
PhSPEAR1 Participates in Regulating the Branch Development of Petunia


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Isolation of PhSPEAR1
2.3. Bioinformatic Analysis
2.4. Subcellular Localization Analysis
2.5. qRT-PCR Analysis
2.6. Construction and Transformation of 35S::PhSPEAR1
2.7. Arabidopsis Transformation
2.8. Phenotype Analysis and Statistical Analysis
2.9. Sample Preparation and Plant Hormone Extraction
2.10. Statistical Analysis
3. Results
3.1. Identification of SPEAR1 in Petunia
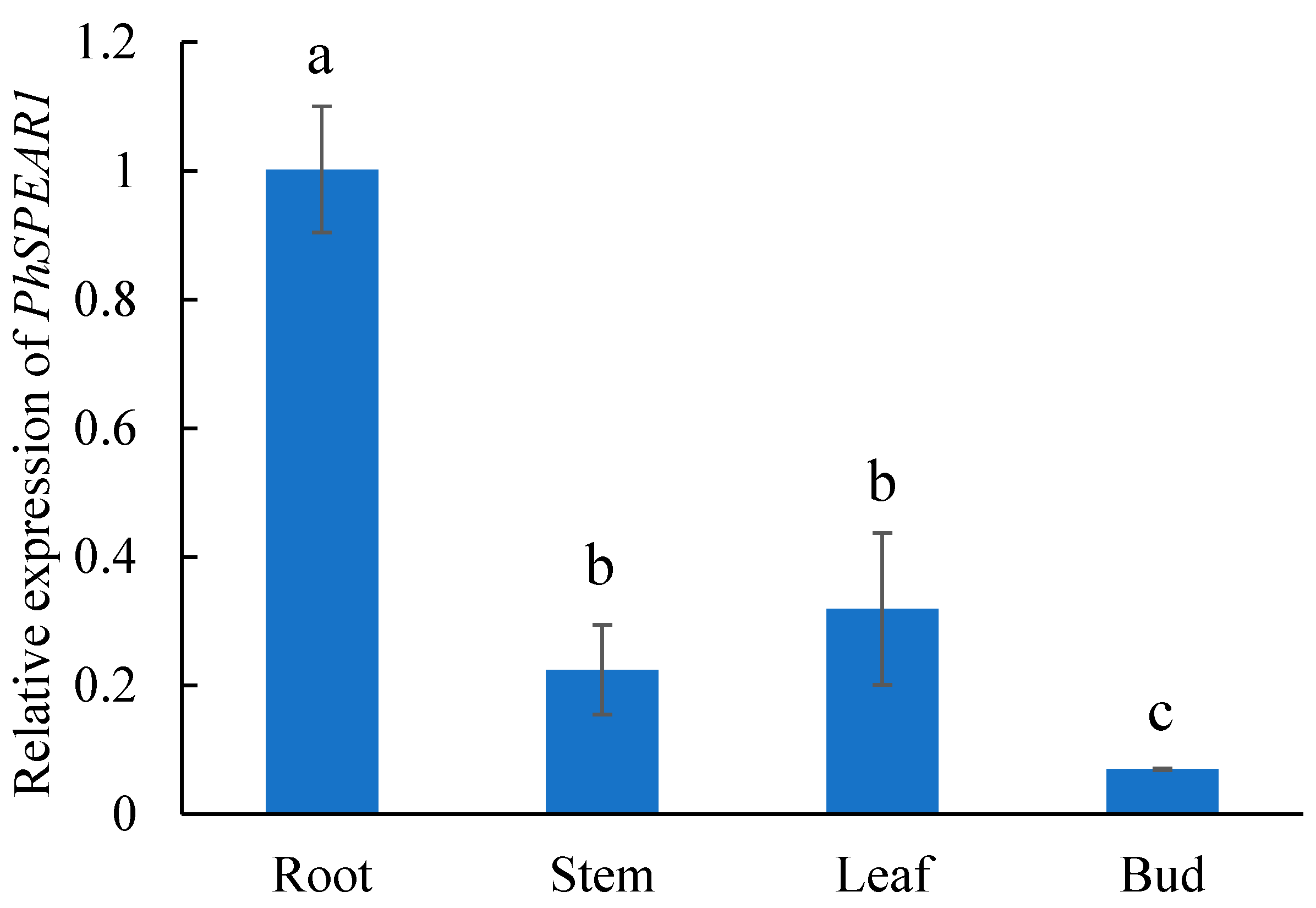
3.2. Tissue-Specific Expression Analysis of PhSPEAR1
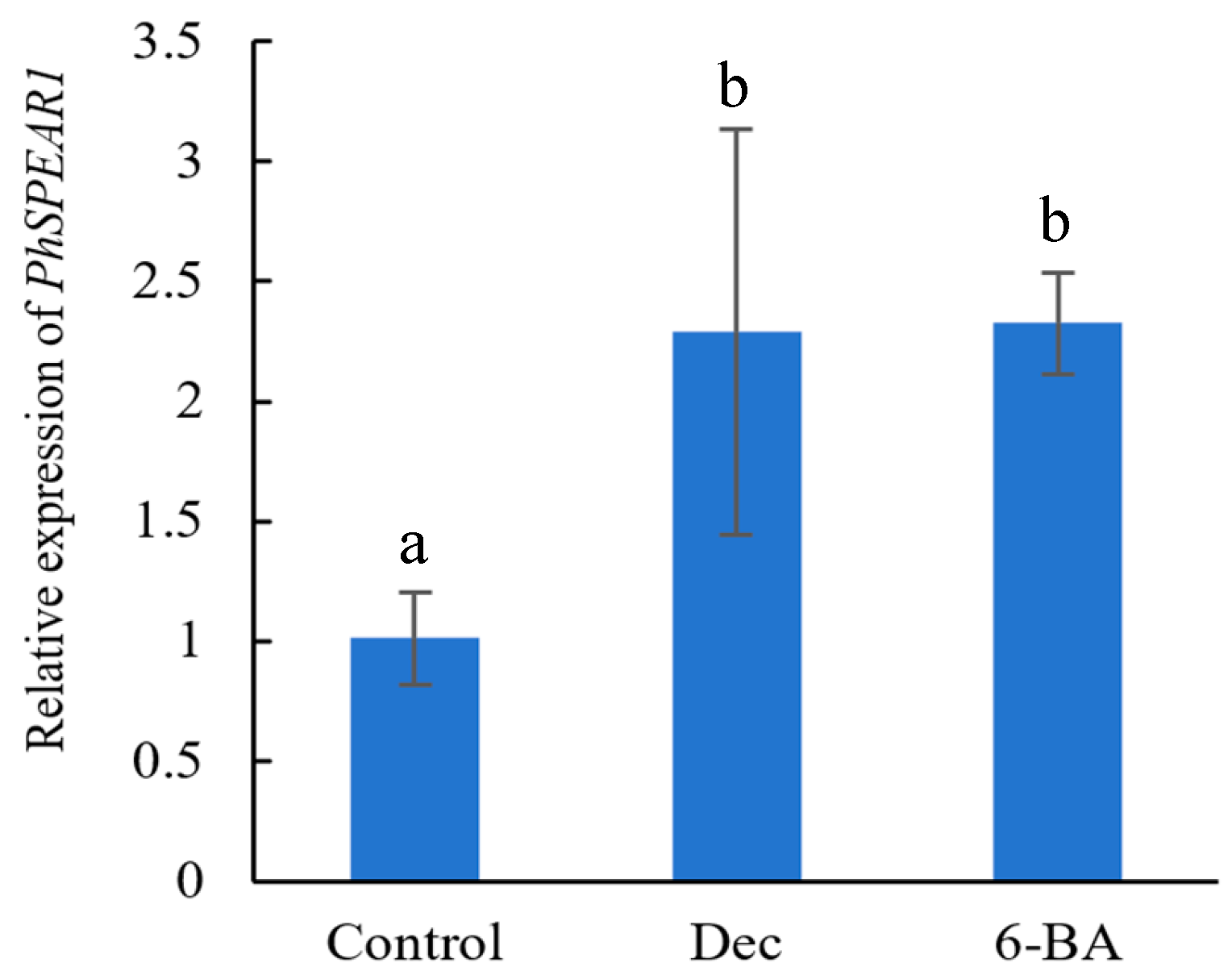
3.3. Expression Analysis of PhSPEAR1 under Branch-Inducing Treatments
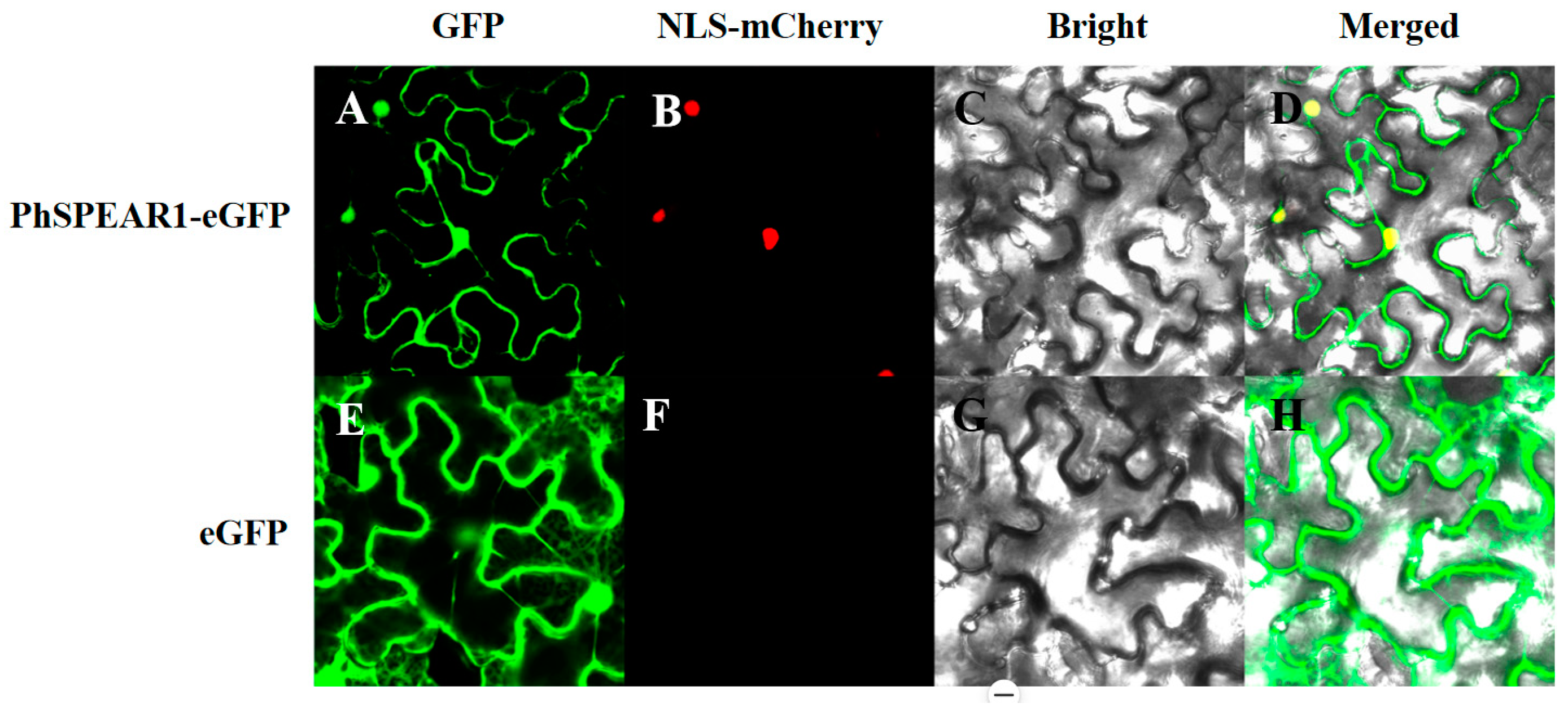
3.4. Subcellular Localization Analysis of PhSPEAR1
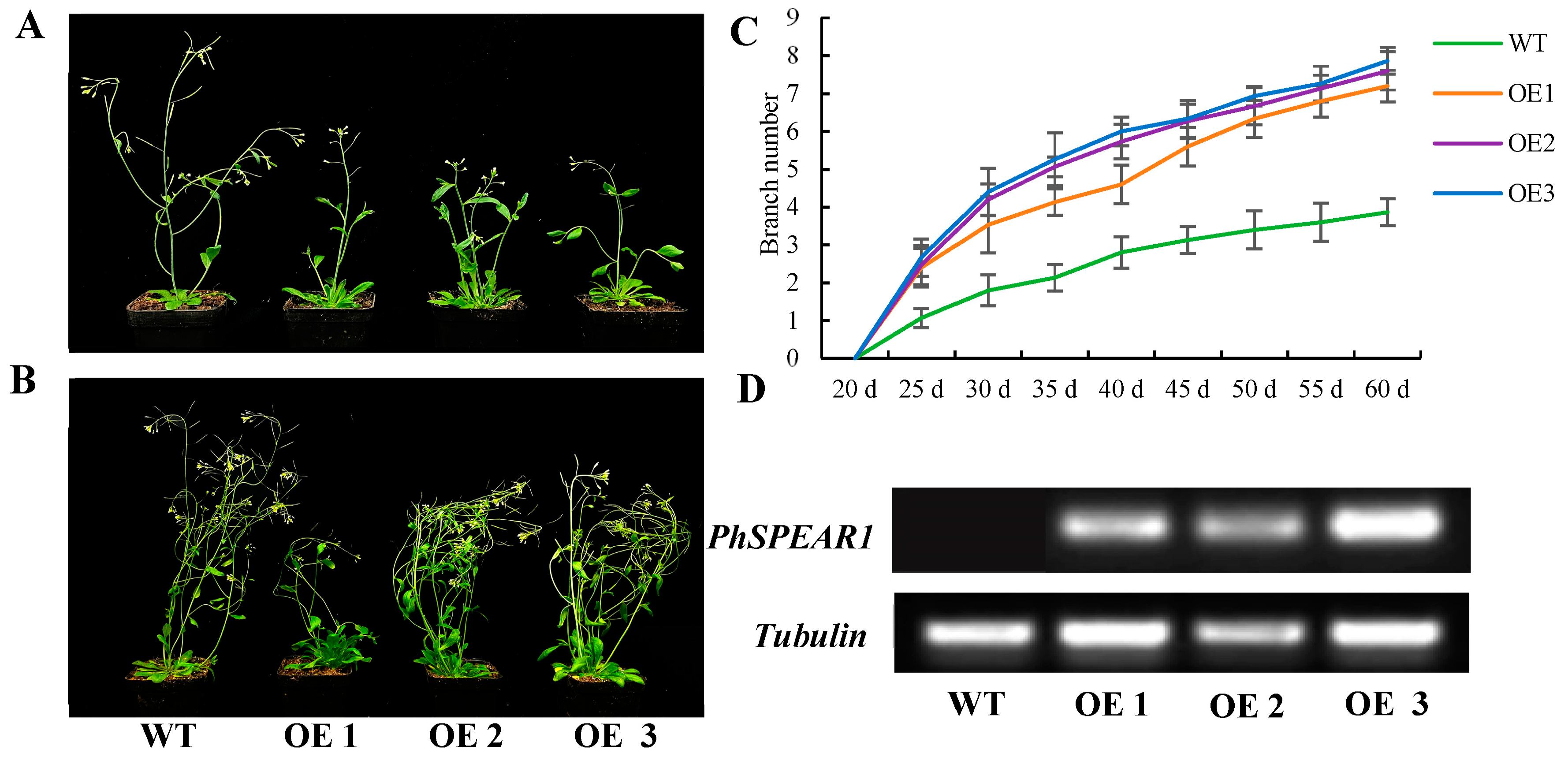
3.5. Overexpression of PhSPEAR1 Increases Branch Number in Arabidopsis
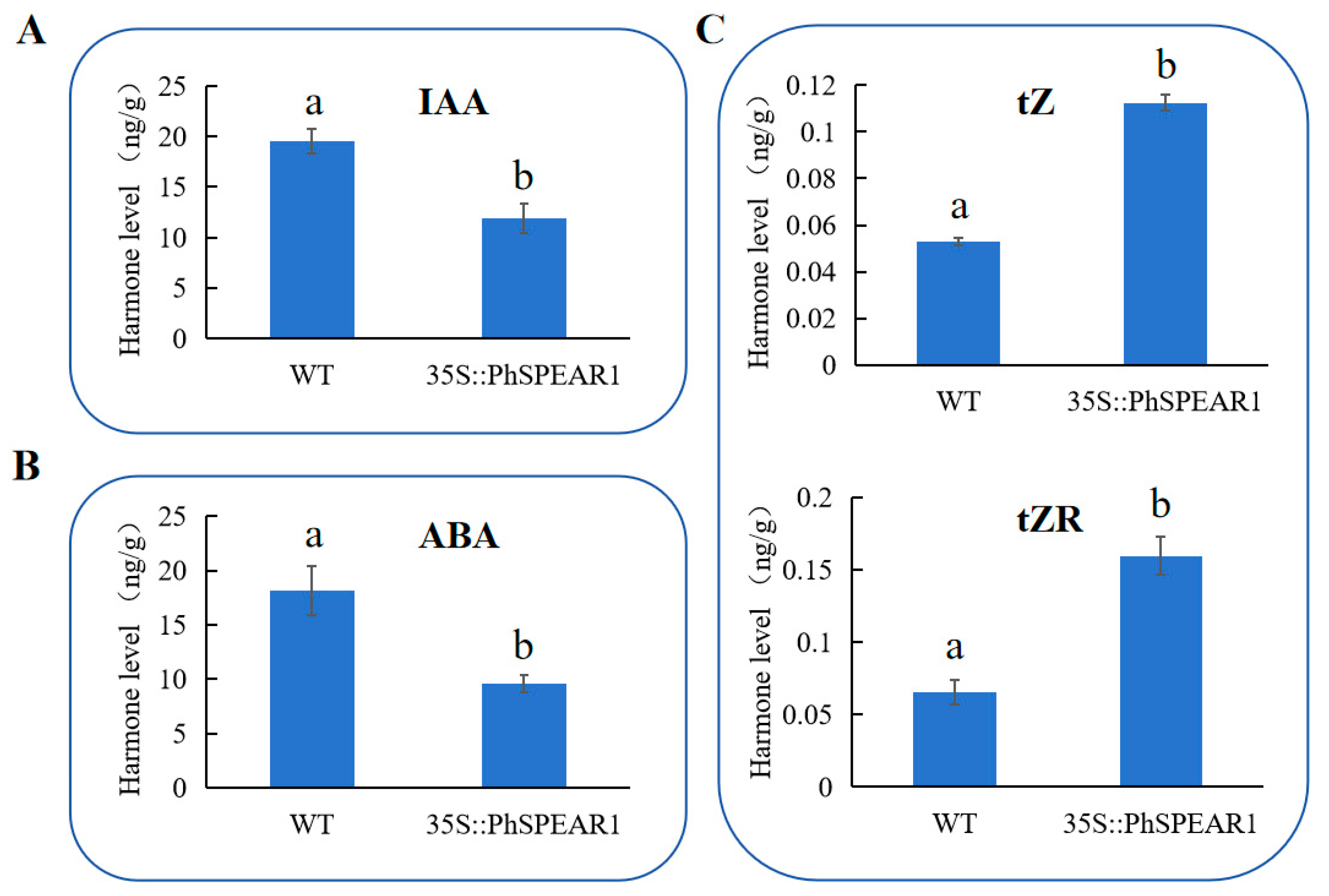
3.6. PhSPEAR1 Regulates the Synthesis of CKs, IAA, and ABA
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hou, M.; Wu, D.; Li, Y.; Tao, W.; Chao, L.; Zhang, Y. The role of auxin in nitrogen-modulated shoot branching. Plant Signal. Behav. 2021, 16, 1885888. [Google Scholar] [CrossRef] [PubMed]
