
Viticultural Performance of Hybrids and Vitis vinifera Varieties Established in Annapolis Valley (Nova Scotia)



Abstract
1. Introduction
2. Materials and Methods
2.1. Site of Study and Plant Material
2.2. Climate and Soil Conditions
2.3. Plant Phenology Determinations
2.4. Viticultural Parameters and Shoot Growth
2.5. Plant Water Status and δ13C
2.6. Plant Nutrient Status
2.7. Must Determinations
2.8. Statistical Analysis
3. Results
3.1. Vine Phenology
3.2. Petiole Nutrient Content
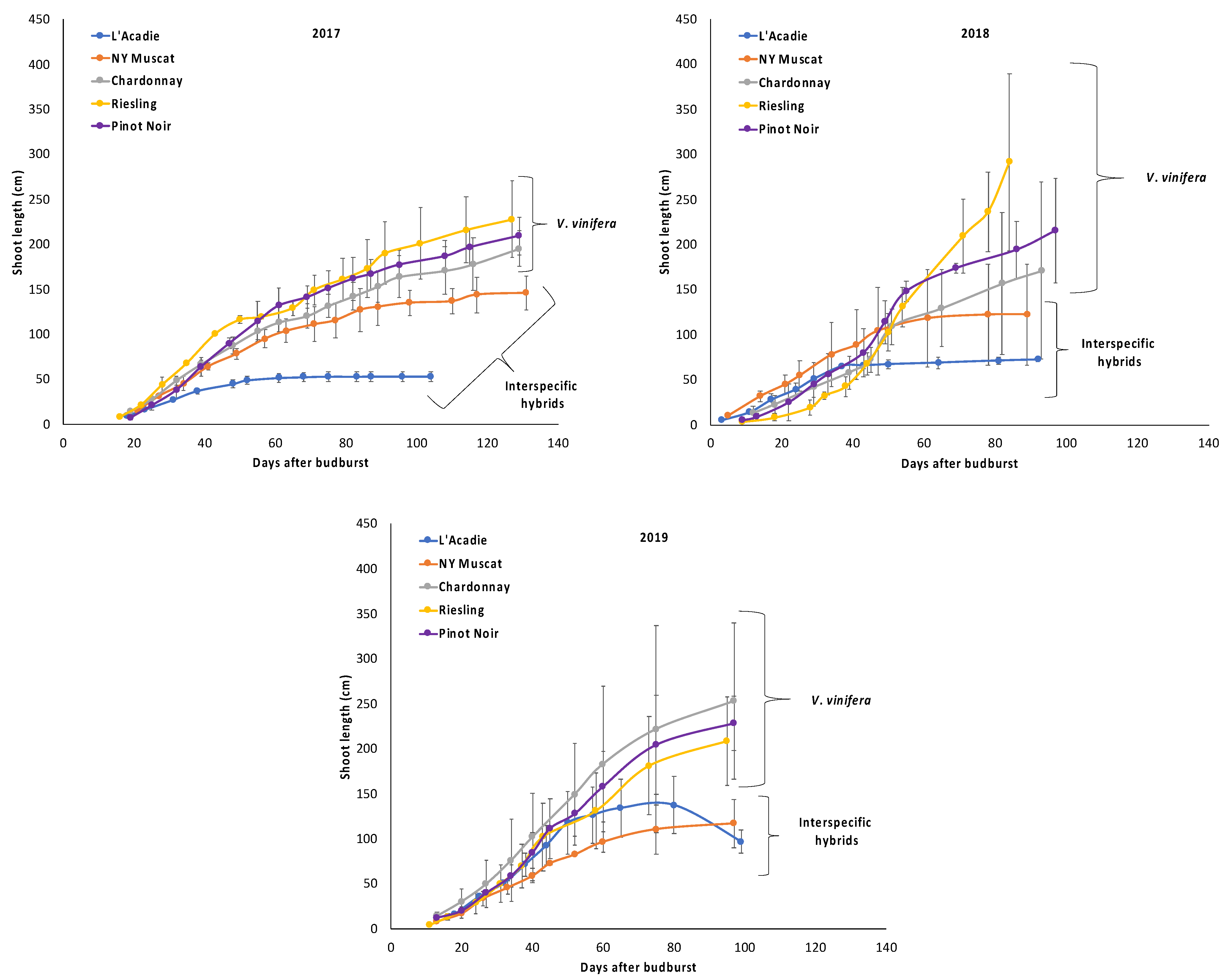
3.3. Shoot Growth Developing
3.4. Yield and Water Status
3.5. Must Physico-Chemical Parameters
3.6. Principal Component Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Naugler, C. The Tangled Vine: Winegrowing in Nova Scotia; Blue Frog: Bridgewater, NS, Canada, 2004. [Google Scholar]
- Frank, A. The Economic Impact of the Wine and Grape Industry in Canada 2015; Canada Vintners Association Winery and Grower Alliance of Ontario British Columbia Wine Institute Winery Association of Nova Scotia: Ottawa, ON, Canada, 2017. [Google Scholar]
- Naugler, C.; Wright, B. Wamboldt’s Nova Scotia Winegrowers’ Guide; Blue Frog: Bridgewater NS, Canada, 2006. [Google Scholar]
- Provost, C.; Pedneault, K. The organic vineyard as a balanced ecosystem: Improved organic grape management and impacts on wine quality. Sci. Hortic. 2016, 208, 43–56. [Google Scholar] [CrossRef]
- Nicolle, P.; Marcotte, C.; Angers, P.; Pedneault, K. Pomace limits tannin retention in Frontenac wines. Food Chem. 2019, 277, 438–447. [Google Scholar] [CrossRef] [PubMed]
- Pedneault, K.; Provost, C. Fungus resistant grape varieties as a suitable alternative for organic wine production: Benefits, limits, and challenges. Sci. Hortic. 2016, 208, 57–77. [Google Scholar] [CrossRef]
- Slegers, A.; Angers, P.; Ouellet, É.; Truchon, T.; Pedneault, K. Volatile compounds from grape skin, juice and wine from five interspecific hybridgrape cultivars grown in Québec (Canada) for wine production. Molecules 2015, 20, 10980–11016. [Google Scholar] [CrossRef] [PubMed]
- Pedneault, K.; Dorais, M.; Angers, P. Flavor of cold-hardy grapes: Impact of berry maturity and environmental conditions. J. Agric. Food Chem. 2013, 61, 10418–10438. [Google Scholar] [CrossRef]
- Pickering, G.J.; Soleas, G.J. Influence of variety, wine style, vintage and viticultural area on selected chemical parameters of Canadian Icewine. J. Food Agric. Environ. 2007, 5, 97–101. [Google Scholar]
- Mansfield, A.K.; Vickers, Z.M. Characterization of the aroma of red Frontenac table wines by descriptive analysis. Am. J. Enol. Vitic. 2009, 60, 435–441. [Google Scholar]
- Nicolle, P.; Marcotte, C.; Angers, P.; Pedneault, K. Co-fermentation of red grapes and white pomace: A natural and economical process to modulate hybrid wine composition. Food Chem. 2018, 242, 481–490. [Google Scholar] [CrossRef]
