
A Genome-Wide Analysis of the WUSCHEL-Related Homeobox Transcription Factor Family Reveals Its Differential Expression Patterns, Response to Drought Stress, and Localization in Sweet Cherry (Prunus avium L.)


Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Culture Conditions
2.2. WOX Gene Identification in Sweet Cherry and Bioinformatics Analysis
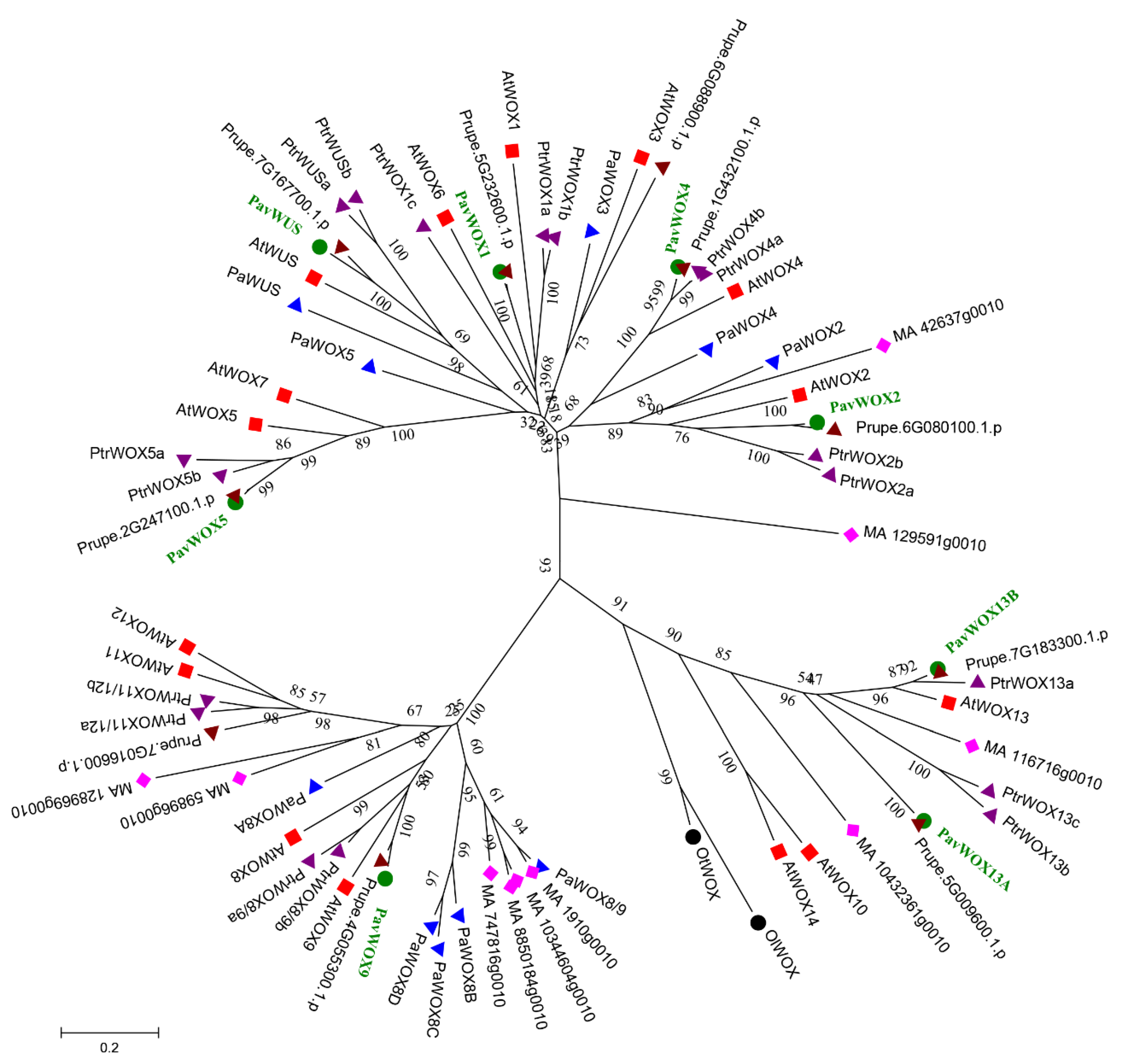
2.3. Phylogenetic Analyses
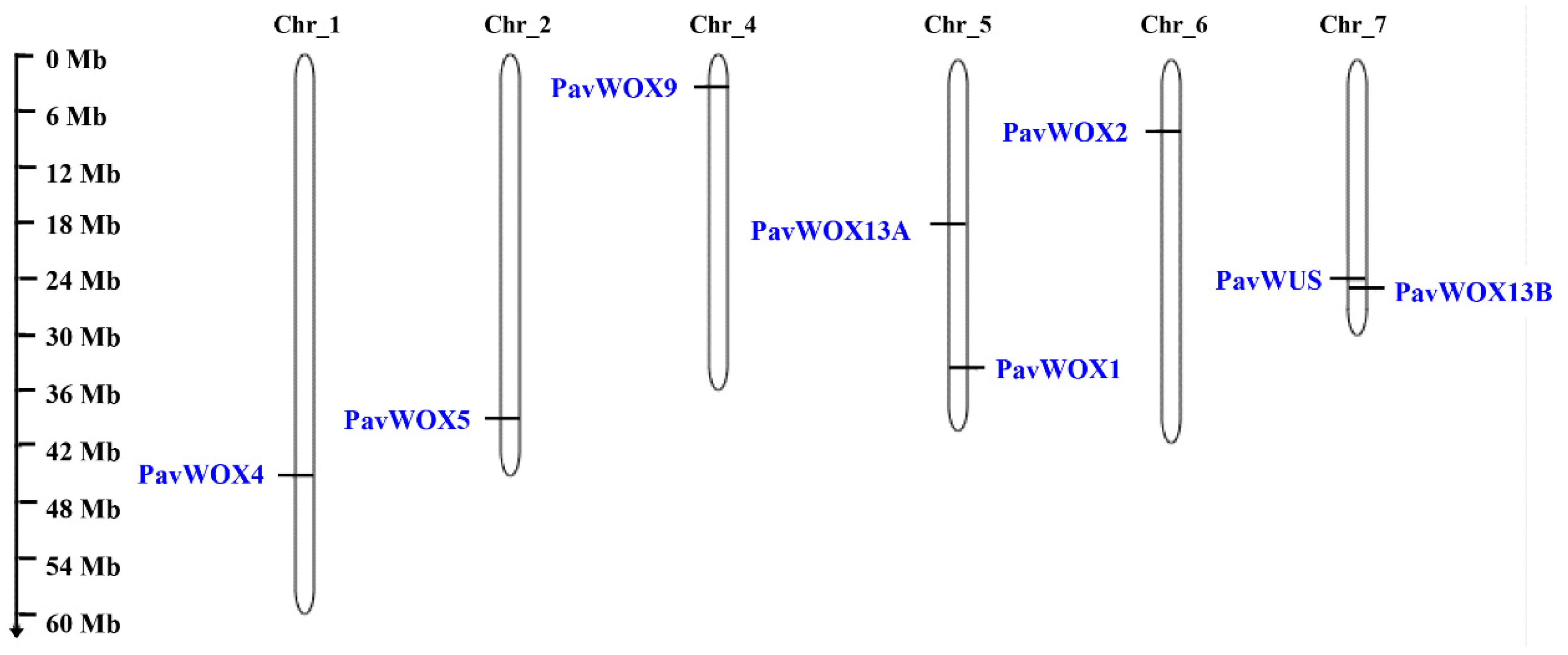
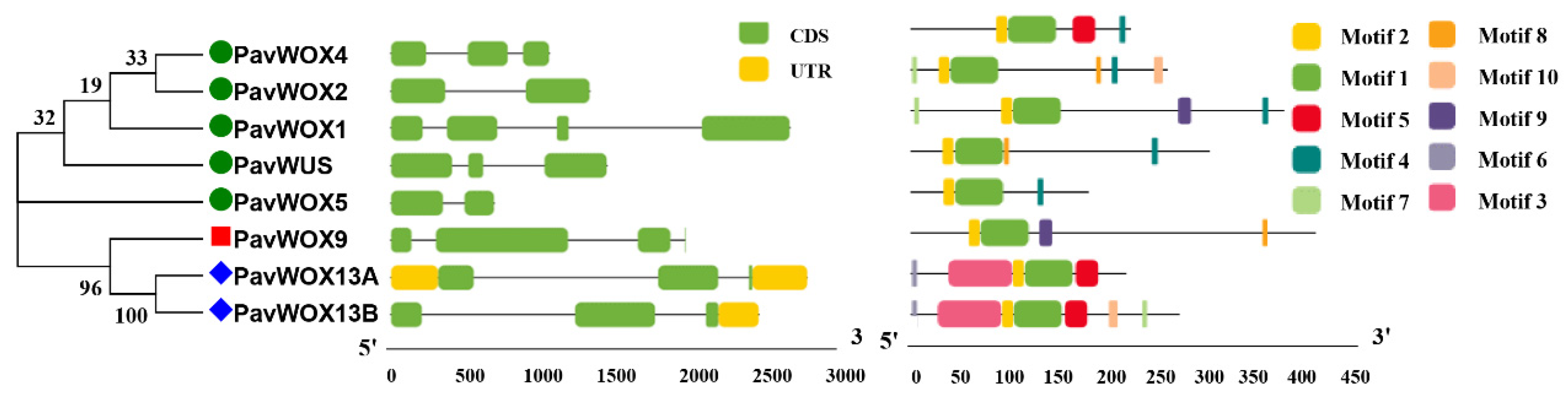
2.4. Analyses of Gene Structures, Conserved Motifs, and Chromosomal Locations
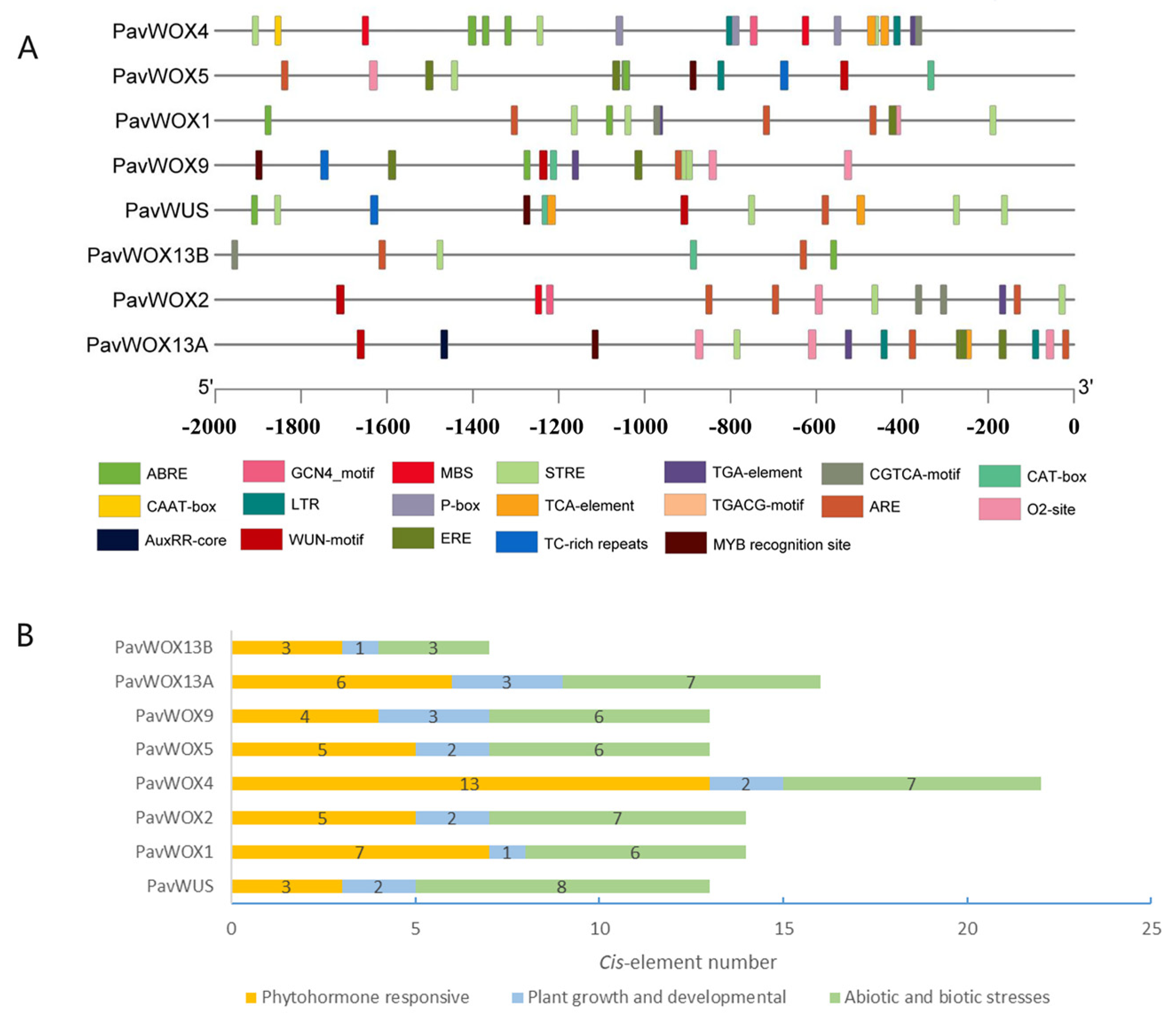
2.5. Promoter Analysis
2.6. Gene Expression Analysis
2.7. RNA Isolation, cDNA Synthesis, and Gene Cloning
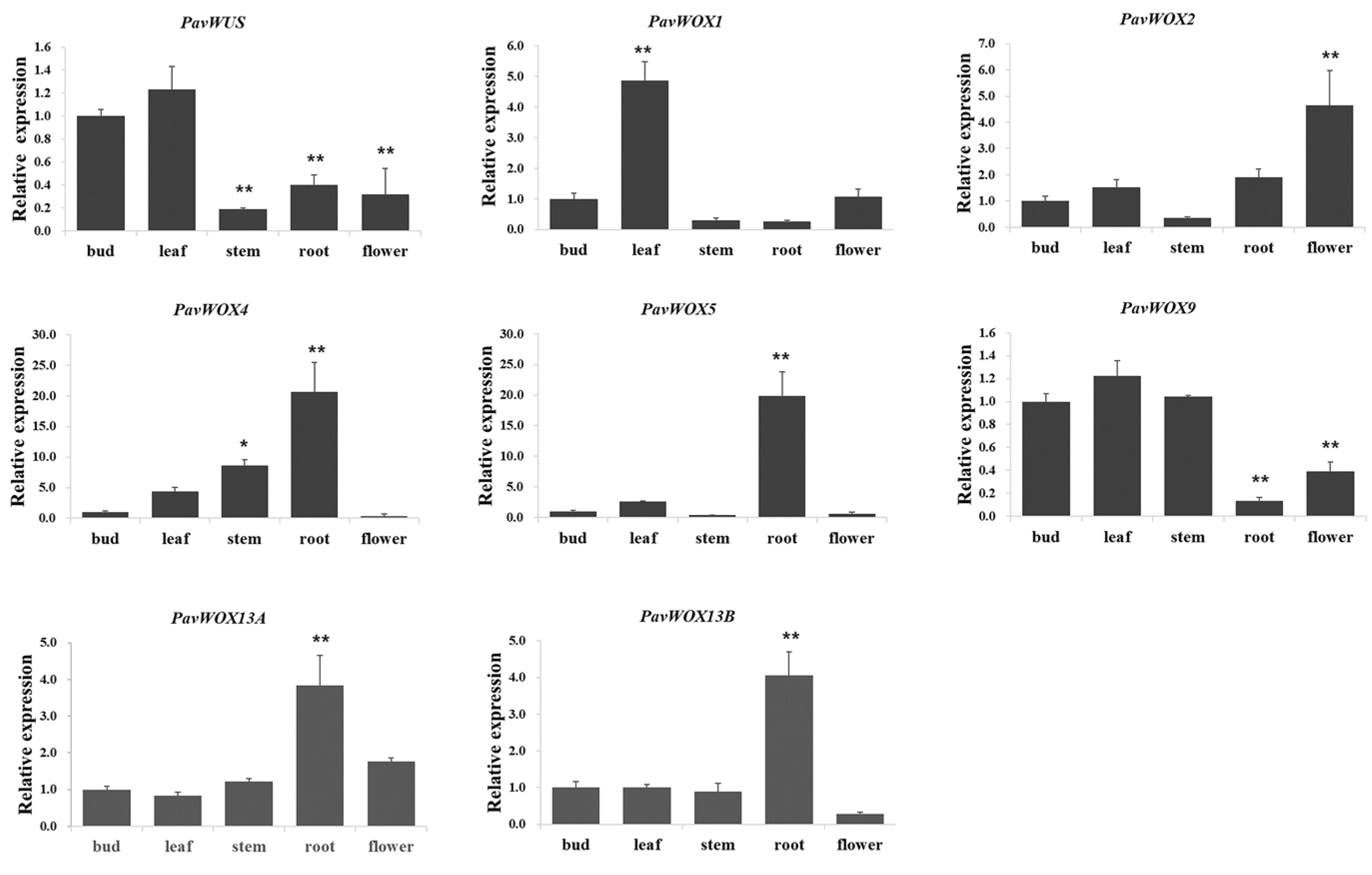
2.8. Expression Analysis of PavWOX by qRT-PCR
2.9. Plasmid Construction and Subcellular Localization of PavWOX13A
2.10. Data Analysis Method
3. Results
3.1. WOX Gene Family Identification within Sweet Cherry
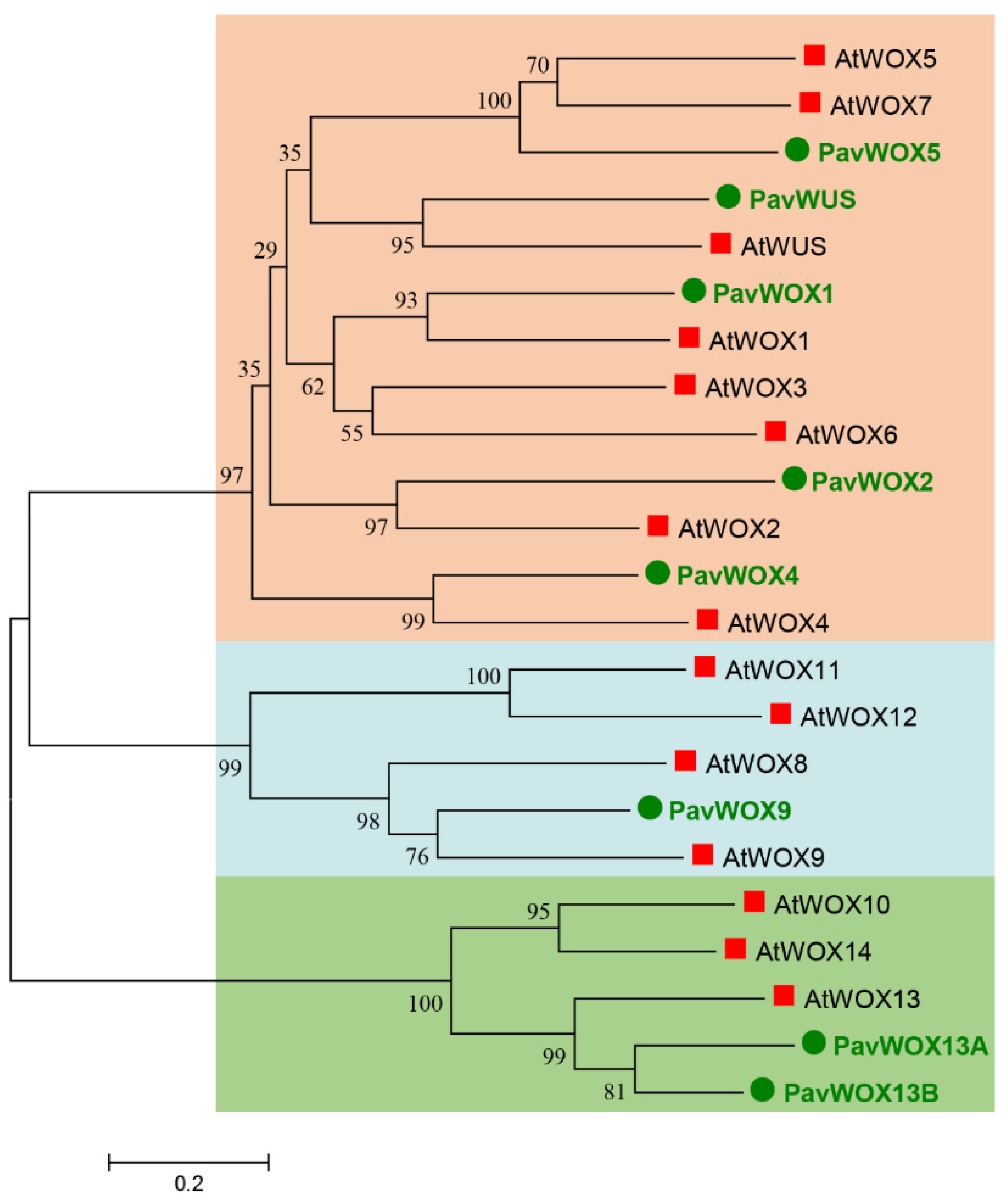
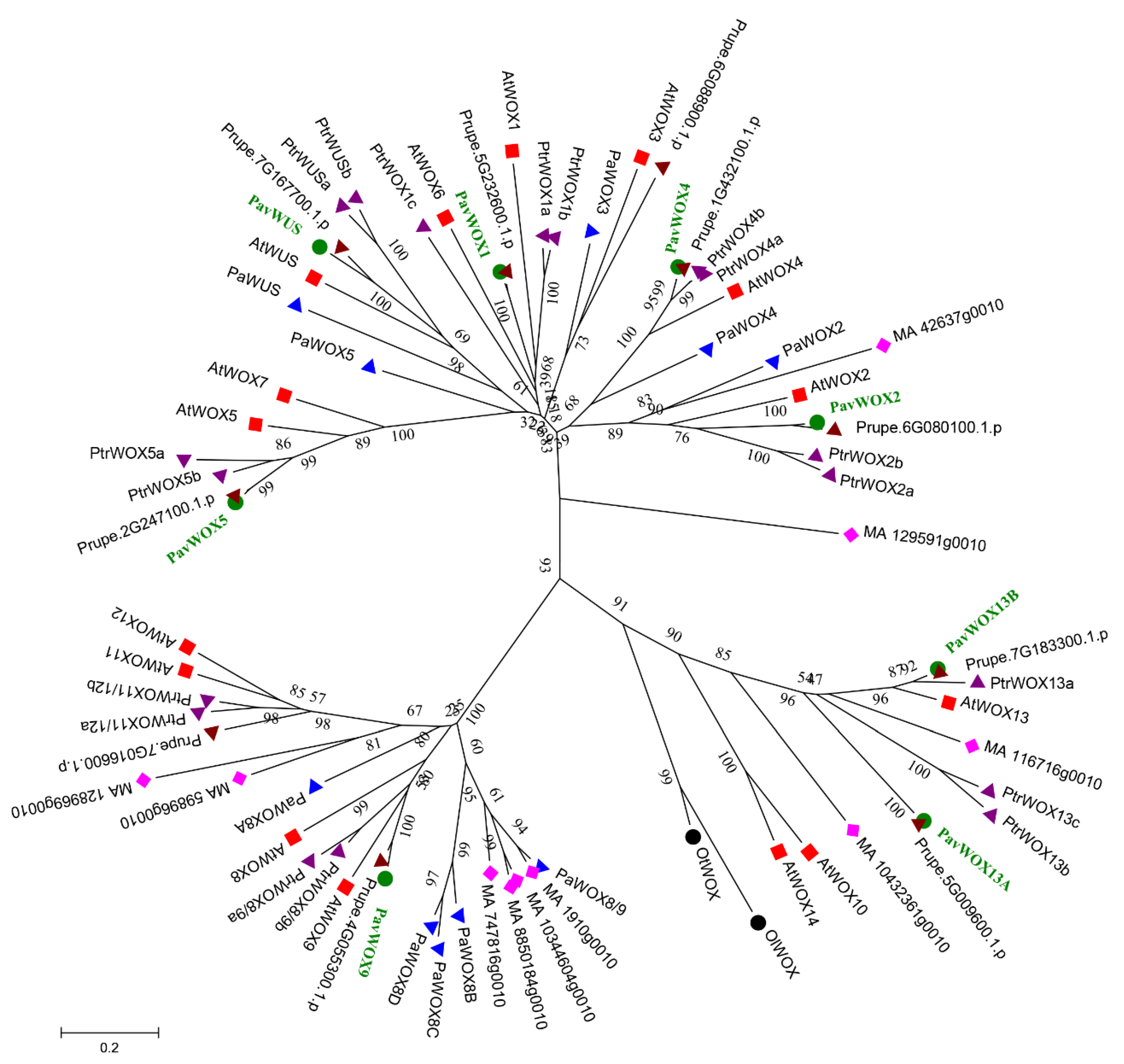
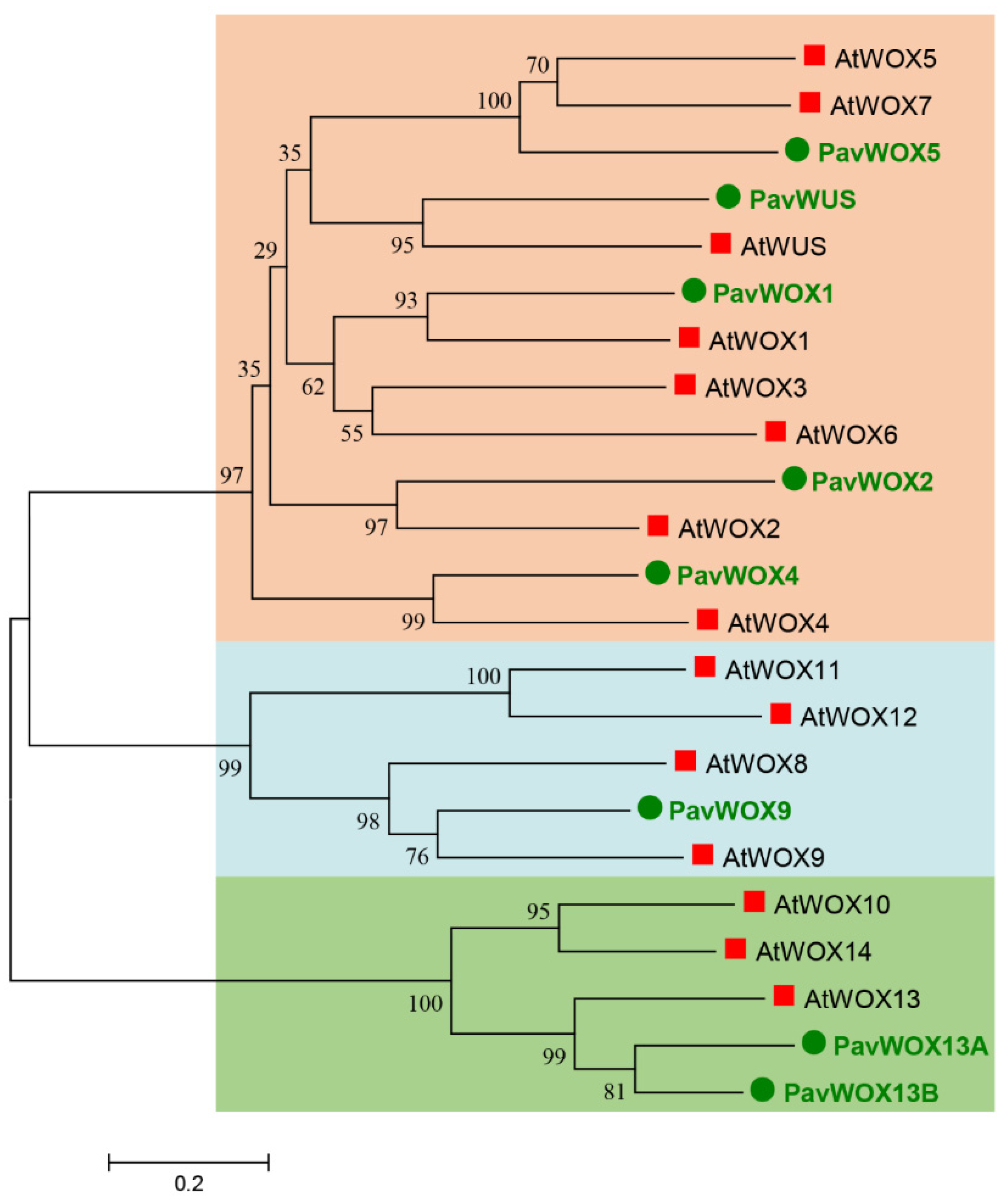
3.2. Phylogenetic Analyses of the WOX Gene Family
3.3. WOX Gene Structures and Conserved Motifs within Sweet Cherry
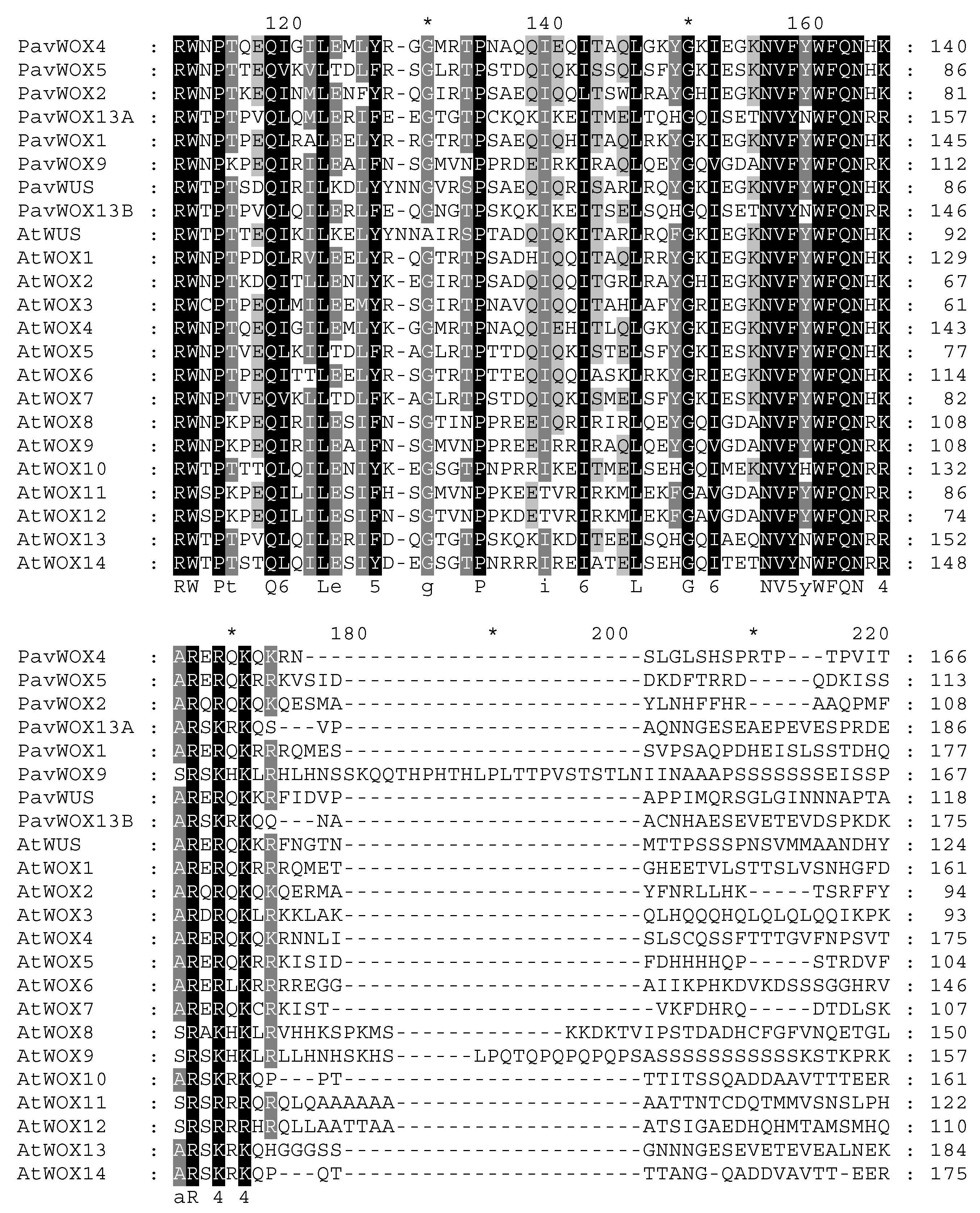
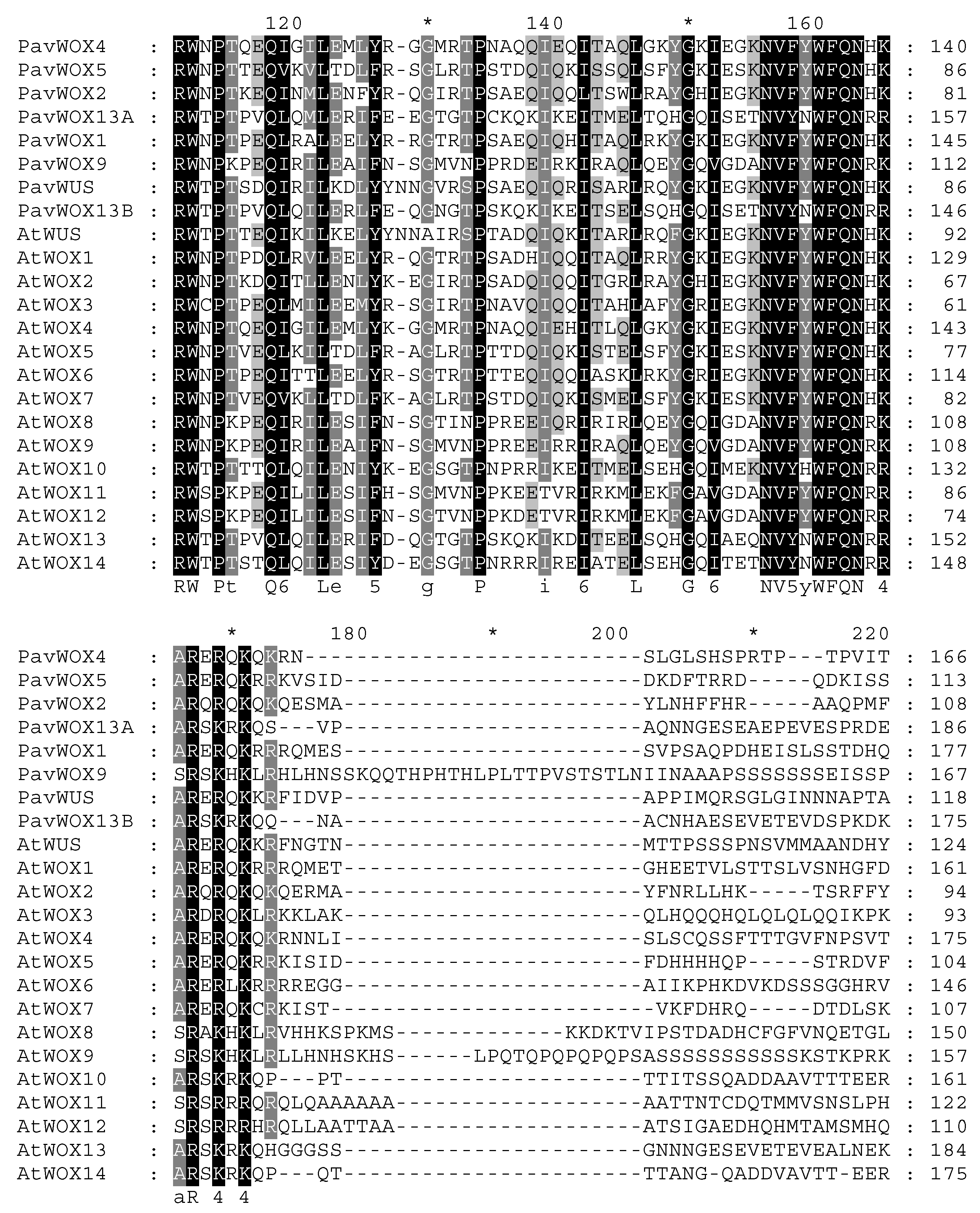
3.4. Multiple Sequence Alignment Analysis on WOX Proteins within Sweet Cherry
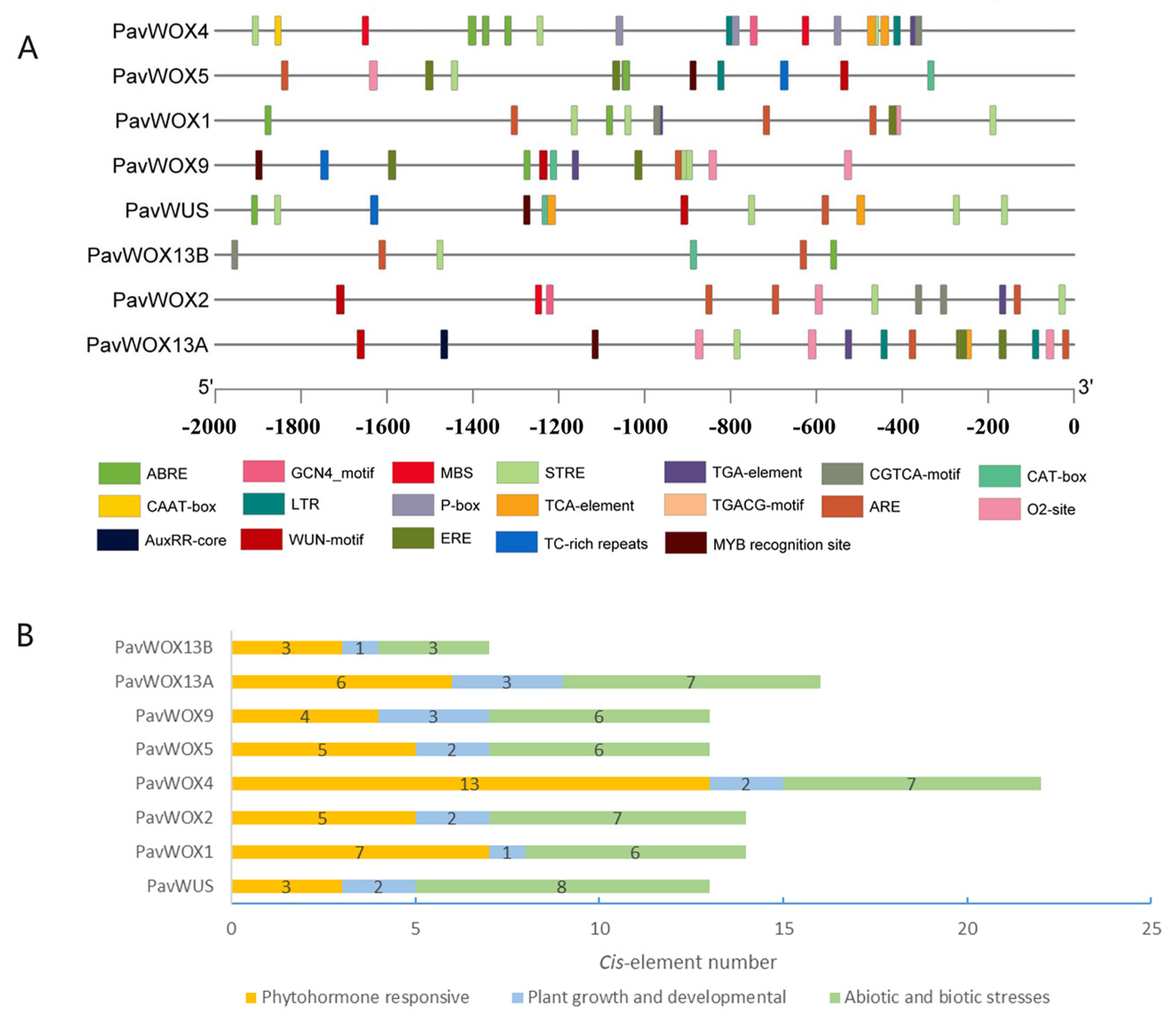
3.5. Promoter Component Analysis of WOX Genes in Sweet Cherry
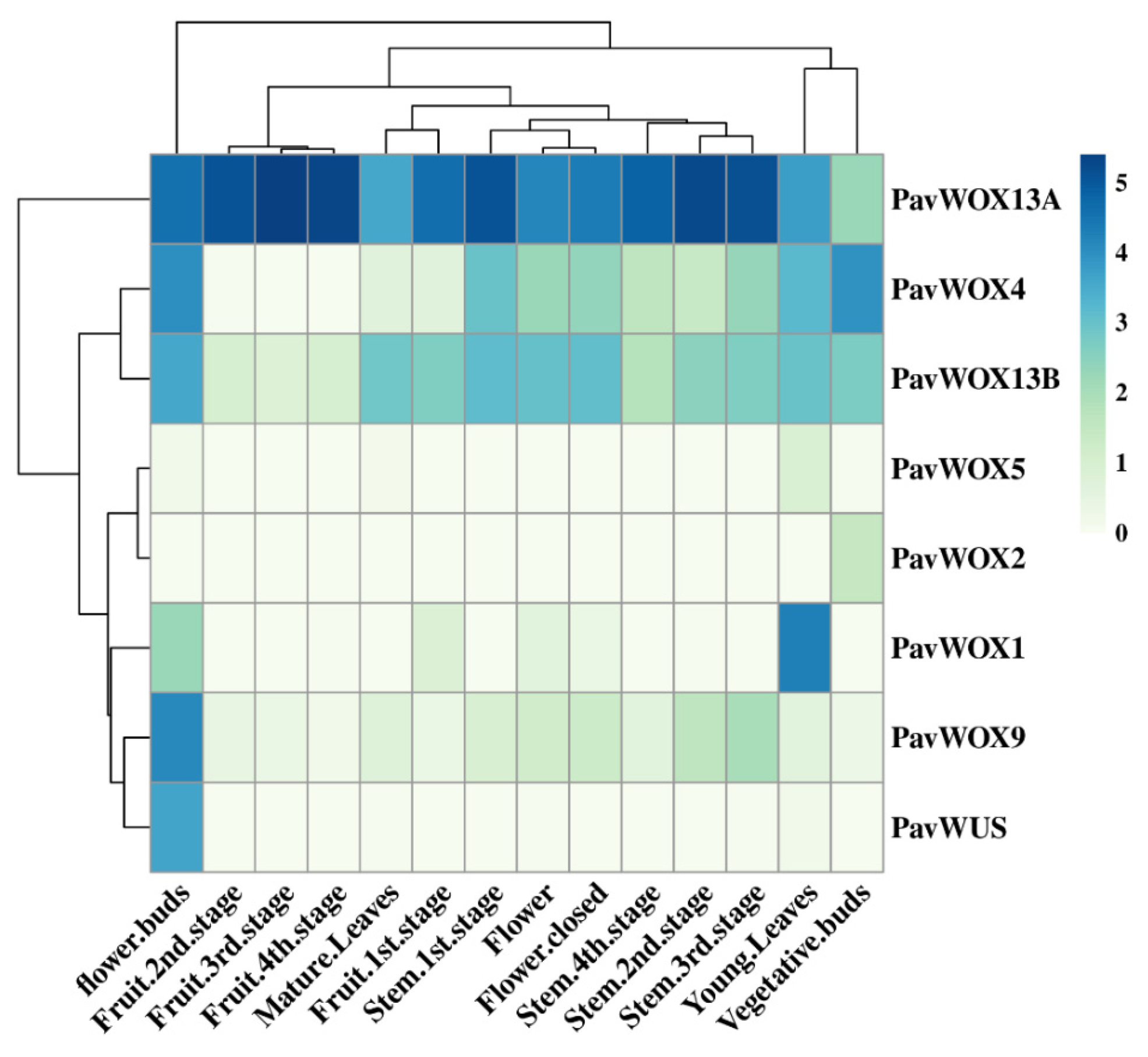
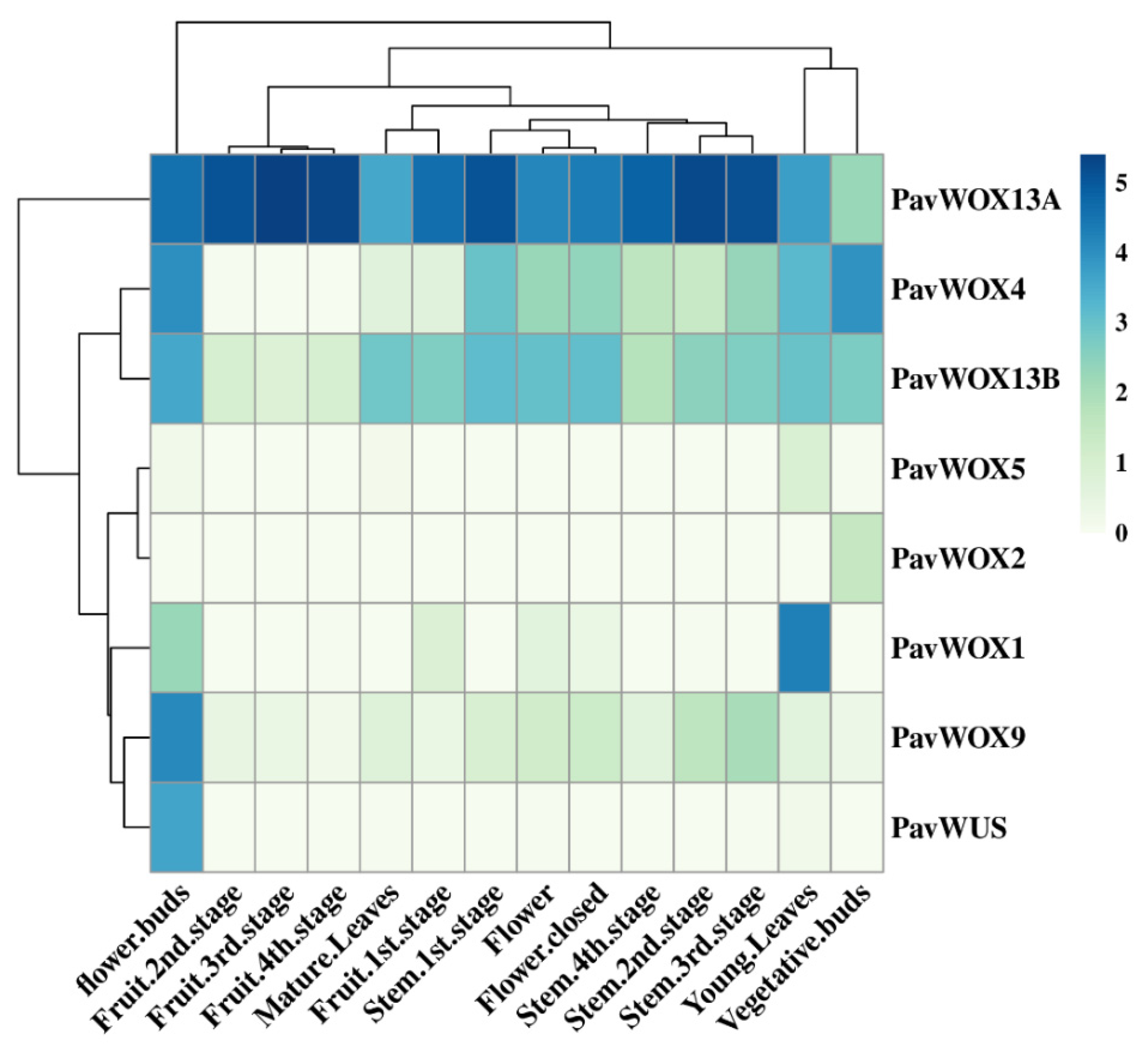
3.6. Differential WOX Gene Expression within Sweet Cherry
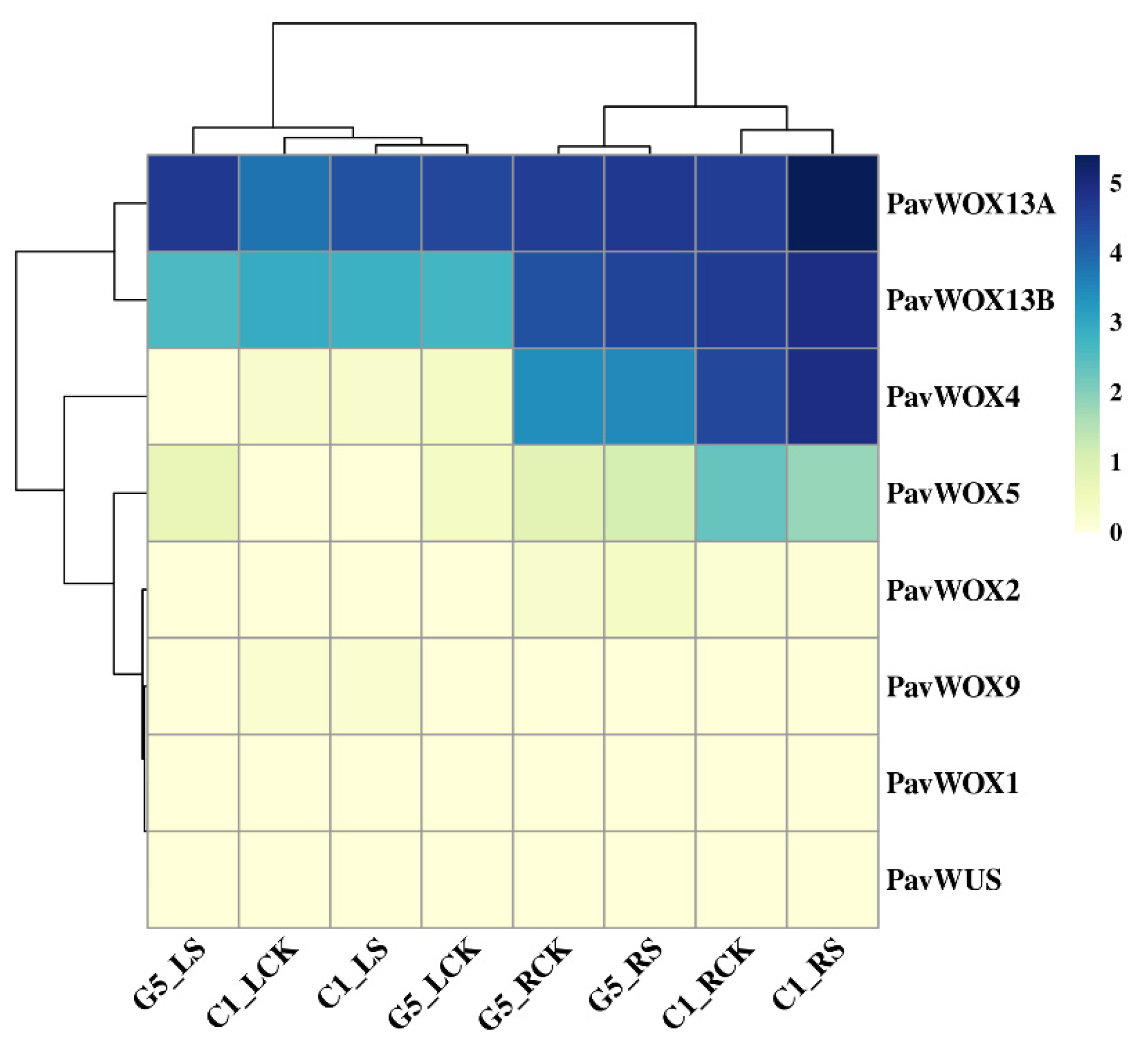