- Del Rosario Cárdenas-Aquino, M.; Sarria-Guzmán, Y.; Martínez-Antonio, A. Review: Isoprenoid and aromatic cytokinins in shoot branching. Plant Sci. 2022, 319, 111240. [Google Scholar] [CrossRef] [PubMed]
- Dun, E.A.; Brewer, P.B.; Gillam, E.M.J.; Beveridge, C.A. Strigolactones and Shoot Branching: What Is the Real Hormone and How Does It Work? Plant Cell Physiol. 2023, 64, 967–983. [Google Scholar] [CrossRef] [PubMed]
- Katyayini, N.U.; Rinne, P.L.H.; Tarkowská, D.; Strnad, M.; van der Schoot, C. Dual Role of Gibberellin in Perennial Shoot Branching: Inhibition and Activation. Front. Plant Sci. 2020, 11, 736. [Google Scholar] [CrossRef]
- Salam, B.B.; Barbier, F.; Danieli, R.; Teper-Bamnolker, P.; Ziv, C.; Spíchal, L.; Aruchamy, K.; Shnaider, Y.; Leibman, D.; Shaya, F.; et al. Sucrose promotes stem branching through cytokinin. Plant Physiol. 2021, 185, 1708–1721. [Google Scholar] [CrossRef]
- Fichtner, F.; Barbier, F.F.; Annunziata, M.G.; Feil, R.; Olas, J.J.; Mueller-Roeber, B.; Stitt, M.; Beveridge, C.A.; Lunn, J.E. Regulation of shoot branching in arabidopsis by trehalose 6-phosphate. New Phytol. 2021, 229, 2135–2151. [Google Scholar] [CrossRef]