- Diez-Zamudio, F.; Laytte, R.; Grallert, C.; Gutiérrez-Gamboa, G. Nutritional status differentially affect yield and must composition of hybrids and V. vinifera varieties established under cold climate conditions. Ciência e Técnica Vitivinícola 2021, 36, 89–103. [Google Scholar] [CrossRef]
- Winkler, A.J.; Cook, J.A.; Kliewer, W.M.; Lider, A.L.; Cerruti, L. General Viticulture; University of California Press: Berkeley, CA, USA, 1974. [Google Scholar]
- Huglin, P. Nouveau mode d’évaluation des possibilités héliothermiques d’un milieu viticole. Comptes Rendus l’Académie d’Agriculture France 1978, 64, 1117–1126. [Google Scholar] [CrossRef][Green Version]
- Tonietto, J.; Carbonneau, A. A multicriteria climatic classification system for grape-growing regions worldwide. Agric. For. Meteorol. 2004, 124, 81–97. [Google Scholar] [CrossRef]
- Jones, G.V. Climate and terroir: Impacts of climate change variability and change on wine. GeoSci. Canada 2005, 73, 1–14. [Google Scholar] [CrossRef]
- Gashu, K.; Sikron Persi, N.; Drori, E.; Harcavi, E.; Agam, N.; Bustan, A.; Fait, A. Temperature shift between vineyards modulates berry phenology and primary metabolism in a varietal collection of wine grapevine. Front. Plant. Sci. 2020, 17, 588739. [Google Scholar] [CrossRef] [PubMed]
- Coombe, B.G. Growth stages of the grapevine: Adoption of a system for identifying grapevine growth stages. Aust. J. Grape Wine Res. 1995, 1, 104–110. [Google Scholar] [CrossRef]
- Spitzke, M.E.; Fauhl-Hassek, C. Determination of the 13C/12C ratios of ethanol and higher alcohols in wine by GC-C-IRMS analysis. Eur. Food Res. Technol. 2010, 231, 247–257. [Google Scholar] [CrossRef]
- AOAC International. Official Methods of Analysis of AOAC International 18th Edition; 4.2.08 AOAC Official Method 990.03, Combustion Method; AOAC International: Rockville, MD, USA, 2006. [Google Scholar]
- AOAC International. Official Methods of Analysis of AOAC International 18th Edition Annual Meeting; AOAC Official Method 968.08, modified method using ICP-OES; AOAC International: Rockville, MD, USA, 2005. [Google Scholar]
- OIV Compendium of Internationals Methods of Wine and Must Analysis. Physical and Chemical Analysis. 2003. Available online: https://www.oiv.int/en/technical-standards-and-documents/methods-of-analysis/compendium-of-international-methods-of-analysis-of-wines-and-musts-2-vol (accessed on 7 July 2021).
- OIV Compendium of Internationals Methods of Wine and Must Analysis. Organic acids method OIV-MA-AS313-04 2009. Available online: https://docplayer.fr/12139221-Compendium-of-international-methods-of-wine-and-must-analysis-international-organisation-of-vine-and-wine.html (accessed on 7 July 2021).
