
Study on Dormant and Germination Characteristics of Chinese Olive (Canarium album) Seeds

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Observation of Seed Morphology
2.3. Seed Water Absorption Rate
2.4. Seed Inhibitor Activity Test
2.4.1. Preparation of Crude Extracts from Different Parts of Chinese Olive Seeds
2.4.2. The Inhibitory Effect of Crude Extracts from Different Parts of Chinese Olive Seeds
2.5. Seed Germination Test
2.6. Data Analysis
3. Results
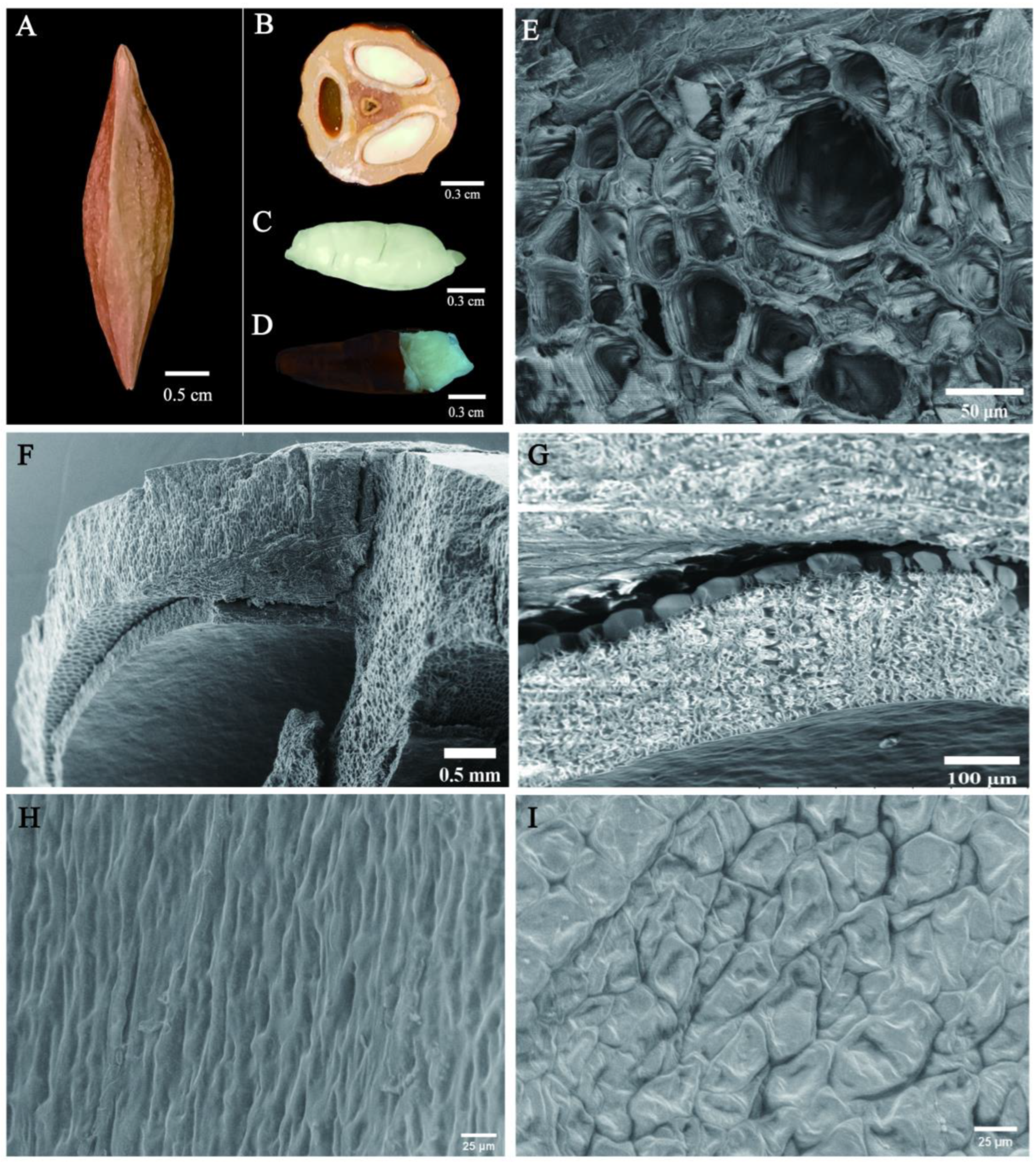
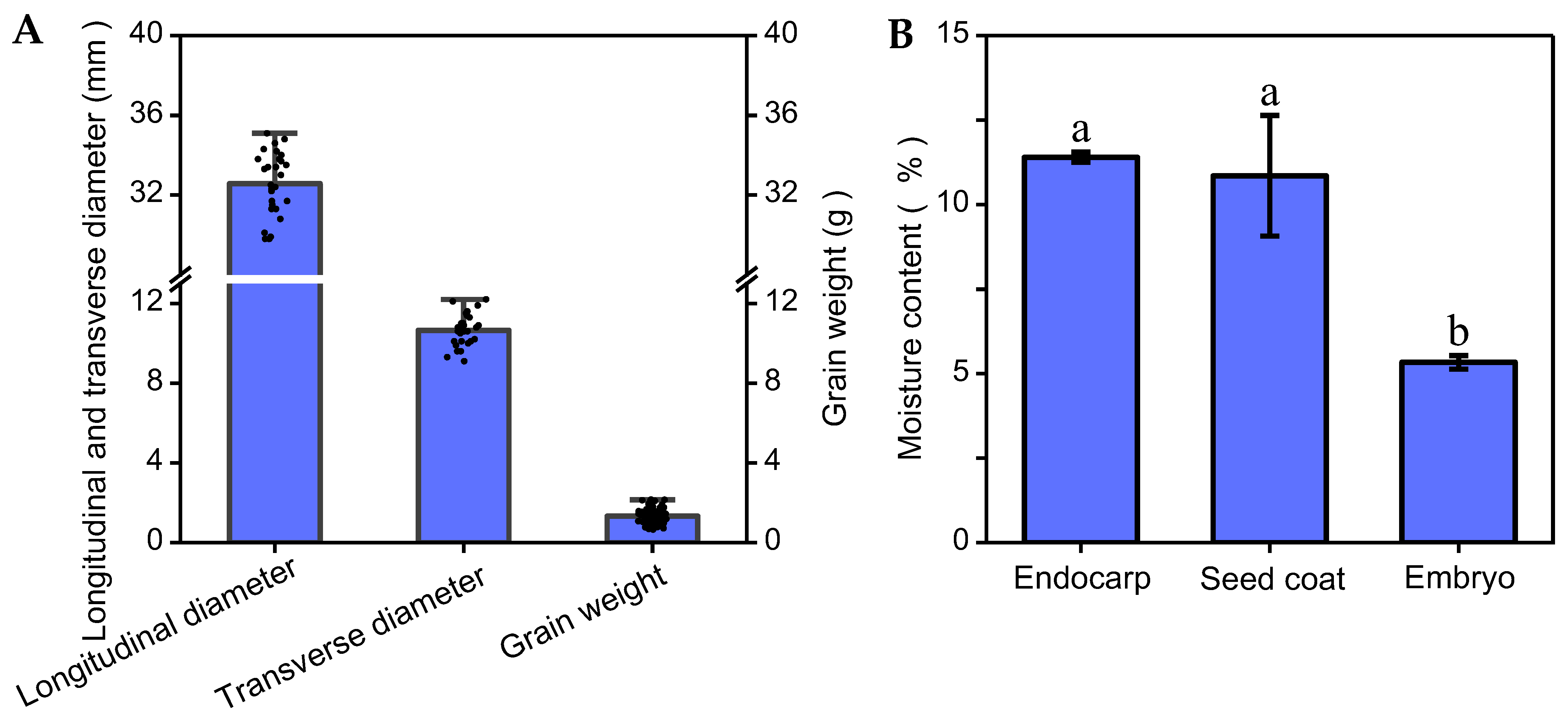
3.1. Morphological Characteristics of Chinese Olive Seeds
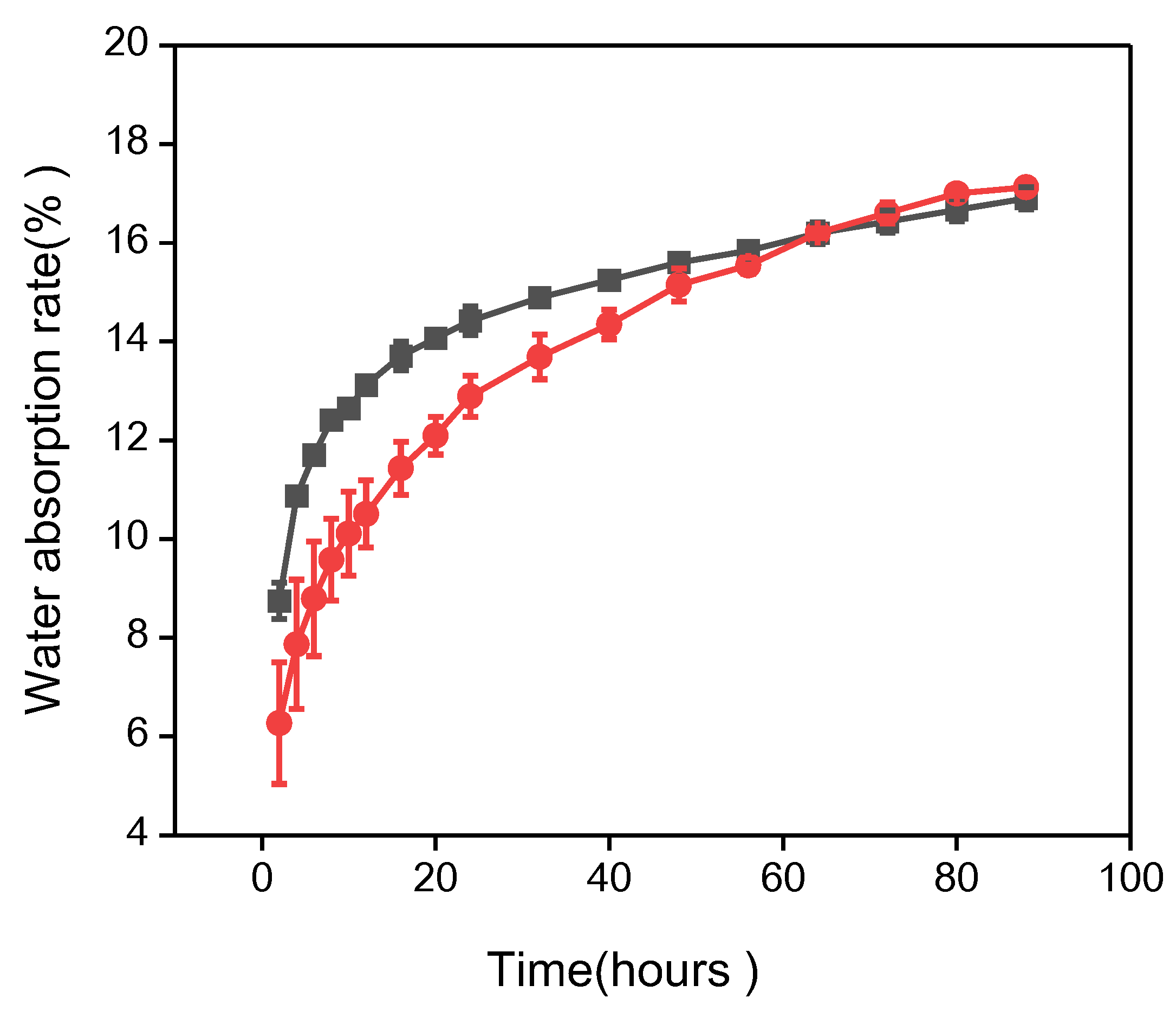
3.2. Analysis of Seed Water Absorption
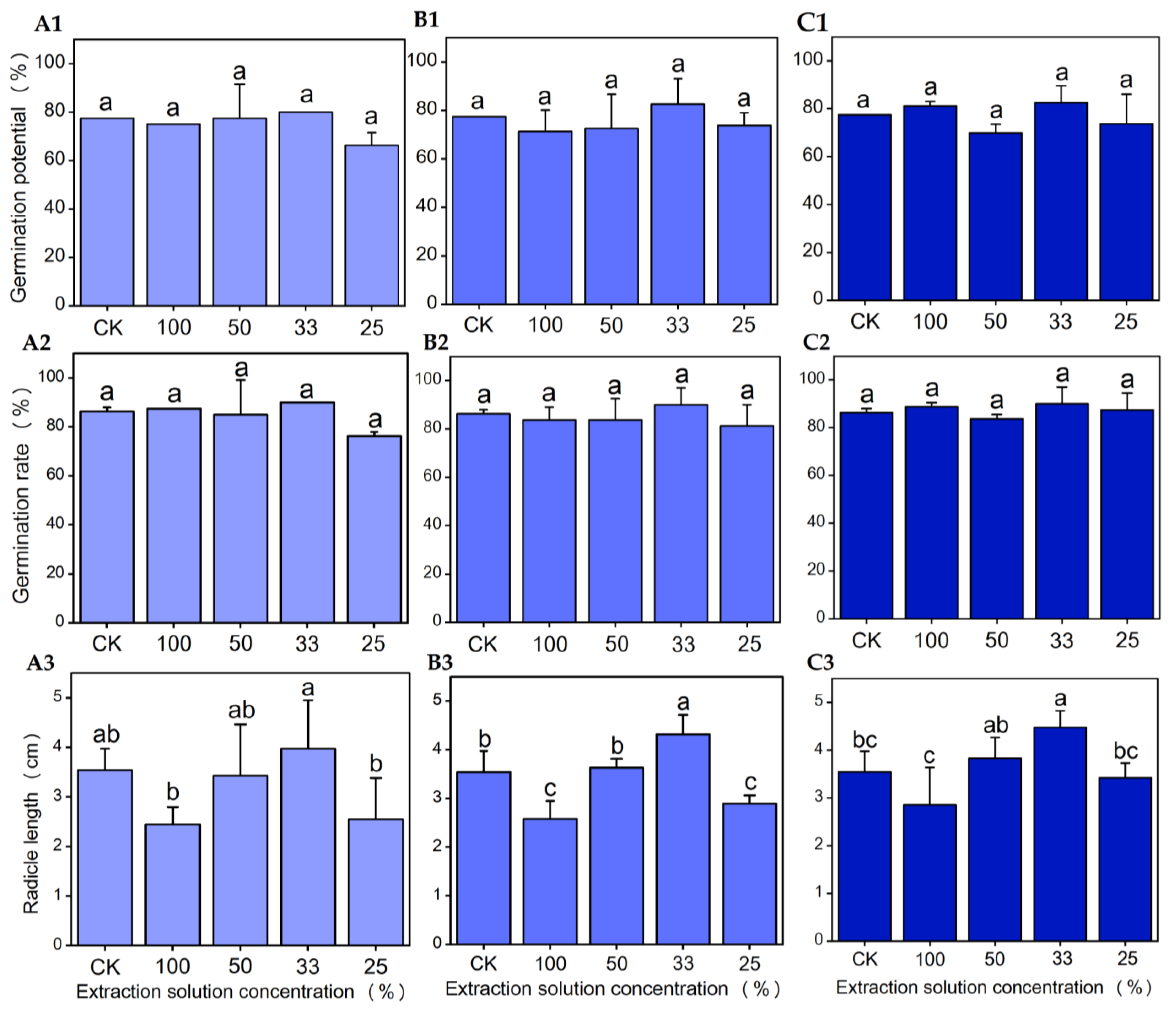
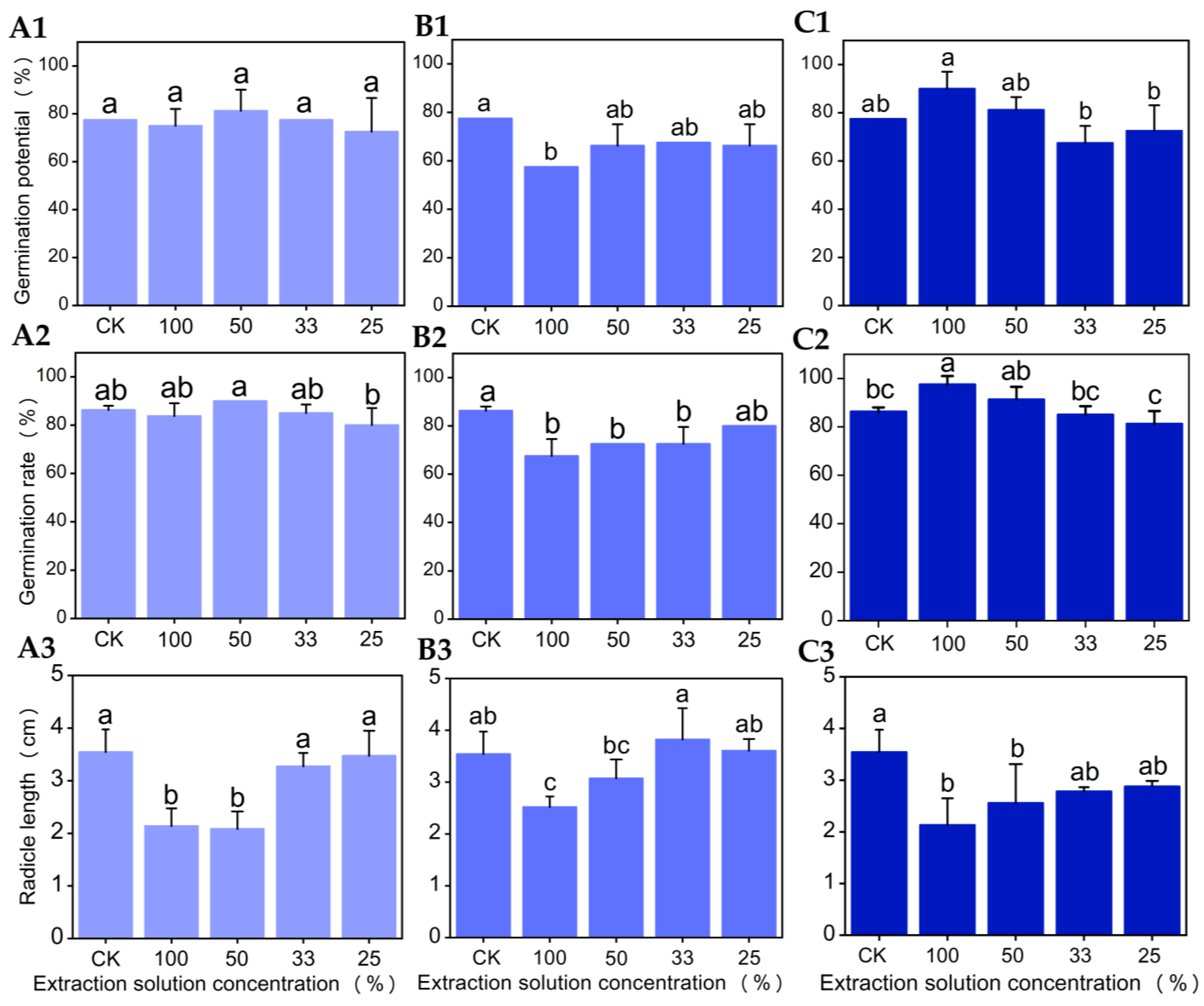
3.3. Effects of Seed Extracts from Different Parts of Chinese Olive on Seed Germination and Radicle Growth of Chinese Cabbage
3.3.1. Effects of Water Extract from Different Parts of Chinese Olive Seeds on Seed Germination and Radicle Growth of Chinese Cabbage
3.3.2. Effects of Methanol Extracts from Different Parts of Chinese Olive Seeds on Seed Germination and Radicle Growth of Chinese Cabbage
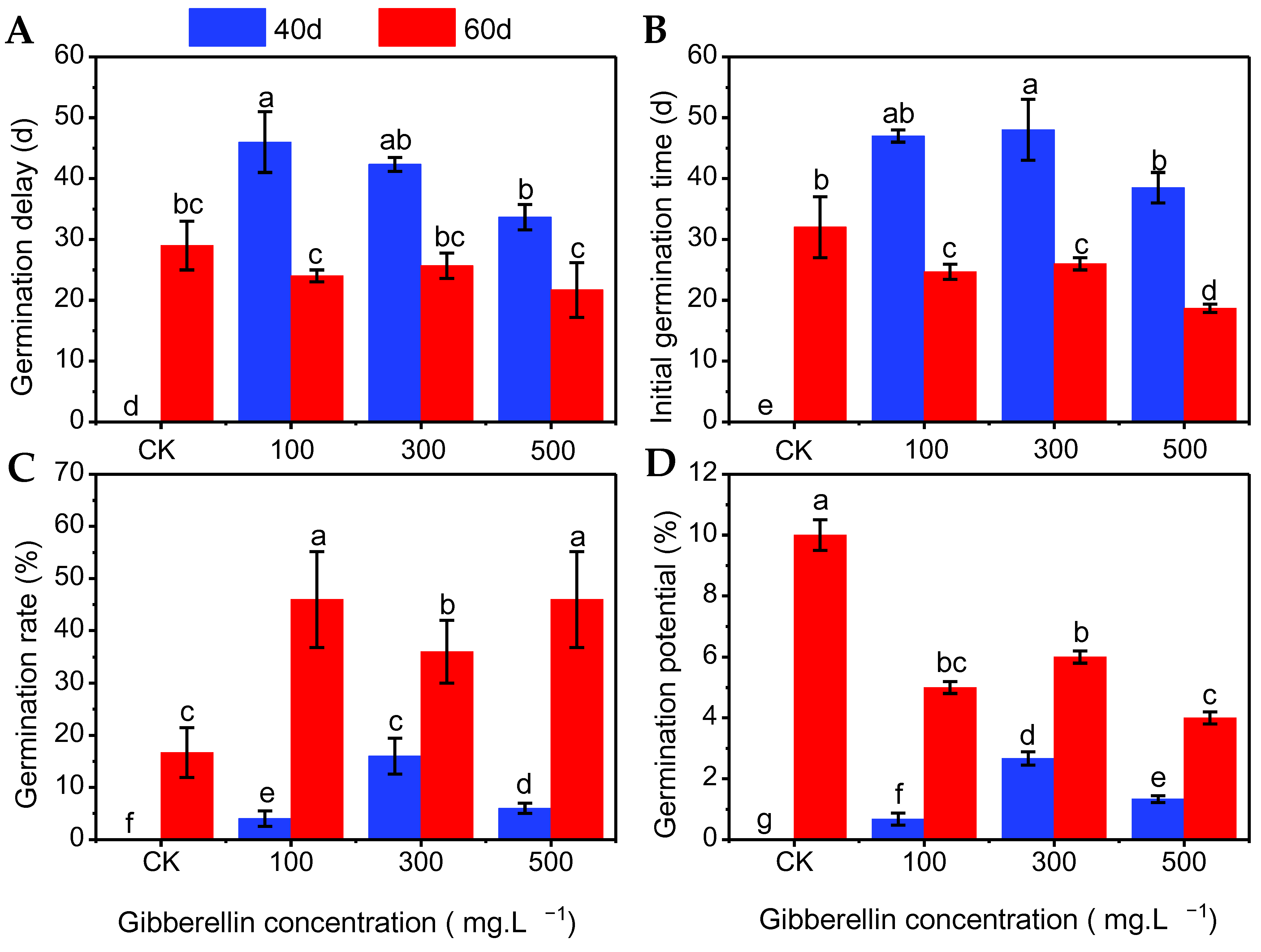
3.4. Effects of Gibberellin Combined with Low-Temperature Stratification on Seed Germination of Chinese Olive
3.5. Main Effect Analysis of Germination Index of Chinese Olive Seeds Treated with Low-Temperature Stratification and Gibberellin
4. Discussion
4.1. Seed Dormancy Characteristics
4.2. Seed Dormancy Release
