Abstract
An experiment was conducted with two-year-old ‘Fino 95’ lemon plants (Citrus limon Burm. f. cv. Fino) grafted on C. macrophylla (Citrus macrophylla Wester) to study the effects of salinity on plants inoculated with arbuscular mycorrhizal fungi (AMF). Half of the inoculated (+AM) and non-inoculated (−AM) plants were irrigated with half-strength Hoagland solution, and the remainder were inoculated with half-strength Hoagland solution + 30 mM NaCl. Ninety-eight days later, results showed that AMF had alleviated the negative effect of salinity on growth. Inoculation with AMF provided some protection against the damage that salinity caused on cellular membranes and improved the plant water status and turgor under saline conditions (Ψx and Π increased by 16% and 48%, respectively). The responses of mineral nutrition to salinity and AMF treatments were complex. P concentrations in the leaves and roots of +AM plants were lower than in those of −AM plants, but inoculation improved Ca2+ (by 20%), Mg2+ (24%), Fe2+ (21%), and Zn2+ (7%) nutrition in roots and also the Mg2+/Na+ ratio in leaves (33%), reducing the antagonistic effect of Na+ on Mg2+ nutrition in salt-treated plants. AMF could protect plants against salt stress through the maintenance of the gas exchange capacity and due to a better antioxidant response. All these positive effects of AMF contributed to mitigating the harmful effects of salinity stress on the plant growth performance of lemon trees grafted on C. macrophylla rootstock under salinity conditions.
1. Introduction
Citrus species stand out as the most productive and widely consumed fruit globally, constituting a vital component of the economy as an evergreen tree cultivated in 140 countries that span across various regions [1]. These regions can be predominantly found in warm temperate zones [2], which offer favorable conditions for citrus cultivation, albeit they are often characterized by arid, hot summers. A prime example of this is the south-eastern region of Spain, renowned for its significant citrus output, attributed to a warm Mediterranean climate and a fertile soil, which collectively foster an optimal growing environment [3,4]. However, the low precipitation levels of these areas create the necessity of implementing supplementary irrigation systems. Many of these arid and semi-arid regions grapple with groundwater salinization issues stemming from aquifer overexploitation and seawater intrusion. As a consequence, the available irrigation water presents with subpar quality due to the high concentrations of soluble salts, posing challenges for citrus cultivation, as citrus trees are particularly sensitive to salinity [5,6]. Moreover, in the Mediterranean regions, the temperature rise induced by climate change is projected to increase evapotranspiration, decrease rainfall, and, consequently, elevate irrigation demands for this crop, which already exhibits high irrigation requirements. With climate change becoming increasingly widespread, the diminishing freshwater resources pose a threat to the sustainability of irrigated agriculture and, therefore, to food security [7,8]. Thus, in citrus-producing regions, climate change is anticipated to exacerbate the already significant issues of drought and salinity stress in citrus.
On the other hand, in the northern hemisphere, fresh lemon (Citrus limon [L.] Osbeck) is mainly produced in the Mediterranean area. Specifically, in Spain, it is mainly grown in the southeastern region of the country, which is responsible for almost 90% of its production [9]. Among commercial citrus rootstocks, Citrus macrophylla is one of the most commonly used in lemon orchards in southeastern Spain, since growers used it to replace sour orange, which is strongly affected by the tristeza disease [10]. Due to its low vigor compared with other rootstocks, it presents a higher drought-stress tolerance, and its use is recommended for regions such as southeastern Spain, where the availability of water is not assured [11].
Salinity poses a significant environmental constraint that limits the productivity of citrus, classified as a salt-sensitive crop. The response of citrus to soil salinity varies depending on the rootstock, the scion, the soil composition, the irrigation method, and the climatic conditions. Moderate levels of salinity can disrupt physiological processes, diminish growth, and reduce fruit yield in citrus trees [5,12,13,14]. Salinity can have an impact on citrus plants through various mechanisms, including the inhibition of CO2 assimilation due to elevated concentrations of Cl− and Na+ in leaves [15]. Additionally, it can lead to nutrient imbalances by impeding the uptake of essential elements such as Ca2+, Mg2+, and K+ [16], while also triggering an increase in intracellular inorganic and organic solutes as the plant seeks osmotic adjustment to maintain leaf turgor [17]. Saline stress prompts plants to restrict stomatal opening, partly through abscisic acid (ABA) signaling, resulting in a decreased photosynthetic activity [18,19]. This alteration in photosynthesis disrupts the electron transport chain and heightens the production of reactive oxygen species. Variations in the response to salinity among citrus varieties and rootstocks have been linked to differences in their capacity to regulate transpiration and photosynthesis [20].
Mycorrhizal fungi, which develop a symbiotic relationship with plant roots, are believed to alleviate salt stress in host plants. Arbuscular mycorrhizal fungi (AMF) have been shown to enhance citrus plant growth and vigor, mitigating the damage caused by soil salinization [21,22,23,24,25,26,27]. The increased plant growth under salinity following mycorrhizal colonization with Rhizophagus irregularis, Funneliformis mosseae, or Paraglomus occultum described in these studies has been attributed to a more efficient nutrient uptake, particularly in the case of phosphorus (P), and physiological processes such as increased carbon dioxide exchange rates, transpiration, stomatal conductance, and water use efficiency, as well as an improved osmotic adjustment through organic and inorganic osmolytes, scavenging reactive oxygen species (ROS) through antioxidant enzymes and antioxidants, soil structural improvements, etc.
The research hypothesis of this study is that the inoculation of AMF can mitigate the adverse effects of salinity on citrus plants, particularly when considering both the rootstock and the scion components in commercial orchards. This hypothesis is based on previous findings indicating that AMF colonization has alleviated NaCl stress in commonly used citrus rootstocks in Spain [22,23]. However, in commercial orchards, citrus trees comprise both rootstock and scion. The novelty of this study is the use of AMF to alleviate the negative effects of salinity in typical commercial citrus plants. For this purpose, we used grafted citrus plants and evaluated the growth, mineral nutrition, and physiological responses of the plant in two-year-old lemon trees grafted on C. macrophylla, one of the rootstocks that had been previously studied, which is considered as a Cl− and Na+ accumulator under saline conditions [28,29]. This study aims to provide a comprehensive understanding of how AMF inoculation can alleviate the effects of salinity on commercial citrus plants that are commonly used in citrus orchards. The findings could offer valuable insights for developing sustainable agricultural practices, providing citrus growers with tools to enhance crop productivity and quality under saline conditions, and thereby addressing a significant challenge in the Mediterranean agriculture.
2. Materials and Methods
2.1. Plant Culture and Experimental Design
Uniformly sized, two-year-old ‘Fino 95’ lemon trees (Citrus limon [L.] Burm. fil.), grafted on C. macrophylla rootstock and obtained from a commercial nursery, were transplanted to plastic pots (3 L) on 15 March 2021. Soil was collected from the Estación Experimental de Torreblanca (Murcia, Spain), sieved (5 mm), diluted with quartz sand (1:3, soil/sand, v/v) and then sterilized in an autoclave for 1 h at 100 °C three times on alternating days. The sterilization bags with the substrate were closed and, after each sterilization step, they were stored in a clean and cool place. Original soil was analyzed by preparing a saturated paste with the following characteristics: organic carbon 1.38%; organic matter 2.37%; organic N 0.12%; C–N ratio 11.2; Olsen P 72.1 mg L−1. The nutrient concentrations (g L−1) were as follows: K 0.23; Ca 0.59; Mg 0.17; NO3− 0.62; SO42− 1.19; HCO3− 0.18; Cl− 1.08. Plants were placed in a greenhouse providing average day/night temperatures of 30–32/13–15 °C. Relative humidity was maintained at approximately 75%.
