Abstract
The storage roots of orange-fleshed sweet potato contain high levels of polyphenols and carotenoids. Little information is available on changes in the content and composition of these secondary metabolites during the growth stages of the root system. We investigated changes in carotenoids, polyphenols, and anthocyanins in the root system of the orange-fleshed sweet potato ‘Tamaakane’ from the post-planting stage. Carotenoids and polyphenols accumulated immediately after root-system formation at 15 days after transplanting (DAT). The levels of carotenoids increased by 45 DAT and decreased once the storage root enlargement began, but increased rapidly thereafter. After 90 DAT, β-carotene accounted for >90% of the total carotenoids, and the frequency remained stable until maturity. Total polyphenol content increased significantly towards 45 DAT and then gradually decreased as the storage roots began to thicken. No anthocyanin was detected in the root systems of ‘Tamaakane’ at any stage. The content and composition of these secondary metabolites are discussed in terms of agronomic aspects, as they might contribute to a strategy to protect storage roots while engaging with each other against biotic or abiotic stresses at the growth stage of the root system.
1. Introduction
Sweet potato (Ipomoea batatas (L.) Lam.) is a herbaceous dicotyledonous plant of the Convolvulaceae family, widely grown in subtropical and temperate regions. Mature sweet potatoes have fibrous, thickly pigmented tuberous roots. Fibrous roots develop into storage roots that accumulate starch and form pigments such as carotenoids and anthocyanins through very complex physiological and genetic processes. Recently, the molecular mechanisms of storage root formation have been clarified [1,2,3]. Understanding these molecular mechanisms as well as those of anthocyanin biosynthesis will facilitate plant breeding and/or engineering that promotes the production of anthocyanins, an important group of antioxidants available from pigmented foods [4].
Sweet potato storage roots are a source of energy because they contain carbohydrates, vitamins, and minerals [5]. Another characteristic of sweet potato storage roots is their polyphenol content and composition: one cultivar was found to contain 780 mg·100 g−1 of total polyphenols [6], and another study identified caffeoylquinic acid as the main polyphenol [7]. Polyphenols are generally highly correlated with antioxidant activity [8,9,10] and considered to be involved in the body’s defenses against UV light and pathogens [11]. For instance, we previously revealed how the polyphenol content and its composition in purple-fleshed sweet potatoes vary with growth stage [6,12], increasing rapidly just before the storage root enlarges. This phenomenon may also be a protective mechanism to protect storage roots from biotic or abiotic stresses during the critical transition of storage root enlargement.
Yellow- and orange-fleshed sweet potatoes contain unique carotenoids in addition to polyphenols, and carotenoid contents in mature storage roots of yellow- and orange-fleshed sweet potato cultivars have been investigated [13,14,15,16,17]. The carotenoids in the storage root of sweet potatoes include β-carotene, β-crypthoxanthin, zeaxanthin, violaxanthin, and other unknown carotenoids. More specifically, yellow-fleshed cultivars contain β-carotene 5,8;5′,8′-diepoxide at a rate of 32–51% and β-cryptoxanthin 5,8-epoxide at 11–30%, while orange-fleshed cultivars have a higher percentage of β-carotene at 80–92% [7]. Carotenoid content in plants is determined by carotenoid biosynthesis, and the regulation of this pathway has been widely studied [18]. Numerous carotenoid biosynthesis genes in sweet potato such as phytoene synthase and lycopene epsilon cyclase have been isolated and characterized [19,20,21]. Recently, Ke et al. grew transgenic sweet potato plants (RLE plants) with down-regulated expression of IbLCY-ε and found they had high levels of total carotenoids and β-carotene and tolerated oxidative stress, salt stress, and drought stress surprisingly well [22].
The total amounts and compositions of carotenoids and polyphenols in sweet potatoes are valuable information for managing cultivation methods for resistance to biotic or abiotic stresses and for breeding. There is little information on carotenoids and polyphenols during the growth stages of storage roots of orange-fleshed cultivars. Therefore, in the present study, we investigated the effect of the growth stage of the root system on carotenoid and polyphenol contents and compositions using ‘Tamaakane’, an orange-fleshed cultivar known to have high carotenoid content.
2. Materials and Methods
2.1. Plant Materials
The orange-fleshed sweet potato cultivar ‘Tamaakane’ was used in this experiment. This cultivar is a sweet potato developed by the Kyushu Okinawa Agricultural Research Center and was first released in 2009. This is the first exclusive cultivar with orange flesh for use as a raw planting material in producing shochu, an alcoholic beverage popular in Japan. Cultivar characteristics, including nutritional information, for ‘Tamaakane’ are shown [23]. The yellow-fleshed cultivar ‘Kokei No.14’ was used as a control. Cultivar characteristics, including nutritional information, for ‘Kokei No.14’ are shown [24].
Stem cuttings of each cultivar were transplanted into containers (64 × 23 × 18.5 cm) filled with commercially available culture soil (NAFCO Co., Ltd., Fukuoka, Japan) at 29 May 2021 and grown at the experimental farm at the Faculty of Agriculture, University of Miyazaki (31°49′39.8″ N 131°24′42.4″ E). The sweet potatoes were grown in containers to enable harvesting without damage to the initial thin roots. To investigate the changes in carotenoids, polyphenols, and anthocyanins during root growth, the root system up to 60 days was harvested from containers at four developmental stages (15, 30, 45, and 60 days after transplanting (DAT)) (Figure 1). Root systems for investigation included both adventitious roots and lateral roots. The number of samples in this study was 3, with a minimum of 2 g of root system per sample. In cases where the amount of the root system was insufficient, the nurseries were collected from multiple plants. In addition, stem cuttings of each cultivar were grown in a research plot in Miyazaki, Miyazaki Prefecture, near the experimental field (31°49′55.6″ N 131°18′13.2″ E). To investigate the changes in carotenoids, polyphenols, and anthocyanins during root growth, the root system was harvested from conventionally grown fields at three developmental stages (90, 120, and 150 DAT) (Figure 1). The weather conditions in Miyazaki City, the test site, during this period were as follows: The average monthly temperatures were 23.2 °C in June, 27 °C in August, and 20.8 °C in October. The monthly precipitation was 368.0 mm in June, 465.5 mm in August, and 120.5 mm in October, and the monthly sunshine hours were 107.8 h in June, 168.1 h in August, and 213.6 h in October [25].

Figure 1.
Root system and sections of ‘Tamaakane’ at different growth stages. DAT: days after transplant.
Harvested samples were freeze-dried, ground into powder, and stored at −30 °C until analysis for carotenoids, polyphenols, and anthocyanins. All root systems were frozen to a temperature of −30 °C and then freeze-dried in an FDU-1100 lyophilizer and a DRC-1000 chamber (Tokyo Rikakikai, Tokyo, Japan). The freeze-dried samples were ground into powder and stored at −30 °C until use in the experiments.
2.2. Determination of Total Carotenoid Content
Carotenoids were extracted from the root system and the total carotenoid content was determined according to the method of Ishiguro et al. [13].
We dissolved 0.1 g of freeze-dried powder of the root systems (15–150 DAT) in 0.6 mL of acetone and mixed the suspension with a vortex mixer. The mixture was then centrifuged at 1500× g for 10 min. After the removal of a 0.2 mL aliquot of supernatant, 0.6 mL of acetone was added to the remainder. After re-extraction, 0.6 mL of the supernatant was collected and mixed with the first extract (0.8 mL total). Extracts were diluted accordingly and used for the assay. Total carotenoid content was detected in the wavelength range of 360–500 nm spectrophotometrically with a UV-mini 1240 (Shimadzu, Kyoto, Japan). The total carotenoid content was calculated using the extinction coefficient of E = 2500 at λ max. Measurements were replicated three times, and results are expressed as mg·100 g−1 FW.
2.3. Compositional Analysis of Carotenoids
To investigate the carotenoids in the root system, the carotenoid composition was measured using high-performance liquid chromatography (HPLC) according to the method described by Ishiguro et al. [13] with some modifications. Compositional analysis could not be performed for 15 and 30 DAT roots due to their low volume. For the rest, 0.5 g of the freeze-dried powder of root systems (45–150 DAT) was dissolved in 3 mL of acetone, and the suspension was mixed by a vortex mixer. Then, the mixture was centrifuged at 1500× g for 10 min. A 1 mL aliquot of the supernatant was removed and then 3 mL of acetone was added again. After re-extraction, 3 mL of the supernatant was collected and combined with the first extract (total 4 mL). The extract was then evaporated under nitrogen (60 °C) and the residue was redissolved in 1 mL of tetrahydrofuran containing 0.1% butylated hydroxytoluene (BHT) and filtered through a 0.2 μm membrane filter (Sartorius, Göttingen, Germany).