3.7. PavWOX Expression under Drought Stress
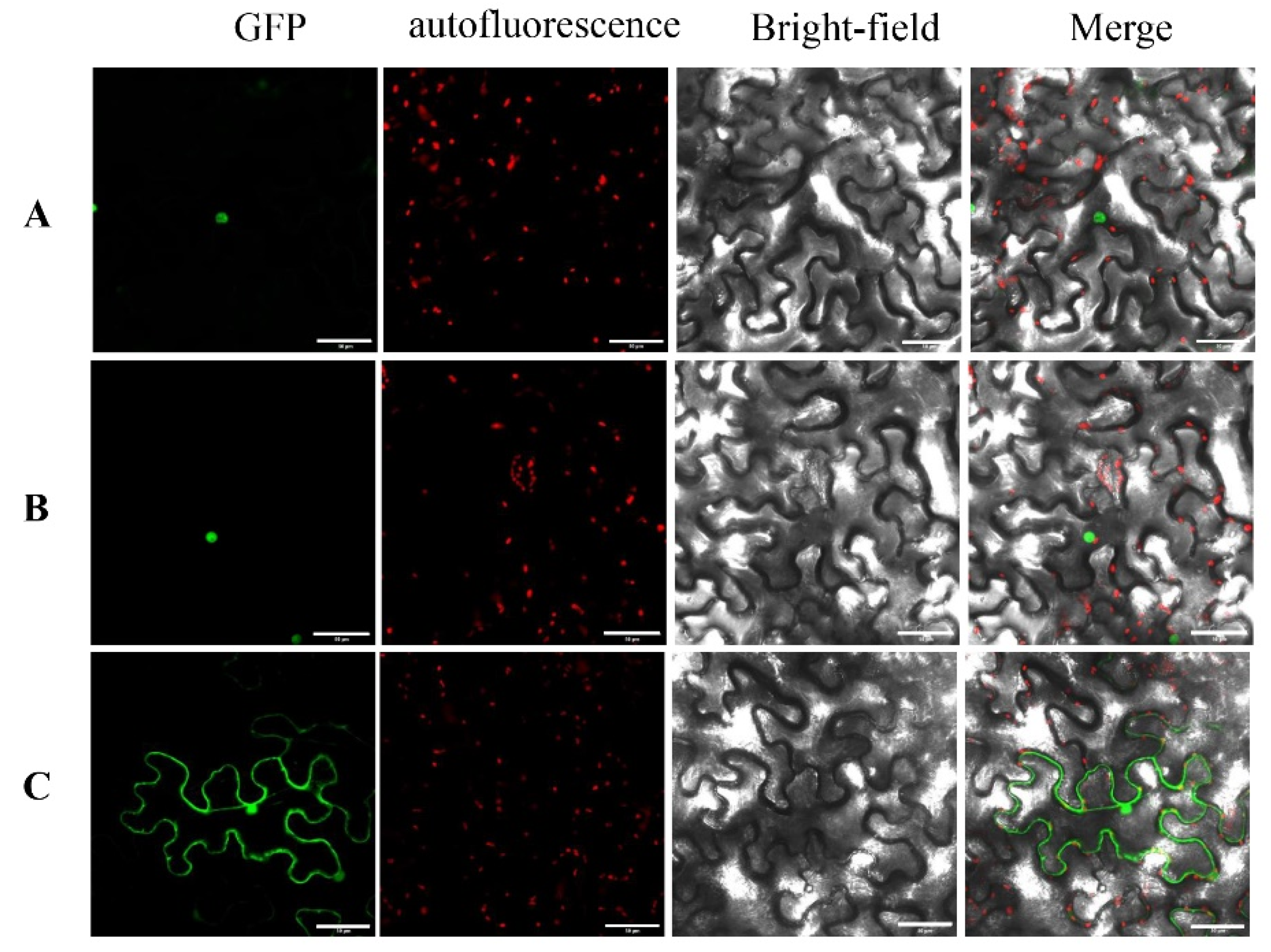
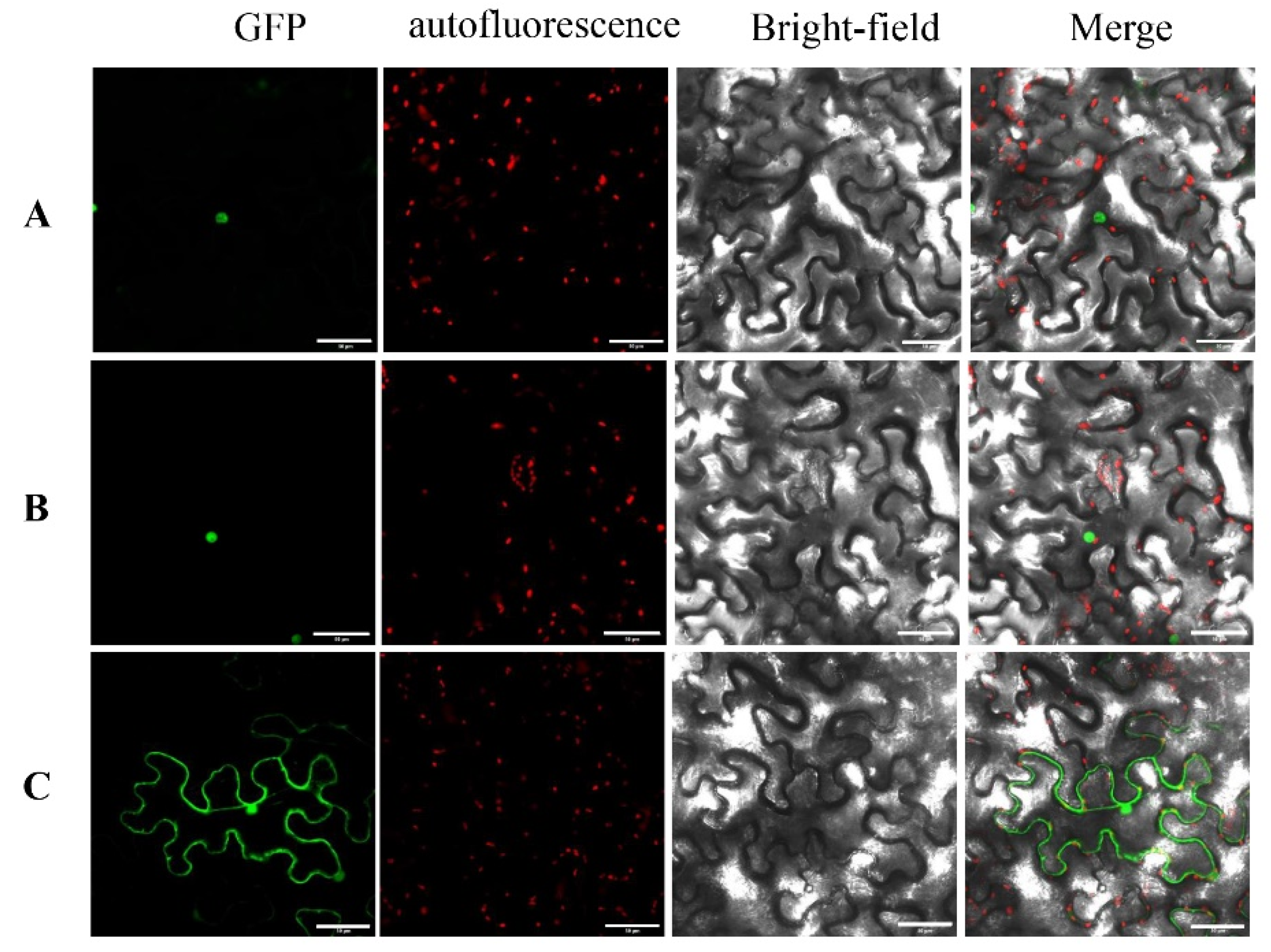
3.8. Nuclear Localization of PavWOX4 and PavWOX13A
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Achim, H.; Rita, G.H.; Bernd, G.; Ananda, S.; Holger, B.; Marita, H.; Thomas, L. Expression dynamics of WOX genes mark cell fate decisions during early embryonic patterning in Arabidopsis thaliana. Development 2004, 131, 657–668. [Google Scholar]
- Graaff, E.V.D.; Laux, T.; Rensing, S.A. The WUS homeobox-containing (WOX) protein family. Genome Biol. 2009, 10, 248. [Google Scholar]
- Chen, S.K.; Kurdyukov, S.; Kereszt, A.; Wang, X.D.; Gresshoff, P.M.; Rose, R.J.J.P. The association of homeobox gene expression with stem cell formation and morphogenesis in cultured Medicago truncatula. Planta 2009, 230, 827–840. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.; Huang, Y.; Zhu, N.; Zhao, Y. The rice WUSCHEL-related homeobox genes are involved in reproductive organ development, hormone signaling and abiotic stress response. Gene 2014, 549, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Wang, R.; Liu, Z.; Wu, X.; Wang, J. Genome-wide identification and analysis of the WUSCHEL-related homeobox (WOX) gene family in allotetraploid Brassica napus reveals changes in WOX genes during polyploidization. BMC Genom. 2019, 20, 317. [Google Scholar] [CrossRef] [PubMed]
- Rathour, M.; Sharma, A.; Kaur, A.; Upadhyay, S.K.J.H. Genome-wide characterization and expression and co-expression analysis suggested diverse functions of WOX genes in bread wheat. Heliyon 2020, 6, e05762. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zong, J.; Liu, J.; Yin, J.; Zhang, D. Genome-wide analysis of WOX gene family in rice, sorghum, maize, Arabidopsis and poplar. J. Integr. Plant Biol. 2010, 52, 1016–1026. [Google Scholar] [CrossRef]
