

Preliminary Study on Programmed Cell Death during Calyx Abscission of Korla Fragrant Pear
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Description of the Experimental Site and Plant Material
2.2. Sampling Methods
2.3. Investigation of Korla Fragrant Pear Calyx Abscission Patterns
2.4. Characteristics of PCD during Korla Fragrant Pear Calyx Abscission
2.4.1. Cellular Ultrastructure Observation
2.4.2. DAPI Fluorescence with TUNEL Assay
2.4.3. DNA Fragmentation Assay
2.5. Regulatory Signals of PCD during Korla Fragrant Pear Calyx Abscission
2.5.1. Determination of Endogenous Hormones
2.5.2. ROS Determination
2.6. Statistical Analysis
3. Results
3.1. Apparent Appearance of Korla Fragrant Pear during Calyx Abscission
3.1.1. Changes in External Morphological Characteristics during Calyx Abscission
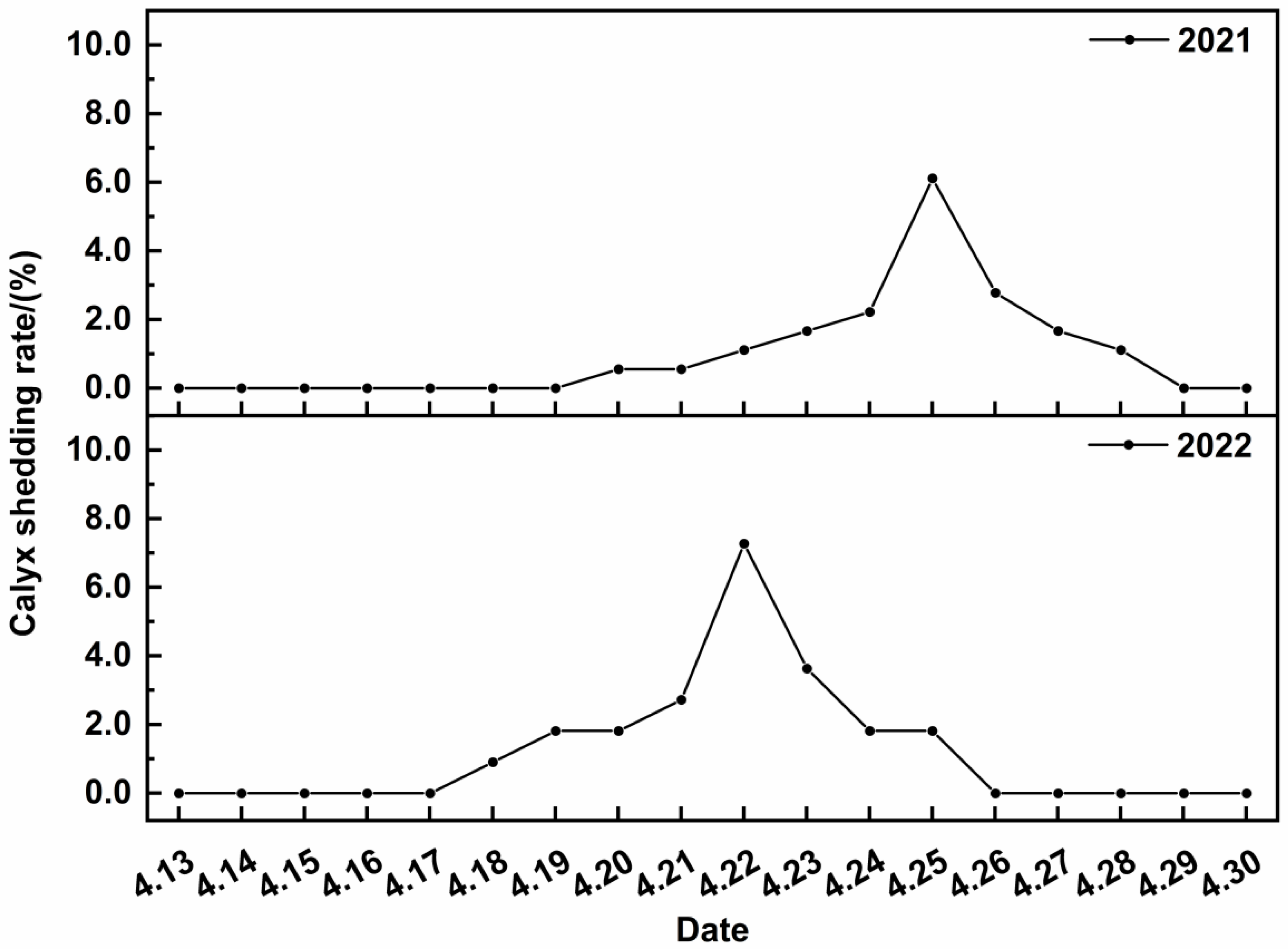
3.1.2. Calyx Abscission Patterns in 2021 and 2022
3.2. Characteristics of Programmed Cell Death during Korla Fragrant Pear Calyx Abscission
3.2.1. Cellular Ultrastructure
3.2.2. DAPI Fluorescence and TUNEL Assay
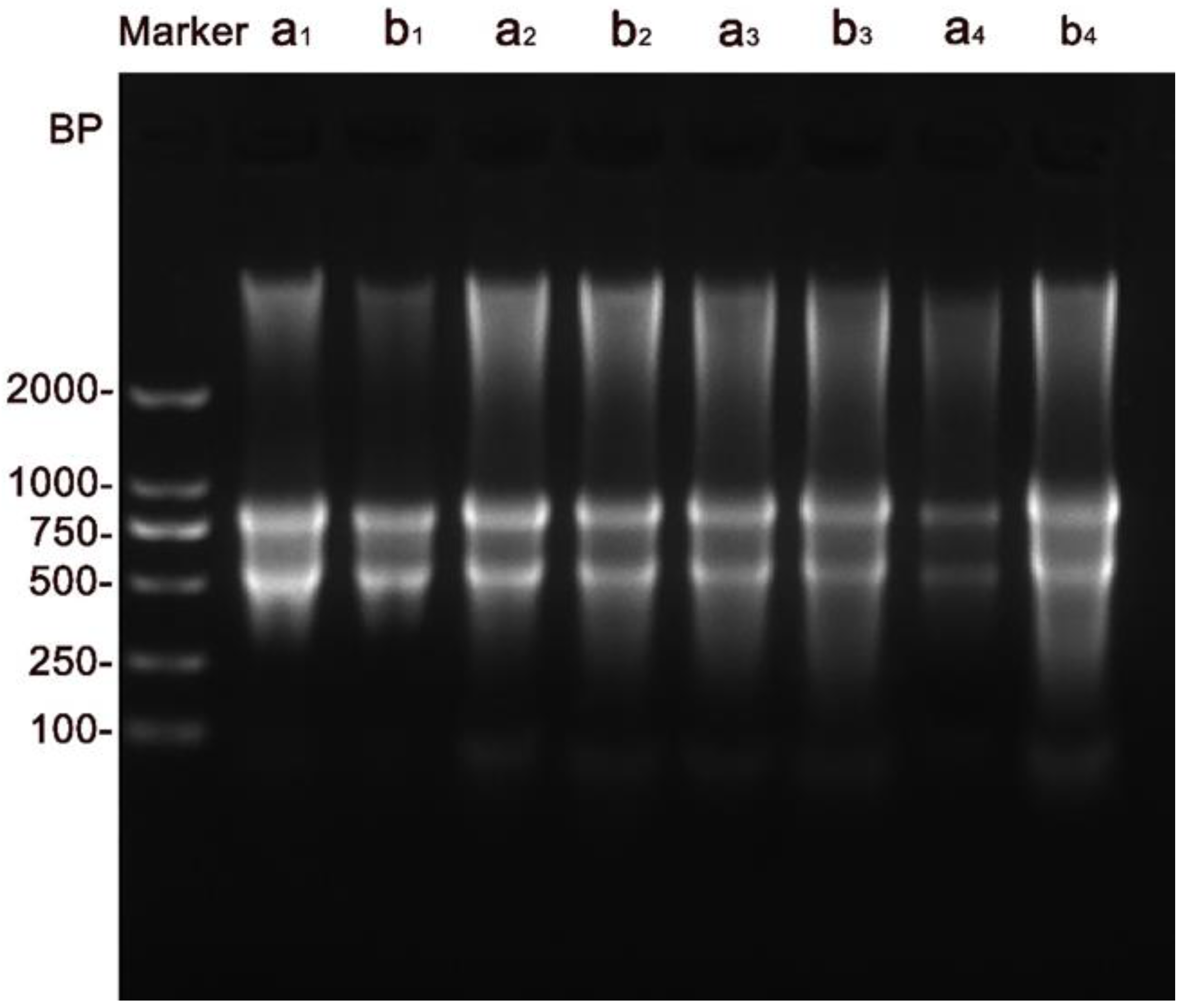
3.2.3. DNA Fragmentation Assay
3.3. PCD-Regulated Signals during Korla Fragrant Pear Calyx Abscission
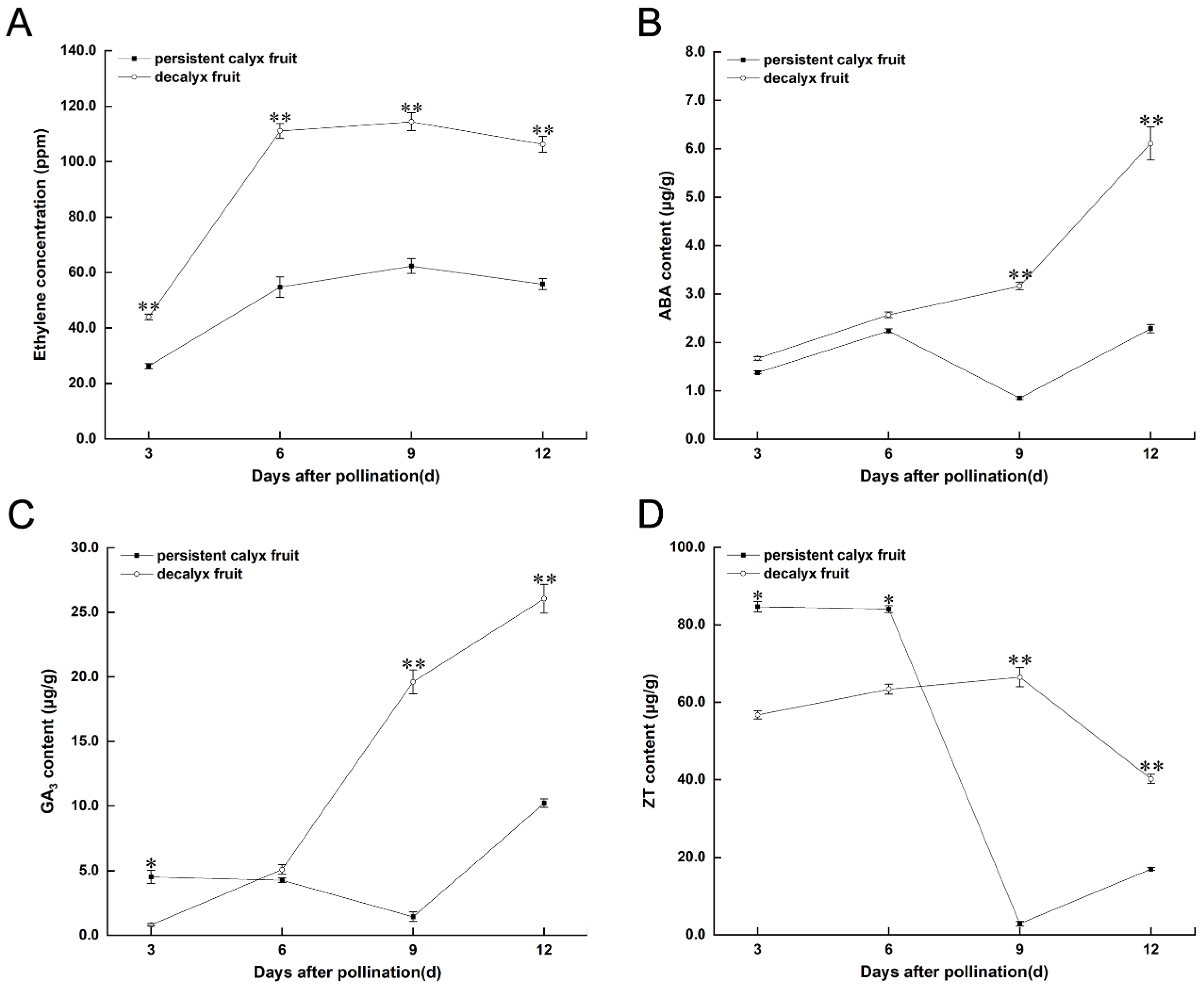
3.3.1. Endogenous Hormone
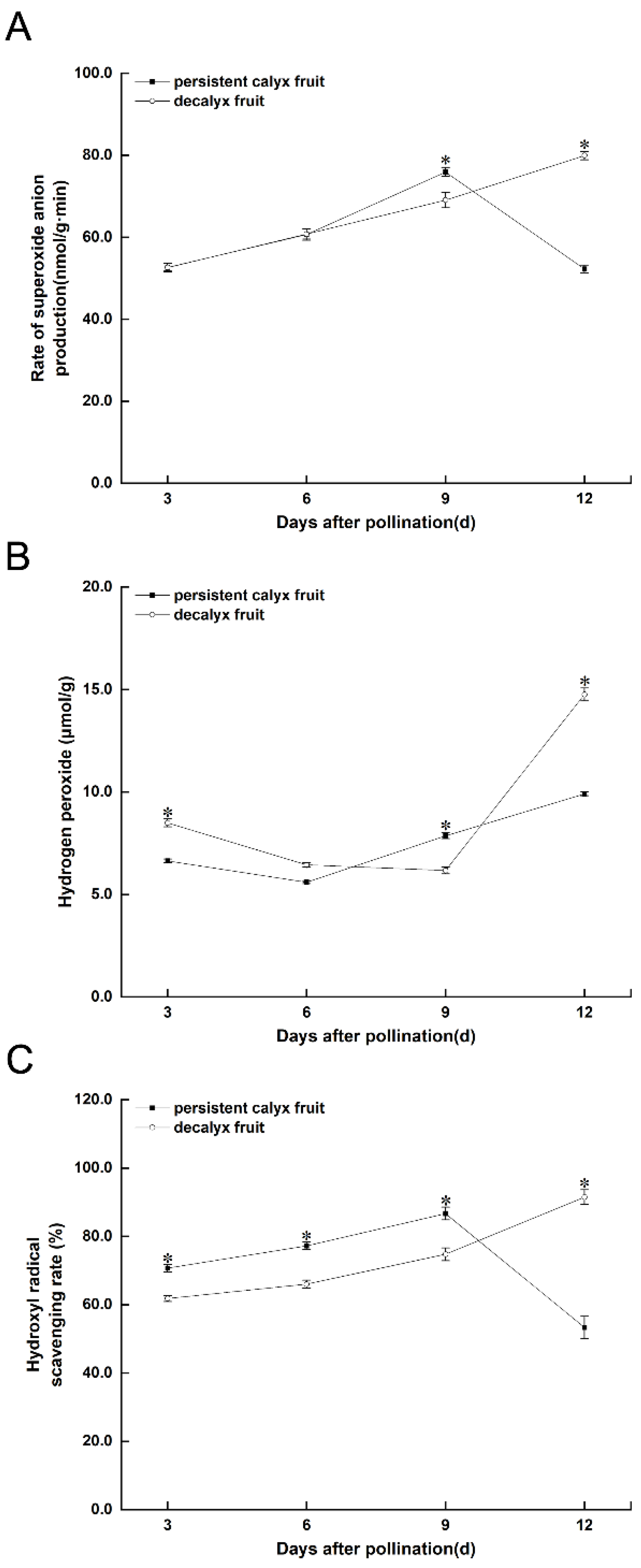
3.3.2. Reactive Oxygen Species
4. Discussion
4.1. Apparent Characteristics of Calyx Abscission in Korla Fragrant Pear
4.2. PCD Was Involved in Calyx Abscission in Korla Fragrant Pear
4.3. Relationship between PCD and Plant Hormone during Calyx Abscission in Korla Fragrant Pear
4.4. Relationship between PCD and Reactive Oxygen Species during Calyx Abscission in Korla Fragrant Pear