Twenty-five grams of Rhizophagus irregularis (Błaszk., Wubet, Renker & Buscot) C. Walker & A. Schüßler and Funneliformis mosseae (T.H. Nicolson & Gerd.) C. Walker & A. Schüßler in equal proportions (a mix of spores, mycorrhized roots, and substrate) as the mycorrhizal inoculum, propagated with the hybrid of Sorghum bicolor (L.) Moench and Sorghum sudanense (Piper) Hitch (Sorghum bicolor × sudanense) as the trap plant, were used to inoculate plants. In half of the pots (+AM), the inoculum was placed just around the root system and the pot was filled with the substrate. The inoculum was supplied by the Mycology-Mycorrhizas Laboratory, Department of Plant Biology, of the University of Murcia (Spain). Non-inoculated plants (−AM) were watered every 2 days with 500 mL of half-strength modified Hoagland’s solution [30] to maintain the moisture content of the substrate. Inoculated plants (+AM) were also watered regularly with the same solution, but without P. Seedlings were acclimated for 3 months and then were subjected to salinity treatments. For both the +AM and −AM seedlings, half of them were selected randomly for the non-salt treatment (0 mM NaCl), and the other half received the salt stress treatment (30 mM NaCl). The salt stress treatment was applied in steps of 10 mM NaCl per day to avoid an osmotic shock.
Plants were placed randomly in the greenhouse, and the experiment was laid out in a completely randomized block design. Experimental treatments consisted of factorial combinations of two components: mycorrhization (−AM and +AM) and salinity (0 and 30 mM NaCl). Each of the four treatments had 10 replicates, giving a total of 40 pots. Trees were harvested on 20 September 2021, 98 days after starting the NaCl treatments.
2.2. Determination of Physiological Parameters
Gas interchange parameters (net CO2 assimilation rate [A], transpiration [E], and stomatal conductance [gs]) were measured one day before harvest using an infrared gas analyzer Li-6400 (Li-Cor, Lincoln, NE, USA) equipped with a broad leaf chamber (6.0 cm2). All of the measurements were taken at a molar air flow of 300 μmol mol−1, a reference CO2 concentration of 400 μmol mol−1, and a photosynthetic photon flux density of 1200 μmol m−2 s−1. The intrinsic gas exchange efficiency was calculated as A/gs.
Stem water potential (Ψx) was measured at noon (12:00–14:00) with a Schölander-type pressure chamber (3000; Soil Moisture Equipment Corp., Santa Barbara, CA, USA; [31]) using mature leaves that had been enclosed in aluminum-foil-covered plastic envelopes at least 2 h before measurement. After Ψx measurement, leaves were tightly enclosed in aluminum foil and frozen at −18 °C. After thawing, leaf osmotic potential (Π) was measured in expressed cell sap collected from a syringe at 25 ± 1 °C and placed in an osmometer (Digital Osmometer, Wescor, Logan, UT, USA). Similar fresh leaves were hydrated by placing their petioles in distilled water at 4 °C for 24 h; after they were frozen at −18 °C, osmotic potential at full turgor (Π100) was determined by the same procedure that has been previously described.
2.3. Plant Growth
The relative growth rate (RGR) of the trunk section of the scion variety was calculated as in Tattini et al. [32], measuring the trunk when salinity treatments started and at the end of the experiment: RGR = (ln M2 − ln M1)/(t2 − t1), where M is the trunk section of the measured variety and t2 − t1 is the time elapsed between measurements.
At harvest, plant roots were carefully separated from the substrate and washed with distilled water. The fresh root systems were divided into two parts: one of them was used for the determination of mycorrhization, and the other was oven-dried to calculate the dry weight (DW) and analyze the nutrients. The leaves and stems were separated and weighed fresh. The total leaf area was measured using an Area Meter LI-3100C (LI-COR, Lincoln, NE, USA). The relative water content (RWC) of the leaves was measured. After the roots and stems were oven-dried at 60 °C for 48 h (until constant weight) and the leaves were freeze-dried, their DWs were determined.
2.4. Plant Analysis
The dried plant tissues were ground and stored for chemical analysis. They were ashed at 550 °C, ashes were dissolved in 0.7 N HNO3, and then P, Na, K, Mg, Ca, Fe, Cu, Zn, and Mn were analyzed by an inductively coupled plasma optical emission spectrometer Varian MPX Vista (Varian, Palo Alto, CA, USA). Chloride was extracted with bi-distilled water, using the method of Guilliam [33], and determined by ionic chromatography with a Dionex ICS-3000 (Dionex, Sunnyvale, CA, USA) ion chromatograph equipped with a conductivity detector and an AS11-HC anion exchange column.
Quaternary ammonium compounds (QAC) were measured by the Grieve and Grattan [34] method. Samples were extracted with deionized H2O and, after dilution with 2 N H2SO4, periodide crystals were formed with cold KI-I2 reagent. The absorbance at 365 nm was measured after periodide crystals were dissolved in 1,2-dichloroethane.
Free proline was analyzed by a modification of the acid ninhydrin method [35]. Fifty mg of freeze-dried leaves were extracted with 10 mL of 5-sulfosalicylic acid, shaken for 30 min with an orbital shaker, and centrifuged at 5000 rpm for 15 min. After the addition of glacial acetic acid and acid ninhydrin, color was developed at 100 °C, the chromophore was extracted with toluene, and the absorbance was measured at 520 nm.
Lipid peroxidation was determined by measuring malondialdehyde (MDA) using the Heath and Packer [36] method. Dry tissue was homogenized with a thiobarbituric acid–trichloroacetic acid mixture. After 45 min at 95 °C and centrifugation at 4000× g for 35 min, the absorbance was read at 532 and 600 nm to correct the unspecific turbidity.
2.5. Determination of Mycorrhizal Colonization
Root samples from every tree were taken from the middle part of the root system to make an estimation of mycorrhizal colonization. They were cleaned and stained with trypan blue according to the method of Phillips and Haymann [37] but using lactic acid instead of lactophenol. One hundred root segments per plant were mounted on slides, squashed by pressing on the coverslips, and quantified for AM colonization according to McGonigle et al. [38].
2.6. Statistical Analysis
Data were analyzed using analysis of variance (ANOVA) procedures with the SPSS software package (SPSS 7.5.1 for Windows, standard version, 1996). A two-way ANOVA procedure was used to discriminate the effects of the inoculation with AMF, the salinity level, and their interaction.
3. Results
3.1. Mycorrhizal Colonization and Plant Growth Analysis
Trees inoculated with AMF showed a high percentage of root colonization, with no significant effect of the salt level on the percentages of colonization (Table 1, Figure 1). There was no mycorrhizal colonization in the non-inoculated plants.

Table 1.
Root colonization, vesicular colonization, and arbuscular colonization of roots of ‘Fino 95’ lemon plants grafted on C. macrophylla and grown at two levels of salinity under greenhouse conditions.
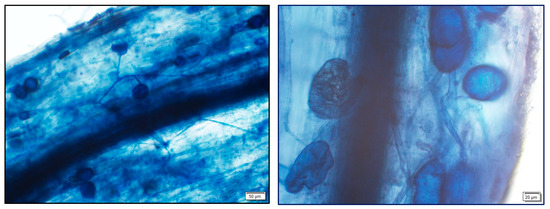
Figure 1.
Arbuscular mycorrhizal colonization of the mycorrhizal roots, details of hyphae and vesicles.
Salinity decreased the plant growth of the lemon trees grafted on C. macrophylla rootstock in a significant way, but this effect was different in −AM and +AM plants (Figure 2). Whereas salinity decreased the growth of −AM plants (23% of plant dry weight and 35% of RGR trunk section; Figure 2), no growth reduction was observed in +AM plants.
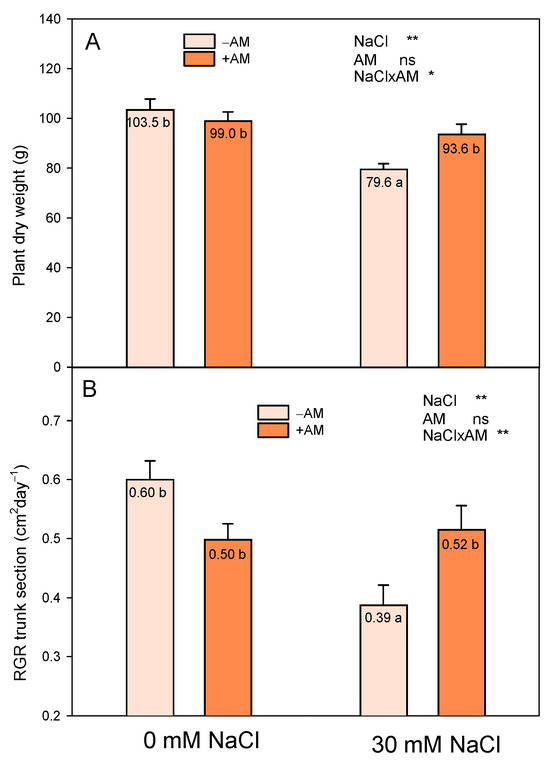
Figure 2.