- Shafique Khan, F.; Gan, R.-F.; Zhang, Z.M.; Hu, J.Z. J, C.G. Genome-Wide Identification and Expression Profiling of the WOX Gene Family in Citrus sinensis and Functional Analysis of a CsWUS Member. Int. J. Mol. Sci. 2021, 22, 4919. [Google Scholar] [CrossRef]
- Liu, B.; Wang, L.; Zhang, J.; Li, J.; Zheng, H.; Chen, J.; Lu, M. WUSCHEL-related Homeobox genes in Populus tomentosa: Diversified expression patterns and a functional similarity in adventitious root formation. BMC Genom. 2014, 15, 296. [Google Scholar] [CrossRef]
- Zhu, T.; Moschou, P.N.; Alvarez, J.M.; Sohlberg, J.J.; von Arnold, S. WUSCHEL-RELATED HOMEOBOX 8/9 is important for proper embryo patterning in the gymnosperm Norway spruce. J. Exp. Bot. 2014, 65, 6543–6552. [Google Scholar] [CrossRef]
- Alvarez, J.M.; Bueno, N.; Cañas, R.A.; Avila, C.; Cánovas, F.M.; Ordás, R.J. Analysis of the WUSCHEL-RELATED HOMEOBOX gene family in Pinus pinaster: New insights into the gene family evolution. Plant Physiol. Biochem. 2017, 123, 304. [Google Scholar] [CrossRef] [PubMed]
- Lian, G.; Ding, Z.; Wang, Q.; Zhang, D.; Xu, J. Origins and Evolution of WUSCHEL-Related Homeobox Protein Family in Plant Kingdom. Sci. World J. 2014, 2014, 534140. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Liu, D.; Xia, Y.; Li, Z.; Jing, D.; Du, J.; Niu, N.; Ma, S.; Wang, J.; Song, Y.; et al. Identification of the WUSCHEL-Related Homeobox (WOX) Gene Family, and Interaction and Functional Analysis of TaWOX9 and TaWUS in Wheat. Int. J. Mol. Sci. 2020, 21, 1581. [Google Scholar] [CrossRef] [PubMed]
