
Expression of Cystathionine-β-Synthase (CBS) Genes in Grape under Various Abiotic Stresses and Subcellular Localization of VvCBS27

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Stress Treatment
2.2. Identification and Characterization of Grape CBS Genes
2.3. Construction and Phylogenetic Analysis of the CBS Gene Family in Grape
2.4. Collinearity and Protein Interaction Analysis of the CBS Gene Family in Grape
2.5. Subcellular Localization and Protein Secondary Structure Analysis of CBS Gene in Grape
2.6. Prediction of cis-Acting Elements and Analysis of Tissue Differential Expression Pattern of CBS Genes in Grape
2.7. Gene Expression Analysis by RT-qPCR
2.8. Cloning and Vector Construction of the VvCBS27 Gene
2.9. Subcellular Localization of VvCBS27 Protein
3. Results
3.1. Analysis of Physicochemical Properties and Chromosome Mapping of the CBS Gene Family in Grape
3.2. Gene Structure and Phylogenetic Analysis of the CBS Gene Family in Grape
3.3. Subcellular Localization and Secondary Structure Analysis of the CBS Gene Family in Grape
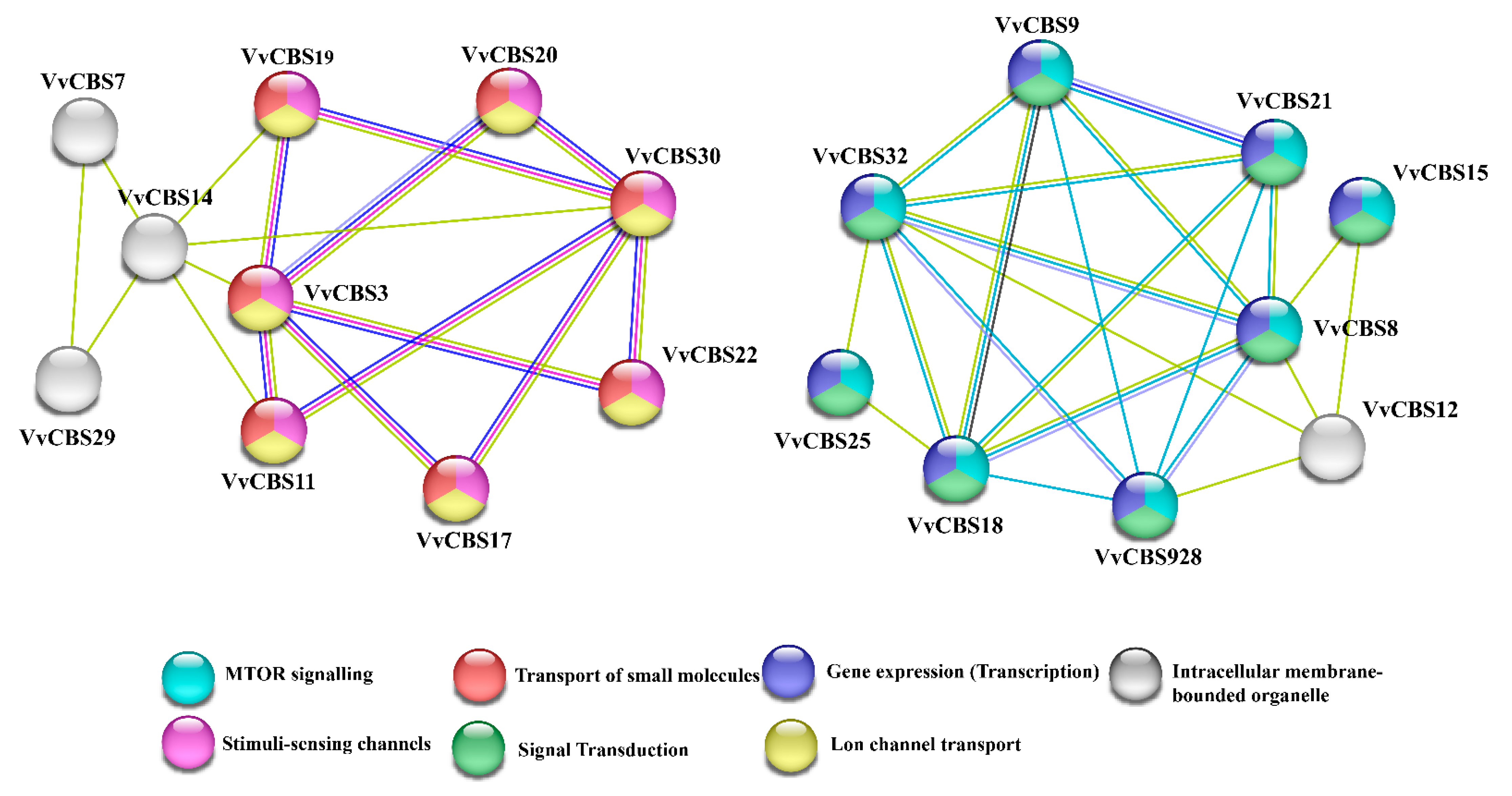
3.4. Collinearity and Protein Interaction Analysis of the CBS Gene Family in Grape
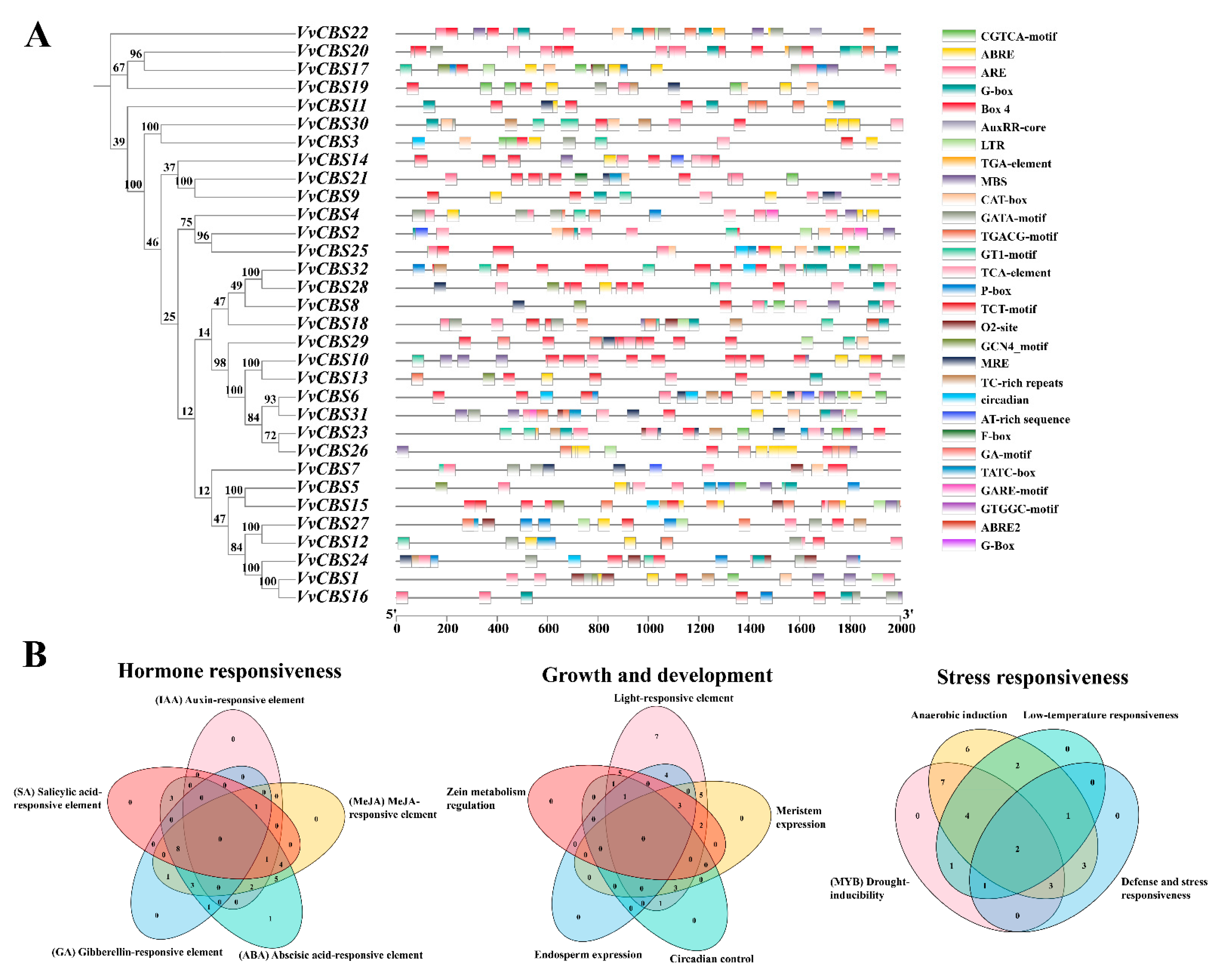
3.5. Prediction of cis-Acting Elements in the VvCBS Gene Family
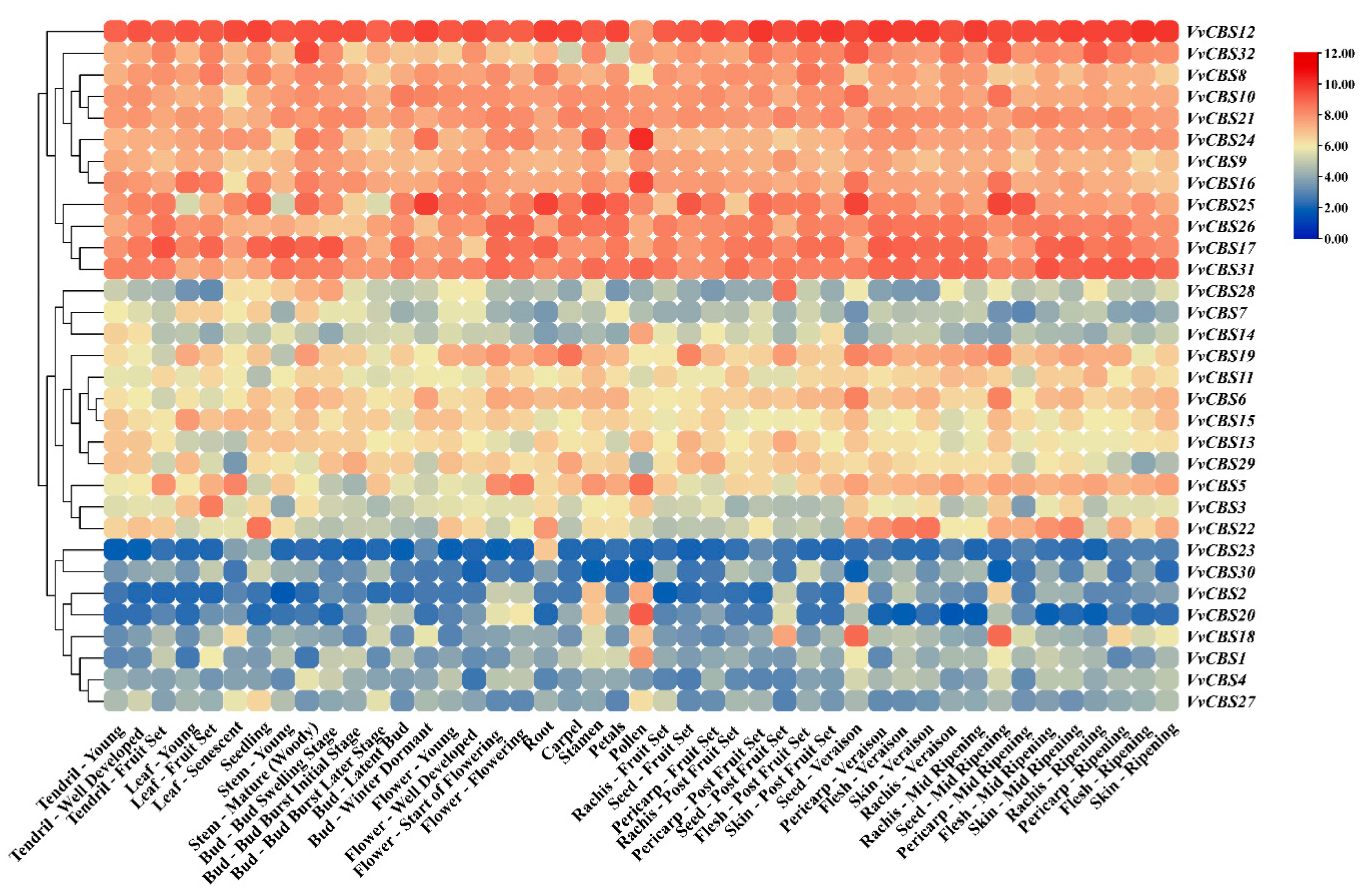
3.6. Analysis of Expression Pattern of CBS Gene Family in Grape
3.7. Expression of VvCBS in Grape under Abiotic Stress Treatment
3.8. VvCBS27 Gene Cloning and Subcellular Localization
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bateman, A. The structure of a domain common to archaebacteria and the homocystinuria disease protein. Trends Biochem. Sci. 1997, 22, 12–13. [Google Scholar] [CrossRef] [PubMed]
- Ignoul, S.; Eggermont, J. CBS domains: Structure, function, and pathology in human proteins. Am. J. Physiol. Cell Physiol. 2005, 289, C1369–C1378. [Google Scholar] [CrossRef] [PubMed]
- Ereño-Orbea, J.; Oyenarte, I.; Martínez-Cruz, L.A. CBS domains: Ligand binding sites and conformational variability. Arch. Biochem. Biophys. 2013, 540, 70–81. [Google Scholar] [CrossRef]