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Locato, V.; De Gara, L. Programmed Cell Death in Plants: An Overview. In Plant Programmed Cell Death: Methods and Protocols; Humana Press: New York, NY, USA, 2018; pp. 1–8. [Google Scholar]
- Huysmans, M.; Buono, R.A.; Skorzinski, N.; Radio, M.C.; De Winter, F.; Parizot, B.; Mertens, J.; Karimi, M.; Fendrych, M.; Nowack, M.K. NAC Transcription Factors ANAC087 and ANAC046 Control Distinct Aspects of Programmed Cell Death in the Arabidopsis Columella and Lateral Root Cap. Plant Cell 2018, 30, 2197–2213. [Google Scholar] [CrossRef] [PubMed]
- Parveen, S.; Altaf, F.; Farooq, S.; Lone, M.L.; Haq, A.U.; Tahir, I. The Swansong of Petal Cell Death: Insights into the mechanism and regulation of ethylene-mediated flower senescence. J. Exp. Bot. 2023, 74, 3961–3974. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zhang, X.; Guo, W.; Fu, K.; Li, C.; Li, C. Impact of Ethylene and Abscisic Acid on Programmed Cell Death in Endosperm and Grain Filling of Wheat (Triticum aestivum L.). J. Plant Growth Regul. 2023, 42, 4275–4289. [Google Scholar] [CrossRef]
- Huang, L.; Liu, Y.; Wang, X.; Jiang, C.; Zhao, Y.; Lu, M.; Zhang, J. Peroxisome-Mediated Reactive Oxygen Species Signals Modulate Programmed Cell Death in Plants. Int. J. Mol. Sci. 2022, 23, 10087. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.; Zhao, X.; Li, W.; Hussain, J.; Qi, G.; Liu, S. Calcium Signaling in Plant Programmed Cell Death. Cells 2021, 10, 1089. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Liu, Y.; Liang, Y.; Lu, J.; Jiang, C.; Fei, Z.; Jiang, C.Z.; Ma, C.; Gao, J. Rosa hybrida Rh ERF 1 and Rh ERF 4 Mediate Ethylene-and Auxin-Regulated Petal Abscission by Influencing Pectin Degradation. Plant J. 2019, 99, 1159–1171. [Google Scholar] [CrossRef] [PubMed]