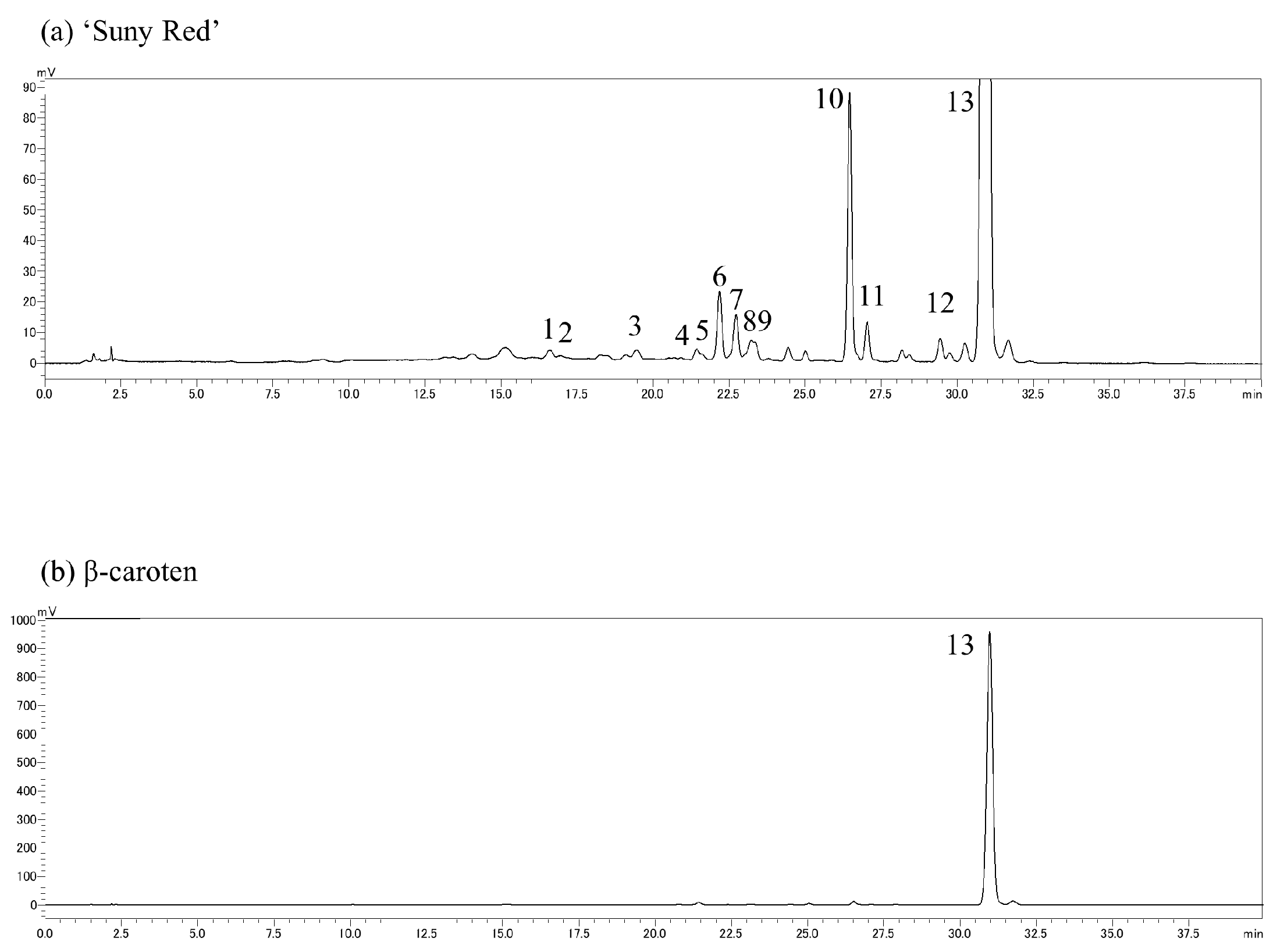
The extracts were analyzed by reverse-phase HPLC using the Prominence (Shimadzu, Kyoto, Japan) and LC solution systems (Shimadzu, Kyoto, Japan) with a Synergi Hydro-RP column (Particle size; 4 µm, inside diameter; 3 mm, length; 250 mm) (Phenomenex, Torrance, CA, USA). Solvent A was acetonitrile/chloroform/methanol/trimethylamine at ratios of 90:10:0.05, and solvent B was acetonitrile/ultrapure water/trimethylamine at 75:15:10:0.05; other chromatographic conditions were as follows: column temperature, 35 °C; detection at 430 nm; flow rate, 0.6 mL·100 g−1 FW. The binary gradient was as follows: 0–100% B (0–20 min), 100% B (20–40 min), 100–0% B (40–41 min). The carotenoid composition was estimated using the extract from ‘Sunny Red’ as a standard (analyzed by Ishiguro et al. [13]) with a real reagent (β-carotene), and their chromatograms are shown in Figure 2. The results are expressed as percentages, setting the total carotenoid content equal to 100. The measurements were replicated three times.
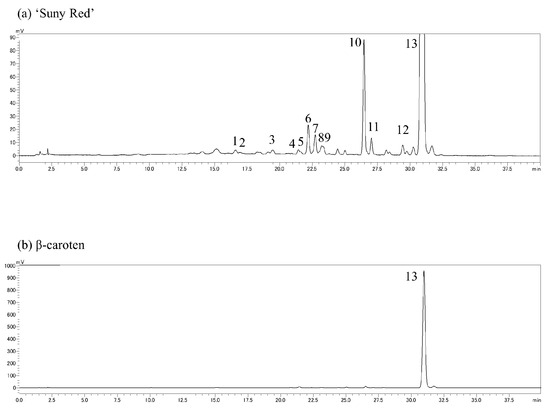
Figure 2.
Chromatograms of (a) carotenoids extracted from ‘Suny Red’ and (b) real reagent β-carotene. (1) Unknown, (2) ipomoeaxanthin C1, (3) ipomoeaxanthin C2, (4) β-cryptoxanthin 5,8;5′,8′-diepoxide, (5) β-cryptoxanthin 5,8-epoxide, (6) unknown, (7) β-carotene 5,8;5′,8′-diepoxide(cis), (8) β-carotene 5,8;5′,8′-diepoxide, (9) β-carotene 5,8;5′,8′-diepoxide, (10) unknown, (11) β-carotene 5,8-epoxide, (12) unknown, and (13) β-carotene.
2.4. Compositional Analysis of Polyphenols
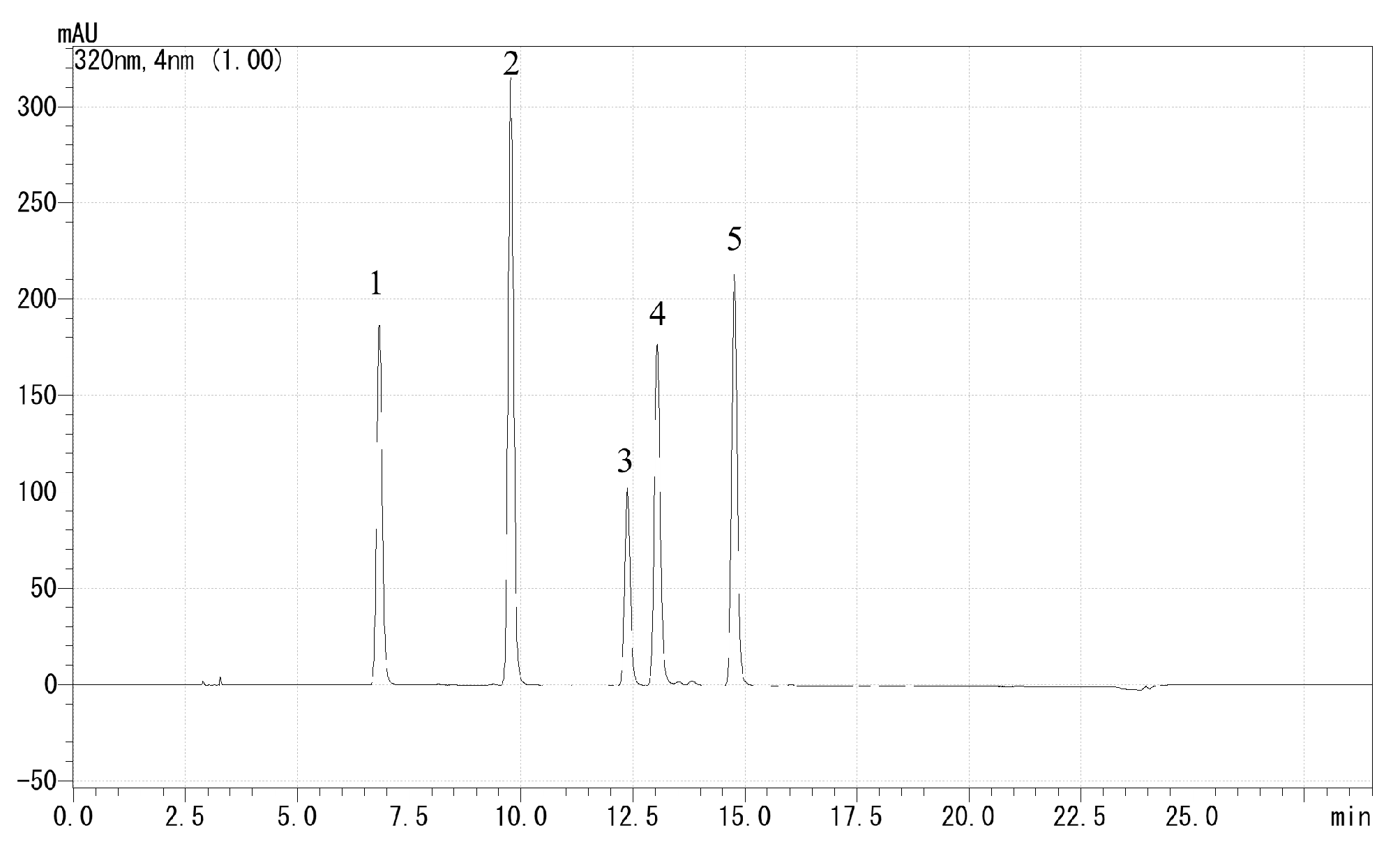
HPLC was used to determine the total polyphenol content and composition, and individual polyphenols in the samples were separated and quantified. First, we dissolved 0.02 g of freeze-dried root powder into 5 mL of 80% methanol and analyzed it through a 0.2 µm membrane filter. The extracts were analyzed by reverse-phase HPLC using the UF-Amino (Shimadzu, Kyoto, Japan) and LC solution systems (Shimadzu, Kyoto, Japan) with an ODS-3 column (particle size, 5 µm; inside diameter, 4.6 mm; length, 250 mm) (GL Sciences, Tokyo, Japan). Solvent A was 100% ethanol and solvent B was 20 mM KH2PO4 (pH 2.4); other chromatographic conditions were as follows: column temperature, 40 °C; detection at 320 nm; flow rate, 1.0 mL·100 g−1 FW. The binary gradient was as follows: 85–68% B (0–12 min), 68% B (12–15 min), 50–55% B (15–20 min), 85% B (20–29 min). Retention times and spectra were compared with pure standards of chlorogenic acid, caffeic acid, 3,4-dicaffeoylquinic, 3,5-dicaffeoylquinic acid, and 4,5-dicaffeoylquinic acid, and the chromatogram of pure standards is shown in Figure 3. To investigate the content of each individual polyphenol, four concentrations of pure standards were prepared at 5, 10, 20, and 50 ppm. The sum of the five polyphenols was used as the total polyphenol content and expressed as mg·100 g−1 FW; the results of the composition are expressed as percentages. Measurements were replicated three times.