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- He, Z.; Xia, W. Nutritional composition of the kernels from Canarium album L. Food Chem. 2007, 102, 808–811. [Google Scholar] [CrossRef]
- Editorial Committee of Flora of China, Chinese Academy of Sciences. Flora of China; Science Press: Beijing, China, 1997; Volume 43, pp. 17–27. (In Chinese) [Google Scholar]
- Chang, Q.; Su, M.; Chen, Q. The advance on the research of chemical constituents and pharmacological activities of Chinese olive. Chin. J. Trop. Crop. 2013, 34, 1610–1616. (In Chinese) [Google Scholar]
- Baskin, J.M.; Baskin, C.C. A classification system for seed dormancy. Seed Sci. Res. 2004, 14, 1–16. [Google Scholar] [CrossRef]
- Nonogaki, H. Seed germination and dormancy: The classic story, new puzzles, and evolution. J. Integr. Plant Biol. 2019, 61, 541–563. [Google Scholar] [CrossRef]
- Shu, K.; Liu, X.D.; Xie, Q.; He, Z.H. Two faces of one seed: Hormonal regulation of dormancy and germination. Mol. Plant 2016, 9, 34–45. [Google Scholar] [CrossRef]
- Zhang, W. Study on Dormancy Releasing Mechanism of Tulipa Thianschanica Regel Seeds; Shenyang Agricultural University: Shenyang, China, 2020. (In Chinese) [Google Scholar]
- Debeaujon, I.; Koornneef, M. Gibberellin requirement for arabidopsis seed germination is determined both by testa characteristics and embryonic abscisic acid. Plant Physiol. 2000, 122, 415–424. [Google Scholar] [CrossRef]
- Guan, S.X.; Li, Y.S.; Zeng, W.Q.; Lin, J.W.; Zhan, H.; Han, X.Y.; Zhang, X.L.; Lu, X.J. A gibberellin, abscisic acid, and delay of germination 1 interaction network regulates critical developmental transitions in model plant arabidopsis thaliana—A review. Appl. Ecol. Environ. Res. 2021, 19, 4699–4720. [Google Scholar] [CrossRef]