- Anashkin, V.A.; Baykov, A.A.; Lahti, R. Enzymes regulated via cystathionine β-synthase domains. Biochemistry 2017, 82, 1079–1087. [Google Scholar] [CrossRef] [PubMed]
- Kemp, B.E. Bateman domains and adenosine derivatives form a binding contract. J. Clin. Investig. 2004, 113, 182–184. [Google Scholar] [CrossRef]
- Zhang, R.; Evans, G.; Rotella, F.J.; Westbrook, E.M.; Beno, D.; Huberman, E.; Joachimiak, A.; Collart, F.R. Characteristics and crystal structure of bacterial inosine-5 ‘-monophosphate dehydrogenase. Biochemistry 1999, 38, 4691–4700. [Google Scholar] [CrossRef]
- Baykov, A.A.; Tuominen, H.K.; Lahti, R. The CBS domain: A protein module with an emerging prominent role in regulation. ACS Chem. Biol. 2011, 6, 1156–1163. [Google Scholar] [CrossRef]
- Hackenberg, C.; Hakanpää, J.; Cai, F.; Antonyuk, S.; Eigner, C.; Meissner, S.; Laitaoja, M.; Jänis, J.; Kerfeld, C.A.; Dittmann, E.; et al. Structural and functional insights into the unique CBS-CP12 fusion protein family in cyanobacteria. Proc. Natl. Acad. Ences 2018, 115, 7141–7146. [Google Scholar] [CrossRef]
- Xiao, B.; Heath, R.; Saiu, P.; Leiper, F.C.; Leone, P.; Jing, C.; Walker, P.A.; Haire, L.; Eccleston, J.F.; Davis, C.T.; et al. Structural basis for AMP binding to mammalian AMP-activated protein kinase. Nature 2007, 449, 496–500. [Google Scholar] [CrossRef]
- Yoo, K.S.; Ok, S.H.; Jeong, B.C.; Jung, K.W.; Cui, M.H.; Hyoung, S.; Lee, M.R.; Song, H.K.; Shin, J.S. Single cystathionine β-synthase domain–containing proteins modulate development by regulating the thioredoxin system in Arabidopsis. Plant Cell 2011, 23, 3577–3594. [Google Scholar] [CrossRef]
- Ke, X.; Xiao, H.; Peng, Y.; Wang, J.; Lv, Q.; Wang, X. Phosphoenolpyruvate reallocation links nitrogen fixation rates to root nodule energy state. Science 2022, 378, 971–977. [Google Scholar] [CrossRef] [PubMed]
- Sintchak, M.D.; Fleming, M.A.; Futer, O.; Raybuck, S.A.; Chambers, S.P.; Caron, P.R.; Murcko, M.A.; Wilson, K.P. Structure and mechanism of inosine monophosphate dehydrogenase in complex with the immunosuppressant mycophenolic acid. Cell 1996, 85, 921–930. [Google Scholar] [CrossRef] [PubMed]