- de Jong, M.; Tavares, H.; Pasam, R.K.; Butler, R.; Ward, S.; George, G.; Melnyk, C.W.; Challis, R.; Kover, P.X.; Leyser, O. Natural variation in Arabidopsis shoot branching plasticity in response to nitrate supply affects fitness. PLoS Genet. 2019, 15, e1008366. [Google Scholar] [CrossRef]
- Holalu, S.V.; Finlayson, S.A. The ratio of red light to far red light alters Arabidopsis axillary bud growth and abscisic acid signalling before stem auxin changes. J. Exp. Bot. 2017, 68, 943–952. [Google Scholar] [CrossRef]
- Yang, Y.; Ahmad, S.; Yang, Q.; Yuan, C.; Zhang, Q. Decapitation Experiments Combined with the Transcriptome Analysis Reveal the Mechanism of High Temperature on Chrysanthemum Axillary Bud Formation. Int. J. Mol. Sci. 2021, 22, 9704. [Google Scholar] [CrossRef]
- Semeradova, H.; Montesinos, J.C.; Benkova, E. All Roads Lead to Auxin: Post-translational Regulation of Auxin Transport by Multiple Hormonal Pathways. Plant Commun. 2020, 1, 100048. [Google Scholar] [CrossRef]
- Dun, E.A.; Brewer, P.B.; Beveridge, C.A. Strigolactones: Discovery of the elusive shoot branching hormone. Trends Plant Sci. 2009, 14, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Müller, D.; Leyser, O. Auxin, cytokinin and the control of shoot branching. Ann. Bot. 2011, 107, 1203–1212. [Google Scholar] [CrossRef]