- Nelson, S.K.K.; Kanno, Y.; Seo, M.; Steber, C.M.M. Seed Dormancy loss from dry after-ripening is associated with increasing gibberellin hormone levels in Arabidopsis thaliana. Front. Plant Sci. 2023, 14, 1145414. [Google Scholar] [CrossRef]
- Matilla, A.J.; Matilla-Vazquez, M.A. Involvement of ethylene in seed physiology. Plant Sci. 2008, 175, 87–97. [Google Scholar] [CrossRef]
- Linkies, A.; Graeber, K.; Knight, C.; Leubner-Metzger, G. The evolution of seeds. New Phytol. 2010, 186, 817–831. [Google Scholar] [CrossRef]
- Li, S.; Shi, T.; Kong, F.; Ma, Q.; Mao, Y.; Yin, T. Methods for breaking the dormancy of eastern redbud (Cercis canadensis) seeds. Seed Sci. Technol. 2013, 41, 27–35. [Google Scholar] [CrossRef]
- Bian, F.; Su, J.; Liu, W.; Li, S. Dormancy release and germination of Taxus yunnanensis seeds during wet sand storage. Sci. Rep. 2018, 8, 3205. [Google Scholar] [CrossRef] [PubMed]
- Vasil’eva, O.Y. Reproduction systems of representatives of the Genus rosa L. under conditions of continental climate. Contemp. Probl. Ecol. 2009, 2, 361–368. [Google Scholar] [CrossRef]
- Nadeem, M.; Riaz, A.; Younis, A.; Akond, M.; Farooq, A.; Tariq, U. Improved Technique for treating seed dormancy to enhance germination in Rosa × Hybrida. Turk. J. Bot. 2013, 37, 521–529. [Google Scholar] [CrossRef]
- Dai, J. Several treatment experiments on Chinese olive seedling cultivation. Southeast Hortic. 1996, 5–7. (In Chinese) [Google Scholar]