- Buey, R.M.; Fernández-Justel, D.; Marcos-Alcalde, Í.; Winter, G.; Gómez-Puertas, P.; de Pereda, J.M.; Luis Revuelta, J. A nucleotide-controlled conformational switch modulates the activity of eukaryotic IMP dehydrogenases. Sci. Rep. 2017, 7, 2648. [Google Scholar] [CrossRef] [PubMed]
- Schmidt-Rose, T.; Jentsch, T.J. Reconstitution of functional voltage-gated chloride channels from complementary fragments of CLC-1. J. Biol. Chem. 1997, 272, 20515–20521. [Google Scholar] [CrossRef] [PubMed]
- Kushwaha, H.R.; Singh, A.K.; Sopory, S.K.; Singla-Pareek, S.L.; Pareek, A. Genome wide expression analysis of CBS domain containing proteins in Arabidopsis thaliana (L.) Heynh and Oryza sativa L. reveals their developmental and stress regulation. BMC Genomics 2009, 10, 200. [Google Scholar] [CrossRef]
- Crozet, P.; Margalha, L.; Confraria, A.; Rodrigues, A.; Martinho, C.; Adamo, M.; Elias, C.A.; Baena-González, E. Mechanisms of regulation of SNF1/AMPK/SnRK1 protein kinases. Front. Plant Sci. 2014, 5, 190. [Google Scholar] [CrossRef] [PubMed]
- Shahbaaz, M.; Ahmad, F.; Imtaiyaz Hassan, M. Structure-based functional annotation of putative conserved proteins having lyase activity from Haemophilus influenzae. 3 Biotech 2015, 5, 317–336. [Google Scholar] [CrossRef]
- Scott, J.W.; Hawley, S.A.; Green, K.A.; Anis, M.; Stewart, G.; Scullion, G.A.; Norman, D.G.; Hardie, D.G. CBS domains form energy-sensing modules whose binding of adenosine ligands is disrupted by disease mutations. J. Clin. Investig. 2004, 113, 274–284. [Google Scholar] [CrossRef]
- Giménez-Mascarell, P.; González-Recio, I.; Fernández-Rodríguez, C.; Oyenarte, I.; Müller, D.; Martínez-Chantar, M.L.; Martínez-Cruz, L.A. Current structural knowledge on the CNNM family of magnesium transport mediators. Int. J. Mol. Sci. 2019, 20, 1135. [Google Scholar] [CrossRef]
- Xiao, B.; Sanders, M.J.; Underwood, E.; Heath, R.; Mayer, F.V.; Carmena, D.; Jing, C.; Walker, P.A.; Eccleston, J.F.; Haire, L.F.; et al. Structure of mammalian AMPK and its regulation by ADP. Nature 2011, 472, 230–233. [Google Scholar] [CrossRef]