- Meir, S.; Philosoph-Hadas, S.; Sundaresan, S.; Selvaraj, K.V.; Burd, S.; Ophir, R.; Kochanek, B.; Reid, M.S.; Jiang, C.-Z.; Lers, A. Microarray Analysis of the Abscission-Related Transcriptome in the Tomato Flower Abscission Zone in Response to Auxin Depletion. Plant Physiol. 2010, 154, 1929–1956. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Zhang, C.; Lai, D.; Sun, Y.; Samma, M.K.; Zhang, J.; Shen, W. Hydrogen Sulfide Delays GA-Triggered Programmed Cell Death in Wheat Aleurone Layers by the Modulation of Glutathione Homeostasis and Heme Oxygenase-1 Expression. J. Plant Physiol. 2014, 171, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Cortleven, A.; Leuendorf, J.E.; Frank, M.; Pezzetta, D.; Bolt, S.; Schmülling, T. Cytokinin Action in Response to Abiotic and Biotic Stresses in Plants. Plant Cell Environ. 2019, 42, 998–1018. [Google Scholar] [CrossRef]
- Ye, C.; Zheng, S.; Jiang, D.; Lu, J.; Huang, Z.; Liu, Z.; Zhou, H.; Zhuang, C.; Li, J. Initiation and Execution of Programmed Cell Death and Regulation of Reactive Oxygen Species in Plants. Int. J. Mol. Sci. 2021, 22, 12942. [Google Scholar] [CrossRef]
- Wang, Y.; Shirakawa, M.; Ito, T. Dynamic Changes in Reactive Oxygen Species in the Shoot Apex Contribute to Stem Cell Death in Arabidopsis thaliana. Int. J. Mol. Sci. 2022, 23, 3864. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Tian, X.; Wu, H.; Hao, X.; Gao, B.; Zhang, H.; Feng, J.; Han, L. H2O2 Signaling Modulates Glycoprotein-1 Induced Programmed Cell Death in Tobacco Suspension Cells. Pestic. Biochem. Physiol. 