
Peanut Root Exudates Suppress Fusarium solani and Modulate the Microbial Community Structure of Rhizosphere in Grape Replant Soil


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.1.1. Plant Materials
2.1.2. Soil Materials
2.1.3. Strain Material
2.2. Experimental Design
2.2.1. Collection of Peanut Root Secretion
2.2.2. Peanut Root Exudates Treatment in Grape Replant Soil
2.2.3. Pot Experiment
2.3. The Effect of Peanut Secretions on the Growth of Fusarium solani
2.4. Soil Bacterial and Fungal Community Analyses
2.5. Determination of Physiological Indexes
2.6. Statistical Analysis
3. Results
3.1. Effects of Peanut Root Exudates on the Growth and Physiological Index of Grapevine Seedlings
3.2. Effects of Peanut Root Exudates on the Growth of Harmful Fungus (Fusarium solanum)
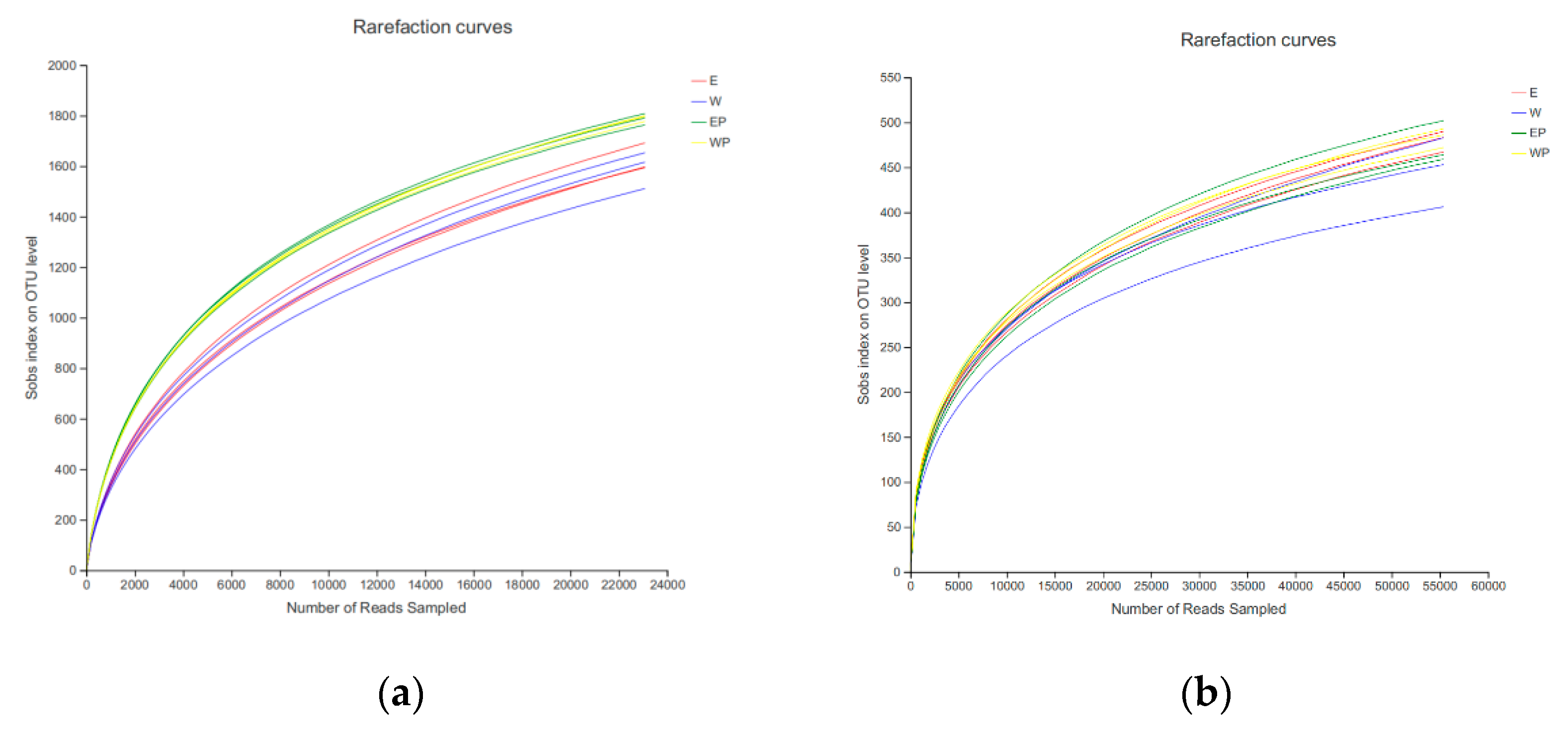
3.3. Composition and Diversity of Bacteria and Fungi in Grapevine Replanted Soil following Application of Peanut Root Exudates
3.4. Bacteria and Fungi Co-Occurrence Network
4. Discussion
4.1. Peanut Root Exudates Can Promote the Growth of Grapevine Seedlings in Replanting Soil
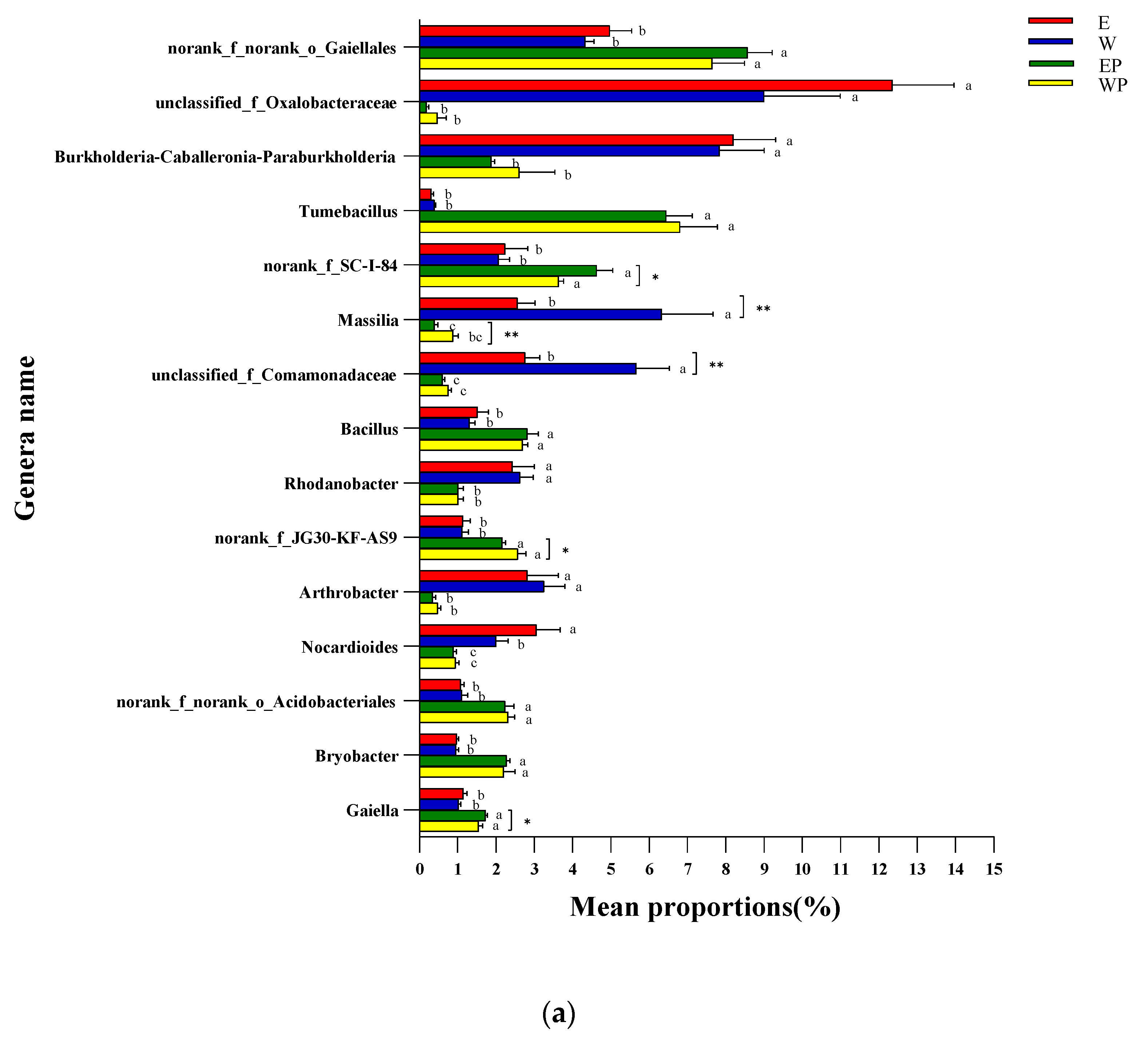