- Skoutelas, D.; Ricardo-da-Silva, J.M.; Laureano, O. Validation and comparison of formol and FT-IR methods for assimilable nitrogen in vine grapes. S. Afr. J. Enol. Vitic. 2011, 32, 262–266. [Google Scholar] [CrossRef][Green Version]
- Poitou, X.; Thibon, C.; Darriet, P. 1,8-Cineole in French red wines: Evidence for a contribution related to its various origins. J. Agric. Food Chem. 2017, 65, 383–393. [Google Scholar] [CrossRef]
- Leeuwen Van, C.; Roby, J.-P.; Rességuier, L. de Soil-related terroir factors: A review. OENO One 2018, 52, 173–188. [Google Scholar] [CrossRef]
- Clarke, S.J.; Lamont, K.J.; Pan, H.Y.; Barry, L.A.; Hall, A.; Rogiers, S.Y. Spring root-zone temperature regulates root growth, nutrient uptake and shoot growth dynamics in grapevines. Aust. J. Grape Wine Res. 2015, 21, 479–489. [Google Scholar] [CrossRef]
- Jones, G.V.; White, M.A.; Cooper, O.R.; Storchmann, K. Climate change and global wine quality. Clim. Change 2005, 73, 319–343. [Google Scholar] [CrossRef]
- Fisher, K.H.; Jamieson, A.R. L’acadie, a cold hardy, white wine grape cultivar for low heat unit regions. Acta Hortic. 2000, 528, 563–567. [Google Scholar] [CrossRef]
- Londo, J.P.; Johnson, L.M. Variation in the chilling requirement and budburst rate of wild Vitis species. Environ. Exp. Bot. 2014, 106, 138–147. [Google Scholar] [CrossRef]
- Schnabel, B.J.; Wample, R.L. Dormancy and cold hardiness in Vitis vinifera L. cv. white Riesling as influenced by photoperiod and temperature. Am. J. Enol. Vitic. 1987, 38, 265–272. [Google Scholar]
- Hamman, R.A.; Dami, I.-E.; Walsh, T.M.; Stushnoff, C. Seasonal carbohydrate changes and cold hardiness of Chardonnay and Riesling grapevines. Am. J. Enol. Vitic. 1996, 47, 31–36. [Google Scholar]
- Ferguson, J.C.; Moyer, M.M.; Mills, L.J.; Hoogenboom, G.; Keller, M. Modeling dormant bud cold hardiness and budbreak in twenty-three Vitis genotypes reveals variation by region of origin. Am. J. Enol. Vitic. 2014, 65, 59–71. [Google Scholar] [CrossRef]
- Péros, J.-P.; Torregrosa, L.; Berger, G. Variability among Vitis vinifera cultivars in micropropagation, organogenesis and antibiotic sensitivity. J. Exp. Bot. 1998, 49, 171–179. [Google Scholar] [CrossRef]
- Keller, M.; Arnink, K.J.; Hrazdina, G. Interaction of nitrogen availability during bloom and light intensity during veraison. I. Effects on grapevine growth, fruit development, and ripening. Am. J. Enol. Vitic. 1998, 49, 333–340. [Google Scholar]
- Keller, M.; Kummer, M.; Vasconcelos, M.C. Soil nitrogen utilisation for growth and gas exchange by grapevines in response to nitrogen supply and rootstock. Aust. J. Grape Wine Res. 2001, 7, 2–11. [Google Scholar] [CrossRef]