- Gomez-Roldan, V.; Fermas, S.; Brewer, P.B.; Puech-Pagès, V.; Dun, E.A.; Pillot, J.P.; Letisse, F.; Matusova, R.; Danoun, S.; Portais, J.C.; et al. Strigolactone inhibition of shoot branching. Nature 2008, 455, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Sasse, J.; Simon, S.; Gübeli, C.; Liu, G.W.; Cheng, X.; Friml, J.; Bouwmeester, H.; Martinoia, E.; Borghi, L. Asymmetric localizations of the ABC transporter PaPDR1 trace paths of directional strigolactone transport. Curr. Biol. 2015, 25, 647–655. [Google Scholar] [CrossRef]
- Seale, M.; Bennett, T.; Leyser, O. BRC1 expression regulates bud activation potential but is not necessary or sufficient for bud growth inhibition in Arabidopsis. Development 2017, 144, 1661–1673. [Google Scholar] [CrossRef]
- Yao, C.; Finlayson, S.A. Abscisic Acid Is a General Negative Regulator of Arabidopsis Axillary Bud Growth. Plant Physiol. 2015, 169, 611–626. [Google Scholar] [CrossRef] [PubMed]
- Cline, M.G.; Oh, C. A reappraisal of the role of abscisic acid and its interaction with auxin in apical dominance. Ann. Bot. 2006, 98, 891–897. [Google Scholar] [CrossRef] [PubMed]