- Si, X.; Wang, W.; Wang, K.; Liu, Y.; Bai, J.; Meng, Y.; Zhang, X.; Liu, H. A Sheathed Spike Gene, TaWUS-like Inhibits Stem Elongation in Common Wheat by Regulating Hormone Levels. Int. J. Mol. Sci. 2021, 22, 11210. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Jia, H.; Sun, P.; Zhang, J.; Xia, Y.; Hu, J.; Wang, L.; Lu, M. The WUSCHELa (PtoWUSa) is Involved in Developmental Plasticity of Adventitious Root in Poplar. Genes 2020, 11, 176. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Runions, A.; Mentink, R.A.; Kierzkowski, D.; Karady, M.; Hashemi, B.; Huijser, P.; Strauss, S.; Gan, X.; Ljung, K.; et al. A WOX/Auxin Biosynthesis Module Controls Growth to Shape Leaf Form. Curr. Biol. 2020, 30, 4857–4868.e6. [Google Scholar] [CrossRef] [PubMed]
- Conklin, P.A.; Johnston, R.; Conlon, B.R.; Shimizu, R.; Scanlon, M.J. Plant homeodomain proteins provide a mechanism for how leaves grow wide. Development 2020, 147, dev193623. [Google Scholar] [CrossRef] [PubMed]
- Zhu, T.; Moschou, P.N.; Alvarez, J.M.; Sohlberg, J.J.; von Arnold, S. WUSCHEL-RELATED HOMEOBOX 2 is important for protoderm and suspensor development in the gymnosperm Norway spruce. BMC Plant Biol. 2016, 16, 19. [Google Scholar] [CrossRef] [PubMed]
- Hassani, S.B.; Trontin, J.F.; Raschke, J.; Zoglauer, K.; Rupps, A. Constitutive Overexpression of a Conifer WOX2 Homolog Affects Somatic Embryo Development in Pinus pinaster and Promotes Somatic Embryogenesis and Organogenesis in Arabidopsis Seedlings. Front. Plant Sci. 2022, 13, 838421. [Google Scholar] [CrossRef]