- Messiga, A.J.; Gallant, K.S.; Sharifi, M.; Hammermeister, A.; Fuller, K.; Tango, M.; Fillmore, S. Grape yield and quality response to cover crops and amendments in a vineyard in Nova Scotia, Canada. Am. J. Enol. Vitic. 2016, 67, 77–85. [Google Scholar] [CrossRef]
- Hébert-Haché, A.; Inglis, D.; Kemp, B.; Willwerth, J.J. Clone and rootstock interactions influence the cold hardiness of Vitis vinifera cvs. Riesling and Sauvignon blanc. Am. J. Enol. Vitic. 2020, 72, 126–136. [Google Scholar] [CrossRef]
- Farhat, N.; Elkhouni, A.; Zorrig, W.; Smaoui, A.; Abdelly, C.; Rabhi, M. Effects of magnesium deficiency on photosynthesis and carbohydrate partitioning. Acta Physiol. Plant 2016, 38, 145. [Google Scholar] [CrossRef]
- Mullins, M.G.; Bouquet, A.; Williams, L.E. Biology of the Grapevine; Cambridge University Press: Cambridge, UK, 1992. [Google Scholar]
- El-Razek, E.A.; Treutter, D.; Saleh, M.M.S.; El-Shamma, M.; Amera, A.F.; Abdel-Hamid, N. Effect of nitrogen and potassium fertilization on productivity and fruit quality of ‘crimson seedless’ grape. Agric. Biol. J. North Am. 2011, 2, 330–340. [Google Scholar] [CrossRef]
- Thomidis, T.; Zioziou, E.; Koundouras, S.; Karagiannidis, C.; Navrozidis, I.; Nikolaou, N. Effects of nitrogen and irrigation on the quality of grapes and the susceptibility to Botrytis bunch rot. Sci. Hortic. 2016, 212, 60–68. [Google Scholar] [CrossRef]
- Baldwin, J.G. The effect of some cultural practices on nitrogen and fruitfulness in the Sultana vine. Am. J. Enol. Vitic. 1966, 17, 58–62. [Google Scholar]
- Vasconcelos, M.C.; Greven, M.; Winefield, C.S.; Trought, M.C.T.; Raw, V. The flowering process of Vitis vinifera: A review. Am. J. Enol. Vitic. 2009, 60, 411–434. [Google Scholar]
- Ewart, A.; Kliewer, W.M. Effects of controlled day and night temperatures and nitrogen on fruit-set, ovule fertility, and fruit composition of several wine grape cultivars. Am. J. Enol. Vitic. 1977, 28, 88–95. [Google Scholar]
- Riesterer-Loper, J.; Workmaster, B.A.; Atucha, A. Impact of fruit zone sunlight exposure on ripening profiles of cold climate interspecific hybrid winegrapes. Am. J. Enol. Vitic. 2019, 70, 286–296. [Google Scholar] [CrossRef]
- Vystavna, Y.; Rätsep, R.; Klymenko, N.; Drozd, O.; Pidlisnyuk, V.; Klymenko, M. Comparison of soil-to-root transfer and translocation coefficients of trace elements in vines of Chardonnay and Muscat white grown in the same vineyard. Sci. Hortic. 2015, 192, 89–96. [Google Scholar] [CrossRef]
- Vystavna, Y.; Zaichenko, L.; Klimenko, N.; Rätsep, R. Trace metals transfer during vine cultivation and winemaking processes. J. Sci. Food Agric. 2017, 97, 4520–4525. [Google Scholar] [CrossRef]