- Tuan, P.; Yamasaki, Y.; Kanno, Y.; Mitsunori, S.; Ayele, B.T. Transcriptomics of cytokinin and auxin metabolism and signaling genes during seed maturation in dormant and non-dormant wheat genotypes. Sci. Rep. 2019, 9, 3983. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H. Practical methods for rapid seed germination from seed coat-imposed dormancy of Prunus yedoensis. Sci. Hortic. 2019, 243, 451–456. [Google Scholar] [CrossRef]
- Ye, Q.; Wang, W.; Xie, Q.; Chen, Q. Dormancy physiology and stratification effects of Chinese olive [Canarium album (Lour.) Raeusch.] Seeds. Chin. J. Trop. Crop. 2021, 42, 1646–1653. (In Chinese) [Google Scholar]
- Tang, Q.; Chen, Y.; Jiang, L.; Chen, J.; Li, C.; Zeng, W.; Liu, Q.; Li, P. Characterization and mechanism of seed dormancy in Symplocos paniculata. Front. Plant Sci. 2024, 14, 1322238. [Google Scholar] [CrossRef]
- Li, T.; Min, X. Dormancy characteristics and germination requirements of Phoebe bournei seed. Sci. Hortic. 2020, 260, 108903. [Google Scholar] [CrossRef]
- Zhang, W.; Qu, L.W.; Zhao, J.; Xue, L.; Dai, H.P.; Xing, G.M.; Lei, J.J. Practical methods for breaking seed dormancy in a wild ornamental tulip species Tulipa thianschanica Regel. Agronomy 2020, 10, 1765. [Google Scholar] [CrossRef]
- Tang, S.; Cui, N.; Feng, Y.; Zhang, J.; Liu, Y. Effect of gibberellin concentration on seed germination of Acer mono Maxim. Chin. J. Appl. Environ. Biol. 2021, 27, 555–559. (In Chinese) [Google Scholar] [CrossRef]