- Arend, M.; Schnitzler, J.P.; Ehlting, B.; Hänsch, R.; Lange, T.; Rennenberg, H.; Himmelbach, A.; Grill, E.; Fromm, J. Expression of the Arabidopsis mutant abi1 gene alters abscisic acid sensitivity, stomatal development, and growth morphology in gray poplars. Plant Physiol. 2009, 151, 2110–2119. [Google Scholar] [CrossRef]
- Chen, G.H.; Sun, J.Y.; Liu, M.; Liu, J.; Yang, W.C. SPOROCYTELESS is a novel embryophyte-specific transcription repressor that interacts with TPL and TCP proteins in Arabidopsis. J. Genet. Genom. 2014, 41, 617–625. [Google Scholar] [CrossRef]
- Tao, Q.; Guo, D.; Wei, B.; Zhang, F.; Pang, C.; Jiang, H.; Zhang, J.; Wei, T.; Gu, H.; Qu, L.J.; et al. The TIE1 transcriptional repressor links TCP transcription factors with TOPLESS/TOPLESS-RELATED corepressors and modulates leaf development in Arabidopsis. Plant Cell 2013, 25, 421–437. [Google Scholar] [CrossRef]
- He, Q.; Yuan, R.; Zhang, T.; An, F.; Wang, N.; Lan, J.; Wang, X.; Zhang, Z.; Pan, Y.; Wang, X.; et al. Arabidopsis TIE1 and TIE2 transcriptional repressors dampen cytokinin response during root development. Sci. Adv. 2022, 8, eabn5057. [Google Scholar] [CrossRef] [PubMed]