Effect of inoculation with AMF (−AM non-inoculated plants, +AM inoculated plants) on total plant biomass (A) and RGR trunk section (B) of ‘Fino 95’ lemon plants grown at two levels of salinity (0 and 30 mM NaCl) under greenhouse conditions. * and ** indicate significant differences at the 0.05 and 0.01 levels of probability, respectively. Non-significant differences at the 0.05 level of probability are indicated as ns. Different letters indicate significant differences between treatments according to Duncan’s multiple range test at the 95% confidence level.
3.2. Physiological Changes
Our study revealed the change in leaf water potential in response to NaCl treatment and mycorrhizal inoculation (Table 2). Salinity significantly decreased the Ψx in both −AM and +AM plants. The reduction in Ψx due to salinity was greater in −AM plants (67%) than in +AM plants (19%), so AMF inoculation improved plant water status under saline conditions, increasing Ψx with regard to −AM plants (Table 2). In this experiment, only +AM plants under 30 mM NaCl slightly decreased their Π in order to maintain turgor potential (Table 2). In fact, salinity treatment reduced turgor in −AM plants, and only +AM plants maintained turgor values that were similar to those of the control (Table 2). The significant decrease in the osmotic potential at full turgor (Π100) by salinity observed in +AM plants highlighted the active solute accumulation in these plants (Table 2).
Table 2.
Effect of the inoculation with AMF (−AM non-inoculated plants, +AM inoculated plants) on water potential (Ψx, MPa), osmotic potential (Π, MPa), turgor potential (P, MPa), osmotic potential at full turgor (Π100, MPa), and malondialdehyde concentration (MDA, µmol kg−1 DW) of ‘Fino 95’ lemon plants grown at two levels of salinity (0 and 30 mM NaCl) under greenhouse conditions.
In our experiment, ‘Fino 95’ lemon plants did not accumulate proline as part of their osmotic response to the salinity treatment, and the increase in Π in +AM plants treated with 30 mM NaCl was partly due to a higher accumulation of QAC than in non-salinized plants (Figure 3).
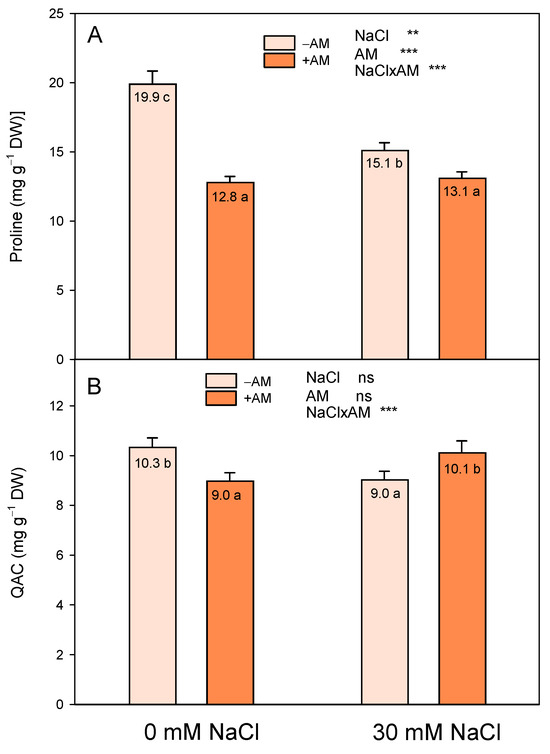
Figure 3.
Effect of inoculation with AMF (−AM non-inoculated plants, +AM inoculated plants) on the leaf concentrations of proline (A) and quaternary ammonium compounds (QAC) (B) of ‘Fino 95’ lemon plants grown at two levels of salinity (0 and 30 mM NaCl) under greenhouse conditions. ** and *** indicate significant differences at the 0.01 and 0.001 levels of probability, respectively. Non-significant differences at the 0.05 level of probability are indicated as ns. Different letters indicate significant differences between treatments according to Duncan’s multiple range test at the 95% confidence level.
In our experiment, MDA data showed that salinity produced a significantly higher lipid peroxidation in −AM plants, whereas oxidative damage due to salinity was reduced in +AM plants (Table 2).
3.3. Nutritional Changes
Under saline conditions, citrus plants transported Cl− to leaves, as shown in the present research by the increase in foliar Cl− in these conditions (Table 3). However, no effect of the AMF on the foliar Cl− concentrations under control or saline conditions was found in our experiment (Table 3). Salinity increased the root Na+ concentrations and did not reduce K+ in roots nor in leaves. AMF did not modify Na+ concentrations in roots or leaves but increased the foliar K+ concentration under non-saline conditions (Table 3 and Table 4). However, this surge was not enough to increase the K+/Na+ ratio in the leaves of control plants. Similarly, the decrease in foliar K+ due to AMF under saline conditions was not enough to reduce the K+/Na+ ratio of these plants. However, the K+/Na+ ratio was increased by AMF under non-saline conditions but was not modified with 30 mM NaCl (Table 4). As AMF did not increase the abundance of K+ in roots or leaves nor did they decrease that of Na+ under salinity, K+/Na+ ratios were not higher in +AM plants under saline conditions.
Table 3.
Effect of inoculation with AMF (−AM non-inoculated plants, +AM inoculated plants) on the leaf nutrient concentrations and leaf ionic balance of ‘Fino 95’ lemon plants grown at two levels of salinity (0 and 30 mM NaCl) under greenhouse conditions. Cl−, Na+, P, and K expressed as mmol kg−1 DW; Cu and Mn expressed as mg kg−1 DW.
Table 4.
Effect of inoculation with AMF (−AM non-inoculated plants, +AM inoculated plants) on the root nutrient concentrations and root ionic balance of ‘Fino 95’ lemon plants grown at two levels of salinity (0 and 30 mM NaCl) under greenhouse conditions. Cl−, Na+, P, and K expressed as mmol kg−1 DW; Cu and Mn expressed as mg kg−1 DW.
Inoculated plants of this experiment had higher root concentrations of Ca2+ and Mg2+ than −AM plants for both salinity treatments (Figure 4). Nonetheless, in plants exposed to salinity, the increase in these nutrients due to AMF was not enough to improve the Ca2+/Na+ and Mg2+/Na+ ratios of the roots (Table 4). Although the Ca2+ and Mg2+ concentrations of mycorrhizal lemon roots were increased under saline conditions, the root Na+ levels were also heightened (although not significantly) and, as a consequence, the Ca2+/Na+ and Mg2+/Na+ ratios of the +AM and −AM plants were similar (Table 4). However, the Mg2+/Na+ ratio increased in the leaves of mycorrhizal lemon plants exposed to salt stress.
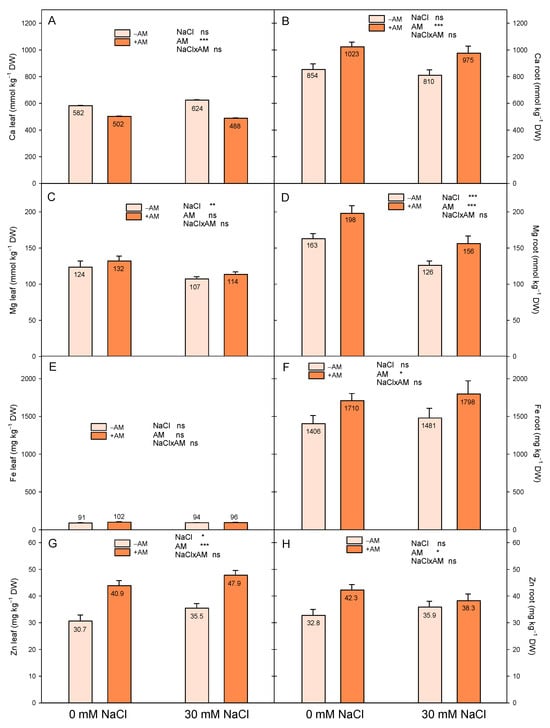
Figure 4.