- Song, D.; Jaganathan, G.K.; Han, Y.; Liu, B. Seed dormancy in Camellia sinensis L. (Theaceae): Effects of cold-stratification and exogenous gibberellic acid application on germination. Botany 2016, 95, 147–152. [Google Scholar] [CrossRef]
- Sajeev, N.; Koornneef, M.; Bentsink, L. A commitment for life: Decades of unraveling the molecular mechanisms behind seed dormancy and germination. Plant Cell 2024. [Google Scholar] [CrossRef] [PubMed]
- Francoz, E.; Lepiniec, L.; North, H.M. Seed coats as an alternative molecular factory: Thinking outside the box. Plant Reprod. 2018, 31, 327–342. [Google Scholar] [CrossRef] [PubMed]
- Huo, S.; Zhang, X.; Xiang, Z.; Yan, Q.; Zhang, J.; Yu, Z.; Peng, F.; Xiong, W. Study on the rates of sucking up moisture with maize seeds during their germination. J. Maize Sci. 2004, 12, 54–56, 59. (In Chinese) [Google Scholar]
- Song, S.; Tang, C.; Lei, H.; Jiang, X.; Wang, W.; Cheng, H. Research progress of seed dormancy and germination regulation. Acta Agron. Sin. 2024, 50, 1–15. (In Chinese) [Google Scholar] [CrossRef]
- Blake, P.S.; Taylor, J.M.; Finch-Savage, W.E. Identification of abscisic acid, indole-3-acetic acid, jasmonic acid, indole-3-acetonitrile, methyl jasmonate and gibberellins in developing, dormant and stratified seeds of ash (Fraxinus excelsior). Plant Growth Regul. 2002, 37, 119–125. [Google Scholar] [CrossRef]
- Kucera, B.; Cohn, M.A.; Leubner-Metzger, G. Plant hormone interactions during seed dormancy release and germination. Seed Sci. Res. 2005, 15, 281–307. [Google Scholar] [CrossRef]