2021, 171, 104697. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Yang, H.; Ren, D.; Li, Y. Activation of ZmMKK10, a Maize Mitogen-Activated Protein Kinase Kinase, Induces Ethylene-Dependent Cell Death. Plant Sci. 2017, 264, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Yan, P.; Zheng, Q.; Wang, Z.; Chen, Q.; Wang, Y. Changes in the Metabolome and Nutritional Quality of Pulp from Three Types of Korla Fragrant Pears with Different Appearances as Revealed by Widely Targeted Metabolomics. Plants 2023, 12, 3981. [Google Scholar] [CrossRef] [PubMed]
- Lers, A.; Sonego, L.; Green, P.J.; Burd, S. Suppression of LX Ribonuclease in Tomato Results in a Delay of Leaf Senescence and Abscission. Plant Physiol. 2006, 142, 710–721. [Google Scholar] [CrossRef] [PubMed]
- Bar-Dror, T.; Dermastia, M.; Kladnik, A.; Žnidarič, M.T.; Novak, M.P.; Meir, S.; Burd, S.; Philosoph-Hadas, S.; Ori, N.; Sonego, L.; et al. Programmed Cell Death Occurs Asymmetrically during Abscission in Tomato. Plant Cell 2011, 23, 4146–4163. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, J.M.; Morcillo, R.L.; Vierheilig, H.; Ocampo, J.A.; Ludwig-Müller, J.; Garrido, J.M.G. Mycorrhization of the Notabilis and Sitiens Tomato Mutants in Relation to Abscisic Acid and Ethylene Contents. J. Plant Physiol. 2010, 167, 606–613. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Chen, R.; Ma, J. Detection of Antioxidant Enzymes and DNA Dynamic Changes During Senescence of Eucalyptuses Phellogen Zone. Acta Bot. Boreali-Occident. Sin. 2012, 32, 940–947. [Google Scholar]