- Breuninger, H.; Rikirsch, E.; Hermann, M.; Ueda, M.; Laux, T. Differential expression of WOX genes mediates apical-basal axis formation in the Arabidopsis embryo. Dev. Cell 2008, 14, 867–876. [Google Scholar] [CrossRef]
- Hendelman, A.; Zebell, S.; Rodriguez-Leal, D.; Dukler, N.; Robitaille, G.; Wu, X.; Kostyun, J.; Tal, L.; Wang, P.; Bartlett, M.E.; et al. Conserved pleiotropy of an ancient plant homeobox gene uncovered by cis-regulatory dissection. Cell 2021, 184, 1724–1739.e1716. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Xu, L. Transcription Factors WOX11/12 Directly Activate WOX5/7 to Promote Root Primordia Initiation and Organogenesis. Plant Physiol. 2016, 172, 2363–2373. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, J.; Jia, H.; Liu, B.; Sun, P.; Hu, J.; Wang, L.; Lu, M. The WUSCHEL-related homeobox 5a (PtoWOX5a) is involved in adventitious root development in poplar. Tree Physiol. 2017, 38, 139–153. [Google Scholar] [CrossRef] [PubMed]
- Deveaux, Y.; Toffano-Nioche, C.; Claisse, G.; Thareau, V.; Morin, H.; Laufs, P.; Moreau, H.; Kreis, M.; Lecharny, A. Genes of the most conserved WOX clade in plants affect root and flower development in Arabidopsis. BMC Evol. Biol. 2008, 8, 291. [Google Scholar] [CrossRef]
- Park, S.O.; Hwang, S.; Hauser, B.A. The phenotype of Arabidopsis ovule mutants mimics the morphology of primitive seed plants. Proc. Biol. Sci. 2004, 271, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Gu, R.; Song, X.; Liu, X.; Yan, L.; Zhou, Z.; Zhang, X. Genome-wide analysis of CsWOX transcription factor gene family in cucumber (Cucumis sativus L.). Sci. Rep. 2020, 10, 6216. [Google Scholar] [CrossRef] [PubMed]
