hassler, S.C.; Baysal-Gurel, F. Methods for management of soilborne diseases in crop production. Agriculture 2020, 10, 16. [Google Scholar] [CrossRef]
- Ortega-Urquieta, M.E.; Valenzuela-Ruíz, V.; Mitra, D.; Hyder, S.; Elsheery, N.I.; Kumar Das Mohapatra, D.; Parra-Cota, F.I.; de los Santos-Villalobos, S. Draft Genome Sequence of Priestia sp. Strain TSO9, a Plant Growth-Promoting Bacterium Associated with Wheat (Triticum turgidum subsp. durum) in the Yaqui Valley, Mexico. Plants 2022, 11, 2231. [Google Scholar] [CrossRef] [PubMed]
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. CheckM: Assessing the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.H.; Ha, S.M.; Lim, J.; Kwon, S.; Chun, J. A large-scale evaluation of algorithms to calculate average nucleotide identity. Antonie Van Leeuwenhoek 2017, 110, 1281–1286. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.P.; Göker, M. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinform. 2013, 14, 60. [Google Scholar] [CrossRef] [PubMed]
- Blin, K.; Shaw, S.; Augustijn, H.E.; Reitz, Z.L.; Biermann, F.; Alanjary, M.; Fetter, A.; Terlouw, B.R.; Metcalf, W.W.; Helfrich, E.J.N.; et al. antiSMASH 7.0: New and improved predictions for detection, regulation, chemical structures and visualisation. Nucleic Acids Res. 2023, 51, W46–W50. [Google Scholar] [CrossRef] [PubMed]
- de los Santos-Villalobos, S.; Díaz-Rodríguez, A.M.; Ávila-Mascareño, M.F.; Martínez-Vidales, A.D.; Parra-Cota, F.I. COLMENA: A culture collection of native microorganisms for harnessing the agro-biotechnological potential in soils and contributing to food security. Diversity 2021, 13, 337. [Google Scholar] [CrossRef]
- Baard, V.; Bakare, O.O.; Daniel, A.I.; Nkomo, M.; Gokul, A.; Keyster, M.; Klein, A. Biocontrol potential of Bacillus subtilis and Bacillus tequilensis against four fusarium species. Pathogens 2023, 12, 254. [Google Scholar] [CrossRef]
- Rueden, C.T.; Schindelin, J.; Hiner, M.C.; DeZonia, B.E.; Walter, A.E.; Arena, E.T.; Eliceiri, K.W. ImageJ2: ImageJ for the next generation of scientific image data. BMC Bioinform. 2017, 18, 529. [Google Scholar] [CrossRef]
- R Core Team. R version 4.3.0 (2023–04–21 ucrt) “Already Tomorrow” Copyright (C) 2023 The R Foundation for Statistical Computing Platform: x86_64–w64–mingw32/x64 (64–bit). 2023. Available online: https://www.r-project.org/ (accessed on 25 February 2024).
- Lefort, V.; Desper, R.; Gascuel, O. FastME 2.0: A comprehensive, accurate, and fast distance-based phylogeny inference program. Mol. Biol. Evol. 2015, 32, 2798–2800. [Google Scholar] [CrossRef]
- Farris, J.S. Estimating phylogenetic trees from distance matrices. Am. Nat. 1972, 106, 645–667. [Google Scholar] [CrossRef]
- Yi, H.S.; Ahn, Y.R.; Ryu, C.M. Impact of a bacterial volatile 2, 3-butanediol on Bacillus subtilis rhizosphere robustness. Front. Microbiol. 2016, 7, 180263. [Google Scholar] [CrossRef] [PubMed]
- Ayilara, M.S.; Adeleke, B.S.; Babalola, O.O. Bioprospecting and challenges of plant microbiome research for sustainable agriculture, a review on soybean endophytic bacteria. Microb. Ecol. 2023, 85, 1113–1135. [Google Scholar] [CrossRef] [PubMed]
- Collinge, D.B.; Jensen, D.F.; Rabiey, M.; Sarrocco, S.; Shaw, M.W.; Shaw, R.H. Biological control of plant diseases–What has been achieved and what is the direction? Plant Pathol. 2022, 71, 1024–1047. [Google Scholar] [CrossRef]