- Heo, S.; Chung, Y.S. Validation of MADS-Box Genes from Apple Fruit Pedicels during Early Fruit Abscission by Transcriptome Analysis and Real-Time PCR. Genes Genom. 2019, 41, 1241–1251. [Google Scholar] [CrossRef]
- Pei, M.; Niu, J.; Li, C.; Cao, F.; Quan, S. Identification and Expression Analysis of Genes Related to Calyx Persistence in Korla Fragrant Pear. BMC Genom. 2016, 17, 132. [Google Scholar] [CrossRef]
- Kirschner, G.K. A matter of Life and Death: Networks regulating programmed cell death in Arabidopsis. Plant J. 2023, 115, 1463–1464. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Zhou, L.; Quan, S.; Xu, H.; Yang, J.; Niu, J. Integrated Analysis of mRNA-Seq and miRNA-seq in Calyx Abscission Zone of Korla Fragrant Pear Involved in Calyx Persistence. BMC Plant Biol. 2019, 19, 192. [Google Scholar] [CrossRef]
- Ma, H.; Wang, Y.; Wen, X.; Qi, M.; Li, J. Effects of Different Reagent Treatments on the Microstructure of Calyx Tubes with or without Calyx of Korla Fragrant Pear. J. Fruit Sci. 2011, 28, 518–520. [Google Scholar]
- Ma, L.; Zhou, L.; Quan, S.; Xu, H.; Yang, J.; Niu, J. Relationship between Abscission of Calyx of Korla Fragrant Pear and Plant Hormones in Different Parts of the Separation Zone. Plant Physiol. J. 2022, 58, 1369–1380. [Google Scholar]
- Lee, J.; Chen, H.; Lee, G.; Emonet, A.; Kim, S.; Shim, D.; Lee, Y. MSD2-Mediated ROS Metabolism Fine-Tunes the Timing of Floral Organ Abscission in Arabidopsis. New Phytol. 2022, 235, 2466–2480. [Google Scholar] [CrossRef]
- Van Doorn, W.G. Classes of Programmed Cell Death in Plants, Compared to Those in Animals. J. Exp. Bot. 2011, 62, 4749–4761. [Google Scholar] [CrossRef] [PubMed]