- Sajjad, M.; Wei, X.; Liu, L.; Li, F.; Ge, X. Transcriptome Analysis Revealed GhWOX4 Intercedes Myriad Regulatory Pathways to Modulate Drought Tolerance and Vascular Growth in Cotton. Int. J. Mol. Sci. 2021, 22, 898. [Google Scholar] [CrossRef]
- Minh-Thu, P.T.; Kim, J.S.; Chae, S.; Jun, K.M.; Lee, G.S.; Kim, D.E.; Cheing, J.J.; Song, S.I.; Nahm, B.H.; Kim, Y.-K. A WUSCHEL Homeobox Transcription Factor, OsWOX13, Enhances Drought Tolerance and Triggers Early Flowering in Rice. Mol. Cells 2018, 41, 781–798. [Google Scholar] [PubMed]
- Wang, L.Q.; Li, Z.; Wen, S.S.; Wang, J.N.; Zhao, S.T.; Lu, M.Z. WUSCHEL-related homeobox gene PagWOX11/12a responds to drought stress by enhancing root elongation and biomass growth in poplar. J. Exp. Bot. 2019, 71, 1503–1513. [Google Scholar] [CrossRef]
- Liu, R.; Wang, R.; Lu, M.Z.; Wang, L.Q. WUSCHEL-related homeobox gene PagWOX11/12a is involved in drought tolerance through modulating reactive oxygen species scavenging in poplar. Plant Signal. Behav. 2021, 16, 1866312. [Google Scholar] [CrossRef]
- Letunic, I.; Khedkar, S.; Bork, P. SMART: Recent updates, new developments and status in 2020. Nucleic Acids Res. 2020, 49, D458–D460. [Google Scholar] [CrossRef] [PubMed]
- Punta, M.; Coggill, P.; Eberhardt, R.; Finn, R. The Pfam protein families database. Nucleic Acids Res. 2011, 40, D290–D301. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 70 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Jin, J.; Guo, A.Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, W.; Zhu, D.; Hong, P.; Zhang, S.; Xiao, S.; Tan, Y.; Chen, X.; Xu, L.; Zong, X.; et al. Chromosome-scale genome assembly of sweet cherry (Prunus avium L.) cv. Tieton obtained using long-read and Hi-C sequencing. Hortic. Res. 2020, 7, 122. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Wu, Y.; Li, J.; Wang, X.; Zeng, Z.; Xu, J.; Liu, Y.; Feng, J.; Chen, H.; He, Y.; et al. TBtools-II: A “one for all, all for one” bioinformatics platform for biological big-data mining. Mol. Plant 2023, 16, 1733–1742. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general-purpose read summarization program. Bioinformatics 2013, 30, 923–930. [Google Scholar] [CrossRef]
- Cao, Y.; Han, Y.; Meng, D.; Li, G.; Li, D.; Abdullah, M.; Lin, Y.; Cai, Y. Genome-Wide Analysis Suggests the Relaxed Purifying Selection Affect the Evolution of WOX Genes in Pyrus bretschneideri, Prunus persica, Prunus mume, and Fragaria vesca. Front. Genet. 2017, 8, 78. [Google Scholar] [CrossRef]
- Xu, X.Z.; Che, Q.Q.; Cheng, C.X.; Yuan, Y.B.; Wang, Y.Z. Genome-wide identification of WOX gene family in apple and a functional analysis of MdWOX4b during adventitious root formation. J. Integr. Agric. 2022, 21, 1332–1345. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Genomic Position | Protein Length (aa) | Molecular Weight (Da) | Isoelectric Point | Subcellular Localization |
|---|---|---|---|---|---|---|
| PavWUS | FUN_038736 | chr_7: 24441104–24442540 | 305 | 33,845.92 | 5.91 | Nucleus. |
| PavWOX1 | FUN_026584 | chr_5: 34401315–34403970 | 403 | 45,466.65 | 8.49 | Nucleus. |
| PavWOX2 | FUN_019165 | chr_6: 7919462–7920784 | 261 | 29,121.23 | 7.71 | Nucleus. |
| PavWOX4 | FUN_005951 | chr_1: 47077176–47078228 | 224 | 25,281.56 | 9.30 | Nucleus. |
| PavWOX5 | FUN_012296 | chr_2: 40510721–40511406 | 180 | 20,439.96 | 9.30 | Nucleus. |