- Graeber, K.; Nakabayashi, K.; Miatton, E.; Leubner-Metzger, G.; Soppe, W.J.J. Molecular mechanisms of seed dormancy. Plant Cell Environ. 2012, 35, 1769–1786. [Google Scholar] [CrossRef]
- White, C.N.; Rivin, C.J. Gibberellins and Seed Development in Maize. Ii. Gibberellin synthesis inhibition enhances abscisic acid signaling in cultured embryos. Plant Physiol. 2000, 122, 1089–1097. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Hu, P.; Huang, M.; Tang, Y.; Li, Y.; Li, L.; Hou, X. The Nf-Yc-Rgl2 module integrates GA and ABA signalling to regulate seed germination in Arabidopsis. Nat. Commun. 2016, 7, 12768. [Google Scholar] [CrossRef] [PubMed]
- Carruggio, F.; Onofri, A.; Impelluso, C.; Giusso del Galdo, G.; Scopece, G.; Cristaudo, A. Seed dormancy breaking and germination in Bituminaria basaltica and B. bituminosa (Fabaceae). Plants 2020, 9, 1110. [Google Scholar] [CrossRef] [PubMed]
- Shi, F.; Cao, Y.; Gao, Y.; Qiu, Y.; Lu, Y.; Han, B.; Shen, Y. The impact of magnetic field and gibberellin treatment on the release of dormancy and internal nutrient transformation in Tilia miqueliana Maxim. Seeds. For. 2024, 15, 311. [Google Scholar] [CrossRef]
- Bhat, I.A.; Magray, J.A.; Guleria, K.; Fayaz, M.; Ul Qadir, R.; Ganie, A.H.; Kaloo, Z.A. Conservation implications of seed germination studies of Aquilegia fragrans Benth.: A native and endangered medicinal plant species of kashmir himalaya. S. Afr. J. Bot. 2024, 166, 151–160. [Google Scholar] [CrossRef]
- Yuan, Z.; Wang, C.; Li, S.; Li, X.; Tai, F. Effects of Different plant hormones or peg seed soaking on maize resistance to drought stress. Can. J. Plant Sci. 2014, 94, 1491–1499. [Google Scholar] [CrossRef]