- Hardie, D.G.; Hawley, S.A. AMP-activated protein kinase: The energy charge hypothesis revisited. Bioessays 2001, 23, 1112–1119. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.Y.; Cang, J.; Lu, Q.W.; Tian, Y.; Song, C.H.; Ren, Z.P.; Sun, X.Z.; Fan, X.P.; Wang, D.J.; Fu, L.S. Genome-Wide Analysis of CBS gene Family in Hexaploid Wheat. J. Triticeae Crops. 2020, 23, 421–433. (In Chinese) [Google Scholar]
- Liu, H.; Wang, Q.; Xie, L.; Xu, K.; Zhang, F.; Ruan, X.; Li, L.; Tan, G. Genome-wide identification of cystathionine beta synthase genes in wheat and its relationship with anther male sterility under heat stress. Front. Plant Sci. 2022, 13, 1061472. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.R.; Li, J.; Lin, J.X.; Xu, H.M.; Chu, N.; Wang, Q.N.; Gao, S.J. Genome-wide characterization of cys-tathionine-β-synthase domain-containing proteins in sugarcane reveals their role in defense responses under multiple stressors. Front. Plant Sci. 2022, 13, 985653. [Google Scholar] [CrossRef]
- Chen, Z.Q.; Li, G.; Yan, J.W.; Guo, X.R.; Hu, C.Q.; Wang, Y.; Wang, F.; Tian, D.G. Identification and characterisation of CBS gene family in maize. Jiangsu Agric. Sci 2024, 52, 41–49. (In Chinese) [Google Scholar]
- Jung, K.W.; Kim, Y.Y.; Yoo, K.S.; Ok, S.H.; Cui, M.H.; Jeong, B.C.; Yoo, S.D.; Jeung, J.U.; Shin, J.S. A cystathionine-β-synthase domain-containing protein, CBSX2, regulates endothecial secondary cell wall thickening in anther development. Plant Cell Physiol. 2013, 54, 195–208. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.S.; So, W.M.; Kim, S.Y.; Noh, M.; Hyoung, S.; Yoo, K.S.; Shin, J.S. CBSX3-Trxo-2 regulates ROS generation of mitochondrial complex II (succinate dehydrogenase) in Arabidopsis. Plant Sci. 2020, 294, 110458. [Google Scholar] [CrossRef]
- Wang, X.; Ren, X.; Zhu, L.; He, G.C. OsBi1, a rice gene, encodes a novel protein with a CBS-like domain and its expression is induced in responses to herbivore feeding. Plant Sci. 2004, 166, 1581–1588. [Google Scholar] [CrossRef]