Effect of inoculation with AMF (−AM non-inoculated plants, +AM inoculated plants) on the leaf (A,C,E,G) and root (B,D,F,H) concentrations of Ca (A,B), Mg (C,D), Fe (E,F), and Zn (G,H) of ‘Fino 95’ lemon plants grown at two levels of salinity (0 and 30 mM NaCl) under greenhouse conditions. Ca and Mg expressed as mmol kg−1 DW; Fe and Zn expressed as mg kg−1 DW. *, ** and *** indicate significant differences at the 0.05, 0.01 and 0.001 levels of probability, respectively. Non-significant differences at the 0.05 level of probability are indicated as ns.
The concentrations of P and Mn in the roots and leaves of +AM plants were lower than those found in −AM plants (Table 3 and Table 4). With regard to the Fe concentration, +AM plants had increased root levels of Fe, in both the control and saline treatments, but their leaf Fe concentration was not modified (Figure 4). On the other hand, Zn was significantly increased by AMF in both roots and leaves of control and salinized plants (Figure 4).
3.4. Gas Interchange Parameters
The present study indicated an inhibitive effect of salt stress on the photosynthetic rate and stomatal conductance (Figure 5). However, this effect of salinity on A and gs depended on the inoculation, since both parameters were significantly decreased by the exposure to 30 mM NaCl in −AM plants, remaining unaltered in +AM plants (Figure 5). We observed similar gs values in +AM and −AM plants under saline conditions and a lower reduction in gs in +AM plants than in −AM plants with increasing salinity. On the other hand, inoculation significantly decreased the photosynthetic capacity of the plants under control conditions (Figure 5). No effects were observed on the transpiration rate (E) nor on the intrinsic photosynthetic efficiency (A/gs).
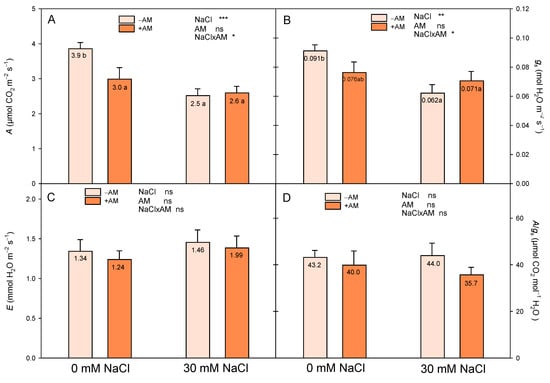
Figure 5.
Effect of inoculation with AMF (−AM non-inoculated plants, +AM inoculated plants) on the net CO2 assimilation rate (A, µmol CO2 m−2s−1) (A), stomatal conductance (gs, mol H2O m−2s−1) (B), transpiration (E, mmol H2O m−2s−1) (C), and intrinsic gas exchange efficiency (A/gs, µmol CO2 mol−1 H2O) (D) of ‘Fino 95’ lemon plants grown at two levels of salinity (0 and 30 mM NaCl) under greenhouse conditions. *, ** and *** indicate significant differences at the 0.05, 0.01 and 0.001 levels of probability, respectively. Non-significant differences at the 0.05 level of probability are indicated as ns. Different letters indicate significant differences between treatments according to Duncan’s multiple range test at the 95% confidence level.
4. Discussion
Some previous reports in citrus plants showed a decrease in root mycorrhizal colonization with salinity when compared with no-NaCl treatment [39,40,41], which was attributed to both a reduced hyphal extension of AMF and a decreased availability of inoculum sporulation due to a poorer development of spores. However, the salinity levels used in these previous studies were much higher (75, 100, or even 150 mM NaCl) than those used in this experiment (30 mM NaCl), and we only observed a slight and non-significant decrease in the arbuscular colonization due to salinity.
Our results indicated that AMF mitigated the harmful effects of salinity stress the on plant growth performance of lemon trees grafted on C. macrophylla rootstock. A similar positive resistance to soil salinity was previously observed in citrus plants by some authors when R. irregularis and/or F. mosseae were used [23,25,27,41]. In all of these studies, it was concluded that mycorrhizal citrus plants had a better growth than non-mycorrhizal ones under salinity conditions. A meta-analysis of the published studies to determine how AMF influences plant responses under salt-stressed conditions indicated that mycorrhizal inoculation had a significant impact on some physiological and biochemical variables of the plant related to plant growth, photosynthesis, and defense against oxidative damage under salinity stress [42]. In this meta-analysis, it was confirmed that AMF inoculation increases the biomass of host plants and that the effect of AMF on plant growth is more prominent under salinity stress than under normal conditions. The different mechanisms that AMF utilizes to enhance salt tolerance in host plants have been summarized in several reviews [43,44,45,46,47,48]: enhancing nutrient uptake, inhibiting a high uptake of Na+ and Cl− and their transport to plant shoots, improving water uptake, accumulating proline and polyamines or having a better oxidant response. Other physiological mechanisms induced by AMF may include osmotic adjustment, which helps to maintain the leaf turgor pressure, and effects on photosynthesis, transpiration, stomatal conductance, and water use efficiency. In our experiment, some of these mechanisms have been involved in the beneficial effect of AMF on the growth of ‘Fino 95’ lemon plants under saline conditions.
First of all, a positive effect of AMF on the plant water status was observed. Under salinity stress, this is the first cause of plant growth suppression. The reduction in Ψx in both −AM and +AM plants by salinity is associated with the reduction in water potential in the root environment and, therefore, with the water absorption capacity of the plant. The lower reduction in Ψx due to salinity in +AM plants indicated that AMF inoculation improved plant water status under saline conditions. Changes in leaf water potential under saline conditions stimulated by AMF have been previously described [41,43,47]. The higher absorption of water of +AM plants could be explained by an improved hydraulic conductivity attributed to AMF-induced altered root morphology, as well as their ability to explore soil well beyond depletion due to the external hyphae, which can help the host plant to absorb more water from soils [49,50]. Furthermore, the better water status in +AM plants may be also explained by the AMF-regulated expression of aquaporin (AQP) genes present in the leaves and roots of salt-stressed plants. AQPs are the crucial pathway to transport water and nutrients, and AMF upregulated their expression in the host under salinity conditions, revealing the important regulation of plant water by AMF [41,51]. Recent studies in trifoliate orange have shown that fourteen AQPs were dramatically induced by AMF inoculation under salt stress [41], revealing that AMF can increase the expression of AQP genes under salt stress, which improves the membrane’s water permeability and stimulates the host’s water transport.
Additionally, mycorrhizal plants under salt stress showed a better osmoregulation. Under saline conditions, plants can use osmoregulation as a mechanism to tolerate salt stress, lowering their osmotic potential due to the accumulation of osmolytes [5,14]. Under saline conditions, +AM plants reduced their Π, and this reduction was due to an active solute accumulation, not to a concentration effect for a possible dehydration of plants. This lowering of Π allowed them to maintain the turgor potential within similar values to those of control plants (Table 2). Some of the key osmolytes that help plants to maintain cell turgor are proline and QAC [52]. A large number of plant species accumulate proline in response to salinity, since it plays a role in their defense against salinity stress. In our experiment, the protection of +AM plants against salinity was not mediated by an increased accumulation of proline, since lower proline concentrations were found in these plants. Some authors have stated that these lower proline concentrations in +AM plants occur because they suffer less stress than −AM plants [53,54]. In fact, in our experiment, +AM plants under salt stress had a better water status, as revealed by their higher Ψx with regard to −AM plants. In addition, the inoculation of AMF has been reported to modulate the biosynthesis of glycine betaine, probably the most thoroughly studied QAC, thereby protecting and improving photosynthetic efficiency [55,56]. In our experiment, the decrease in Π in +AM plants treated with 30 mM NaCl was partly due to a higher accumulation of QAC than in non-salinized plants (Figure 3). Neither Na+ nor Cl− concentrations were involved in the osmotic adjustment observed in +AM plants in the 30 mM NaCl treatment.
With regard to the effect of AMF on the inhibition of a high uptake of Na+ and Cl−, no effects were found. The salinity level used in the present experiment was low, so it is possible that Na+ concentrations in roots and leaves were not high enough for the AMF to regulate Na+ uptake, and the results show no effect of the fungi on the Na+ concentrations in roots or leaves under control or saline conditions (Table 3 and Table 4). Previous studies have suggested the possibility of a buffering effect of AMF on the uptake of Na+ when the content of the latter is within acceptable limits [43,57]. Moreover, the lack of an accumulation of Na+ in shoots in parallel to Cl− in our experiment could be related to the scion vigor. In grafted plants, the rootstock is not the only element that controls the Cl− and Na+ transport to the shoot; the scion may also have an influence on their accumulation in leaves [12,58]. The grafting of lemon scion varieties on sour orange and on C. macrophylla produced variable effects on the Na+ and Cl− concentrations of the leaf [58], so the mechanism by which some rootstocks reduce the concentration of ions seems to depend on the vigor of the scion and on water requirements [12].