- Maante-Kuljus, M.; Rätsep, R.; Moor, U.; Mainla, L.; Põldma, P.; Koort, A.; Karp, K. Effect of vintage and viticultural practices on the phenolic content of hybrid winegrapes in very cool climate. Agriculture 2020, 10, 169. [Google Scholar] [CrossRef]
- Sweetman, C.; Sadras, V.O.; Hancock, R.D.; Soole, K.L.; Ford, C.M. Metabolic effects of elevated temperature on organic acid degradation in ripening Vitis vinifera fruit. J. Exp. Bot. 2014, 65, 5975–5988. [Google Scholar] [CrossRef] [PubMed]
- Rapp, A.; Knipser, W.; Engel, L.; Ullemeyer, H.; Heimann, W. Off-flavour compounds in the berry and wine aroma of grapevine hybrids 1. The strawberry-like flavour. Vitis 1980, 19, 23. [Google Scholar] [CrossRef]
- Shure, K.B.; Acree, T.E. Changes in the odor-active compounds in Vitis labruscana Cv. Concord during growth and development. J. Agric. Food Chem. 1994, 42, 350–353. [Google Scholar] [CrossRef]
- Slegers, A.; Angers, P.; Pedneault, K. Volatile compounds from must and wines from five white grape varieties. J. Food Chem. Nanotechnol. 2017, 3, 8–18. [Google Scholar] [CrossRef]
- Gutiérrez-Gamboa, G.; Alañón-Sánchez, N.; Mateluna-Cuadra, R.; Verdugo-Vásquez, N. An overview about the impacts of agricultural practices on grape nitrogen composition: Current research approaches. Food Res. Int. 2020, 136, 109477. [Google Scholar] [CrossRef]
- Ugliano, M.; Henschke, P.A.; Herderich, M.J.; Pretorius, I.S. Nitrogen management is critical for wine flavour and style. Aust. New Zeal. Wine Ind. J. 2007, 22, 24–30. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Variety | Species | Season of Planting | Plantation Distance | Plant Characteristics | ||
|---|---|---|---|---|---|---|
| between Rows | between Plants | Clone | Rootstock | |||
| L’Acadie | Vitis interspecific crossing | 2005 | 3.1 | 0.9 | Unknown | Own rooted |
| New York Muscat | Vitis interspecific crossing | 1997 | 3.0 | 1.2 | Unknown | Own rooted |
| Chardonnay | V. vinifera L., ssp. vinifera | 2003–2006 | 3.1 | 1.1 | 76 | 3309 Couderc |
| White Riesling | V. vinifera L., ssp. vinifera | 2013–2015 | 2.6 | 1.4 | 21B | 3309 Couderc |
| Pinot Noir | V. vinifera L., ssp. vinifera | 2003–2006 | 3.1 | 1.1 | 115 | 3309 Couderc |
| Season | Bioclimatic Indices | Frost Free Period | Precipitation mm | |||||
|---|---|---|---|---|---|---|---|---|
| WI | HI | GST (°C) | CI (°C) | Spring | Fall | Frost Free Days | ||
| 2017 | 1169 | 1579 | 14.7 | 9.8 | 22 April 2017 | 10 November 2017 | 202 | 683 |
| 2018 | 1143 | 1535 | 14.2 | 7.2 | 5 June 18 | 24 September 18 | 111 | 898 |
| 2019 | 960 | 1515 | 12.5 | 7.7 | 4 May 2019 | 25 October 19 | 174 | 633 |
| The Timing of Phenological Events (Dates) | The Intervals between Phenological Phases (Days) | ||||||
|---|---|---|---|---|---|---|---|
| Budburst (BB) | Bloom (Bl) | Véraison (Vér) | Harvest (Har) | BB to Bl | Bl to Vér | Vér to Har | |
| Variety | |||||||