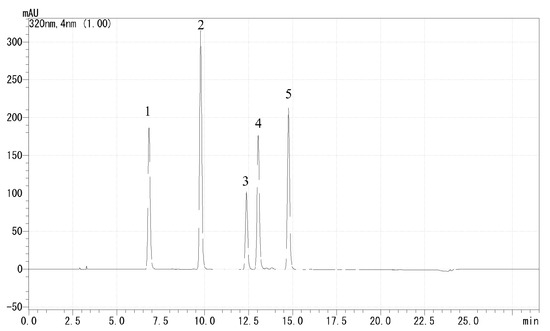
Figure 3.
Chromatogram of polyphenol pure standards: (1) chlorogenic acid, (2) caffeic acid, (3) 3,4-dicaffeoylquinic acid, (4) 3,5-dicaffeoylquinic acid, and (5) 4,5-dicaffeoylquinic acid.
2.5. Compositional Analysis of Anthocyanin
Total anthocyanins were measured using a micro plate reader according to the pH differential method [26]. A 0.04 g sample of lyophilized powder was dissolved in 2 mL of 100% methanol and sonicated for 15 min. Finally, the extract was passed through a 0.2 μm membrane filter and analyzed for total anthocyanins.
Briefly, absorbance of the extract was measured at 510 and 700 nm in buffers at pH 1.0 (hydrochloric acid–potassium chloride, 0.2 N) and 4.5 (acetate acid–sodium acetate, 1 M), respectively. Anthocyanin content was calculated using a molar extinction coefficient of 29,600 (Cy-glu; cyanidin-3-glucoside) and absorbance of A = [(A510 − A700) pH1.0 − (A510 − A700) pH4.5]. The results are expressed as mg·100 g−1 FW. Measurements were replicated three times.
2.6. Statistical Analysis
All results were evaluated for statistical significance using Tukey’s multiple range test with Excel (Microsoft Office Home and Business 2016) and Excel statistics used for statistical analysis. Differences were considered significant at the 5% level as determined by Tukey’s multiple range test.
3. Results
3.1. Total Carotenoid Contents
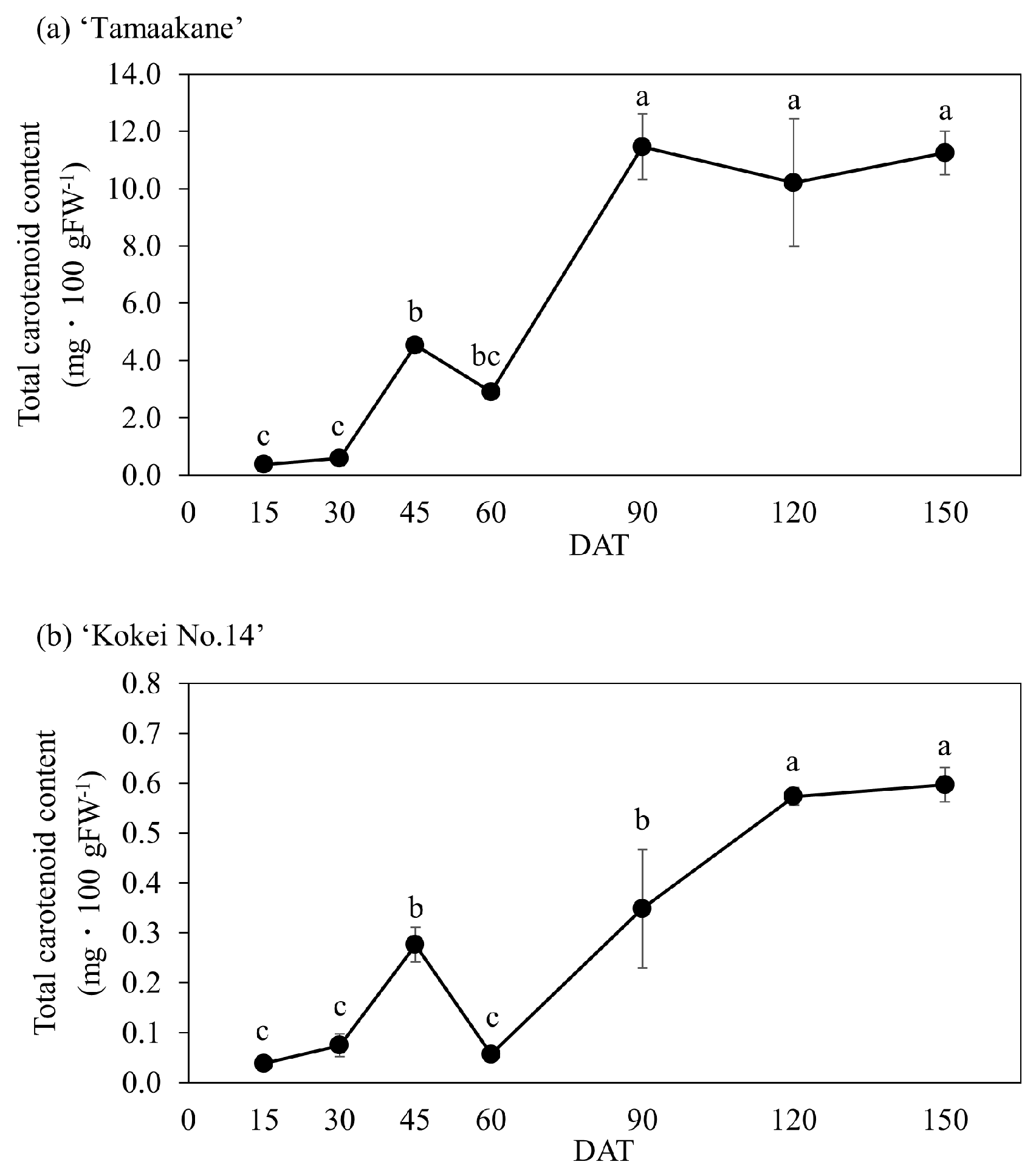
The total carotenoid content differed between the orange-fleshed cultivar ‘Tamaakane’ and the yellow-fleshed cultivar ‘Kokei No.14’ at all growth stages. Parts of the root system of ‘Tamaakane’ had already accumulated carotenoids at 15 DAT (Figure 1). The total carotenoid content at 15 DAT was 0.37 mg·100 g−1 FW, and a clear orange color was observed inside the storage roots at 30 DAT. Thereafter, the total carotenoid content decreased temporarily to 2.91 mg·100 g−1 FW at 60 DAT when the storage roots began to enlarge, but it then rapidly increased to 11.46 mg·100 g−1 FW at 90 DAT (Figure 4a). The content level then remained flat from 90 to 150 DAT (Figure 4a).
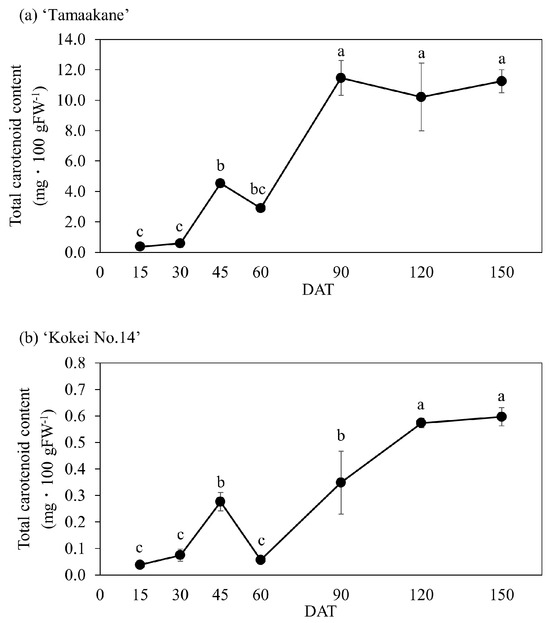
Figure 4.