However, AMF can modify the altered nutrient uptake that can occur due to the high concentration of Na+ and Cl−. Maintaining a high K+/Na+ ratio in inoculated plants is important for proper metabolic functioning [59]. In citrus plants, mycorrhizal symbiosis enhances salinity tolerance through the selective absorption of K+/Na+ but not through their selective transport from roots to leaves, which is reduced by mycorrhizal treatment under salt stress [60]. In our experiment, since AMF did not increase the abundance of K+ in roots or leaves nor did they decrease that of Na+ under salinity, plant inoculation offered no advantage with regard to maintaining higher K+/Na+ ratios in +AM plants under saline conditions. On the other hand, it has been shown that AMF selectively takes up other elements, such as Mg2+ and Ca2+, while diminishing the uptake of potentially toxic Na+ [21,57,61] to keep the internal Mg2+/Na+ and Ca2+/Na+ ratios within narrow limits that help to maintain the integrity of the membrane and protect the host plants against salt damage. Although Ca2+ and Mg2+ concentrations in mycorrhizal lemon roots were increased under saline conditions, Na+ levels in the root were also heightened and, as a consequence, the rise in these nutrients was not enough to improve the Ca2+/Na+ and Mg2+/Na+ ratios of the roots. However, the Mg2+/Na+ ratio increased in the leaves of mycorrhizal lemon plants exposed to salt stress, which suggests that mycorrhizal colonization alleviated salt stress by reducing the antagonistic effect of Na+ [62].
The positive effect of AMF under saline conditions can also be related to the concentration of some micronutrients. An excessive amount of Na+ and Cl− in the soil contributes to limiting the availability of Fe, Zn, and Cu [63]. However, extraradical hyphae can bridge the zone of nutrient depletion adjacent to the root and thus increase the availability of elements of low mobility, such as Zn, by transporting them from more distant sites across the hyphae to the plant root [64,65]. This can be the reason why Zn concentration in both roots and leaves of control and salinized plants was significantly increased by AMF (Table 3 and Table 4). In addition, AMF contributed to better Fe nutrition in both the control and saline treatments. On the contrary, although mycorrhizal hyphae also increased the availability of the immobile Cu in the soil by translocating it from more remote sites through the hyphae to the plant root [64], we found no effect of AMF on the Cu concentration in roots and leaves. The lower Mn concentrations found in +AM plants could be due to a reduction in Mn uptake by hyphae in +AM plants. The solubility of Mn in the soil may decrease due to changes in the oxidation state influenced by the pH of the mycorrhizosphere, or to a decreased Mn4+ reducing potential in the rhizosphere of mycorrhizal plants, likely attributable to a smaller population of Mn-reducing organisms [66,67].
Another important nutrient that can be modified by AMF is P. Mycorrhizal association increases the acquisition of P by the host plant [68], and the improved growth of mycorrhizal plants in saline conditions is primarily related to a mycorrhiza-mediated enhancement of the host plant P nutrition [23,69,70]. In the present experiment, +AM plants were not fertilized with P, and we expected results to be similar to those previously found in other experiments where +AM plants had not been fertilized with P; besides that, they had high P concentrations because of the P input coming from the fungus [22,71]. All of these experiments were conducted in pots containing substrate composed by soil/sand (1:3) where, due to the high levels of CaCO3 (19%, typical of southeastern Spain), a significant portion of the high soil P concentration (72 ppm Olsen P) could be in insoluble form. However, we anticipated that the fungal hyphae secretion of acid and alkaline phosphatases [42] would release and solubilize enough P to provide an adequate P nutrition throughout the experiment, as has been observed in previous experiments [22,71]. Contrary to preceding results, the concentrations of P in the roots and leaves of +AM plants were lower than those found in −AM plants (Table 3 and Table 4). However, +AM plants grown without P fertilization were able to take up high amounts of P (25.7 mg kg−1 DW, Table 3). Even if these P concentrations were below the optimum range of P for adult citrus trees (37–50 mg kg−1 DW, [72]), they were sufficient to sustain the proper growth of citrus lemon plants (Figure 2) and adequate to support the healthy growth of young citrus seedlings without any deficiency symptoms. In a previous experiment performed with C. macrophylla plants fed with 0 mM of P, foliar concentrations of P in both +AM and −AM plants subjected to P starvation were even lower than those found in this experiment, and no visual deficiency symptoms were found in plants [73]. The results found in our experiment showed that, even if AMF can counteract to some extent the imposed P starvation, the application of a regular P fertilization to −AM plants certainly affected the P availability and may explain the results obtained with this particular combination of plants and AMF. In light of these results and due to the P concentrations found in +AM plants, we suspect that this P starvation could have produced a new stress situation; although this circumstance did not affect plant growth, it probably modified the balanced AMF/plant relationship.
Another physiological mechanism that AMF may alter under salinity stress is photosynthesis. Salt stress impedes photosynthesis and brings an enormous decline in crop productivity. However, our results showed that AMF could protect plants against salt stress through the maintenance of the gas exchange capacity, since there was no decrease in A due to salinity. According to previous results, we observed similar gs values in +AM and −AM plants under saline conditions and a lower reduction in gs in +AM plants than in −AM plants with increasing salinity, which suggests a decreased reduction in CO2 diffusion through the stomata and, hence, a better water status in +AM plants [74,75]. Consistent with our results, a positive effect of AMF colonization on the gas exchange capacity of salinity-exposed citrus plants was previously observed [22,24,76]. The positive role of AMF symbiosis in the alleviation of the negative effects of salinity on the photosynthetic capacity of the plant has been summarized in several reports [42,47,74]: an improvement in water status that results in maintaining a higher stomatal conductance and, consequently, a better CO2 assimilation; due to a higher expression of Rubisco, an increase in its activity, which allows the abatement of the intercellular CO2 concentration and ensures the protection of the photosynthetic capacity; maintenance of the integrity of PSII as a result of a higher concentration of polyamines and glycine betaine, which stabilizes PSII pigment–protein complexes, as well as the protection of the activities of enzymes such as Rubisco; the maintenance of a higher chlorophyll and carotenoid concentration by improving the uptake of Mg2+, enhancing photosynthetic efficiency via a proficient conversion of harvested light into chemical energy; and the minimization of NPQ. In our experiment, salinity did not decrease the photosynthetic rate in +AM plants, probably due to some of these advantages that AMF provides to plants to alleviate the effects of salinity. In this way, +AM plants under 30 mM NaCl improved their water status with regard to –AM plants (Table 2), and they had higher gs than −AM plants (Figure 5); additionally, +AM plants under salinity treatment had a higher QAC concentration (Figure 3), which probably helped to maintain the integrity of PSII; and finally, the higher concentration of Mg2+ in the plant (Figure 4) could be probably related with a higher photosynthesis rate through a higher chlorophyll concentration.
On the other hand, inoculation significantly decreased the photosynthetic capacity of the plants under control conditions, probably due to the fact that the relationship established between the plants and AMF under non-saline conditions was not a mutualistic relationship as balanced as those we found in previous studies with the same rootstock [22]. For a given plant species, this relationship could differ among growth stages. Some plant species show a much greater benefit from an association with an AM fungus in the seedling stage than being adult plants [75]. In our previous studies, the colonization of C. macrophylla plants by R. irregularis and F. mosseae significantly improved gaseous interchange parameters at the seedling stage [22]. In the present experiment, we used older C. macrophylla plants; moreover, these plants were grafted with ‘Fino 95’ lemon, which could have had an influence on their response to AMF. In addition to all of this, the AMF–host plant relations are context-dependent and can be mediated by environmental conditions [76,77]. As Jin et al. [78] pointed out, the C cost analyses of plants suggest that the effectiveness of AMF varies from mutualistic to parasitic, which depends primarily on the soil nutrient availability. Under our P starvation conditions, AMF did not improve plant growth in non-saline conditions, even when plants received most likely a major part of P via the AMF pathway, as was previously found [79,80,81]. Under these conditions, the photosynthesis rate was reduced, so the amount of C available for the plant and the fungus also decreased, reducing plant growth. The C-for-nutrient trade is the key factor in predicting the outcome of AMF symbioses, but our results showed a complex “cost/benefit” analysis.