| L’Acadie | 23-May ± 0.55 a | 08-July ± 1.05 a | 01-Sept ± 1.32 a | 12-Oct ± 2.11 a | 45.67 a | 55.22 a | 40.89 ab |
| NY Muscat | 25-May ± 0.88 abc | 12-July ± 1.67 ab | 13-Sept ± 2.10 b | 18-Oct ± 3.37 ab | 47.48 ab | 63.08 b | 35.45 a |
| Chardonnay | 25-May ± 0.88 ab | 15-July ± 1.67 b | 9-Sept ± 2.11 b | 22-Oct ± 3.37 bc | 50.98 b | 56.83 ab | 42.95 ab |
| Riesling | 27-May ± 0.67 c | 15-July ± 1.28 b | 12-Sept ± 1.62 b | 28-Oct ± 2.58 c | 48.83 ab | 59.00 ab | 46.17 b |
| Pinot Noir | 27-May ± 0.88 bc | 13-July ± 1.67 b | 11-Sept ± 2.11 b | 20-Oct ± 3.37 abc | 47.48 ab | 59.33 ab | 38.95 ab |
| Seasons | |||||||
| 2017 | 19-May ± 0.50 a | 10-July ± 0.95 a | 06-Sept ± 1.20 a | 18-Oct ± 1.92a | 51.77 b | 57.59 a | 41.73 a |
| 2018 | 20-May ± 0.85 a | 12-July ± 1.62 a | 05-Sept ± 2.04 a | 15-Oct ± 3.26 a | 52.56 b | 55.36 a | 40.28 a |
| 2019 | 06-Jun ± 0.50 b | 16-July ± 0.96 b | 17-Sept ± 1.20 b | 28-Oct ± 1.92 b | 39.95 a | 63.13 b | 40.64 a |
| Significance | |||||||
| Variety | 0.0027 | 0.0034 | 0.0002 | 0.0023 | 0.2134 | 0.1901 | 0.155 |
| Season | <0.0001 | 0.0017 | <0.0001 | 0.0015 | <0.0001 | 0.0204 | 0.89 |
| N (%) | Ca (%) | K (%) | Mg (%) | P (%) | Na (%) | B (ppm) | Fe (ppm) | Zn (ppm) | |
|---|---|---|---|---|---|---|---|---|---|
| Variety (V) | |||||||||
| L’Acadie | 1.08 ab | 1.34 ab | 1.34 a | 0.62 c | 0.37 b | 0.03 a | 30.30 a | 46.18 ab | 32.09 a |
| NY Muscat | 0.71 a | 1.19 a | 1.68 a | 0.29 a | 0.19 a | 0.04 a | 27.18 a | 31.21 a | 48.73 a |
| Chardonnay | 1.57 b | 1.26 ab | 1.31 a | 0.41 ab | 0.26 ab | 0.03 a | 36.11 b | 48.46 ab | 58.61 a |
| Riesling | 1.28 b | 1.31 ab | 2.56 b | 0.46 b | 0.37 b | 0.03 a | 41.71 c | 54.92 ab | 48.61 a |
| Pinot Noir | 1.48 b | 1.38 b | 1.59 a | 0.54 bc | 0.19 a | 0.03 a | 35.68 b | 76.02 b | 50.41 a |
| Seasons (S) | |||||||||
| 2017 | 0.87 a | 1.41 b | 1.39 a | 0.47 a | 0.26 a | 0.04 b | 34.51 ab | 46.58 a | 60.86 b |
| 2018 | 1.10 a | 1.23 a | 1.87 a | 0.46 a | 0.32 a | 0.02 a | 32.12 a | 42.74 a | 44.71 ab |
| 2019 | 1.70 b | 1.26 a | 1.82 a | 0.46 a | 0.25 a | 0.04 b | 35.96 b | 64.76 a | 37.49 a |
| Significance | |||||||||
| V | 0.0133 | 0.1159 | 0.0336 | 0.0012 | 0.0059 | 0.4828 | <0.0001 | 0.256 | 0.28 |
| S | 0.0005 | 0.0096 | 0.2597 | 0.9886 | 0.2392 | 0.016 | 0.0442 | 0.2865 | 0.0793 |
| V × S | 0.1773 | 0.5852 | 0.856 | 0.9445 | 0.3361 | 0.1089 | 0.2673 | 0.2737 | 0.8549 |
| Bunch Weight (g) | Weight of 200 Berries (g) | Yield Per Plant (kg) | 𝜹13C | ΨStem (Mpa) | |
|---|---|---|---|---|---|
| Variety | |||||
| L’Acadie | 86.09 a | 289.60 a | 1.05 a | −27.82 a | −0.62 a |
| NY Muscat | 132.19 b | 598.565 c | 1.25 ab | −28.58 a | −0.60 a |
| Chardonnay | 191.13c | 344.44 b | 2.06 ab | −27.76 a | −0.40 a |
