Abstract
The choice of training system impacts the architecture and physiological characteristics of grapevines, affecting grape production and quality. Continuous studies are necessary to optimize viticulture production in Brazil. This study aimed to evaluate the effects of rootstocks and different training systems on the production and quality of ‘IAC 138-22 Máximo’ and ‘BRS Violeta’ grapevines for juice and wine. The experiment was conducted over two productive cycles (2019/2020 and 2020/2021) in an experimental vineyard at the Advanced Center for Fruit Research as part of the Agronomic Institute (IAC), in Jundiaí, São Paulo, Brazil (23°06′ S, 46°55′ W, and 745 m altitude). For each cultivar, a randomized block design in a 2 × 2 factorial scheme was used, with two rootstocks (‘IAC 766 Campinas’ and 106-8 ‘Mgt’) and two training systems (low and high trellises), with five blocks of three plants per experimental plot. In both cycles, the gas exchange and grapevine production, the chemical characteristics of the grape juice (must), and the chemical compounds in the berry skins were evaluated. The rootstocks and training systems influenced the variables evaluated in both cultivars, with the high trellis system providing the best results. This approach increased the photosynthetic rate, improved water-use efficiency, elevated grapevine production, and enhanced fruit quality. Therefore, its use is recommended for training grapevines. Regarding rootstocks, the best scion–rootstock affinity was found between hybrid grapevines and ‘IAC 766 Campinas’.
1. Introduction
The Brazilian viticulture landscape is diverse, driven by the genetic breeding of grapevines for greater productivity and adaptation to the tropical climate [1]. In 2023, Brazil produced 1,719,630 tons of grapes over 76,747 hectares [2]. The regional diversity in national viticulture includes different production cycles, harvest times, cultivars, and management practices, requiring studies on specific cultivation techniques. The Brazilian grape market is divided between fresh consumption (table grapes) and processing (grape juice and wines), with a predominance of non-vinifera grapes (Vitis labrusca and hybrids). The diversity of cultivars results in grapes with distinct characteristics, influenced by the polyphenolic content and the search for high-quality antioxidant products.
Genetic breeding has enabled the development of hybrid cultivars, such as ‘IAC 138-22 Máximo’ and ‘BRS Violeta’ [3,4], both red and cultivated for processing juice and wines.
These cultivars are aimed at meeting the grape juice and derivatives market, characterized by high productivity and lower sensitivity to major fungal diseases compared to Vitis vinifera. However, studies are needed to understand the performance of these cultivars grafted on different rootstocks and grown using different training systems, as well as to evaluate their productive characteristics and fruit quality.
Given climate change, the selection of rootstocks and scion varieties becomes crucial for viticulture, as it allows better adaptation of the grapevines to adverse conditions and increases their resistance to abiotic stresses [5]. This strategic choice optimizes the use of water and nutrients, maintaining grape productivity and quality, which is essential for the sustainability of the viticulture sector [5].
The use of rootstocks in viticulture not only protects against damage caused by Phylloxera (Daktulosphaira vitifoliae), but also enables cultivation in situations involving different abiotic factors and adverse soil conditions (the presence of pests and high salinity), which directly affect vegetative development, the duration of the phenological phases, and production quality [6].
The interaction between scion varieties and rootstocks is widely studied under different edaphoclimatic conditions, aiming to optimize vegetative growth, productivity, and grape quality [7,8,9,10,11].
With the expansion of viticulture areas, the choice of the vineyard training system directly influences the architecture and physiological characteristics of grapevines, as well as the quantitative and qualitative aspects of the fruits [12,13]. Although the three-wire trellis system is the most commonly used, new variations have emerged to increase the leaf area, favoring physiological characteristics that increase production and improve fruit quality, especially in terms of phenolic compounds and sugars, which are fundamental to the quality of juices and wines [14].
Among the training system alternatives, there is the high trellis system, with four wires, providing a greater height for the vegetative canopy. The interaction of solar radiation with the grapevine canopy structure directly influences its productivity, affecting the light distribution, carbon assimilation, and water deficit [15]. Adequate foliage is essential for photosynthesis, without creating excessive shading that could hinder grape development and ripening [16,17]. During maturation, proper light exposure is essential for the synthesis and accumulation of anthocyanins and flavanols, which are positively related to solar radiation exposure [18,19].
Studies on training systems used in grape processing have shown significant variations in grape and wine or juice production and quality [12,14,20,21]. However, for the cultivars ‘IAC 138-22 Máximo’ and ‘BRS Violeta’, in subtropical conditions, there are no reports in the literature. Grapes are recognized for their antioxidant properties due to phenolic compounds, such as flavonoids and anthocyanins, which play important roles in the human diet and in the prevention of cardiovascular, cancerous, and neurological diseases [22,23,24]. In addition to improving nutritional quality, these compounds enhance the commercial quality of red grapes, being fundamental for their color and external appearance [25].
In this context, the present work aimed to evaluate the effects of interactions between rootstocks (‘IAC 766 Campinas’ and 106-8 ‘Mgt’) and training systems (low and high trellises), focusing on improving the productivity and quality of ‘IAC 138-22 Máximo’ and ‘BRS Violeta’ grapevine fruits.
2. Materials and Methods
2.1. Installation and Vineyard Management, and Location of the Experimental Area
The experiment was conducted over two productive cycles (2019/2020 and 2020/2021) in a vineyard in its 9th and 10th year of production, located at the Advanced Center for Fruit Research as part of the Agronomic Institute (IAC), Jundiaí, São Paulo, Brazil (23°06′ S, 46°55′ W, with an average altitude of 745 m). According to the Köppen classification, the region’s climate is classified as Cfb, with an average annual rainfall of 1400 mm, an average annual temperature of 19.5 °C, and a relative humidity of 70.6%. The soil in the experimental area is classified as Dystrophic Red Cambisol [26].
In both experimental years, short pruning was performed, with one bud, followed by the application of 5% hydrogen cyanamide (Dormex®, BASF, Ludwigshafen am Rhein, Germany). After budburst, only one productive shoot per spur was maintained, with disbudding and tying of the shoots to wires, thinning, and defoliation. Additionally, cultural practices, such as weeding, and applications of herbicides, fungicides, and insecticides, were carried out.
2.2. Experimental Design and Treatments
The cultivars ‘IAC 138-22 Máximo’ (‘Seibel 11342’ × ‘Syrah’) and ‘BRS Violeta’ (‘BRS Rúbea’ × ‘IAC 1398-21’) were evaluated over two productive cycles in an experimental vineyard located at the Advanced Center for Fruit Research as part of the Agronomic Institute (IAC), Jundiaí, São Paulo, Brazil. For each cultivar, a randomized block design was used in a 2 × 2 factorial arrangement, with two rootstocks (‘IAC 766 Campinas’ and 106-8 ‘Mgt’) and two training systems: low trellis with three wires situated at 1, 1.3, and 1.6 m from the ground, and high trellis with four wires situated at 1, 1.3, 1.6, and 2 m from the ground, with five blocks and three plants per experimental plot.
2.3. Harvest and Evaluations
Harvests were performed according to the physiological maturation of each cultivar, occurring at 140 days for ‘BRS Violeta’ and 146 days for ‘IAC 138-22 Máximo’ in the first cycle, and at 153 days for both cultivars in the second cycle (Figure 1).
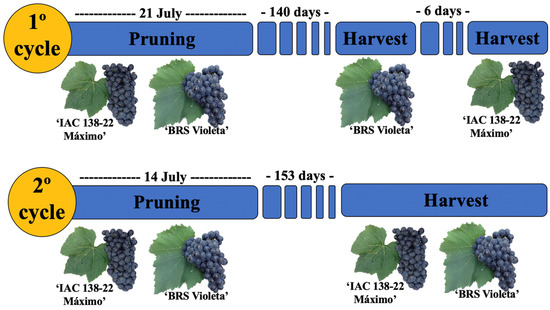
Figure 1.
Pruning and harvest times for Brazilian hybrid grapes in two productive cycles, in Jundiaí, São Paulo, Brazil.