Change in the total carotenoid content of the root system of (a) ‘Tamaakane’ and (b) ‘Kokei No.14’. Each data represent mean ± S.D. (n = 3). Different letters represent significant differences at 5% level as determined by Tukey’s multiple range test. DAT: days after transplant.
In contrast, carotenoids in the control cultivar ‘Kokei No.14’ were detected at 15 DAT, as in ‘Tamaakane’, but in very small amounts. The total carotenoid content of ‘Kokei No.14’ was 0.04 mg·100 g−1 FW at 15 DAT and it increased up to 45 DAT, but like the data for ‘Tamaakane’ it had decreased at 60 DAT when the storage roots began to enlarge. Thereafter, the content rapidly increased to 120 DAT and was maintained at that level to 150 DAT (0.60 mg·100 g−1 FW) (Figure 4b). As noted above, our observations confirmed that carotenoids had already accumulated at 15 DAT in both cultivars and rapidly increased after 60 DAT.
3.2. Carotenoid Composition
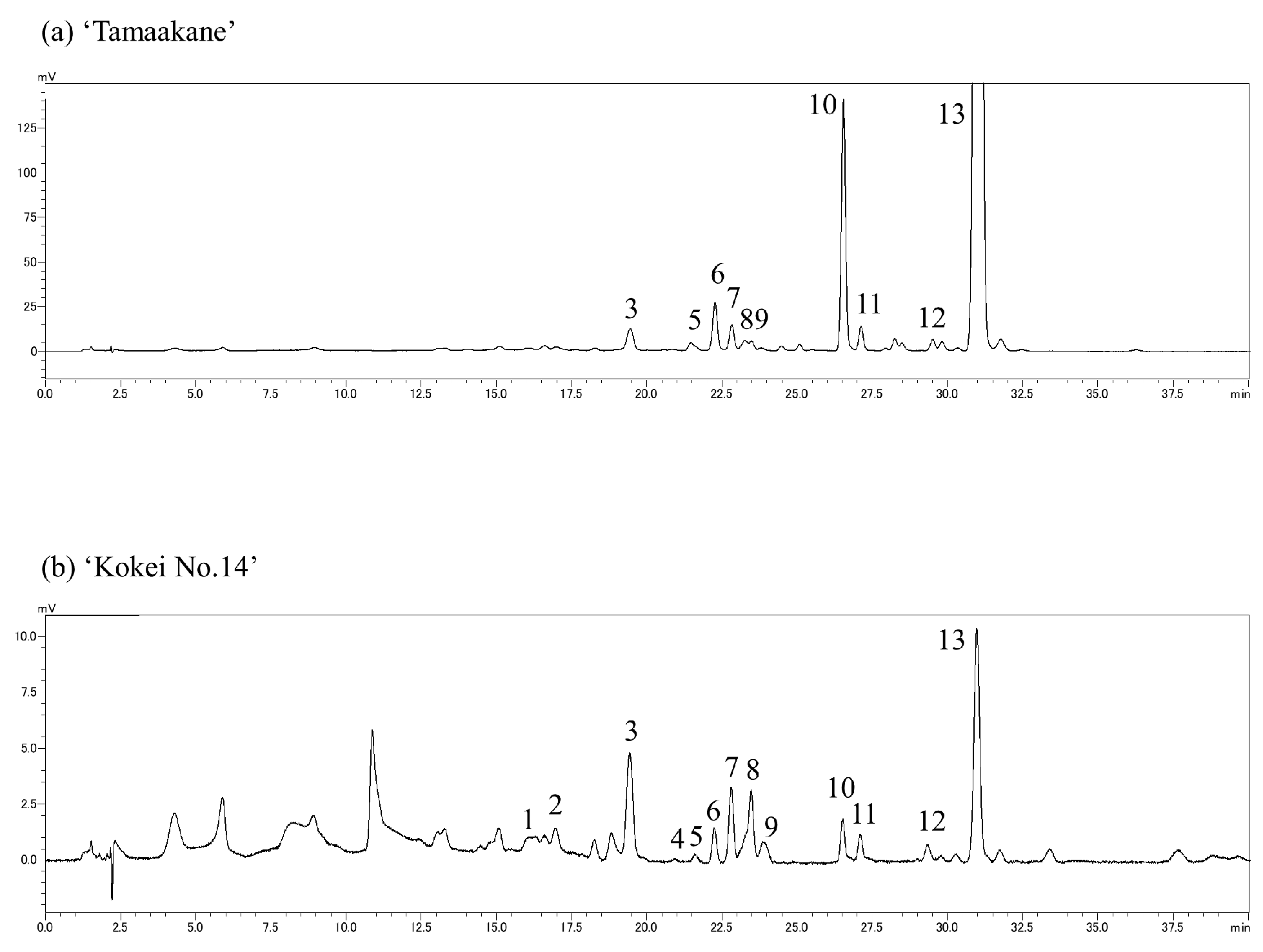
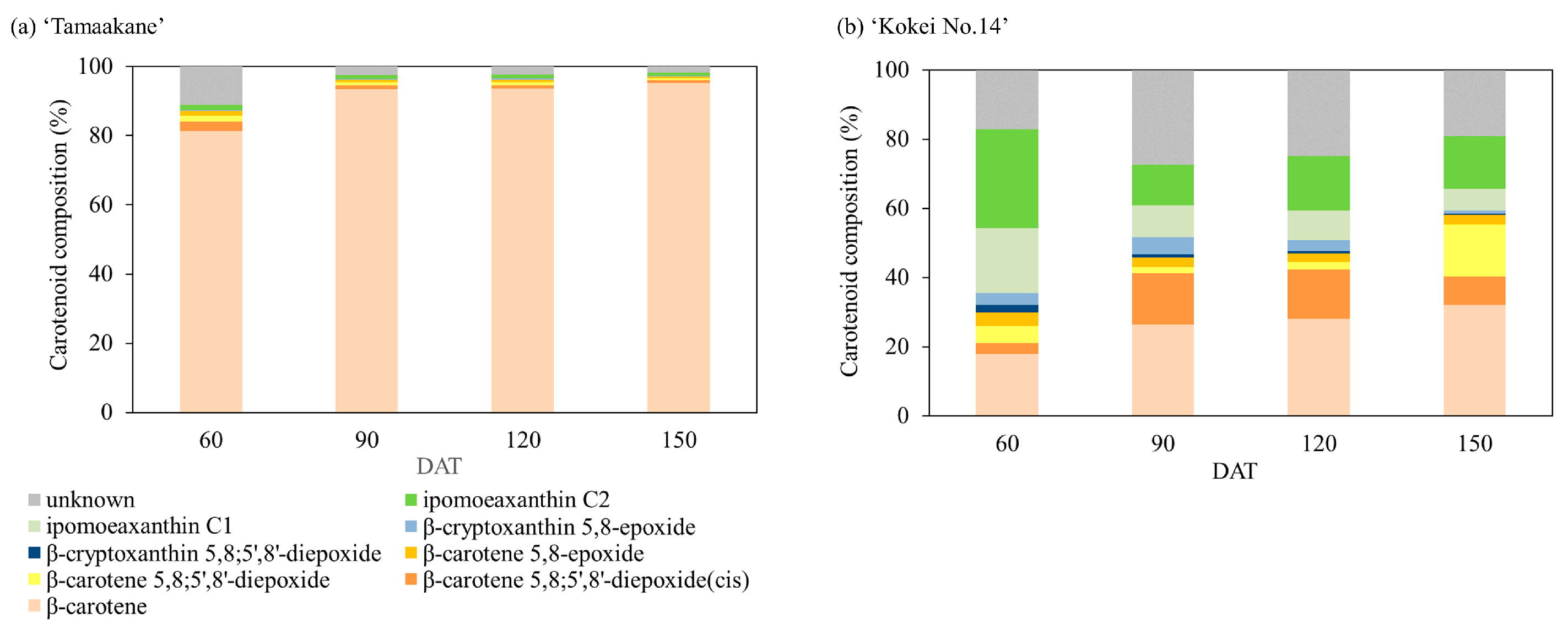
We evaluated the carotenoid compositions of the storage root of the ‘Tamaakane’ and ‘Kokei No.14’ cultivars and identified 10 and 13 peaks, respectively (Figure 5). These peaks were ipomeaxanthin, β-carotene, β-cryptoxanthin, and its epoxy compounds. Figure 6 depicts the frequency of these carotenoids. In the storage root of ‘Tamaakane’ at 60 DAT, β-carotene accounted for 81.4% of the total carotenoids. Small amounts of 2.7% β-carotene 5,8;5′,8′-diepoxide(cis), 1.7% β-carotene 5,8;5′,8′-diepoxide, 1.4% β-carotene 5,8-epoxide, 0.2% β-cryptoxanthin 5,8-epoxide, and 1.5% ipomoeaxanthin C2 were also observed. After 90 DAT, β-carotene accounted for >90% of the total carotenoids, and the frequency remained stable until maturity. Other similar carotenoids were also detected in small amounts after 90 DAT.