- Ni, X.-L.; Gui, M.-Y.; Tan, L.-L.; Zhu, Q.; Liu, W.-Z.; Li, C.-X. Programmed Cell Death and Aerenchyma Formation in Water-Logged Sunflower Stems and Its Promotion by Ethylene and ROS. Front. Plant Sci. 2019, 9, 1928. [Google Scholar] [CrossRef]
- Wang, M.; Oppedijk, B.J.; Lu, X.; Van Duijn, B.; Schilperoort, R.A. Apoptosis in Barley Aleurone during Germination and Its Inhibition by Abscisic Acid. Plant Mol. Biol. 1996, 32, 1125–1134. [Google Scholar] [CrossRef] [PubMed]
- Ni, X.L.; Su, H.; Zhou, Y.F.; Wang, F.H.; Liu, W.Z. Leaf-Shape Remodeling: Programmed Cell Death in Fistular Leaves of a Llium fistulosum. Physiol. Plant. 2015, 153, 419–431. [Google Scholar] [CrossRef]
- Feng, Y.-Q.; Shen, Y.-Y.; Qin, L.; Cao, Q.-Q.; Han, Z.-H. Short catkin1, a Novel Mutant of Castanea mollissima, is Associated with Programmed Cell Death during Chestnut Staminate Flower Differentiation. Sci. Hortic. 2011, 130, 431–435. [Google Scholar] [CrossRef]
- Trobacher, C.P. Ethylene and Programmed Cell Death in Plants. Botany 2009, 87, 757–769. [Google Scholar] [CrossRef]
- Steffens, B.; Sauter, M. Epidermal Cell Death in Rice is Regulated by Ethylene, Gibberellin, and Abscisic Acid. Plant Physiol. 2005, 139, 713–721. [Google Scholar] [CrossRef] [PubMed]
- Orzáez, D.; Granell, A. DNA Fragmentation is Regulated by Ethylene during Carpel Senescence in Pisum sativum. Plant J. 1997, 11, 137–144. [Google Scholar] [CrossRef]
- Li, N.; Han, X.; Feng, D.; Yuan, D.; Huang, L.-J. Signaling Crosstalk between Salicylic Acid and Ethylene/Jasmonate in Plant Defense: Do We Understand What They Are Whispering? Int. J. Mol. Sci. 2019, 20, 671. [Google Scholar] [CrossRef] [PubMed]