2.4. Gas Exchange, Water-Use Efficiency, SPAD Index, and Total Chlorophyll
The gas exchange was evaluated at the full flowering stage of the vines, using an open photosynthesis system, with an infrared CO2 and water vapor analyzer (Infrared Gas Analyzer—IRGA, model LI-6400, Li-Cor). During the evaluations, the ambient CO2 concentration was used as a reference, along with data collection on the temperature and relative humidity. The CO2 assimilation rate (A, μmol CO2 m−2 s−1), transpiration rate (E, mmol H2O m−2 s−1), internal CO2 concentration (Ci), and stomatal conductance (gs) were calculated according to the method proposed by [27].
The water-use efficiency (WUE, μmol CO2 (mmol H2O)−1) was determined by the ratio of CO2 assimilation to the transpiration rate. The electron transport rate (ETR) was calculated following the method by [28]. The SPAD index was evaluated on four leaves per plant, with readings taken at three points per leaf using a SPAD meter (Model 502, Konica Minolta, Tokyo, Japan). The total chlorophyll concentration was determined using 100 g of fresh leaf material [29].
2.5. Number of Bunches per Plant, Productivity, and Physicochemical Composition of Grape Must
After harvesting, the grape bunches were counted and weighed to determine the number of bunches per plant, the fresh mass of the bunches, and the yield per plant. Subsequently, based on the number and fresh mass of the bunches and the cultivation stand, the productivity (t ha−1) was estimated.
For the physicochemical analyses of grape must, 250 berries per experimental plot (n = 250) were used. The must was obtained by pressing the berries, and the soluble solids (SS) content was evaluated by direct refractometry, using an Atago® digital refractometer (Schmidt Haensch, Berlin, Germany) with automatic temperature compensation, with results expressed in °Brix. The pH was measured by a direct reading of the must using a Micronal B-274 pH meter (Mettler Toledo, Barueri, Brazil). The titratable acidity (TA) was determined by titration, using 5 g of must, diluted in 100 mL of distilled water, titrating with a standardized 0.1 N sodium hydroxide solution, using phenolphthalein as an indicator, until the color change endpoint, with results expressed as a percentage of tartaric acid. The reducing sugars were determined by the colorimetric method of Somogyi-Nelson, based on a standard glucose curve and readings taken at 510 nm in a BEL Photonics® SP 2000 UV/vis spectrophotometer [30], with the results expressed as a percentage of reducing sugars per mL of must.
2.6. Biochemical Composition of Grape Skins
For the analyses of the total phenolic compounds [31] and total monomeric anthocyanins [32], as well as the antioxidant activity via DPPH (2,2-diphenyl-1-picrylhydrazyl) [33], modified by [34] and FRAP [35], 200 berries per experimental plot (n = 1000 berries per treatment) were collected. The berries were halved, the seeds were removed, and the skins and pulp were separated and frozen in liquid nitrogen. Approximately 100 g of frozen fresh skin was manually pulverized in a porcelain mortar with a pestle and stored at −80 °C until the analyses. All the analyses were performed in triplicate.
2.7. Statistical Analysis
The means obtained from the two production cycles for each cultivar were subjected to analysis of variance (ANOVA) to determine the effects of the rootstocks and training systems and their interactions. Subsequently, the means were compared using the Tukey test (p < 0.05), utilizing the Sisvar 6.0 software [36]. Additionally, to characterize the interaction between the rootstocks and the training systems and to assess the correlation of the variables, principal component analysis (PCA) was performed using Statistical Analysis Software 4.0 (SAS).
3. Results
3.1. Impact of Rootstocks and Training Systems on ‘IAC 138-22 Maximo’ Grape Variety: Physiological, Biochemical Parameters and Yield
Significant interactions were observed between rootstocks and training systems concerning the electron transport rate (ETR), stomatal conductance (gs), transpiration rate (E), water-use efficiency (WUE), assimilation rate (A), and internal carbon concentration (Ci) in the ‘IAC 138-22 Máximo’ grape variety (Table 1).

Table 1.
Electron transport rate (ETR), stomatal conductance (gs), transpiration rate (E), water-use efficiency (WUE), CO2 assimilation rate (A), and internal carbon concentration (Ci) of the ‘IAC 138-22 Máximo’ vine on different rootstocks and training systems.
A higher ETR was achieved with the combination of the rootstock 106-8 ‘Mgt’ and the high espalier, as well as with the rootstock ‘IAC 766 Campinas’ and the low espalier. This suggests that these specific combinations are more efficient in capturing and utilizing light energy for electron transport, leading to increased production of ATP and NADPH, which are essential for the synthesis of photoassimilates.
Stomatal conductance (gs) is another vital parameter, referring to the rate of CO2 influx and water vapor efflux through the stomata. An increase in gs, as observed in vines grafted onto the rootstock 106-8 ‘Mgt’ with the low espalier training system, allows for greater CO2 intake, thereby increasing the assimilation rate (A). It was also noted that the transpiration rate (E), which measures water loss through the stomata, increased as well, potentially aiding in maintaining leaf temperature and ensuring continuous water and nutrient flow.
The increased gs and E in vines grafted onto the rootstock 106-8 ‘Mgt’ may have contributed to the lower water-use efficiency (WUE) in this combination of rootstock and training system, indicating that, despite the increased photosynthesis, water was not used as efficiently, which may result in lower biomass accumulation.
In both training systems, vines grafted onto the rootstock ‘IAC 766 Campinas’ exhibited a higher internal carbon concentration (Ci), suggesting that despite the high stomatal conductance (gs), CO2 fixation may not have been as efficient due to lower Rubisco activity and reduced water-use efficiency (WUE). Another hypothesis is that mesophyll cells consume CO2 during photosynthetic assimilation, consequently resulting in lower CO2 concentration in the intercellular airspace compared to the ambient air outside the leaf. The internal carbon concentration (Ci) reflects the concentration of CO2 within the leaves and indicates the balance between CO2 fixation and its entry through the stomata.
Thus, it was observed that the correct choice of rootstock and training system can optimize photosynthesis and water use in vines, resulting in more efficient and productive plants. These interactions directly influence physiological parameters, such as the ETR, gs, E, WUE, A, and Ci, and consequently, the agronomic performance of the ‘IAC 138-22 Máximo’ vines.
No interaction was observed between the rootstocks and training systems for the SPAD index and total chlorophyll content in the ‘IAC 138-22 Máximo’ vine, with only rootstocks showing a significant isolated effect. Higher SPAD indices and chlorophyll contents were obtained with the use of the rootstock 106-8 ‘Mgt’ (Table 2). Vines with higher SPAD indices exhibit greater green coloration intensity, as a result of higher concentrations of photosynthetic pigments, particularly chlorophylls. This aspect is particularly important for grapes intended for processing, where a high SPAD index and chlorophyll content enhance light utilization during photosynthesis, increasing carbohydrate accumulation, which will be converted into sugars in the berries and energy for the next production cycle.
Table 2.
SPAD index and total chlorophyll of the ‘IAC 138-22 Máximo’ vine on different rootstocks and training systems.
Associating these results with photosynthetic parameters, it can be inferred that vines with a higher chlorophyll content and SPAD index, as observed with the rootstock 106-8 ‘Mgt’, exhibit greater photosynthetic efficiency. This is reflected in a higher ETR and CO2 assimilation rate (A), promoting better photosynthetic performance.
No significant interaction was observed between rootstocks and training systems for the productive and physicochemical variables of the ‘IAC 138-22 Máximo’ grape must. Therefore, the isolated effect of the variables was assessed. Significant differences were found for the production, yield, cluster fresh weight, soluble solids, and pH, among the training systems. Except for pH, all the variables showed higher results when the ‘IAC 138-22 Máximo’ vine was trained with a high espalier (Table 3).
Table 3.
Number and fresh weight of clusters, yield, productivity, soluble solids, pH, titratable acidity (TA), SS/TA ratio, and reducing sugars of the ‘IAC 138-22 Máximo’ vine on different rootstocks and training systems.