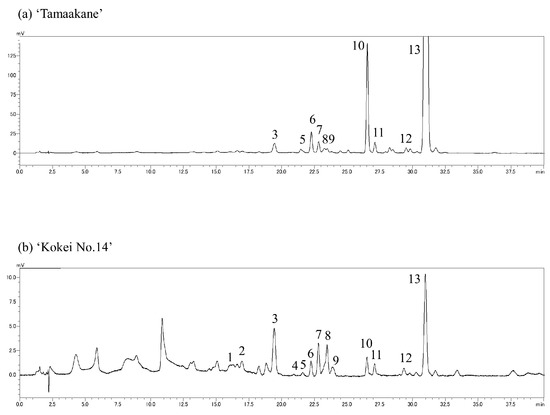
Figure 5.
Chromatograms of carotenoids extracted from (a) ‘Tamaakane’ and (b) ‘Kokei No.14’: (1) unknown, (2) ipomoeaxanthin C1, (3) ipomoeaxanthin C2, (4) β-cryptoxanthin 5,8;5′,8′-diepoxide, (5) β-cryptoxanthin 5,8-epoxide, (6) unknown, (7) β-carotene 5,8;5′,8′-diepoxide(cis), (8) β-carotene 5,8;5′,8′-diepoxide, (9) β-carotene 5,8;5′,8′-diepoxide, (10) unknown, (11) β-carotene 5,8-epoxide, (12) unknown, and (13) β-carotene.
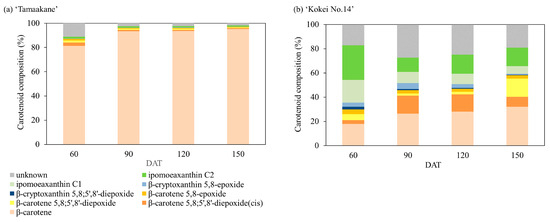
Figure 6.
Change in the percentage of carotenoids extracted from (a) ‘Tamaakane’ and (b) ‘Kokei No.14’. Each data represent mean values (n = 3). DAT: days after transplant.
3.3. Polyphenol Content and Composition
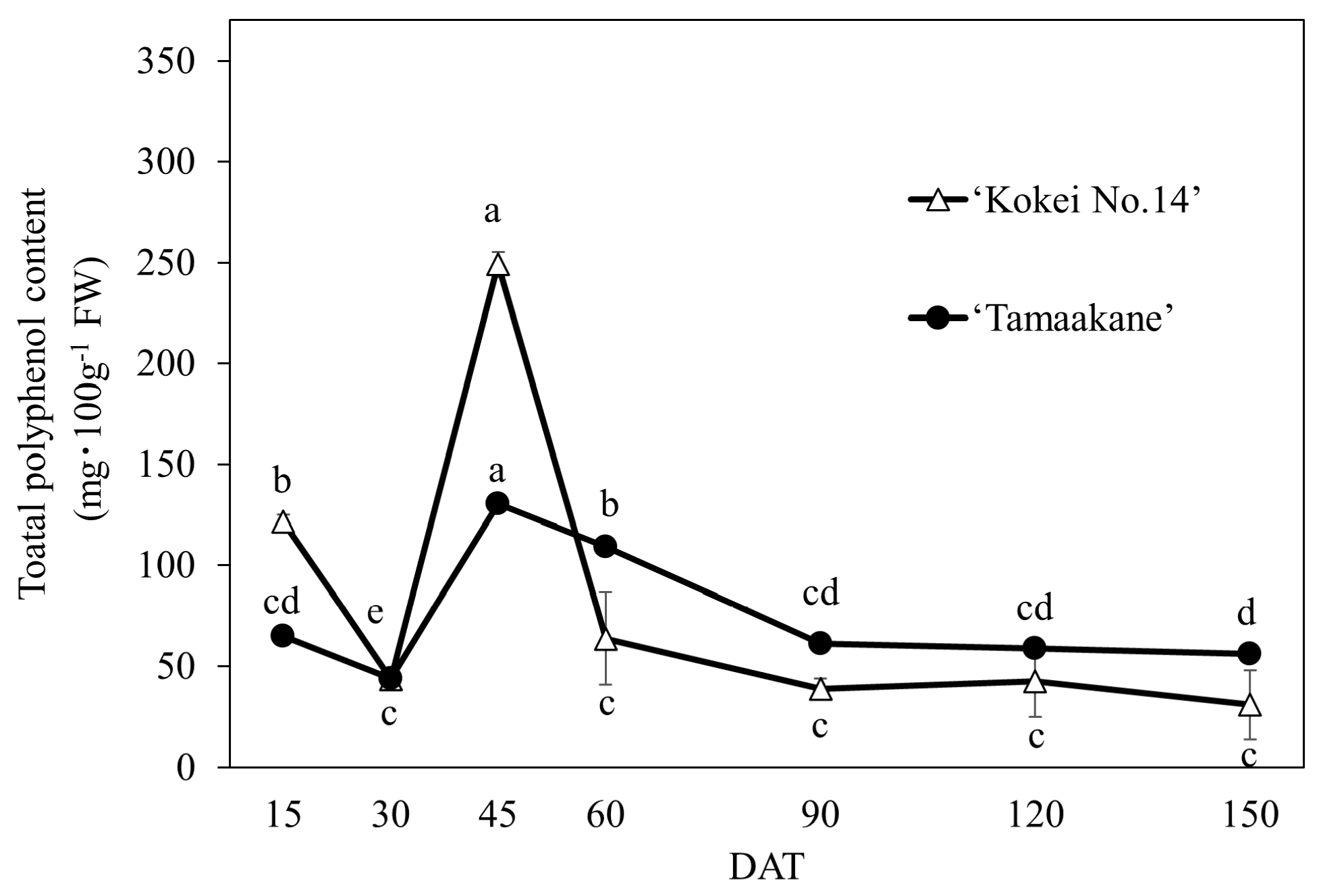
The polyphenol extracts of ‘Tamaakane’ and ‘Kokei No.14’ were analyzed by HPLC, and five different peaks were eluted (Figure 7). Chromatographic patterns did not differ by cultivar, but individual peak areas were different. The total polyphenol content calculated from the peak area is shown in Figure 8. The trends of the total polyphenol content in the root system were similar in the two cultivars, although differences in the concentration were observed at each growth stage. In the root system of ‘Tamaakane’, polyphenols were already detected at 15 DAT (64.76 mg·100 gFW−1), and although their levels had decreased significantly by 30 DAT, they rapidly increased to 130.25 mg·100 gFW−1 by 45 DAT. The content then decreased significantly as the storage roots enlarged, and after 90 DAT, the content remained stable at a low level. In contrast, polyphenols were detected at 15 DAT in the root system of ‘Kokei No.14’ as well as in ‘Tamaakane’ (121.54 mg·100 gFW−1), which rapidly increased to 249.13 mg·100 gFW−1 at 45 DAT, and then the content decreased significantly as the storage roots enlarged.
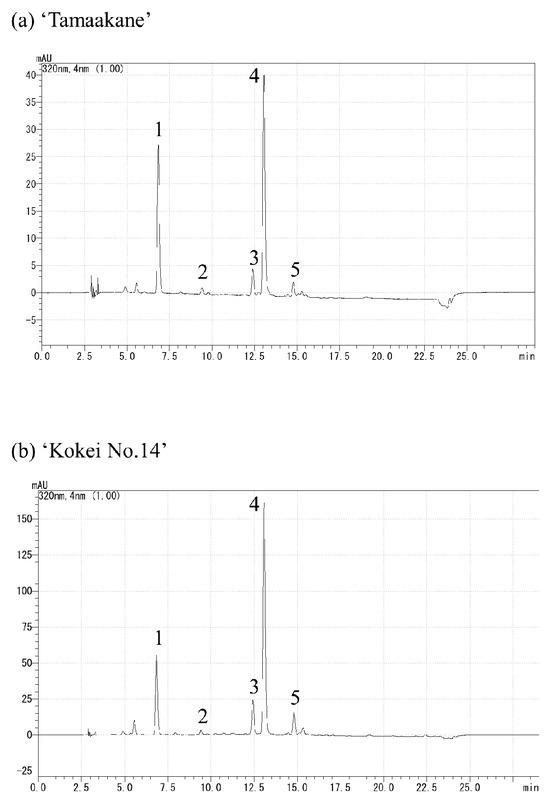
Figure 7.
Chromatograms of polyphenols extracted from (a) ‘Tamaakane’ and (b) ‘Kokei No.14’: (1) chlorogenic acid, (2) caffeic acid, (3)3,4-dicaffeoylquinic acid, (4) 3,5-dicaffeoylquinic acid, and (5) 4,5-dicaffeoylquinic acid.
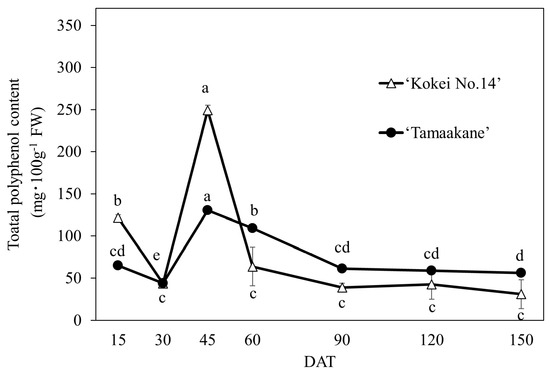
Figure 8.