Finally, as per our results, AMF mitigated the harmful effects of salinity stress on plant growth due to a better antioxidant response. Exposure to salt stress can lead to a detrimental impact on membrane integrity due to the generation of reactive oxygen species (ROS), which induce the release of electrolytes from cellular membranes, consequently disrupting the functionality of those membranes and upsetting the metabolic balance [82]. The lipid membrane of cells is damaged by oxidative burst, resulting in lipid peroxidation that can be measured by the content of malondialdehyde (MDA), the final product. In our experiment, no accumulation of MDA was observed in inoculated plants by salinity, so AMF alleviates salt-induced oxidative damage by protecting membrane lipids from ROS oxidation, as previously observed [83,84]. Furthermore, AMF-inoculated plants have higher levels of polyunsaturated fatty acids in membranes, thus enhancing plasma membrane integrity, reducing cellular component leakage, and leading to decreased ROS production [55].
5. Conclusions
The results of this study support the research hypothesis that the inoculation of AMF can mitigate the adverse effects of salinity on citrus plants, particularly in terms of plant growth and physiological responses. Whereas AMF did not consistently improve mineral nutrition or gas exchange parameters, their positive impact on plant growth under saline conditions is noteworthy. However, the complex nature of the relationship between AMF and citrus plants suggests that further research is needed to fully understand their interaction, particularly considering factors such as plant age and the influence of the scion on the rootstock response. Future studies should focus on exploring the chemical signaling between the root and the aerial parts to comprehensively understand the effect of the rootstock on the scion. This will provide deeper insights into the intricate dynamics of AMF–plant interactions and their implications for improving citrus cultivation practices.
This research underscores the significance of utilizing AMFs as viable options as biofertilizers in the cultivation of citrus trees, especially in regions where saline water is commonly used for irrigation. By harnessing the symbiotic relationship between plants and AMFs, citrus growers can mitigate the detrimental effects of salinity on crop productivity, offering valuable insights for developing sustainable agricultural practices in the Mediterranean regions. Future research endeavors should explore the optimization of the symbiotic relationship between AMFs and citrus plants to maximize their potential benefits and address the remaining challenges in Mediterranean agriculture.
Author Contributions
Conceptualization, J.M.N., O.P.-T., and A.M.; methodology, J.M.N., O.P.-T., and A.M.; formal analysis, J.M.N.; investigation, J.M.N., O.P.-T., and A.M.; data curation, J.M.N. and O.P.-T.; writing—original draft preparation, J.M.N.; writing—review and editing, J.M.N. and A.M.; funding acquisition, J.M.N. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by 15369/PI/10 Fundación Séneca (Region of Murcia, Spain) and POI07-12 (Instituto Murciano de Investigación y Desarrollo Agrario y Medioambiental), as well as the FEDER Funds.
Data Availability Statement
Data will be made available upon request. The raw data supporting the conclusions of this article will be made available by the authors on request.
Acknowledgments
We acknowledge Eva Arques for her help with the analysis..
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Rasera, J.B.; da Silva, R.F.; Mourão Filho, F.D.A.A.; Delbem, A.C.B.; Saraiva, A.M.; Sentelhas, P.C.; Marques, P.A.A. Climate change and citriculture: A bibliometric analysis. Agronomy 2023, 13, 723. [Google Scholar] [CrossRef]
- Abobatta, W.F. Citriculture and climate change. Mod. Concepts Dev. Agron. 2020, 6, 000639. [Google Scholar] [CrossRef]
- Navarro, L. The Spanish citrus industry. Acta Hortic. 2015, 1065, 41–48. [Google Scholar] [CrossRef]
- Ben Amor, R.; de Miguel Gomez, M.D. Competitiveness of Spanish orange sector in the Mediterranean area. Acta Hortic. 2020, 1292, 23–30. [Google Scholar] [CrossRef]
- Maas, E.V. Salinity and citriculture. Tree Physiol. 1993, 12, 195–216. [Google Scholar] [CrossRef] [PubMed]
- Grattan, S.R.; Díaz, F.J.; Pedrero, F.; Vivaldi, G.A. Assessing the suitability of saline wastewaters for irrigation of Citrus spp.: Emphasis on boron and specific-ion interactions. Agric. Water Manag. 2015, 157, 48–58. [Google Scholar] [CrossRef]
- Soares, D.; Paço, T.A.; Rolim, J. Assessing climate change impacts on irrigation water requirements under Mediterranean conditions—A review of the methodological approaches focusing on maize crop. Agronomy 2023, 13, 117. [Google Scholar] [CrossRef]
- Yetik, A.K.; Sen, B. Evaluation of the impacts of climate change on irrigation requirements of maize by CROPWAT Model. Gesunde Pflanz. 2022, 75, 1297–1305. [Google Scholar] [CrossRef]
- MAGRAMA. Anuario de Estadística Agraria 2021; Ministerio de Agricultura, Pesca y Alimentación, Madrid, Spain. Available online: https://www.mapa.gob.es/es/estadistica/temas/estadistica-diqital/ (accessed on 10 October 2023).
- Pérez-Pérez, J.G.; Porras, I.; Garcia-Lidon, A.; Botía, P.; Garcia-Sanchez, F. ‘Fino’ lemon clones compared with two other lemon varieties on two rootstocks in Murcia (Spain). Sci. Hortic. 2005, 106, 530–538. [Google Scholar] [CrossRef]
- Robles, J.M.; Botía, P.; Pérez-Pérez, J.G. Sour orange rootstock increases water productivity in deficit irrigated ‘Verna’ lemon trees compared with Citrus macrophylla. Agric. Water Manag. 2017, 186, 98–107. [Google Scholar] [CrossRef]
- Storey, R.; Walker, R.R. Citrus and salinity. Sci. Hortic. 1999, 78, 39–81. [Google Scholar] [CrossRef]
- García-Sánchez, F.; Jifon, J.; Carvajal, M.; Syvertsen, J.P. Gas exchange, chlorophyll and nutrient contents in relation to Na+ and Cl− accumulation in ‘Sunburst’ mandarin grafted on different rootstock. Plant Sci. 2002, 162, 705–712. [Google Scholar] [CrossRef]
- Syvertsen, J.P.; Garcia-Sanchez, F. Multiple abiotic stresses occurring with salinity stress in citrus. Environ. Exp. Bot. 2014, 103, 128–137. [Google Scholar] [CrossRef]
- García-Sánchez, F.; Syvertsen, J.P. Salinity tolerance of Cleopatra mandarin and Carrizo citrange citrus rootstocks seedlings is affected by CO2 enrichment during growth. J. Amer. Soc. Hort. Sci. 2006, 131, 24–31. [Google Scholar] [CrossRef]
- Romero-Aranda, R.; Moya, L.; Tadeo, F.R.; Legaz, F.; Primo-Millo, E.; Talón, M. Physiological and anatomical disturbances induced by chloride salts in sensitive and tolerant citrus: Beneficial and detrimental effects of cations. Plant Cell Environ. 1998, 21, 1243–1253. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanism of salinity tolerance. Ann. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Cadenas, A.; Vives, V.; Zandalinas, S.I.; Manzi, M.; Sánchez-Pérez, A.M.; Pérez-Clemente, R.M.; Arbona, V. Abscisic acid: A versatile phytohormone in plant signaling and beyond. Curr. Protein Pept. Sci. 2015, 16, 413–434. [Google Scholar] [CrossRef] [PubMed]