The yield of the vines trained with a high espalier was 38.52% higher than those trained with a low espalier, demonstrating greater compatibility of the cultivar with this training system, possibly due to its high vigor. The ‘IAC 138-22 Máximo’ vine requires a system that allows for better distribution of the branches and the vegetative canopy, thus providing more efficient leaf distribution and greater solar radiation capture. This benefits the photosynthetic process, increasing photoassimilate production and, consequently, the yield. Additionally, this training system resulted in heavier clusters, which directly contributes to the observed higher yield.
The high espalier system promotes an increase in the soluble solids in the must of the ‘IAC 138-22 Máximo’ grape (15.25 °Brix). Although the training system did not influence the titratable acidity and the SS/TA ratio, the low espalier system produced fruit with an acidity of 0.98% tartaric acid and an SS/TA ratio of 14.91, values below the requirements in Brazilian legislation, which stipulates a maximum acidity of 0.9% tartaric acid and a minimum SS/TA ratio of 15 for grapes intended for processing. This result was also observed when the vines were grafted onto the rootstock 106-8 ‘Mgt’, which had a titratable acidity of 1.02% tartaric acid and an SS/TA ratio of 14.17.
When a grape cultivar exhibits high acidity, low soluble solids content, or any other undesirable chemical characteristic, it becomes crucial to diversify cultivars for blending purposes. This aims to balance the potential limitations of each cultivar, thus meeting the required standards for beverage production and enhancing the quality of juices and wines.
The high espalier system, by providing better canopy distribution, supports a higher ETR and assimilation rate, optimizing light utilization and increasing ATP and NADPH production. This not only improves photosynthesis, but also enhances the efficiency of photoassimilate production, resulting in a higher soluble solids content and better grape must quality. Therefore, choosing an appropriate training system, such as the high espalier, in combination with efficient rootstocks, is crucial for maximizing the productivity and quality of the ‘IAC 138-22 Máximo’ vine.
A well-distributed canopy allows more leaves to receive direct light, increasing the CO2 assimilation rate and light-use efficiency. Increased photosynthetic activity promotes a higher concentration of carbohydrates, which are utilized for the growth and development of the clusters, explaining the greater cluster fresh weight and higher soluble solids concentration. Thus, the high espalier offers optimal conditions to maximize the photosynthetic efficiency and productivity of the ‘IAC 138-22 Máximo’ vine.
The rootstock ‘IAC 766 Campinas’ provided the ‘IAC 138-22 Máximo’ grape must with lower titratable acidity, increasing the SS/TA ratio and improving the grape flavor.
Improvements to the photosynthetic efficiency and carbohydrate production of grapevines are essential for fruit growth and development. Consequently, the increased absorption and availability of nutrients, common to vigorous rootstocks, may lead to reduced titratable acidity, balancing the SS/TA ratio and enhancing the flavor and quality of the must.
Moreover, the greater vigor of the ‘IAC 766 Campinas’ rootstock may be associated with its better adaptation to various environmental and soil conditions, promoting more balanced and healthy vine growth. This results in a more efficient distribution of resources within the plant, contributing to a more uniform development of the aerial parts and, particularly, the clusters, leading to better overall fruit quality.
Significant interactions were observed between rootstocks and training systems for the total phenolic compounds, monomeric anthocyanins, and antioxidant activity (DPPH and FRAP) of the ‘IAC 138-22 Máximo’ grapevine (Table 4). The combination of the high espalier with the ‘IAC 766 Campinas’ rootstock resulted in higher levels of monomeric anthocyanins and total phenolic compounds in the grapes, increasing the antioxidant activity as expressed by DPPH and FRAP values (Table 4).
Table 4.
Total phenolic compounds, total monomeric anthocyanins, and antioxidant activity (DPPH and FRAP) in the skin of ‘IAC 138-22 Máximo’ grapes on different rootstocks and training systems.
The presence of high levels of phenolic compounds, anthocyanins, and antioxidant activity in the berry skins of ‘IAC 138-22 Máximo’ grapes, especially when combined with the ‘IAC 766 Campinas’ rootstock and the high espalier system, offers numerous health benefits. Anthocyanins and flavonoids are positively related to vine exposure to solar radiation, which justifies the high antioxidant activity observed in grapes cultivated with this combination of training system and rootstock.
The high antioxidant activity observed, as measured by the DPPH and FRAP methods, indicates that these grapes have a significant capacity to combat cellular damage caused by reactive oxygen species.
The higher concentration of these compounds in the combination of the high espalier and the ‘IAC 766 Campinas’ rootstock suggests an increased synthesis of flavonoids.
The combination of the high espalier and the ‘IAC 766 Campinas’ rootstock proved effective in maximizing the levels of these bioactive compounds in the grape skins. This training system and rootstock favor light exposure and canopy distribution, optimizing photosynthesis [14] and secondary metabolite production.
Consequently, the increase in phenolic compounds and anthocyanins not only improves the functional and sensory quality of the grapes, but also enhances the health benefits, making these grapes highly beneficial for both fresh consumption and for the production of juices and wines. Therefore, the choice of appropriate training systems and rootstocks is crucial for maximizing the beneficial chemical and biochemical attributes of ‘IAC 138-22 Máximo’ grapes for consumers.
3.2. Impact of Rootstock and Training Systems on ‘BRS Violeta’ Grape Variety: Physiological, Biochemical Parameters and Yield
For the variables, namely the electron transport rate (ETR), stomatal conductance (gs), CO2 assimilation rate (A), internal carbon concentration (Ci), and SPAD index of the ‘BRS Violeta’ vine, a significant interaction between the rootstocks and training systems was observed (Table 5). The combination of ‘IAC 766 Campinas’ with the low espalier resulted in reduced energy dissipation and a higher ETR, indicating a less stressful condition for the scion cultivar and resulting in less wear on the vines. This combination also increased the CO2 assimilation rate and reduced the internal carbon concentration, reinforcing the benefits of this combination for vine cultivation (Table 5).
Table 5.
Electron transport rate (ETR), stomatal conductance (gs), assimilation rate (A), internal carbon concentration (Ci), and SPAD index of the ‘BRS Violeta’ vine on different rootstocks and training systems.
The lower energy dissipation observed with the combination of ‘IAC 766 Campinas’ and a low espalier suggests that this interaction optimizes the capture of light energy, directing it more efficiently towards photosynthetic processes.
Furthermore, the reduction in the internal carbon concentration (Ci) observed with this interaction suggests that the available CO2 is rapidly fixed during photosynthesis, preventing CO2 accumulation in leaf cells and potentially reducing oxidative stress [27,28].
For the SPAD index, a significant interaction was observed, with the ‘IAC 766 Campinas’ rootstock providing higher SPAD values in ‘BRS Violeta’ vines, regardless of whether it was combined with a low or high espalier. The SPAD index is an indirect indicator of chlorophyll content, with high SPAD values being associated with greater photosynthetic capacity of the plant.
The presence of higher chlorophyll content suggests that the ‘IAC 766 Campinas’ rootstock is particularly effective in enhancing the photosynthetic efficiency of ‘BRS Violeta’, providing optimal conditions for the production of ATP and NADPH, necessary for CO2 assimilation.
The combination of the ‘IAC 766 Campinas’ rootstock with both training systems (low or high espalier) maximizes the SPAD index, contributing to a higher ETR, gs, and A. The increased ETR facilitates electron transfer between the photosystems, while a higher gs enhances the gas exchange and CO2 uptake. The combination of these physiological and biochemical characteristics optimizes photosynthesis, promoting more vigorous growth of ‘BRS Violeta’ vines.
There were no significant interactions between the training systems and rootstocks in terms of the transpiration rate (E), water-use efficiency (WUE), and chlorophyll content; therefore, these variables were analyzed separately (Table 6). For the photosynthetic pigments, no significant differences were observed in any of the evaluated variables.
Table 6.
Transpiration rate (E), water-use efficiency (WUE), and total chlorophyll of the ‘BRS Violeta’ vine on different rootstocks and training systems.