Change in the total polyphenol content of the root system of ‘Tamaakane’ and ‘Kokei No.14’. Each data represent mean ± standard deviation (n = 3). Different letters represent significant differences at 5% level as determined by Tukey’s multiple range test. DAT: days after transplant.
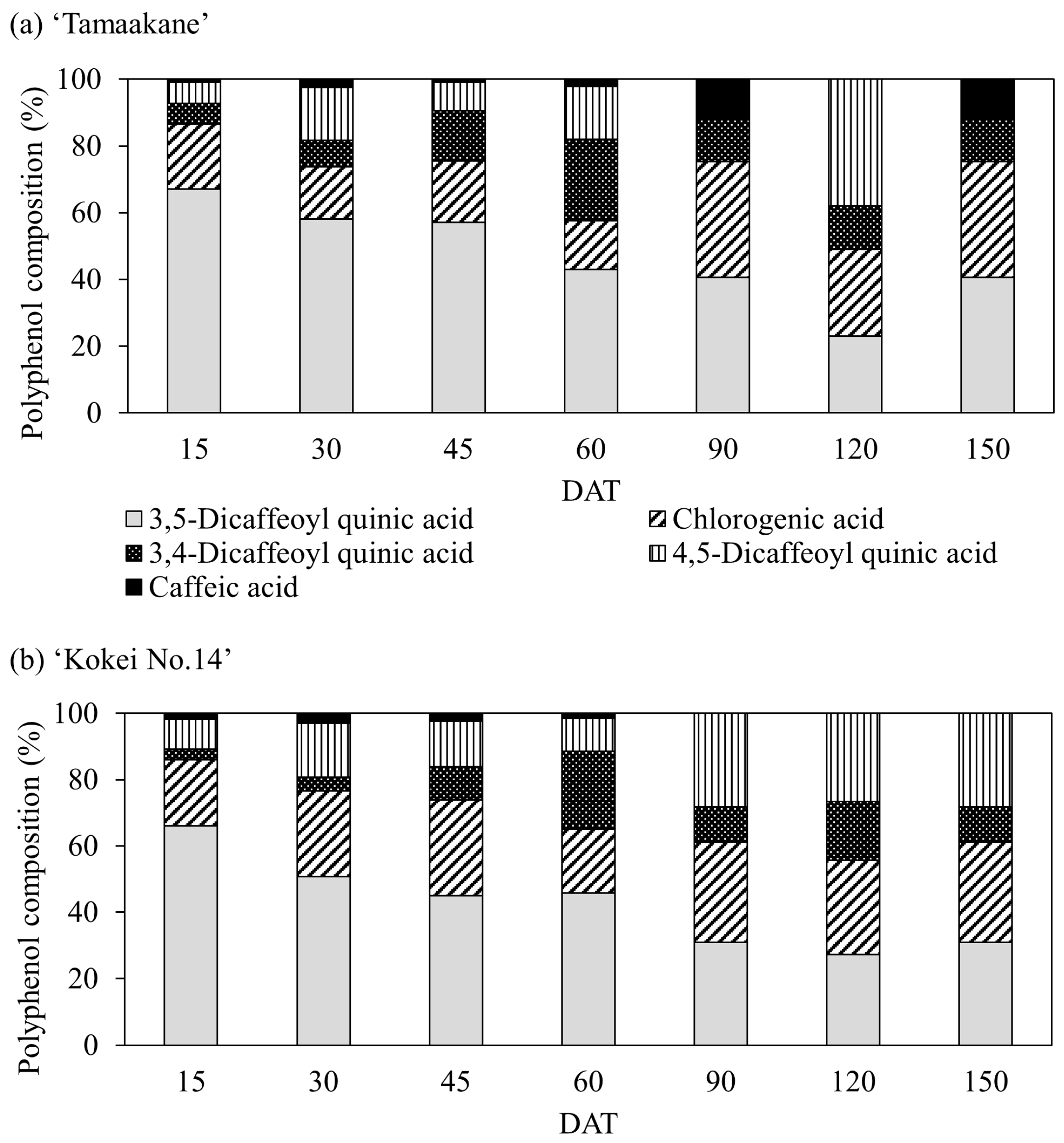
We also detected five polyphenols (3,5-dicaffeoylquinic acid, chlorogenic acid, 3,4-dicaffeoylquinic acid, 4,5-dicaffeoylquinic acid, and caffeic acid) in the root systems of all stages of both cultivars (Figure 9). In the root systems of ‘Tamaakane’ and ‘Kokei No.14’ 15 DAT, 3,5-dicaffeoylquinic acid was the most predominant polyphenol, accounting for 66.1% and 67.1% of all polyphenols, respectively. Thereafter, as the root systems of both cultivars grew, the percentage of the 3,5-dicaffeoylquinic acid content decreased, and at the time of harvest, the percentage of chlorogenic acid content tended to increase.
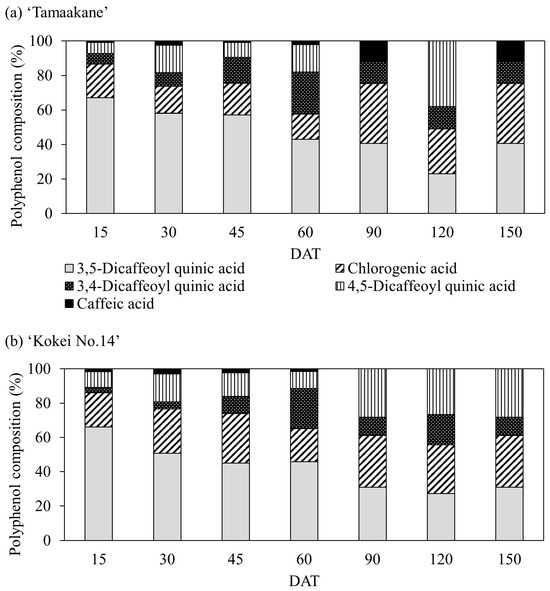
Figure 9.
Change in the percentage of polyphenols extracted from (a) ‘Tamaakane’ and (b) ‘Kokei No.14’. Each data represent mean values (n = 3). DAT: days after transplant.
3.4. Total Anthocyanin Content
No anthocyanin was detected in the root systems of ‘Tamaakane’ at any stage. In ‘Kokei No.14’, slight amounts of anthocyanin in the skin were detected at 30 DAT (0.01 mg·100 g−1 FW), 45 DAT (0.08 mg·100 g−1 FW), and 60 DAT (0.04 mg·100 g−1 FW). Anthocyanin was not detected at other stages such as 15 DAT and maturation stages.
4. Discussion
Polyphenols (including anthocyanins, flavonols, and stilbene derivatives) and carotenoids involved in the biosynthesis of abscisic acid are a class of specialized metabolites that play important physiological roles throughout the plant life cycle, including responses to biotic or abiotic stresses [27]. Polyphenols and carotenoids accumulate in response to various stresses and help plants adapt to unfavorable environments [28,29]. The concentrations of polyphenols and carotenoids in plant tissues are thus good indicators that could be used to predict the degree of plants’ biotic or abiotic stress tolerance, which varies widely among plant species under a range of external factors.
Although there is a significant amount of information regarding the accumulation of polyphenols and carotenoids during the growth of plant leaves, stems, and fruits, there is relatively little information about the accumulations of polyphenols and carotenoids in the root system. A unique accumulation pattern of polyphenols was recently observed in the sweet potato root system, which accumulates large amounts of anthocyanins [6,12]. However, we have found no published data regarding the trends of polyphenols in cultivars that specifically accumulate carotenoids. The present investigation examined the effects of polyphenols, carotenoids, and their composition on the growth stage of the root system using ‘Tamaakane’, an orange-fleshed cultivar that accumulates large amounts of β-carotene.
At 15 DAT, not only polyphenols but also carotenoids were already accumulated in the adventitious roots and lateral roots of the orange-fleshed cultivar ‘Tamaakane’, just after the root system had developed. The carotenoid content then rapidly increased up to 45 DAT along with the polyphenol domain and then decreased once the storage roots began to enlarge. At 45 DAT, when the storage roots begin to enlarge, the root system of ‘Tamaakane’ had accumulated 15 times more carotenoids than the control cultivar, ‘Kokei No.14’. The contents of polyphenols in the roots of ‘Tamaakane’, which accumulated a large amount of carotenoids, were only about half those of ‘Kokei No.14’.
Generally, from early June to early July is the rainy season in the Southern Kyushu region where this study was conducted. Then, from mid-July onward, the summer season is in full swing, and plant bodies are exposed to high temperatures (45–60 DAT; see weather data listed in Section 2). In addition, higher temperatures increase the activity of pests and diseases. Our research group reported that in several purple-fleshed cultivars, the total polyphenol content rapidly increased towards 45 DAT and decreased rapidly as storage roots began to expand [12]. From these results, we consider the transition point from pre-swelling to storage roots to be 30 DAT, that flavonoid synthesis predominated over polyphenol synthesis at the tipping point, and that the polyphenol content would become high at 45 DAT. In other words, it is presumed that the total polyphenol content of storage roots is increased and the plant body acquires resistance to various stresses before entering the critical period when they are exposed to high temperatures and drought stress and their storage roots are enlarged (around 45 DAT). Similar changes in total polyphenol content were observed in the orange-fleshed cultivar ‘Tamaakane’, which does not express anthocyanins, which were the focus of this study. This suggests that the trend in total polyphenol content of storage roots is a common feature of sweet potato. However, the marked change in total polyphenol content was not as pronounced as in the yellow-fleshed ‘Kokei No.14’, which was tested as a control.