- Lombardi, L.; Mariotti, L.; Picciarelli, P.; Ceccarelli, N.; Lorenzi, R. Ethylene Produced by the Endosperm is Involved in the Regulation of Nucellus Programmed Cell Death in Sechium edule Sw. Plant Sci. 2012, 187, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Wu, D.; Li, S.; Guo, N.; Gao, J.; Sun, X.; Cai, Y. Transcriptomic Profiling Identifies Differentially Expressed Genes Associated with Programmed Cell Death of Nucellar Cells in Ginkgo biloba L. BMC Plant Biol. 2019, 19, 91. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Peng, J.; Wen, X.; Guo, H. Ethylene-Insensitive3 is a Senescence-associated Gene that Accelerates Age-dependent Leaf Senescence by Directly Repressing miR164 Transcription in Arabidopsis. Plant Cell 2013, 25, 3311–3328. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Xiao, Y.; Deng, X.; Zhang, H.; Li, T.; Chen, H. Exogenous Hydrogen Peroxide Contributes to Heme Oxygenase-1 Delaying Programmed Cell Death in Isolated Aleurone Layers of Rice Subjected to Drought Stress in a cGMP-dependent Manner. Front. Plant Sci. 2018, 9, 84. [Google Scholar] [CrossRef] [PubMed]
- Gechev, T.S.; Hille, J. Hydrogen Peroxide as a Signal Controlling Plant Programmed Cell Death. J. Cell Biol. 2005, 168, 17–20. [Google Scholar] [CrossRef]
- Baxter, A.; Mittler, R.; Suzuki, N. ROS as Key Players in Plant Stress Signalling. J. Exp. Bot. 2013, 65, 1229–1240. [Google Scholar] [CrossRef]