The ‘BRS Violeta’ hybrid grown with a low espalier has higher water-use efficiency (WUE), particularly when combined with the 106-8 ‘Mgt’ rootstock, which also resulted in a lower transpiration rate (E). Since the WUE indicates the plant’s capacity to assimilate a greater amount of carbon dioxide with less water lost through transpiration, a higher water-use efficiency implies a lower E, which can contribute to better carbohydrate synthesis efficiency and reduced vulnerability to water stress.
The increased WUE observed with the low espalier and the 106-8 ‘Mgt’ rootstock suggests an optimization in the plant’s water balance, allowing efficient CO2 assimilation without excessive water loss through transpiration (E). This trait is particularly advantageous in environments with limited water resources, where the ability to maintain efficient photosynthesis and carbohydrate production with reduced water consumption is crucial. Therefore, selecting training systems and rootstocks that maximize the WUE can significantly improve the sustainability and productivity of the ‘BRS Violeta’ vine under water stress conditions.
No significant interactions were observed between rootstocks and training systems for any of the productive or physicochemical variables of the ‘BRS Violeta’ grape must. Therefore, the factors were evaluated individually (Table 7). Training systems only influenced the soluble solids (SS) content and titratable acidity (TA) of the must.
Table 7.
Number and fresh weight of the clusters, yield, productivity, soluble solids, pH, titratable acidity (TA), SS/TA ratio, and reducing sugars of the ‘BRS Violeta’ vine on the different rootstocks and training systems.
The highest soluble solids (SS) content (16.32 °Brix) and the lowest titratable acidity (TA) (0.68% tartaric acid) were obtained from grapes of vines trained on a high espalier. The SS content in grapes is primarily composed of sugars (glucose and fructose) and, along with the TA, are direct indicators of fruit quality, closely linked to photosynthetic metabolism and the plant’s resource-use efficiency. The higher SS content achieved with a high espalier suggests better photosynthetic efficiency, as the high espalier configuration allows for improved light distribution over the canopy, enhancing light capture and CO2 assimilation (A). This efficiency is highlighted by the elevated values of the electron transport rate (ETR), stomatal conductance (gs), and SPAD index, which together contribute to increased sugar production in the fruits (Table 5).
The lower TA observed in grapes from a high espalier can be explained by the increased CO2 assimilation and greater Rubisco enzyme activity, which channels more carbon into sugar production, reducing the relative concentration of organic acids in the must. This combination is ideal for producing grapes with a more balanced and attractive sensory profile for winemaking.
The ‘IAC 766 Campinas’ rootstock was noted for promoting higher production and productivity in the ‘BRS Violeta’ vine. The increase in these variables can be attributed to several physiological advantages provided by this rootstock; hence, it is recognized for its superior vigor, translating into greater water and nutrient absorption capacity from the soil. This is crucial to meet the high metabolic demands associated with photosynthesis and plant growth.
Thus, the greater photosynthetic efficiency of vines grafted onto ‘IAC 766 Campinas’ is evidenced by the higher values of the ETR, gs, and CO2 assimilation rate (A) (Table 5). These parameters indicate that the vines have a greater capacity to capture and utilize solar light for photosynthesis, resulting in higher production of ATP and NADPH, which are essential for carbohydrate synthesis and increased vine production.
Vines grafted onto this rootstock showed higher SS and pH values, indicating an optimal balance between sugar accumulation and must pH. The superior capacity of ‘IAC 766 Campinas’ to support cultivation conditions and maximize carbohydrate production makes it a recommended choice for the ‘BRS Violeta’ vine. The higher CO2 assimilation rate (A) observed with the use of this rootstock indicates that more carbon is being fixed and converted into sugars and other photoassimilates, which are used for fruit growth and development.
Moreover, the improved regulation of must pH with ‘IAC 766 Campinas’ indicates a balance between sugar synthesis and organic acid accumulation, resulting in higher quality fruit for wine and juice production. Specifically concerning pH, studies suggest that the effects of rootstocks may be related to their capacity for potassium extraction from the soil.
In both grapes and beverages, such as grape juice and wine, pH measurement is crucial as it is directly related to anthocyanin stability, which affects the color intensity of these beverages. Therefore, the choice of rootstock not only impacts vine productivity and health, but also the final quality of the grape-derived products, highlighting the importance of ‘IAC 766 Campinas’ in optimizing the production and quality of ‘BRS Violeta’ grapes.
There were interactions among the factors evaluated for the secondary metabolites in the skin of ‘BRS Violeta’ grapes. The combination of using the ‘IAC 766 Campinas’ rootstock with the high espalier resulted in higher levels of total phenolic compounds and monomeric anthocyanins, as well as greater antioxidant activity, using both methods (Table 8).
Table 8.
Total phenolic compounds, total monomeric anthocyanins, and antioxidant activity (DPPH and FRAP) in the skin of ‘BRS Violeta’ grape in different rootstocks and training systems.
The higher concentration of phenolic compounds obtained with the high espalier can be attributed to better sunlight distribution, which promotes increased photosynthetic activity and synthesis of secondary metabolites, such as phenolic compounds.
This same interaction between the rootstock and the training system also resulted in increased anthocyanin content in the grape skins (Table 8). Biochemically, anthocyanins are pigments responsible for grape coloration and play a significant role in antioxidant activity.
Greater light exposure with a high espalier can induce the expression of genes related to anthocyanin biosynthesis, resulting in higher concentrations of these pigments. Enhanced photosynthetic efficiency, evidenced by the high ETR and increased production of ATP and NADPH (Table 5), also contributes to the increased synthesis of anthocyanins.
Anthocyanins, along with other phenolic components present in ‘BRS Violeta’ grapes, are of extreme importance in the production of red wines and, particularly, juices. Therefore, the correct choice of training system and rootstock, which have greater affinity with the ‘BRS Violeta’ cultivar, is essential to maximize the accumulation potential of antioxidant compounds in grapes.
Vines trained with a high espalier and grafted onto the ‘IAC 766 Campinas’ rootstock exhibited higher antioxidant activity (DPPH: 639.87 µmol g−1 of grape skin; and FRAP: 43.84 µmol Fe kg−1 of grape skin). These results indicate that the increased synthesis of phenolic compounds and anthocyanins as a result of a high espalier not only enhances the nutritional quality of the grapes, but also boosts their antioxidant capacity.
Therefore, the choice of training system and rootstock has a significant impact on the phytochemical composition and antioxidant activity of ‘BRS Violeta’ grapes. The high espalier, combined with the ‘IAC 766 Campinas’ rootstock, proved to be the most effective combination for maximizing the concentration of phenolic compounds, anthocyanins, and antioxidant activity. This combination not only improves the nutritional and sensory quality of the grapes, but also enhances the photosynthetic efficiency and resistance to environmental stresses, promoting more sustainable and high-quality production.
3.3. Principal Component Analysis (PCA)
Principal Component Analysis (PCA) of the ‘IAC 138-22 Máximo’ vine reveals that the first two principal components (PCA 1 and PCA 2, Figure 2A) account for 85.03% of the total variance in the data. It is observed that parameters related to photosynthetic efficiency, such as the SPAD index, internal carbon concentration (Ci), assimilation rate (A), and electron transport rate (ETR), are positively correlated and associated with the ‘IAC 766 Campinas’ rootstock and the high espalier. This indicates that this combination promotes better conditions for photosynthesis, resulting in higher production of photoassimilates and higher levels of phenolic compounds and anthocyanins, which are crucial for the nutritional and sensory quality of the grapes and wines.
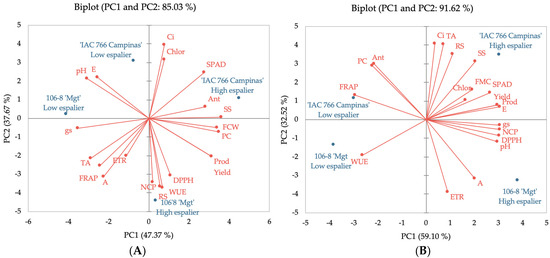
Figure 2.