Anthocyanins are synthesized from the general phenylpropanoid pathway by the action of metabolites that are loosely associated with the cytoplasmic surface of the endoplasmic reticulum and ultimately accumulate in the vacuole [30]. Carotenoids are synthesized in plastids located in the cytoplasm and accumulate as they are [31]. At certain developmental stages of storage roots, different secondary metabolite biosynthetic systems would be activated simultaneously, involving each other. It may be a plant strategy to ensure resistance to biotic and abiotic stresses, among others. Investigating the relationship between storage root development and the regulatory systems of secondary metabolite expression would be meaningful in elucidating plant resistance to stress.
Our analyses also revealed that in the root systems (immature storage roots) of ‘Tamaakane’ and ‘Kokei No.14’ at 60 DAT—a time point at which storage root enlargement is observed in most nurseries—the polyphenol composition was similar in the two cultivars, with 3,5-dicaffeoylquinic acid and chlorogenic acid being the main components. However, the total carotenoid content at 60 DAT differed significantly in composition as well as in its content. In particular, the ratio of β-carotene was completely different between the cultivars.
Chlorogenic acid and its derivatives, which are considered antioxidants, are known to be involved in resistance not only to biotic stresses against insects and pathogens but also to abiotic stresses such as harmful environmental factors [32,33]. It has also been reported that the increase in β-carotene, one of the carotenoids, improved strong antioxidant capacity and salt stress tolerance [18,34]. Aromatic and medicinal plants tend to produce large amounts of phenolics, carotenoids, and other secondary metabolites in response to salinity [35,36,37]. Sweet potatoes may also acquire resistance to biotic or abiotic stresses during the critical period when they are exposed to high temperatures and drought stress and their storage roots are enlarged, with the respective biosynthetic systems of polyphenols and carotenoids engaging each other.
The present study clarified for the first time the changes in total polyphenol and total carotenoid contents and their composition during the root system development of orange-fleshed sweet potato varieties. In particular, the trend in the total polyphenol content was similar to that of purple-fleshed varieties, and considering the above-cited findings, this may be one of the defense strategies against biotic and abiotic stress. Pesticide-free production as well as organic production are receiving renewed attention due to the consumer demand for safe food. Controlling polyphenol and carotenoid contents and their composition in plant bodies might protect food crops from biotic and abiotic stresses and increase the organic acreage.
Author Contributions
Conceptualization, Y.S. and H.K.; methodology, Y.S.; validation, Y.S.; formal analysis, Y.S.; investigation, Y.S.; resources, Y.S.; data curation, Y.S.; writing—original draft preparation, Y.S.; writing—review and editing, H.K., Y.N., T.H. and M.O.; visualization, Y.S.; supervision, H.K., T.H. and M.O.; project administration, H.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Data are contained within the article.
Acknowledgments
This work was supported by JST SPRING, Grant Number JPMJSP2105.
Conflicts of Interest
Author Yosuke Narasako was employed by the company Kushima AoiFarm Co., Ltd. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Suematsu, K.; Tanaka, M.; Isobe, S. Identification of a major QTL for root thickness in diploid wild sweetpotato (Ipomoea trifida) using QTL-seq. Plant Product. Sci. 2022, 25, 120–129. [Google Scholar] [CrossRef]
- Park, H.; Abe, T.; Kunitake, H.; Hirano, T. Characterization of a novel mutant with inhibition of storage root formation in sweet potato. Breed. Sci. 2023, 73, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhu, J.; Sun, L.; Kong, Y.; Chen, J.; Zhu, M.; Xu, T.; Li, Z.; Dong, T. Progress on physiological and molecular mechanisms of storage root formation and development in sweetpotato. Sci. Hortic. 2023, 308, 111588. [Google Scholar] [CrossRef]
- Kim, H.S.; Wang, W.; Kang, L.; Kim, S.E.; Lee, C.J.; Park, S.C.; Park, W.S.; Ahn, M.J.; Kwak, S.S. Metabolic engineering of low-molecular-weight antioxidants in sweetpotato. Plant Biotechnol. Rep. 2020, 14, 193–205. [Google Scholar] [CrossRef]
- Ellong, E.N.; Billard, C.; Adenet, S. Comparison of physicochemical, organoleptic and nutritional abilities of eight sweet potato (Ipomoea batatas) varieties. Food Nutr. Sci. 2014, 5, 42184. [Google Scholar] [CrossRef]
- Nakagawa, S.; Ohmura, R.; Toshima, S.; Park, H.; Narasako, Y.; Hirano, T.; Otani, M.; Kunitake, H. Changes in polyphenols, anthocyanins, and DPPH radical-scavenging activities in sweetpotato (Ipomoea batatas L.) during tuber growth. Sci. Hortic. 2021, 284, 110100. [Google Scholar] [CrossRef]
- Ishiguro, K.; Yahara, S.; Yoshimoto, M. Changes in polyphenolic content and radical-scavenging activity of sweetpotato (Ipomoea batatas L.) during storage at optimal and low temperatures. J. Agric. Food Chem. 2007, 55, 10773–10778. [Google Scholar] [CrossRef] [PubMed]
- Kourouma, V.; Mu, T.H.; Zhang, M.; Sun, H.N. Comparative study on chemical composition, polyphenols, flavonoids, carotenoids and antioxidant activities of various cultivars of sweet potato. Int. J. Food Sci. Technol. 2020, 55, 369–378. [Google Scholar] [CrossRef]
- Li, Q.; Li, Y.; Li, Q.; Chen, Z.; Chen, J.; Geng, S. Evaluation of morphological and phytochemical characteristics of Mesona chinensis populations in southern China. Plant Product. Sci. 2021, 24, 374–387. [Google Scholar] [CrossRef]
- Nagao, N.; Sakuma, Y.; Funakoshi, T.; Itani, T. Variation in antioxidant capacity of the seven azuki bean (Vigna angularis) varieties with different seed coat color. Plant Product. Sci. 2023, 26, 164–173. [Google Scholar] [CrossRef]
- Pinto, G.; Illiano, A.; Carpentieri, A.; Spinelli, M.; Melchiorre, C.; Fontanarosa, C.; Serio, M.; Amoresano, A. Quantification of polyphenols and metals in Chinese tea infusions by mass spectrometry. Foods 2020, 9, 835. [Google Scholar] [CrossRef] [PubMed]
- Setoguchi, Y.; Nakagawa, S.; Ohmura, R.; Toshima, S.; Park, H.; Narasako, Y.; Hirano, T.; Otani, M.; Kunitake, H. Effect of Growth Stages on Anthocyanins and Polyphenols in the Root System of Sweet Potato. Plants 2023, 12, 1907. [Google Scholar] [CrossRef] [PubMed]
- Ishiguro, K.; Yoshinaga, M.; Kai, Y.; Maoka, T.; Yoshimoto, M. Composition, content and antioxidative activity of the carotenoids in yellow-fleshed sweetpotato (Ipomoea batatas L.). Breed. Sci. 2010, 60, 324–329. [Google Scholar] [CrossRef]
- Kim, S.H.; Ahn, Y.O.; Ahn, M.J.; Lee, H.S.; Kwak, S.S. Down-regulation of β-carotene hydroxylase increases β-carotene and total carotenoids enhancing salt stress tolerance in transgenic cultured cells of sweetpotato. Phytochemistry 2012, 74, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Kim, Y.H.; Ahn, Y.O.; Ahn, M.J.; Jeong, J.C.; Lee, H.S.; Kwak, S.S. Downregulation of the lycopene ϵ-cyclase gene increases carotenoid synthesis via the β-branch-specific pathway and enhances salt-stress tolerance in sweetpotato transgenic calli. Physiol. Plant 2013, 147, 432–442. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Ahn, Y.O.; Ahn, M.J.; Jeong, J.C.; Lee, H.S.; Kwak, S.S. Cloning and characterization of an Orange gene that increases carotenoid accumulation and salt stress tolerance in transgenic sweetpotato cultures. Plant Physiol. Biochem. 2013, 70, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Suematsu, K.; Tanaka, M.; Kurata, R.; Kai, Y. Comparative transcriptome analysis implied a ZEP paralog was a key gene involved in carotenoid accumulation in yellow-fleshed sweetpotato. Sci. Rep. 2020, 10, 20607. [Google Scholar] [CrossRef]
- Kang, L.; Park, S.C.; Ji, C.Y.; Kim, H.S.; Lee, H.S.; Kwak, S.S. Metabolic engineering of carotenoids in transgenic sweetpotato. Breed. Sci. 2017, 67, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Ling, Y.; Hong, Z.; Wei, C.; He, S.Z.; Liu, Q.C. Cloning and functional analysis of lycopene ɛ-cyclase (IbLCYe) gene from sweetpotato, Ipomoea batatas (L.) Lam. J. Integr. Agric. 2013, 12, 773–780. [Google Scholar] [CrossRef]
- Li, R.; Zhai, H.; Kang, C.; Liu, D.; He, S.; Liu, Q. De novo transcriptome sequencing of the orange-fleshed sweet potato and analysis of differentially expressed genes related to carotenoid biosynthesis. Int. J. Gen. Eng. Technol. 2015, 2015, 843802. [Google Scholar] [CrossRef]
- Khan, M.Z.; Takemura, M.; Maoka, T.; Otani, M.; Misawa, N. Carotenoid analysis of sweetpotato Ipomoea batatas and functional identification of its lycopene β-and ε-cyclase genes. Z. Naturforsch. C 2016, 71, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Ke, Q.; Kang, L.; Kim, H.S.; Xie, T.; Liu, C.; Ji, C.Y.; Kim, S.H.; Park, W.S.; Ahn, M.J.; Wang, S.; et al. Down-regulation of lycopene ε-cyclase expression in transgenic sweetpotato plants increases the carotenoid content and tolerance to abiotic stress. Plant Sci. 2019, 281, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Katayama, K.; Kobayashi, A.; Sakai, T.; Kuranouchi, T.; Kai, Y. Recent progress in sweetpotato breeding and cultivars for diverse applications in Japan. Breed. Sci. 2017, 67, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Zhao, D.; Xiao, S.; Zhang, A.; Deng, Y.; Dai, X.; Zhou, Z.; Ji, Z.; Cao, Q. Comparative Metabolomic and Transcriptomic Analyses of Phytochemicals in Two Elite Sweet Potato Cultivars for Table Use. Molecules 2022, 27, 8939. [Google Scholar] [CrossRef] [PubMed]
- Historical Weather Data Search/2021 Miyazaki/Japan Meteorological Agency. Available online: https://www.data.jma.go.jp/obd/stats/etrn/view/monthly_s1.php?prec_no=87&block_no=47830&year=2021&month=&day=&view=p1 (accessed on 4 June 2024).