- Rampersad, S.N. Pathogenomics and management of Fusarium diseases in plants. Pathogens 2020, 9, 340. [Google Scholar] [CrossRef] [PubMed]
- Todorović, I.; Moënne-Loccoz, Y.; Raičević, V.; Jovičić-Petrović, J.; Muller, D. Microbial diversity in soils suppressive to Fusarium diseases. Front. Plant Sci. 2023, 14, 1228749. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Selvaraj, J.; Xing, F.; Zhou, L.; Wang, Y.; Song, H.; Tan, X.; Sun, L.; Sangare, L.; Elodie-Folly, Y.M.; et al. Antagonistic action of Bacillus subtilis strain SG6 on Fusarium graminearum. PLoS ONE 2014, 9, e92486. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, V.V.; Santoyo, G.; Godínez, L.J.G.; Chávez, L.A.C.; Cota, F.I.P.; Villalobos, S.d.L.S. Complete genome sequencing of Bacillus cabrialesii TE3T: A plant growth-promoting and biological control agent isolated from wheat (Triticum turgidum subsp. durum) in the Yaqui Valley. Curr. Res. Microb. Sci. 2023, 4, 100193. [Google Scholar]
- Chen, L.; Heng, J.; Qin, S.; Bian, K. A comprehensive understanding of the biocontrol potential of Bacillus velezensis LM2303 against Fusarium head blight. PLoS ONE 2018, 13, e0198560. [Google Scholar] [CrossRef]
- Villa-Rodriguez, E.; Moreno-Ulloa, A.; Castro-Longoria, E.; Parra-Cota, F.I.; de Los Santos-Villalobos, S. Integrated omics approaches for deciphering antifungal metabolites produced by a novel Bacillus species, B. cabrialesii TE3T, against the spot blotch disease of wheat (Triticum turgidum L. subsp. durum). Microbiol. Res. 2021, 251, 126826. [Google Scholar] [CrossRef]
- Gong, A.D.; Li, H.P.; Yuan, Q.S.; Song, X.S.; Yao, W.; He, W.J.; Zhang, J.B.; Liao, Y.C. Antagonistic mechanism of iturin A and plipastatin A from Bacillus amyloliquefaciens S76-3 from wheat spikes against Fusarium graminearum. PLoS ONE 2015, 10, e0116871. [Google Scholar] [CrossRef] [PubMed]
- Boulahouat, S.; Cherif-Silini, H.; Silini, A.; Bouket, A.C.; Luptakova, L.; Alenezi, F.N.; Belbahri, L. Biocontrol Efficiency of Rhizospheric Bacillus against the Plant Pathogen Fusarium oxysporum: A Promising Approach for Sustainable Agriculture. Microbiol. Res. 2023, 14, 892–908. [Google Scholar] [CrossRef]
- Dimopoulou, A.; Theologidis, I.; Benaki, D.; Koukounia, M.; Zervakou, A.; Tzima, A.; Diallinas, G.; Hatzinikolaou, D.G.; Skandalis, N. Direct antibiotic activity of bacillibactin broadens the biocontrol range of Bacillus amyloliquefaciens MBI600. Msphere 2021, 6, e00376-21. [Google Scholar] [CrossRef]
- Ghosh, S.K.; Bera, T.; Chakrabarty, A.M. Microbial siderophore—A boon to agricultural sciences. Biol. Control. 2020, 144, 104214. [Google Scholar] [CrossRef]
- Palmieri, D.; Ianiri, G.; Del Grosso, C.; Barone, G.; De Curtis, F.; Castoria, R.; Lima, G. Advances and perspectives in the use of biocontrol agents against fungal plant diseases. Horticulturae 2022, 8, 577. [Google Scholar] [CrossRef]
- Singh, S.; Kumar, V.; Dhanjal, D.S.; Singh, J. Biological control agents: Diversity, ecological significances, and biotechnological applications. In Natural Bioactive Products in Sustainable Agriculture; Springer: Singapore, 2020; pp. 31–44. [Google Scholar]
- Bonaterra, A.; Badosa, E.; Daranas, N.; Francés, J.; Roselló, G.; Montesinos, E. Bacteria as biological control agents of plant diseases. Microorganisms 2022, 10, 1759. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).