- Bombarely, A.; Moser, M.; Amrad, A.; Bao, M.; Bapaume, L.; Barry, C.S.; Bliek, M.; Boersma, M.R.; Borghi, L.; Bruggmann, R.; et al. Insight into the evolution of the Solanaceae from the parental genomes of Petunia hybrida. Nat. Plants 2016, 2, 16074. [Google Scholar] [CrossRef] [PubMed]
- Hoshikawa, K.; Fujita, S.; Renhu, N.; Ezura, K.; Yamamoto, T.; Nonaka, S.; Ezura, H.; Miura, K. Efficient transient protein expression in tomato cultivars and wild species using agroinfiltration-mediated high expression system. Plant Cell Rep. 2019, 38, 75–84. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef]
- Yang, W.C.; Ye, D.; Xu, J.; Sundaresan, V. The SPOROCYTELESS gene of Arabidopsis is required for initiation of sporogenesis and encodes a novel nuclear protein. Genes Dev. 1999, 13, 2108–2117. [Google Scholar] [CrossRef]
- Wei, B.; Zhang, J.; Pang, C.; Yu, H.; Guo, D.; Jiang, H.; Ding, M.; Chen, Z.; Tao, Q.; Gu, H.; et al. The molecular mechanism of sporocyteless/nozzle in controlling Arabidopsis ovule development. Cell Res. 2015, 25, 121–134. [Google Scholar] [CrossRef]