- Wrolstad, R.E. Color and Pigment Analyses in Fruit Products. Ph.D. Thesis, Agricultural Experiment Station, Oregon State University, Corvallis, OR, USA, 1976; pp. 1–17. [Google Scholar]
- Šamec, D.; Karalija, E.; Šola, I.; Vujčić Bok, V.; Salopek-Sondi, B. The role of polyphenols in abiotic stress response: The influence of molecular structure. Plants 2021, 10, 118. [Google Scholar] [CrossRef] [PubMed]
- Lattanzio, V. Phenolic compounds: Introduction. In Natural Products; Springer: Berlin/Heidelberg, Germany, 2013; pp. 1543–1580. [Google Scholar] [CrossRef]
- Schmidt, É.C.; Felix, M.R.D.L.; Kreusch, M.G.; Pereira, D.T.; Costa, G.B.; Simioni, C.; Ouriques, L.C.; Steiner, N.; Chow, F.; Floh, E.S.L.; et al. Profiles of carotenoids and amino acids and total phenolic compounds of the red alga Pterocladiella capillacea exposed to cadmium and different salinities. J. Appl. Phycol. 2016, 28, 1955–1963. [Google Scholar] [CrossRef]
- Winkel-Shirley, B. Evidence for enzyme complexes in the phenylpropanoid and flavonoid pathways. Physiol. Plant 1999, 107, 142–149. [Google Scholar] [CrossRef]
- Li, L.; Yuan, H. Chromoplast biogenesis and carotenoid accumulation. Arch. Biochem. Biophys. 2013, 539, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Naikoo, M.I.; Dar, M.I.; Raghib, F.; Jaleel, H.; Ahmad, B.; Raina, A.; Khan, F.A.; Naushin, F. Role and regulation of plants phenolics in abiotic stress tolerance: An overview. In Plant Signaling Molecules; Elsevier: Amsterdam, The Netherlands, 2019; pp. 157–168. [Google Scholar] [CrossRef]
- Jha, Y.; Mohamed, H.I. Plant secondary metabolites as a tool to investigate biotic stress tolerance in plants: A review. Gesunde Pflanz. 2022, 74, 771–790. [Google Scholar] [CrossRef]
- Li, R.; Kang, C.; Song, X.; Yu, L.; Liu, D.; He, S.; Zhai, H.; Liu, Q. A ζ-carotene desaturase gene, IbZDS, increases β-carotene and lutein contents and enhances salt tolerance in transgenic sweetpotato. Plant Sci. 2017, 262, 39–51. [Google Scholar] [CrossRef]
- Aghaei, K.; Komatsu, S. Crop and medicinal plants proteomics in response to salt stress. Front. Plant Sci. 2013, 4, 41038. [Google Scholar] [CrossRef]
- Ben Abdallah, S.; Aung, B.; Amyot, L.; Lalin, I.; Lachâal, M.; Karray-Bouraoui, N.; Hannoufa, A. Salt stress (NaCl) affects plant growth and branch pathways of carotenoid and flavonoid biosyntheses in Solanum nigrum. Acta Physiol. Plant 2016, 38, 72. [Google Scholar] [CrossRef]
- Kiani, R.; Arzani, A.; Mirmohammady Maibody, S.A.M. Polyphenols, flavonoids, and antioxidant activity involved in salt tolerance in wheat, Aegilops cylindrica and their amphidiploids. Front. Plant Sci. 2021, 12, 646221. [Google Scholar] [CrossRef]

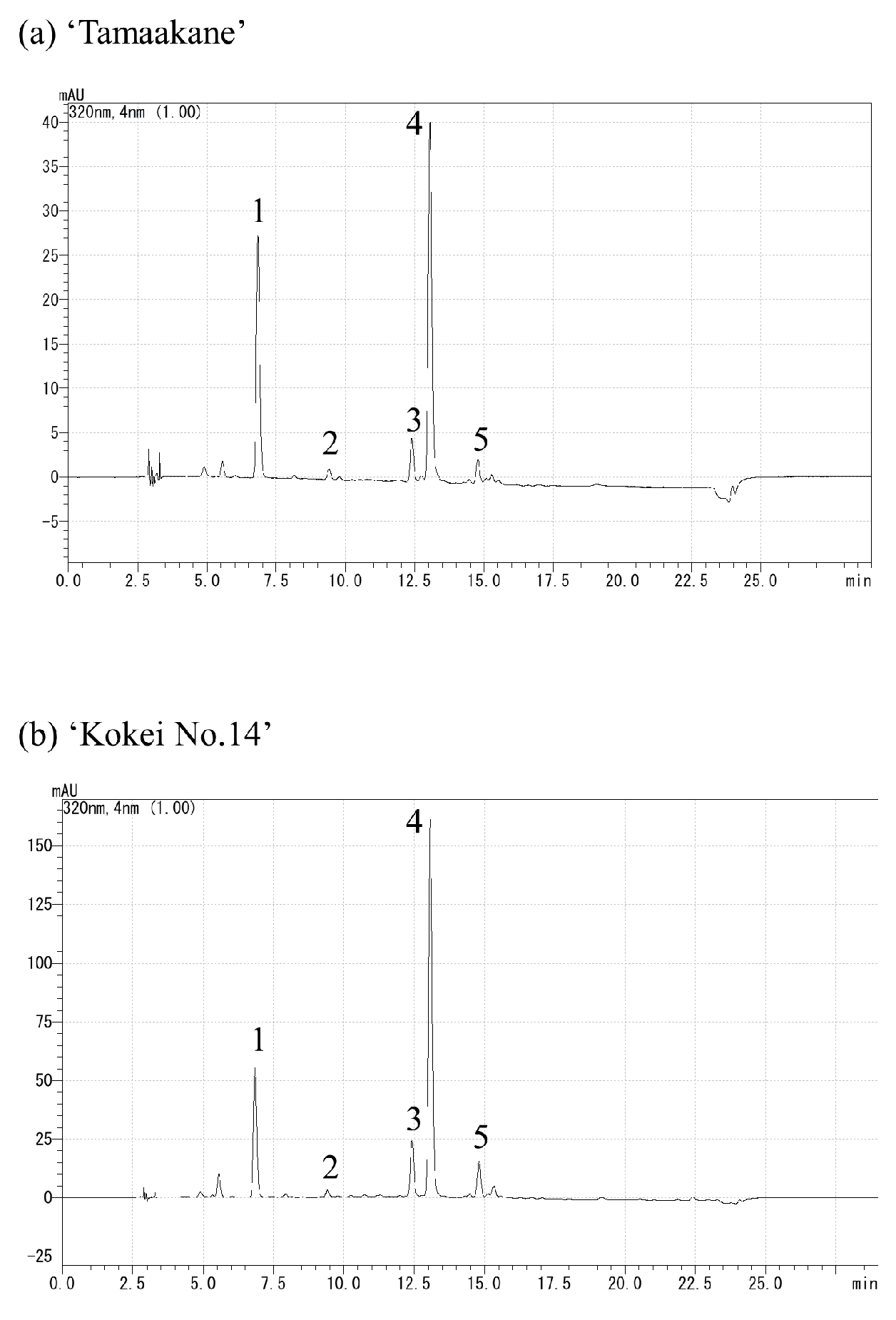
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).