Principal component analysis of hybrid vines on different rootstocks and training systems. (A) ‘IAC 138-22 Máximo’ and (B) ‘BRS Violeta’. Notes: internal carbon concentration (Ci), total chlorophyll (Chlor), anthocyanins (Ant), soluble solids (SS), fresh cluster weight (FCW), phenolic compounds (PC), productivity (Prod), water-use efficiency (WUE), reducing sugars (RS), number of clusters per plant (NCP), electron transport rate (ETR), assimilation rate (A), titratable acidity (TA), stomatal conductance (gs), transpiration rate (E).
Parameters, such as the pH, yield, productivity, and soluble solids, are also positively correlated with a high espalier, highlighting the importance of this training system in maximizing the quality of grape must. The 106-8 ‘Mgt’ rootstock, associated with the low espalier, showed lower efficiency in regard to all these parameters, suggesting reduced physiological and biochemical performance of the ‘IAC 138-22 Máximo’ vines under these conditions.
In the PCA of the ‘BRS Violeta’ vine (Figure 2B), the first two principal components explain 91.62% of the total variance in the data. Similar to ‘IAC 138-22 Máximo’, the photosynthetic efficiency parameters (SPAD, Ci, A, ETR) are strongly associated with the ‘IAC 766 Campinas’ rootstock combined with the high espalier. This rootstock demonstrated notable superiority in maximizing the photosynthetic efficiency and, consequently, the production of phenolic compounds, anthocyanins, and antioxidant activity.
Additionally, must quality parameters, such as the soluble solids (SS) content, yield, and titratable acidity (TA) are positively correlated with the high espalier and the ‘IAC 766 Campinas’ rootstock. This indicates that this combination not only improves the photosynthetic efficiency, but also optimizes the physical and chemical qualities of the grapes. In contrast, the low espalier and the 106-8 ‘Mgt’ rootstock are associated with lower levels of phenolic compounds, anthocyanins, and antioxidant activity (DPPH and FRAP), reflecting the reduced physiological and biochemical performance of the grapes from this combination.
Figure 3 summarizes how different combinations of rootstocks and training systems affect the production and quality of the grapes from the ‘IAC 138-22 Máximo’ and ‘BRS Violeta’ cultivars. The use of the ‘IAC 766 Campinas’ rootstock and the high espalier resulted in increased productivity and improved physiological, physicochemical, and biochemical attributes of the grapes. These results are explained by the higher capacity for water and nutrient absorption, the vigor provided by the rootstocks, and the better light distribution afforded by the high espalier. The integration of these practices can help balance the limitations of each variety, meeting the standards required for juices and wines, and resulting in higher quality products.
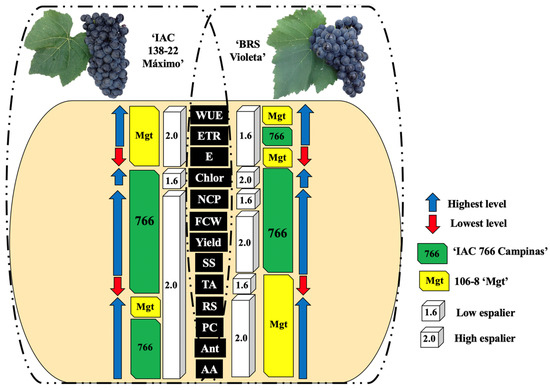
Figure 3.
Graphical abstract of the two grapevine hybrids, ‘IAC 138-22 Maximo’ and ‘BRS Violeta’ and their interaction with the two trellis systems (low 1.6 m and high 2.0 m) and two rootstocks, ‘IAC 766’ (766) and ‘106-8 Mgt’ (Mgt). Notes: water-use efficiency (WUE), electron transport rate (ETR), transpiration rate (E), total chlorophyll (Chlor), number of clusters per plant (NCP), fresh cluster weight (FCW), soluble solids (SS), titratable acidity (TA), reducing sugars (RS), phenolic compounds (PC), total monomeric anthocyanins (Ant), and antioxidant activity (AA).
In summary, the results indicate that the choice of training system and rootstock has a significant impact on the physiology, biochemistry, and final quality of the grapes from the hybrid vines ‘IAC 138-22 Máximo’ and ‘BRS Violeta’. The combination of the high espalier with the ‘IAC 766 Campinas’ rootstock provides the best conditions for maximizing the photosynthetic efficiency and the synthesis of essential biochemical compounds, resulting in grapes of high nutritional and sensory quality. Therefore, this combination is recommended for the optimized production of these hybrids.
4. Discussion
The electron transport rate (ETR) is a fundamental indicator of photosynthetic performance, reflecting the plant’s capacity to transport electrons through the electron transport chain in chloroplasts. This transport is crucial for ATP and NADPH production, which are utilized in the Calvin–Benson cycle for carbon fixation [37,38]. CO2 enters the leaves by diffusing through the stomatal pores on the leaf surface, becoming available as a substrate for photosynthetic assimilation [39]. The sensitivity of vine leaves to solar exposure and water use by plants means that the canopy structure dimensions, geometric forms, and management practices are critical for achieving optimal physiological functioning [40]. Additionally, plants with higher chlorophyll content have a greater capacity to capture solar radiation, which, combined with optimized stomatal conductance (gs) and a higher transpiration rate (E), contributes to increased production of ATP and NADPH, essential for the Calvin–Benson cycle and the production of photoassimilates [37,38].
Physiologically, the improved leaf distribution with a high espalier maximizes solar light interception, which is essential for photosynthesis [14]. These results may be related to the interaction between the scion cultivar and the rootstock, as well as to the different vigor levels provided by the rootstocks to the scion cultivars, since the rootstock ‘IAC 766 Campinas’ is more vigorous than 106-8 ‘Mgt’ [41,42,43]. The use of more vigorous rootstocks, such as ‘IAC 766 Campinas,’ contributes to an increased capacity for water and nutrient absorption and translocation by the roots [44,45], which may result in a greater supply of resources for photosynthesis and plant metabolism. Biochemically, phenolic compounds and monomeric anthocyanins are important secondary metabolites present in grapes, playing crucial roles in the defense against environmental stresses and in the nutritional and sensory quality of the fruit. Phenolic compounds are known for their anti-inflammatory, anticancer, and cardioprotective properties [46,47]. Anthocyanins, in addition to providing the characteristic color to grapes, play an important role in the protection against chronic diseases due to their ability to neutralize free radicals and reduce oxidative stress in the body [48,49,50,51]. Flavonoids are derived from the shikimic and phenolic acid pathways [52].
The high espalier and ‘IAC 766 Campinas’ rootstock favor light exposure and canopy distribution, optimizing photosynthesis [14] and secondary metabolite production. The higher ETR indicates a more efficient transfer of electrons between the photosystems, leading to increased production of ATP and NADPH, which are essential for CO2 assimilation in the Calvin cycle [37,38]. Increased stomatal conductance (gs) enhances gas exchange, increasing CO2 uptake and facilitating a higher assimilation rate (A), which translates into greater photoassimilate production and plant growth [15,19]. The reduction in the internal carbon concentration (Ci) suggests that the available CO2 is rapidly fixed during photosynthesis, preventing CO2 accumulation in leaf cells and potentially reducing oxidative stress [53]. The SPAD index is an indirect indicator of chlorophyll content; high SPAD values are associated with greater photosynthetic capacity of the plant [54]. A higher assimilation rate (A) indicates a greater photosynthetic rate, resulting in increased carbohydrate production and plant growth [54]. Water-use efficiency (WUE), indicates the plant’s capacity to assimilate a greater amount of carbon dioxide with less water lost through transpiration (E) [55]. An optimization in the plant’s water balance allows efficient CO2 assimilation, without excessive water loss through transpiration [55]. The high espalier configuration allows for improved light distribution over the canopy, enhancing light capture and CO2 assimilation (A) [14].