- López-Climent, M.F.; Arbona, V.; Pérez-Clemente, R.M.; Gómez-Cadenas, A. Relationship between salt tolerance and photosynthetic machinery performance in citrus. Environ. Exp. Bot. 2008, 62, 176–184. [Google Scholar] [CrossRef]
- Arbona, V.; López-Climent, M.F.; Pérez-Clemente, R.M.; Gómez-Cadenas, A. Maintenance of a high photosynthetic performance is linked to flooding tolerance in citrus. Environ. Exp. Bot. 2009, 66, 135–142. [Google Scholar] [CrossRef]
- Evelin, H.; Kapoor, R.; Giri, B. Arbuscular mycorrhizal fungi in alleviation of salt stress: A review. Ann. Bot. 2009, 104, 1263–1280. [Google Scholar] [CrossRef]
- Navarro, J.M.; Pérez-Tornero, O.; Morte, A. Alleviation of salt stress in citrus seedlings inoculated with arbuscular mycorrhizal fungi depends on the rootstock salt tolerance. J. Plant Physiol. 2014, 171, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Navarro, J.M.; Andujar, S.; García-Olmos, B.; Rodriguez-Morán, M.; Pérez-Tornero, O.; Morte, A. Arbuscular mycorrhizal fungi influence the response of citrus rootstock seedlings to salinity. Acta Hortic. 2011, 922, 245–252. [Google Scholar] [CrossRef]
- Wu, Q.S.; Zou, Y.N.; He, X.H. Contributions of arbuscular mycorrhizal fungi to growth, photosynthesis, root morphology and ionic balance of citrus seedlings under salt stress. Acta Physiol. Plant. 2010, 32, 297–304. [Google Scholar] [CrossRef]
- Wu, Q.S.; Zou, Y.N.; He, X.H. Mycorrhizal symbiosis enhances tolerance to NaCl stress through selective absorption but not selective transport of K+ over Na+ in trifoliate orange. Sci. Horti. 2013, 160, 366–374. [Google Scholar] [CrossRef]
- Murkute, A.A.; Sharma, S.; Singh, S.K.; Patel, V.B. Response of mycorrhizal citrus rootstock plantlets to salt stress. Indian J. Hort. 2009, 66, 456–460. [Google Scholar]
- Tang, C.; Zhang, Z.; Yu, L.; Li, Y. Research progress of arbuscular mycorrhizal fungi promoting citrus growth. Horticulturae 2023, 9, 1162. [Google Scholar] [CrossRef]
- Conesa, A.; Legua, P.; Navarro, J.M.; Pérez-Tornero, O.; García-Lidón, A.; Porras, I. Recovery of different Citrus rootstock seedlings previously irrigated with saline waters. J. Am. Pomol. Soc. 2011, 65, 158–166. [Google Scholar]
- Nieves, M.; Cerdá, A.; Botella, M.A. Salt tolerance of two lemon scions measured by leaf chloride and sodium accumulation. J. Plant Nutr. 1991, 14, 623–636. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The Water Culture Method for Growing Plants without Soil; Circular—California Agricultural Experiment Station: Berkeley, CA, USA, 1950; p. 347. [Google Scholar]
- Schölander, P.F.; Hammel, H.T.; Bradstreet, E.D.; Hemmingsen, E.A. Sap pressure in plants. Science 1965, 149, 920–922. [Google Scholar] [CrossRef]
- Tattini, M.; Gucci, R.; Romani, A.; Baldi, A.; Everard, J.D. Growth, gas exchange and ion content in Olea europaea plants during salinity and subsequent relief. Physiol. Plant. 1995, 95, 203–210. [Google Scholar] [CrossRef]
- Guilliam, M.G. Rapid measurement of chloride in plant materials. Soil Sci. Soc. Am. Proc. 1971, 35, 512–513. [Google Scholar] [CrossRef]
- Grieve, C.M.; Grattan, S.R. Rapid assay for determination of water soluble quaternary amino compounds. Plant Soil 1983, 70, 303–307. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts. I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Phillips, J.M.; Hayman, D.S. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 1970, 55, 158–161. [Google Scholar] [CrossRef]
- McGonigle, T.P.; Miller, M.H.; Evans, D.G.; Fairchild, G.L.; Swan, J.A. A new method which gives an objective measure of colonization of roots by vesicular-arbuscular mycorrhizal fungi. New Phytol. 1990, 115, 495–501. [Google Scholar] [CrossRef] [PubMed]
- He, J.D.; Li, J.L.; Wu, Q.S. Effects of Rhyzoglomus intraradices on plant growth and root endogenous hormones of trifoliate orange under salt stress. J. Anim. Plant Sci. 2019, 29, 245–250. [Google Scholar]
- Hadian-Deljou, M.; Esna-Ashari, M.; Mirzaieasl, A. Alleviation of salt stress and expression of stress-responsive gene through the symbiosis of arbuscular mycorrhizal fungi with sour orange seedlings. Sci. Hortic. 2020, 268, 109373. [Google Scholar] [CrossRef]
- Cheng, X.F.; Wu, H.H.; Zou, Y.N.; Wu, Q.S.; Kuča, K. Mycorrhizal response strategies of trifoliate orange under well-watered, salt stress, and waterlogging stress by regulating leaf aquaporin expression. Plant Physiol. Biochem. 2021, 162, 27–35. [Google Scholar] [CrossRef]
- Dastogeer, K.M.G.; Zahan, M.I.; Tahjib-Ul-Arif, M.; Akter, M.A.; Okazaki, S. Plant Salinity Tolerance Conferred by Arbuscular Mycorrhizal Fungi and Associated Mechanisms: A Meta-Analysis. Front. Plant Sci. 2020, 11, 588550. [Google Scholar] [CrossRef]
- Ding, Y.E.; Fan, Q.F.; He, J.D.; Wu, H.H.; Zou, Y.N.; Wu, Q.S.; Kuča, K. Effects of mycorrhizas on physiological performance and root TIPs expression in trifoliate orange under salt stress. Arch. Agron. Soil Sci. 2019, 66, 182–192. [Google Scholar] [CrossRef]
- Kumar, A.; Dames, J.F.; Gupta, A.; Sharma, S.; Gilbert, J.A.; Ahmad, P. Current developments in arbuscular mycorrhizal fungi research and its role in salinity stress alleviation: A biotechnological perspective. Crit. Rev. Biotechnol. 2015, 35, 461–474. [Google Scholar] [CrossRef] [PubMed]
- Parijar, M.; Rakshit, A. Arbuscular mycorrhiza: A versatile component for alleviation of salt stress. Nat. Environ. Pollut. Technol. 2016, 15, 417–428. [Google Scholar]
- Hashem, A.; Abd_Allah, E.F.; Alqarawi, A.A.; Egamberdieva, D. Arbuscular Mycorrhizal Fungi and Plant Stress Tolerance. In Plant Microbiome: Stress Response; Egamberdieva, D., Ahmad, P., Eds.; Springer Nature: Singapore, 2018; pp. 81–104. [Google Scholar]
- Evelin, H.; Devi, T.S.; Gupta, S.; Kapoor, R. Mitigation of salinity stress in plants by arbuscular mycorrhizal symbiosis: Current understanding and new challenges. Front. Plant Sci. 2019, 10, 470. [Google Scholar] [CrossRef] [PubMed]
- Ahammed, G.J.; Hajiboland, R. 2024. Introduction to arbuscular mycorrhizal fungi and higher plant symbiosis: Characteristic features, functions, and applications. In Arbuscular Mycorrhizal Fungi and Higher Plants; Ahammed, G.J., Hajiboland, R., Eds.; Springer Nature: Singapore, 2024; pp. 1–18. [Google Scholar]
- Zou, Y.-N.; Srivastava, A.K.; Ni, Q.-D.; Wu, Q.-S. Disruption of mycorrhizal extraradical mycelium and changes in leaf water status and soil aggregate stability in rootbox-grown trifoliate orange. Front. Microbiol. 2015, 6, 203. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.C.; Wang, P.; Wu, Q.H.; Zou, Y.N.; Bao, Q.; Wu, Q.S. Arbuscular mycorrhizas improve plant growth and soil structure in trifoliate orange under salt stress. Archi. Agron. Soil Sci. 2017, 63, 491–500. [Google Scholar] [CrossRef]
- Cheng, H.Q.; Ding, Y.E.; Shu, B.; Zou, Y.N.; Wu, Q.S.; Kuča, K. Plant aquaporin responses to mycorrhizal symbiosis under abiotic stress. Intl. J. Agric. Biol. 2020, 23, 786–794. [Google Scholar]
- Greenway, H.; Munns, R. Mechanisms of salt tolerance on nonhalophytes. Ann. Rev. Plant Physiol. 1980, 31, 149–190. [Google Scholar] [CrossRef]