- Mou, S.; Shi, L.; Lin, W.; Liu, Y.; Shen, L.; Guan, D.; He, S. Over-expression of rice CBS domain containing protein, OsCBSX3, confers rice resistance to Magnaporthe oryzae inoculation. Int. J. Mol. Sci. 2015, 16, 15903–15917. [Google Scholar] [CrossRef]
- Singh, A.K.; Kumar, R.; Pareek, A.; Sopory, S.K.; Singla-Pareek, S.L. Overexpression of rice CBS domain containing protein improves salinity, oxidative, and heavy metal tolerance in transgenic tobacco. Mol. Biotechnol. 2012, 52, 205–216. [Google Scholar] [CrossRef]
- Tomar, S.; Subba, A.; Bala, M.; Singh, A.K.; Pareek, A.; Singla-Pareek, S.L. Genetic conservation of CBS domain containing protein family in Oryza species and their association with abiotic stress responses. Int. J. Mol. Sci. 2022, 23, 1687. [Google Scholar] [CrossRef] [PubMed]
- Hao, Q.; Shang, W.; Zhang, C.; Chen, H.; Chen, L.; Yuan, S.; Chen, S.; Zhang, X.; Zhou, X. Identification and comparative analysis of CBS domain-containing proteins in soybean (Glycine max) and the primary function of GmCBS21 in enhanced tolerance to low nitrogen stress. Int. J. Mol. Sci. 2016, 17, 620. [Google Scholar] [CrossRef] [PubMed]
- Hao, Q.; Yang, Y.; Shan, Z.; Chen, H.; Zhang, C.; Chen, L.; Yuan, S.; Zhang, X.; Chen, S.; Yang, Z.; et al. Genome-wide investigation and expression profiling under abiotic stresses of a soybean unknown function (DUF21) and cystathionine-β-synthase (CBS) domain-containing protein family. Biochem. Genet. 2021, 59, 83–113. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Li, M.; Harrison, M.T.; Fahad, S.; Wei, M.; Li, X.; Yin, L.; Sha, A.; Zhou, M.; Liu, K.; et al. iTRAQ proteomic analysis of Wheat (Triticum aestivum L.) genotypes differing in waterlogging tolerance. Front. Plant Sci. 2022, 13, 890083. [Google Scholar] [CrossRef]
- Dong, A.; Yang, Y.; Liu, S.; Zenda, T.; Liu, X.; Wang, Y.; Duan, H. Comparative proteomics analysis of two maize hybrids revealed drought-stress tolerance mechanisms. Biotechnol. Biotechnol. Equip. 2020, 34, 763–780. [Google Scholar] [CrossRef]