| Riesling | 136.06 b | 325.40 ab | 2.13 b | −28.94 a | −0.46 a |
| Pinot Noir | 110.91 ab | 297.72 ab | 1.07 a | −28.22 a | −0.45 a |
| Seasons | |||||
| 2017 | 133.12 a | 384.68 b | 2.26 b | −28.55 a | −0.41 a |
| 2018 | 137.75 a | 402.43 b | 0.88 a | −27.87 a | −0.40 a |
| 2019 | 122.96 a | 326.33 a | 1.40 a | −28.37 a | −0.71 b |
| Significance | |||||
| Variety | <0.0001 | <0.0001 | 0.0285 | 0.0531 | 0.1971 |
| Season | 0.3928 | 0.0009 | 0.0058 | 0.2982 | 0.004 |
| °Brix | Total Acidity (g L−1 H2SO4) | pH | Malic Acid (g L−1) | YAN (mg L−1) | Organic Nitrogen (mgL−1) | Mineral Nitrogen (mg L−1) | IBMP | |
|---|---|---|---|---|---|---|---|---|
| Variety | ||||||||
| L’Acadie | 17.10 a | 6.08 ab | 2.95 b | 4.71 a | 176.33 a | 156.44 ab | 20.22 a | 0.00 a |
| NY Muscat | 16.86 a | 4.60 a | 3.14 c | 4.21 a | 207.55 a | 188.03 bc | 19.42 a | 0.00 a |
| Chardonnay | 18.09 a | 7.48 bc | 2.96 b | 7.06 b | 301.55 b | 233.78 c | 67.92 b | 0.00 a |
| Riesling | 17.20 a | 8.24 c | 2.74 a | 6.07 b | 182.50 a | 121.17 a | 60.83 b | 0.78 a |
| Pinot Noir | 18.14 a | 7.24 bc | 3.00 b | 7.15 b | 312.80 b | 241.48 c | 71.00 b | 6.95 b |
| Seasons | ||||||||
| 2017 | 18.33 b | 5.06 a | 2.99 a | 4.24 a | 148.35 a | 119.70 a | 28.88 a | 1.89 a |
| 2018 | 17.90 b | 7.36 b | 2.93 a | 6.41 b | 247.75 b | 190.25 b | 57.21 b | 1.54 a |
| 2019 | 16.20 a | 7.77 b | 2.96 a | 6.87 b | 312.35 c | 254.59 c | 57.54 b | 1.34 a |
| Significance | ||||||||
| Variety | 0.5781 | 0.0006 | <0.0001 | 0.001 | 0.0017 | 0.0037 | <0.0001 | 0.0007 |
| Season | 0.0063 | 0.0001 | 0.4458 | 0.0001 | <0.0001 | <0.0001 | 0.0007 | 0.7449 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Diez-Zamudio, F.; Laytte, R.; Grallert, C.; Ivit, N.N.; Gutiérrez-Gamboa, G. Viticultural Performance of Hybrids and Vitis vinifera Varieties Established in Annapolis Valley (Nova Scotia). Horticulturae 2021, 7, 291. https://doi.org/10.3390/horticulturae7090291
Diez-Zamudio F, Laytte R, Grallert C, Ivit NN, Gutiérrez-Gamboa G. Viticultural Performance of Hybrids and Vitis vinifera Varieties Established in Annapolis Valley (Nova Scotia). Horticulturae. 2021; 7(9):291. https://doi.org/10.3390/horticulturae7090291
Chicago/Turabian StyleDiez-Zamudio, Francisco, Rodrigo Laytte, Cecilia Grallert, Nedret Neslihan Ivit, and Gastón Gutiérrez-Gamboa. 2021. "Viticultural Performance of Hybrids and Vitis vinifera Varieties Established in Annapolis Valley (Nova Scotia)" Horticulturae 7, no. 9: 291. https://doi.org/10.3390/horticulturae7090291
APA StyleDiez-Zamudio, F., Laytte, R., Grallert, C., Ivit, N. N., & Gutiérrez-Gamboa, G. (2021). Viticultural Performance of Hybrids and Vitis vinifera Varieties Established in Annapolis Valley (Nova Scotia). Horticulturae, 7(9), 291. https://doi.org/10.3390/horticulturae7090291