- Chahtane, H.; Kim, W.; Lopez-Molina, L. Primary seed dormancy: A temporally multilayered riddle waiting to be unlocked. J. Exp. Bot. 2017, 68, 857–869. [Google Scholar] [CrossRef] [PubMed]
- Yan, A.; Chen, Z. The control of seed dormancy and germination by temperature, light and nitrate. Bot. Rev. 2020, 86, 39–75. [Google Scholar] [CrossRef]
- Yamauchi, Y.; Ogawa, M.; Kuwahara, A.; Hanada, A.; Kamiya, Y.; Yamaguchi, S. Activation of gibberellin biosynthesis and response pathways by low temperature during imbibition of arabidopsis thaliana seeds. Plant Cell 2004, 16, 367–378. [Google Scholar] [CrossRef]
- Hu, X.W.; Huang, X.H.; Wang, Y.R. Hormonal and temperature regulation of seed dormancy and germination in Leymus chinensis. Plant Growth Regul. 2012, 67, 199–207. [Google Scholar] [CrossRef]
- Kushiro, T.; Okamoto, M.; Nakabayashi, K.; Yamagishi, K.; Kitamura, S.; Asami, T.; Hirai, N.; Koshiba, T.; Kamiya, Y.; Nambara, E. The Arabidopsis cytochrome P450cyp707a encodes ABA 8′-hydroxylases: Key enzymes in ABA catabolism. EMBO J. 2004, 23, 1647–1656. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Huang, Y.; Cao, Z.; Li, Y.; Liu, D.; Han, X.; Yin, K.; Liu, Q. New insights into the molecular mechanism of low-temperature stratification on dormancy release and germination of Saposhnikovia divaricata seeds. Braz. J. Bot. 2022, 45, 1183–1198. [Google Scholar] [CrossRef]