- Rabie, G.H.; Almadini, A.M. Role of bioinoculants in development of salt-tolerance of Vicia faba plants under salinity stress. Afr. J. Biotechnol. 2005, 4, 210–222. [Google Scholar]
- Jahromi, F.; Aroca, R.; Porcel, R.; Ruiz-Lozano, J.M. Influence of salinity on the in vitro development of Glomus intraradices and on the in vivo physiological and molecular responses of mycorrhizal lettuce plants. Microb. Ecol. 2008, 55, 45–53. [Google Scholar] [CrossRef]
- Hashem, A.; Abd_Allah, E.F.; Egamberdieva, D.; Ahmad, P. Effect of AM fungi on growth, physio-biochemical attributes, lipid peroxidation, antioxidant enzymes and plant growth regulators in Lycopersicon esculentum mill. Subjected to different concentration of NaCl. Pak. J. Bot. 2015, 47, 327–340. [Google Scholar]
- Scagel, C.F.; Bryla, D.R. Salt exclusion and mycorrhizal symbiosis increase tolerance to NaCl and CaCl2 salinity in ‘Siam Queen’ basil. Hortic. Sci. 2017, 52, 278–287. [Google Scholar] [CrossRef]
- Hammer, E.C.; Nasr, H.; Pallon, J.; Olsson, P.A.; Wallander, H. Elemental composition of arbuscular mycorrhizal fungi at high salinity. Mycorrhiza 2011, 21, 117–129. [Google Scholar] [CrossRef] [PubMed]
- Nieves, M.; Ruiz, D.; Cerdá, A. Influence of Rootstock-Scion Combination in Lemon Trees Salt Tolerance. In Proceedings of the International Society of Citriculture, Acireale, Italy, 8–13 March 1992; pp. 387–390. [Google Scholar]
- Ruiz-Lozano, J.M.; Porcel, R.; Azcón, C.; Aroca, R. Regulation by arbuscular mycorrhizae of the integrated physiological response to salinity in plants: New challenges in physiological and molecular studies. J. Exp. Bot. 2012, 63, 4033–4044. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.Y.; Zou, Y.N.; Zhang, D.J.; Shu, B.; Wu, Q.S. Mycorrhizae and tolerance of abiotic stress in citrus plants. In Biofertilizers for Sustainable Agriculture and Environment; Giri, B., Prasad, R., Wu, Q.S., Varma, A., Eds.; Springer Nature: Cham, Switzerland, 2019; pp. 465–488. [Google Scholar]
- Evelin, H.; Giri, B.; Kapoor, R. Contribution of Glomus intraradices inoculation to nutrient acquisition and mitigation of ionic imbalance in NaCl-stressed Trigonella foenum-graecum. Mycorrhiza 2012, 22, 203–217. [Google Scholar] [CrossRef]
- Giri, B.; Mukerji, K.G. Mycorrhizal inoculant alleviates salt stress in Sesbania aegyptiaca and Sesbania grandiflora under field conditions: Evidence for reduced sodium and improved magnesium uptake. Mycorrhiza 2004, 14, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Grattan, S.R.; Grieve, C.M. Mineral element acquisition and growth response of plants grown in saline environments. Agric. Ecosyst. Environ. 1992, 38, 275–300. [Google Scholar] [CrossRef]
- Linderman, R.G. Vesicular arbuscular mycorrhizae and soil microbial interaction. In Mycorrhizae in Sustainable Agriculture; Bethlenfalvay, G.J., Linderman, R.G., Eds.; ASA Special Publications: Madison, WI, USA, 1992; pp. 45–70. [Google Scholar]
- Ortas, I.; Ortakçi, D.; Kaya, Z.; Çinar, A.; Önelge, N. Mycorrhizal dependence of sour orange in relation to phosphorus and zinc nutrition. J. Plant Nutr. 2002, 24, 1263–1279. [Google Scholar] [CrossRef]
- Kothari, S.K.; Marschner, H.; Romheld, V. Contribution of the VA mycorrhizal hyphae in acquisition of phosphorus and zinc by maize grown in a calcareous soil. Plant Soil. 1991, 131, 177–185. [Google Scholar] [CrossRef]
- Pacovsky, R.S. Micronutrient uptake and distribution in mycorrhizal or phosphorus fertilized soybeans. Plant Soil. 1986, 95, 379–388. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: London, UK, 2008; p. 815. [Google Scholar]
- Al-Karaki, G.N. Growth of mycorrhizal tomato and mineral acquisition under salt stress. Mycorrhiza 2000, 10, 51–54. [Google Scholar] [CrossRef]
- Kumar, A.; Sharma, S.; Mishra, S. Influence of arbuscular mycorrhizal (AM) fungi and salinity on seedlings growth, solute accumulation, and mycorrhizal dependency of Jatropha curcas L. J. Plant Growth Regul. 2010, 29, 297–306. [Google Scholar] [CrossRef]
- Navarro, J.M.; Morte, A. Arbuscular mycorrhizal fungi as biofertilizers to increase the plant quality of Sour-Orange seedlings. Agronomy 2024, 14, 230. [Google Scholar] [CrossRef]
- Sivrastava, A.K.; Singh, S. Leaf and soil nutrient guide in citrus. A review. Agric. Rev. 2004, 25, 235–251. [Google Scholar]
- Navarro, J.M.; Morte, A. Mycorrhizal effectiveness in Citrus macrophylla at low phosphorus fertilization. J. Plant Physiol. 2019, 232, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhang, H.Q.; Zhang, X.L.; Tang, M. Arbuscular mycorrhizal symbiosis alleviates salt stress in black locust through improved photosynthesis, water status, and K+/Na+ homeostasis. Front. Plant Sci. 2017, 8, 1739–1753. [Google Scholar] [CrossRef] [PubMed]
- van der Heijden, M.G.A.; Horton, T.R. Socialism in soil? The importance of mycorrhizal fungal networks for facilitation in natural ecosystems. J. Ecol. 2009, 97, 1139–1150. [Google Scholar] [CrossRef]
- Hoeksema, J.D.; Chaudhary, V.B.; Gehring, C.A.; Johnson, N.C.; Karst, J.; Koide, R.T.; Pringle, A.; Zabinski, C.; Bever, J.D.; Moore, J.C.; et al. A meta-analysis of context-dependency in plant response to inoculation with mycorrhizal fungi. Ecol. Lett. 2010, 13, 394–407. [Google Scholar] [CrossRef]
- Werner, G.D.A.; Kiers, E.T. Partner selection in the mycorrhizal mutualism. New Phytol. 2015, 205, 1437–1442. [Google Scholar] [CrossRef]
- Jin, L.; Wang, Q.; Wang, Q.; Wang, X.; Gange, A.C. Mycorrizal-induced growth depression in plants. Symbiosis 2017, 72, 81–88. [Google Scholar] [CrossRef]
- Li, H.Y.; Smith, S.E.; Holloway, R.E.; Zhu, Y.G.; Smith, F.A. Arbuscular mycorrhizal fungi contribute to phosphorus uptake by wheat grown in a phosphorus-fixing soil even in the absence of positive growth responses. New Phytol. 2006, 172, 536–543. [Google Scholar] [CrossRef] [PubMed]
- Grace, E.J.; Cotsaftis, O.; Tester, M.; Smith, F.A.; Smith, S.E. Arbuscular mycorrhizal inhibition of growth in barley cannot be attributed to extent of colonization, fungal phosphorus uptake or effects on expression of plant phosphate transporter genes. New Phytol. 2009, 181, 938–949. [Google Scholar] [CrossRef] [PubMed]
- Smith, F.A.; Grace, E.J.; Smith, S.E. More than a carbon economy: Nutrient trade and ecological sustainability in facultative arbuscular mycorrhizal symbioses. New Phytol. 2009, 182, 347–358. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Raihan, M.R.H.; Masud, A.A.C.; Rahman, K.; Nowroz, F.; Rahman, M.; Nahar, K.; Fujita, M. Regulation of reactive oxygen species and antioxidant defence in plants under salinity. Int. J. Mol. Sci. 2021, 22, 9326. [Google Scholar] [CrossRef] [PubMed]
- Alqarawi, A.A.; Abd_Allah, E.F.; Hashem, A. Alleviation of salt-induced adverse impact via mycorrhizal fungi in Ephedra aphylla Forssk. J. Plant Interact. 2014, 9, 802–810. [Google Scholar] [CrossRef]
- Yang, Y.; Tang, M.; Sulpice, R.; Chen, H.; Tian, S.; Ban, Y. Arbuscular mycorrhizal fungi alter fractal dimension characteristics of Robinia pseudoacacia L seedlings through regulating plant growth, leaf water status, photosynthesis and nutrient concentration under drought stress. J. Plant Growth Regul. 2014, 3, 612–625. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).