‘IAC 766 Campinas’ is recognized by its strong vigor, translating into greater water and nutrient absorption capacity from the soil [41,42,43,56]. Higher photosynthetic efficiency is directly related to increased sugar production, which constitutes the main components of soluble solids [57].
The greater the affinity of the rootstock for potassium, the higher the pH of the grapes [58]. ‘IAC 766 Campinas’ rootstock has a higher affinity for this nutrient. The effect of rootstocks on the physicochemical composition of grapes may also be related to other factors such as vigor, water and nutrient absorption capacity, disease resistance, and interaction with the scion. These factors directly influence both primary and secondary plant metabolites and, consequently, grape quality [59,60,61].
Anthocyanin stability affects the color intensity of grapes, grape juice, and wine [62]. Anthocyanins are pigments responsible for grape coloration and play a significant role in antioxidant activity [48,49,50,51]. Greater light exposure with a high espalier induces the expression of genes related to anthocyanin biosynthesis, resulting in higher concentrations of these pigments [63]. Phenolic compounds are essential in defending plants against environmental stress, while also contributing significantly to the grapes’ nutritional and sensory qualities [63]. The more vigorous ‘IAC 766 Campinas’ rootstock may enhance nutrient absorption, which in turn could lead to increased synthesis of these compounds [59,60,61]. The phenolic profile of grapes has garnered significant interest from researchers focused on wine and juice production [4,64,65,66]. These compounds, including anthocyanins, are renowned for their health benefits, especially in the prevention of chronic diseases. They act by neutralizing free radicals, namely unstable molecules that can damage cells and are linked to cardiovascular diseases, cancer, and neurodegenerative disorders [67]. Furthermore, the antioxidant properties of these compounds protect plant cells from damage by reactive oxygen species (ROS), enhancing the resistance to abiotic stress and promoting overall plant health [53]. Beyond mitigating oxidative stress, these compounds also improve immune function, reduce inflammation, and safeguard DNA from mutations [63].
The combination of the ‘IAC 766 Campinas’ rootstock with a high espalier is the most effective strategy for maximizing productivity, nutritional, and sensory quality of ‘IAC 138-22 Máximo’ and ‘BRS Violeta’ grapes. This approach simultaneously enhances the photosynthetic efficiency and the concentration of bioactive compounds, making it the ideal choice for viticulturists and the processing industry.
Author Contributions
F.J.D.N., J.D.R. and M.A.T. planned and designed the experiment. F.J.D.N. and C.V.B. performed the plant physiological analyses, chemical, biochemical and enzyme analyses. F.J.D.N., M.F.M. and J.L.H. performed data analyses. F.J.D.N., M.d.S.S. and M.L. created the tables and figures. F.J.D.N., J.D.R., E.O.O., G.P.P.L. and M.d.S.S. wrote and revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Fundação de Amparo à Pesquisa do Estado de São Paulo (Research Support Foundation of the State of São Paulo), FAPESP process no. 2016/07510-2 through granting the scholarship for the first author, several programs that supported the study in setting up the experimental area (FAPESP process no. 2011/03440-6, 2013/8915-5 and 2015/16440-5) and the CNPq for the Research Productivity Scholarship (process no. 307377/2021-0).
Data Availability Statement
The original contributions presented in the study are included in the article material, further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Camargo, U.; Tonietto, J.; Hoffmann, A. Advances in grape culture in Brazil. Rev. Bras. Frutic. 2011, 33, 144–149. [Google Scholar] [CrossRef]
- IBGE—Instituto Brasileiro de Geografia e Estatística. Fruticultura: Área Plantada e Quantidade Produzida. 2024. Available online: http://www.ibge.gov.br (accessed on 15 August 2024).
- Burin, V.M.; Ferreira-Lima, N.; Panceri, C.P.; Bordignon-Luiz, M.T. Bioactive compounds and antioxidant activity of Vitis vinifera and Vitis labrusca grapes: Evaluation of different extraction methods. Microchem. J. 2014, 114, 155–163. [Google Scholar] [CrossRef]
- Lago-Vanzela, E.S.; Procópio, D.P.; Fontes, E.A.F.; Ramos, A.M.; Stringheta, P.C.; Da-Silva, R.; Castillo-Muñoz, N.; Hermosín-Gutiérrez, I. Aging of red wines made from hybrid grape cv. BRS Violeta: Effects of accelerated aging conditions on phenolic composition, color and antioxidant activity. Food Res. Int. 2014, 56, 182–189. [Google Scholar] [CrossRef]
- Santos, J.A.; Fraga, H.; Malheiro, A.C.; Moutinho-Pereira, J.; Dinis, L.-T.; Correia, C.; Moriondo, M.; Leolini, L.; Dibari, C.; Costafreda-Aumedes, S.; et al. A Review of the Potential Climate Change Impacts and Adaptation Options for European Viticulture. Appl. Sci. 2020, 10, 3092. [Google Scholar] [CrossRef]
- Silva, M.J.R.; Paiva, A.P.M.; Pimentel, A.; Sánchez, C.A.P.C.; Callili, D.; Moura, M.; Leonel, S.; Tecchio, M.A. Yield performance of new juice grape varieties grafted onto different rootstocks under tropical conditions. Sci. Hortic. 2018, 241, 194–200. [Google Scholar] [CrossRef]
- Tecchio, M.A.; Teixeira LA, J.; Terra, M.M.; Moura, M.F.; Paioli-Pires, E.J. Extração de nutrientes pela videira ‘Niagara Rosada’ enxertada em diferentes porta-enxertos. Rev. Bras. Frutic. 2011, 33, 736–742. [Google Scholar] [CrossRef]
- Rizk-Alla, M.S.; Sabry, G.H.; Abd-El-Wahab, M.A. Influence of Some Rootstocks on the Performance of Red Globe Grape Cultivar. J. Am. Sci. 2011, 7, 71–81. [Google Scholar]
- Tecchio, M.A.; da Silva, M.J.R.; Sanchez, C.A.P.C.; Callili, D.; Vedoato, B.T.F.; Hernandes, J.L.; Moura, M.F. Yield performance and quality of wine grapes (Vitis vinifera) grafted onto different rootstocks under subtropical conditions. Bragantia 2022, 81, 1. [Google Scholar] [CrossRef]
- Silva, M.J.R.; Paiva, A.P.M.; de Souza, J.F.; Padilha, C.V.d.S.; Basílio, L.S.P.; Lima, M.d.S.; Pereira, G.E.; Corrêa, L.C.; Vianello, F.; Lima, G.P.P.; et al. Phytochemical profile of Brazilian grapes (Vitis labrusca and hybrids) grown on different rootstocks. PLoS ONE 2022, 17, e0275489. [Google Scholar] [CrossRef]
- Sánchez, C.A.P.C.; Tecchio, M.A.; Callili, D.; da Silva, M.J.R.; Basílio, L.S.P.; Leonel, S.; Alonso, J.C.; Lima, G.P.P. Productivity and Physicochemical Properties of the BRS Isis Grape on Various Rootstocks under Subtropical Climatic Conditions. Agriculture 2023, 13, 2113. [Google Scholar] [CrossRef]