- Rojas-Gracia, P.; Roque, E.; Medina, M.; Rochina, M.; Hamza, R.; Angarita-Díaz, M.P.; Moreno, V.; Pérez-Martín, F.; Lozano, R.; Cañas, L.; et al. The parthenocarpic hydra mutant reveals a new function for a SPOROCYTELESS-like gene in the control of fruit set in tomato. New Phytol. 2017, 214, 1198–1212. [Google Scholar] [CrossRef]
- Li, L.C.; Qin, G.J.; Tsuge, T.; Hou, X.H.; Ding, M.Y.; Aoyama, T.; Oka, A.; Chen, Z.; Gu, H.; Zhao, Y.; et al. SPOROCYTELESS modulates YUCCA expression to regulate the development of lateral organs in Arabidopsis. New Phytol. 2008, 179, 751–764. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Huang, J.; Parameswaran, S.; Ito, T.; Seubert, B.; Auer, M.; Rymaszewski, A.; Jia, G.; Owen, H.A.; Zhao, D. The SPOROCYTELESS/NOZZLE gene is involved in controlling stamen identity in Arabidopsis. Plant Physiol. 2009, 151, 1401–1411. [Google Scholar] [CrossRef]
- Sun, D.; Zhang, L.; Yu, Q.; Zhang, J.; Li, P.; Zhang, Y.; Xing, X.; Ding, L.; Fang, W.; Chen, F.; et al. Integrated Signals of Jasmonates, Sugars, Cytokinins and Auxin Influence the Initial Growth of the Second Buds of Chrysanthemum after Decapitation. Biology 2021, 10, 440. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.; Wu, Y.; Zhang, J.; Deng, X.; Wang, T. Transcriptome Analysis Revealed Hormone Pathways and bZIP Genes Responsive to Decapitation in Sunflower. Genes 2022, 13, 1737. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.T.; Liu, D.H.; Luo, Y.; Khan, M.A.; Alam, S.M.; Liu, Y.Z. Transcriptome analysis reveals the key network of axillary bud outgrowth modulated by topping in citrus. Gene 2024, 926, 148623. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, W.; Li, C.; Zhang, Q.; Wang, Y.; Li, X.; Yang, Q.; Dong, L. PhSPEAR1 Participates in Regulating the Branch Development of Petunia. Horticulturae 2024, 10, 912. https://doi.org/10.3390/horticulturae10090912
Wu W, Li C, Zhang Q, Wang Y, Li X, Yang Q, Dong L. PhSPEAR1 Participates in Regulating the Branch Development of Petunia. Horticulturae. 2024; 10(9):912. https://doi.org/10.3390/horticulturae10090912
Chicago/Turabian StyleWu, Wei, Chaoqun Li, Qi Zhang, Yan Wang, Xueying Li, Qin Yang, and Lili Dong. 2024. "PhSPEAR1 Participates in Regulating the Branch Development of Petunia" Horticulturae 10, no. 9: 912. https://doi.org/10.3390/horticulturae10090912
APA StyleWu, W., Li, C., Zhang, Q., Wang, Y., Li, X., Yang, Q., & Dong, L. (2024). PhSPEAR1 Participates in Regulating the Branch Development of Petunia. Horticulturae, 10(9), 912. https://doi.org/10.3390/horticulturae10090912