- Černý, M.; Habánová, H.; Berka, M.; Luklová, M.; Brzobohatý, B. Hydrogen Peroxide: Its Role in Plant Biology and Crosstalk with Signalling Networks. Int. J. Mol. Sci. 2018, 19, 2812. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Liu, M.; Chen, G.; Yuan, L.; Hou, J.; Zhu, S.; Zhang, B.; Li, G.; Pang, X.; Wang, C. TMT-Based Comparative Proteomic Analysis of the Male-Sterile Mutant ms01 sheds Light on Sporopollenin Production and Pollen Development in Wucai (Brassica campestris L.). J. Proteom. 2022, 254, 104475. [Google Scholar] [CrossRef] [PubMed]
- Pan, R.; Han, H.; Medison, M.B.; Abou-Elwafa, S.F.; Liu, Y.; Yang, X.; Zhang, W. Aerenchyma Formation in the Root of Leaf-Vegetable Sweet Potato: Programmed Cell Death Initiated by Ethylene-Mediated H2O2 Accumulation. Physiol. Plant. 2021, 173, 2361–2375. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wen, Y.; Shao, B.; Hao, Z.; Wang, C.; Sun, T.; Han, Y.; Tian, J.; Zhang, F. Preliminary Study on Programmed Cell Death during Calyx Abscission of Korla Fragrant Pear. Horticulturae 2024, 10, 637. https://doi.org/10.3390/horticulturae10060637
Wen Y, Shao B, Hao Z, Wang C, Sun T, Han Y, Tian J, Zhang F. Preliminary Study on Programmed Cell Death during Calyx Abscission of Korla Fragrant Pear. Horticulturae. 2024; 10(6):637. https://doi.org/10.3390/horticulturae10060637
Chicago/Turabian StyleWen, Yue, Baijunjie Shao, Zhichao Hao, Chunfeng Wang, Tianyu Sun, Yutao Han, Jia Tian, and Feng Zhang. 2024. "Preliminary Study on Programmed Cell Death during Calyx Abscission of Korla Fragrant Pear" Horticulturae 10, no. 6: 637. https://doi.org/10.3390/horticulturae10060637
APA StyleWen, Y., Shao, B., Hao, Z., Wang, C., Sun, T., Han, Y., Tian, J., & Zhang, F. (2024). Preliminary Study on Programmed Cell Death during Calyx Abscission of Korla Fragrant Pear. Horticulturae, 10(6), 637. https://doi.org/10.3390/horticulturae10060637