| PavWOX9 | FUN_032123 | chr_4: 3475851–3477812 | 413 | 45,418.52 | 6.87 | Nucleus. |
| PavWOX13A | FUN_024119 | chr_5: 18202603–18205376 | 220 | 25,412.49 | 5.32 | Nucleus. |
| PavWOX13B | FUN_038933 | chr_7: 25450872–25453321 | 273 | 31,202.77 | 5.59 | Nucleus. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, F.; Wang, H.; An, X.; Uwamungu, J.Y. A Genome-Wide Analysis of the WUSCHEL-Related Homeobox Transcription Factor Family Reveals Its Differential Expression Patterns, Response to Drought Stress, and Localization in Sweet Cherry (Prunus avium L.). Horticulturae 2024, 10, 370. https://doi.org/10.3390/horticulturae10040370
Deng F, Wang H, An X, Uwamungu JY. A Genome-Wide Analysis of the WUSCHEL-Related Homeobox Transcription Factor Family Reveals Its Differential Expression Patterns, Response to Drought Stress, and Localization in Sweet Cherry (Prunus avium L.). Horticulturae. 2024; 10(4):370. https://doi.org/10.3390/horticulturae10040370
Chicago/Turabian StyleDeng, Fei, Hongming Wang, Xiaojuan An, and Jean Yves Uwamungu. 2024. "A Genome-Wide Analysis of the WUSCHEL-Related Homeobox Transcription Factor Family Reveals Its Differential Expression Patterns, Response to Drought Stress, and Localization in Sweet Cherry (Prunus avium L.)" Horticulturae 10, no. 4: 370. https://doi.org/10.3390/horticulturae10040370
APA StyleDeng, F., Wang, H., An, X., & Uwamungu, J. Y. (2024). A Genome-Wide Analysis of the WUSCHEL-Related Homeobox Transcription Factor Family Reveals Its Differential Expression Patterns, Response to Drought Stress, and Localization in Sweet Cherry (Prunus avium L.). Horticulturae, 10(4), 370. https://doi.org/10.3390/horticulturae10040370