- Simonetti, L.M.; Sousa, M.C.; Moura, M.F.; Nunes, J.G.d.S.; Diamante, M.S.; da Silva, M.B.; da Silva, M.J.R.; Callili, D.; Lima, G.P.P.; Tecchio, M.A. The influence of different training systems and rootstocks on Sauvignon Blanc grapes. Bragantia 2021, 80, e2021. [Google Scholar] [CrossRef]
- Pimentel Junior, A.; Domingues, F.J.; Basílio, L.S.P.; Monteiro, G.C.; Lima GP, P.; Tecchio, M.A. Training systems improved agronomic characteristics and quality of ‘Niagara Rosada’ table grapes. Bragantia 2023, 82, e20220252. [Google Scholar] [CrossRef]
- Domingues Neto, F.J.; Pimentel Junior, A.; Modesto, L.R.; Moura, M.F.; Putti, F.F.; Boaro, C.S.F.; Ono, E.O.; Rodrigues, J.D.; Tecchio, M.A. Photosynthesis, Biochemical and Yield Performance of Grapevine Hybrids in Two Rootstock and Trellis Height. Horticulturae 2023, 9, 596. [Google Scholar] [CrossRef]
- Del Zozzo, F.; Magnanini, E.; Poni, S. Physiological efficiency of grapevine canopies having varying geometries: Seasonal and diurnal whole canopy gas exchange assessment under well-watered and water deficit conditions. Environ. Exp. Bot. 2024, 221, 105716. [Google Scholar] [CrossRef]
- Wu, Y.; Qiu, T.; Shen, Z.; Wu, Y.; Lu, D.; He, J. Effects of shading on leaf physiology and morphology in the ‘Yinhong’ grape plants. Rev. Bras. Frutic. 2018, 40, e-037. [Google Scholar] [CrossRef]
- Miccichè, D.; de Rosas, M.I.; Ferro, M.V.; Di Lorenzo, R.; Puccio, S.; Pisciotta, A. Effects of artificial canopy shading on vegetative growth and ripening processes of cv. Nero d’Avola (Vitis vinifera L.). Front. Plant Sci. 2023, 14, 1210574. [Google Scholar] [CrossRef]
- Martínez-Lüscher, J.; Brillante, L.; Kurtural, S.K. Flavonol Profile Is a Reliable Indicator to Assess Canopy Architecture and the Exposure of Red Wine Grapes to Solar Radiation. Front. Plant Sci. 2019, 10, 10. [Google Scholar] [CrossRef]
- Yu, R.; Torres, N.; Tanner, J.D.; Kacur, S.M.; Marigliano, L.E.; Zumkeller, M.; Gilmer, J.C.; Gilmer, J.C.; Gilmer, J.C. Adapting wine grape production to climate change through canopy architecture manipulation and irrigation in warm climates. Front. Plant Sci. 2022, 3, 1015574. [Google Scholar] [CrossRef]
- Bavougian, C.M.; Read, P.E.; Walter-Shea, E. Training System Effects on Sunlight Penetration, Canopy Structure, Yield, and Fruit Characteristics of ‘Frontenac’ Grapevine (Vitis spp.). Int. J. Fruit Sci. 2012, 12, 402–409. [Google Scholar] [CrossRef]
- Würz, D.A.; Marcon Filho, J.L.; Allebrandt, R.; Bem, B.D.; Rufato, L.; Kretzschmar, A.A. Agronomic performance of ‘Cabernet Sauvignon’ grapevine in different training systems in high altitude regions of Santa Catarina State, Brazil. Rev. Ciências Agroveterinárias 2019, 18, 73–80. [Google Scholar]
- Machado, T.O.X.; Portugal, I.; Kodel, H.A.C.; Droppa-Almeida, D.; Lima, M.S.; Fathi, F.; Oliveira, M.B.P.P.; Albuquerque-Júnior, R.L.C.; Dariva, C.; Souto, E.B. Therapeutic potential of grape pomace extracts: A review of scientific evidence. Food Biosci. 2024, 60, e104210. [Google Scholar] [CrossRef]
- Wang, H.; Cao, G.; Prior, R.L. Oxygen radical absorbing capacity of anthocyanins. J. Agric. Food Chem. 1997, 45, 304–309. [Google Scholar] [CrossRef]
- Sanchez-Moreno, C. Compuestos polifenolicos: Efectos fisiologicos. Actividad antioxidante. Alimentaria 2002, 329, 29–40. [Google Scholar]
- Rustioni, L.; Rocchi, L.; Failla, O. Effect of anthocyanin absence on white berry grape (Vitis vinifera L.). Vitis 2015, 54, 239–242. [Google Scholar]
- EMBRAPA. Sistema Brasileiro de Classificação do Solo–Brasília: EMBRAPA, Produção de Informação: 5. ed., Rev. e Ampl. 2018, p. 356. Available online: https://www.agroapi.cnptia.embrapa.br/portal/assets/docs/SiBCS-2018-ISBN-9788570358004.pdf (accessed on 19 July 2024).
- Von Caemmerer, S.; Farquhar, G.D. Some relationships between the biochemistry of photosynthesis and the gas exchange of leaves. Planta 1981, 153, 376–387. [Google Scholar] [CrossRef]
- Klughammer, C.; Schreiber, U. Complementary PS II quantum yields calculated from simple fluorescence parameters measured by PAM fluorometry and the Saturation Pulse method. PAM Appl. Notes 2008, 1, 27–35. [Google Scholar]
- Sims, D.A.; Gamon, J.A. Relationships between leaf pigment content and spectral reflectance across a wide range of species, leaf structures and developmental stages. Remote Sens. Environ. 2002, 81, 337–354. [Google Scholar] [CrossRef]
- Nelson, N.A. A photometric adaptation of Somogy method for the determination of glucose. J. Biol. Chem. 1944, 135, 136–175. [Google Scholar]
- Singleton, V.L.; Rossi Junior, J.A. Colorimetry of total phenolics with phosphomolybidic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar] [CrossRef]
- Giusti, M.M.; Wrolstad, R.E. Characterization and measurement of anthocyanins by UV–Visible spectroscopy. Curr. Protoc. Food Anal. Chem. 2001, 3, 1–13. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. Lebensm. Wiss. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Rossetto, M.R.M.; Vianello, F.; Rocha, S.D.; Lima, G.P.P. Antioxidant substances and pesticide in parts of beet organic and conventional manure. Afr. J. Plant Sci. 2009, 3, 245–253. [Google Scholar]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, D.F. Sisvar: A computer statistical analysis system. Ciência Agrotecnologia 2011, 35, 1039–1042. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E.; Møller, I.M.; Murphy, A. Fisiologia e Desenvolvimento Vegetal; Artmed: Porto Alegre, Brazil, 2017; 888p. [Google Scholar]
- Douthe, C.; Gago, J.; Ribas-Carbó, M.; Núñez, R.; Pedrol, N.; Flexas, J. Measuring photosynthesis and respiration with infrared gas analysers. In Advances in Plant Ecophysiology Techniques; Sánchez-Moreiras, A.M., Reigosa, M.J., Eds.; Springer: Cham, Switzerland, 2018; pp. 51–75. [Google Scholar]
- Tominaga, J.; Shimada, H.; Kawamitsu, Y. Direct measurement of intercellular CO2 concentration in a gas-exchange system resolves overestimation using the standard method. J. Exp. Bot. 2018, 69, 1981–1991. [Google Scholar] [CrossRef]
- Hunter, J.K.; Tarricone, L.; Volschenk, C.; Giacalone, C.; Melo, M.S.; Zorer, R. Grapevine physiological response to row orientation-induced spatial radiation and microclimate changes. OENO One 2020, 54, 411–433. [Google Scholar] [CrossRef]
- Santos Neto, J.R.A. A Cultura da Videira; Instituto Agronômico: Campinas, Brazil, 1973; p. 108.
- Pommer, C.V.; Passos, I.R.S.; Terra, M.M.; Pires, E.J.P. Variedades de Videira Para o Estado de São Paulo; Instituto Agronômico, (IAC), Boletim Técnico: Campinas, Brazil, 1997; Volume 166, p. 59.