- Che, L.; Lu, S.; Liang, G.; Gou, H.; Li, M.; Chen, B.; Mao, J. Identification and expression analysis of the grape pentatricopeptide repeat (PPR) gene family in abiotic stress. Physiol. Mol. Biol. Plants. 2022, 28, 1849–1874. [Google Scholar] [CrossRef]
- Airaki, M.; Leterrier, M.; Mateos, R.M.; Valderrama, R.; Chaki, M.; Barroso, J.B.; Del Río, L.A.; Palma, J.M.; Corpas, F.J. Metabolism of reactive oxygen species and reactive nitrogen species in pepper (Capsicum annuum L.) plants under low temperature stress. Plant Cell Environ. 2012, 35, 281–295. [Google Scholar] [CrossRef]
- Basal, O.; Szabó, A.; Veres, S. Physiology of soybean as affected by PEG-induced drought stress. Curr. Plant Biol. 2020, 22, 100135. [Google Scholar] [CrossRef]
- Li, F.; Deng, Y.; Yamamoto, E.; Zhenya, L. Plant omics databases: An online resource guide. In Plant Omics: Advances in Big Data Biology; CABI: Wallingford, UK, 2022; pp. 253–269. [Google Scholar]
- Artimo, P.; Jonnalagedda, M.; Arnold, K. ExPASy: SIB bioinformatics resource portal. Nucleic Acids Res. 2012, 40, W597–W603. [Google Scholar] [CrossRef]
- Ayaz, A.; Huang, H.; Zheng, M.; Zaman, W.; Li, D.; Saqib, S.; Zhao, H.; Lü, S. Molecular cloning and functional analysis of GmLACS2-3 reveals its involvement in cutin and suberin biosynthesis along with abiotic stress tolerance. Int. J. Mol. Sci. 2021, 22, 9175. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.D.; Gao, T.; Chen, J.F.; Yang, J.K.; Huang, H.Y.; Yu, Y.B. The late embryogenesis abundant gene family in tea plant (Camellia sinensis): Genome-wide characterization and expression analysis in response to cold and dehydration stress. Plant Physiol. Bioch. 2018, 135, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Horton, P.; Park, K.J.; Obayashi, T.; Fujita, N.; Harada, H.; Adams-Collier, C.J.; Nakai, K. WoLF PSORT: Protein localization predictor. Nucleic Acids Res. 2007, 35, 585–587. [Google Scholar] [CrossRef] [PubMed]
- Liang, G.P.; Hou, Y.; Wang, H.; Wang, P.; Mao, J.; Hong, B. VaBAM1 weakens cold tolerance by interacting with the negative regulator VaSR1 to suppress β-amylase expression. Int. J. Biol. Macromol. 2023, 225, 1394–1404. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Che, L.; Lu, S.; Gou, H.; Li, M.; Guo, L.; Yang, J.; Mao, J. VvJAZ13 positively regulates cold tolerance in Arabidopsis and grape. Int. J. Mol. Sci. 2024, 25, 4458. [Google Scholar] [CrossRef] [PubMed]