- Amooaghaie, R.; Ahmadi, F. Triangular interplay between ros, aba and ga in dormancy alleviation of bunium persicum seeds by cold stratification. Russ. J. Plant Physiol. 2017, 64, 588–599. [Google Scholar] [CrossRef]
- Gianinetti, A. A travel through landscapes of seed dormancy. Plants 2023, 12, 3963. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | Main Effect of Low-Temperature Stratification (A) | Main Effect of Gibberellin (B) | Interaction (A×B) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| F | p | η2 | F | p | η2 | F | p | η2 | |
| Germination delay | 19.470 | 0.00 | 0.549 | 59.101 | 0.00 | 0.917 | 90.106 | 0.00 | 0.944 |
| Initial germination time | 524.634 | 0.00 | 0.970 | 755.469 | 0.00 | 0.993 | 1447.751 | 0.00 | 0.996 |
| Germination rate | 74.318 | 0.00 | 0.941 | 15.600 | 0.00 | 0.770 | 7.600 | 0.003 | 0.620 |
| Germination potential | 12.944 | 0.00 | 0.735 | 609.961 | 0.00 | 0.992 | 6661.557 | 0.00 | 0.993 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, Q.; Ye, Q.; Liu, T.; Chen, Z.; Chen, Q. Study on Dormant and Germination Characteristics of Chinese Olive (Canarium album) Seeds. Horticulturae 2024, 10, 362. https://doi.org/10.3390/horticulturae10040362
Xie Q, Ye Q, Liu T, Chen Z, Chen Q. Study on Dormant and Germination Characteristics of Chinese Olive (Canarium album) Seeds. Horticulturae. 2024; 10(4):362. https://doi.org/10.3390/horticulturae10040362
Chicago/Turabian StyleXie, Qian, Qinghua Ye, Ting Liu, Zhanhong Chen, and Qingxi Chen. 2024. "Study on Dormant and Germination Characteristics of Chinese Olive (Canarium album) Seeds" Horticulturae 10, no. 4: 362. https://doi.org/10.3390/horticulturae10040362