- Simão, S. Tratado de Fruticulture; FEALQ: Piracicaba, Brazil, 1998; 760p. [Google Scholar]
- Alvarenga, A.A.; Regina, M.A.; Fráguas, J.C.; Silva, A.L.; Souza, C.M.; Cançado GM, A.; Freitas, G.F. Indicação de porta-enxertos de videiras para o sul de Minas Gerais. In Viticultura e Enologia: Atualizando Conceitos; Epamig: Caldas, Brazil, 2002; pp. 243–256. [Google Scholar]
- Pauletto, D.; Mourão Filho FD, A.A.; Kluge, R.A.; Scarpare Filho, J.A. Produção e vigor da videira ‘Niágara Rosada’ relacionados com o porta-enxerto. Pesqui. Agropecuária Bras. 2001, 36, 115–121. [Google Scholar] [CrossRef]
- Decendit, A.; Mamani-Matsuda, M.; Aumont, V.; Waffo-Teguo, P.; Moynet, D.; Boniface, K.; Richard, E.; Krisa, S.; Rambert, J.; Mérillon, J.-M.; et al. Malvidin-3-O-β glucoside, major grape anthocyanin, inhibits human macrophage-derived inflammatory mediators and decreases clinical scores in arthritic rats. Biochem. Pharmacol. 2013, 86, 1461–1467. [Google Scholar] [CrossRef] [PubMed]
- Granato, D.; Carrapeiro, M.d.M.; Fogliano, V.; van Ruth, S.M. Effects of geographical origin, varietal and farming system on the chemical composition and functional properties of purple grape juices: A review. Trends Food Sci. Technol. 2016, 52, 31–48. [Google Scholar] [CrossRef]
- Abe, L.T.; Mota, R.V.; Lajolo, F.M.; Genovese, M.I. Compostos fenólicos e capacidade antioxidante de cultivares de uvas Vitis labrusca L. e Vitis vinifera L. Food Sci. Technol. 2007, 27, 394–400. [Google Scholar] [CrossRef]
- Ferrari, D.; Speciale, A.; Cristani, M.; Fratantonio, D.; Molonia, M.S.; Ranaldi, G.; Saija, A.; Cimino, F. Cyanidin-3-O-glucoside inhibits NF-kB signalling in intestinal epithelial cells exposed to TNF-kB and exerts protective effects via Nrf2 pathway activation. Toxicol. Lett. 2016, 264, 51–58. [Google Scholar] [CrossRef]
- Meng, X.; Maliakal, P.; Lu, H.; Lee, M.J.; Yang, C.S. Urinary and plasma levels of resveratrol and quercetin in humans, mice, and rats after ingestion of pure compounds and grape juice. J. Agric. Food Chem. 2004, 52, 935–942. [Google Scholar] [CrossRef]
- Dani, C.; Oliboni, L.S.; Pasquali, M.A.; Oliveira, M.R.; Umezu, F.M.; Salvador, M.; Moreira, J.C.; Henriques, J.A. Intake of purple grape juice as a hepatoprotective agent in wistar rats. J. Med. Food 2008, 11, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Kumar, G.A.; Kumar, S.; Bhardwaj, R.; Swapnil, P.; Meena, M.; Seth, C.S.; Yadav, A. Recent advancements in multifaceted roles of flavonoids in plant–rhizomicrobiome interactions. Front. Plant Sci. 2024, 14, 1297706. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Li, L.; Wang, S.; Yu, N.; Shan, H.; Shi, Z.; Li, F.; Zhong, X. Effect of gibberellic acid on photosynthesis and oxidative stress response in maize under weak light conditions. Front. Plant Sci. 2023, 14, 1128780. [Google Scholar] [CrossRef]
- Chaimala, A.; Jogloy, S.; Vorasoot, N.; CHolbrook, C.; Kvien, C.K.; Laohasiriwong, S. The variation of relative water content, SPAD chlorophyll meter reading, stomatal conductance, leaf area, and specific leaf area of Jerusalem artichoke genotypes under different durations of terminal drought in tropical region. J. Agron. Crop Sci. 2023, 209, 12–26. [Google Scholar] [CrossRef]
- Alotaibi, M.; El-Hendawy, S.; Mohammed, N.; Alsamin, B.; Refay, Y. Appropriate application methods for salicylic acid and plant nutrients combinations to promote morpho-physiological traits, production, and water use efficiency of wheat under normal and deficit irrigation in an arid climate. Plants 2023, 12, 1368. [Google Scholar] [CrossRef]
- Hartmann, H.T.; Kester, D.E. Propagación de Plantas: Principios y Practicas; Continental: Colonia San Juan Tlihuaca, México, 1990; p. 760. [Google Scholar]
- Baldassi, C.; Berim, A.; Roeder, S.; Losciale, P.; Serra, S.; Gang, D.R.; Musacchi, S. Rootstock and Crop Load Effects on ‘Honeycrisp’Photosynthetic Performance and Carbohydrate Accumulation. Plants 2023, 12, 4035. [Google Scholar] [CrossRef] [PubMed]
- Kodur, S.; Tisdall, J.M.; Clingeleffer, P.R.; Walker, R.R. Regulation of berry quality parameters in ‘Shiraz’ grapevines through rootstocks (Vitis). Vitis 2013, 52, 125–128. [Google Scholar]
- Dias, F.A.N.; da Mota, R.V.; Fávero, A.C.; Purgatto, E.; Shiga, T.M.; de Souza, C.R.; Pimentel, R.M.d.A.; Regina, M.d.A. Videira ‘Syrah’ sobre diferentes portaenxertos em ciclo de inverno no sul de Minas Gerais. Pesqui. Agropecuária Bras. 2012, 47, 208–215. [Google Scholar] [CrossRef]
- Lee, J.; Steenwerth, K.L. ‘Cabernet Sauvignon’ grape anthocyanin increased by soil conservation practices. Sci. Hortic. 2013, 159, 128–133. [Google Scholar] [CrossRef]
- Tecchio, M.A.; Moura, M.F.; Teixeira, L.A.J.; Pires, E.J.P.; Leonel, S. Influence of rootstocks and pruning times on yield and on nutrient content and extraction in ‘Niagara Rosada’ grapevine. Pesqui. Agropecuária Bras. 2014, 49, 340–348. [Google Scholar] [CrossRef]
- Yamamoto, L.Y.; de Assis, A.M.; Roberto, S.R.; Bovolenta, Y.R.; Nixdorf, S.L.; García-Romero, E.; Gómez-Alonso, S.; Hermosín-Gutiérrez, I. Application of abscisic acid (S -ABA) to cv. Isabel grapes (Vitis vinifera × Vitis labrusca) for color improvement: Effects on color, phenolic composition and antioxidant capacity of their grape juice. Food Res. Int. 2015, 77, 572–583. [Google Scholar] [CrossRef]
- Mian, G.; Colautti, A.; Belfiore, N.; Marcuzzo, P.; Tomasi, D.; Bell, L.; Celotti, E. Fertigation affects photosynthesis, modulation of secondary metabolism and sensory profiles of Vitis vinifera cv.“Schioppettino” withered grapes and wines. Sci. Hortic. 2024, 328, 112954. [Google Scholar] [CrossRef]
- Rebello, L.P.G.; Lago-Vanzela, E.S.; Barcia, M.T.; Ramos, A.M.; Stringheta, P.C.; Da-Silva, R.; Castillo-Muñoz, N.; Gómez-Alonso, S.; Hermosín-Gutiérrez, I. Phenolic composition of the berry parts of hybrid grape cultivar BRS Violeta (BRS Rubea × IAC 1398-21) using HPLC-DAD-ESI-MS/MS. Food Res. Int. 2013, 54, 354–366. [Google Scholar] [CrossRef]
- Barcia, M.T.; Pertuzatti, P.B.; Gómez-Alonso, S.; Godoy, H.T.; Hermosín-Gutiérrez, I. Phenolic composition of grape and winemaking by-products of Brazilian hybrid cultivars BRS Violeta and BRS Lorena. Food Chem. 2014, 159, 95–105. [Google Scholar] [CrossRef]
- Lima, M.d.S.; Silani, I.d.S.V.; Toaldo, I.M.; Corrêa, L.C.; Biasoto, A.C.T.; Pereira, G.E.; Bordignon-Luiz, M.T.; Ninow, J.L. Phenolic compounds, organic acids and antioxidant activity of grape juices produced from new Brazilian varieties planted in the Northeast Region of Brazil. Food Chem. 2014, 161, 94–103. [Google Scholar] [CrossRef]
- Caiati, C.; Stanca, A.; Lepera, M.E. Free radicals and obesity-related chronic inflammation contrasted by antioxidants: A new perspective in coronary artery disease. Metabolites 2023, 13, 712. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).