- Hwarari, D.; Guan, Y.L.; Ahmad, B.; Movahedi, A.; Min, T.; Hao, Z.D.; Lu, Y.; Chen, J.H.; Yang, L.M. ICE-CBF-COR signaling cascade and its regulation in plants responding to cold stress. Int. J. Mol. Sci. 2022, 23, 1549. [Google Scholar] [CrossRef]
- Fraga, H.; Santos, J.A.; Moutinho-Pereira, J.; Carlos, C.; Silvestre, J.; Eiras-Dias, J.; Mota, T.; Malheiro, A.C. Statistical modelling of grapevine phenology in Portuguese wine regions: Observed trends and climate change projections. J. Agric. Sci. 2016, 154, 795–811. [Google Scholar] [CrossRef]
- Chen, L.P.; Zhao, Y.; Xu, S.J.; Zhang, Z.Y.; Xu, Y.Y.; Zhang, J.Y.; Chong, K. OsMADS57 together with OsTB1 coordinates transcription of its target OsWRKY94 and D14 to switch its organogenesis to defense for cold adaptation in rice. New Phytol. 2018, 218, 219–231. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, X.; Xu, S.; Li, Y.; Yin, Y. Expression of Cystathionine-β-Synthase (CBS) Genes in Grape under Various Abiotic Stresses and Subcellular Localization of VvCBS27. Horticulturae 2024, 10, 1009. https://doi.org/10.3390/horticulturae10091009
Shi X, Xu S, Li Y, Yin Y. Expression of Cystathionine-β-Synthase (CBS) Genes in Grape under Various Abiotic Stresses and Subcellular Localization of VvCBS27. Horticulturae. 2024; 10(9):1009. https://doi.org/10.3390/horticulturae10091009
Chicago/Turabian StyleShi, Xingyun, Shanshan Xu, Yandong Li, and Yiming Yin. 2024. "Expression of Cystathionine-β-Synthase (CBS) Genes in Grape under Various Abiotic Stresses and Subcellular Localization of VvCBS27" Horticulturae 10, no. 9: 1009. https://doi.org/10.3390/horticulturae10091009
APA StyleShi, X., Xu, S., Li, Y., & Yin, Y. (2024). Expression of Cystathionine-β-Synthase (CBS) Genes in Grape under Various Abiotic Stresses and Subcellular Localization of VvCBS27. Horticulturae, 10(9), 1009. https://doi.org/10.3390/horticulturae10091009