4.2. Root Exudates from Peanuts Can Affect the Compositions and Functions of Bacteria and Fungi in Soil
4.3. The Peanut Root Exudate Changed the Topological Properties of Bacterial and Fungal Co-Occurrence Networks
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Rao, D.; Liu, P.Y.; Zou, L.Y.; Teng, Y.; Yu, H.Y. Microbial dysbiosis together with nutrient imbalance cause the replant problem of upper six flue-cured tobacco in Central Henan. J. Plant Dis. Prot. 2021, 128, 1487–1500. [Google Scholar] [CrossRef]
- Zhang, B.; Li, X.Z.; Wang, F.Q.; Li, M.J.; Zhang, J.Y.; Gu, L.; Zhang, L.J.; Tu, W.Q.; Zhang, Z.Y. Assaying the potential autotoxins and microbial community associated with Rehmannia glutinosa replant problems based on its ‘autotoxic circle’. Plant Soil 2016, 407, 307–322. [Google Scholar] [CrossRef]
- Zhang, Z.Z.; Zhang, Z.D.; Han, X.Y.; Wu, J.H.; Zhang, L.Z.; Wang, J.R.; Gefu, W.P. Specific response mechanism to autotoxicity in melon (Cucumis melo L.) root revealed by physiological analyses combined with transcriptome profiling. Ecotoxicol. Environ. Saf. 2020, 200, 110779. [Google Scholar] [CrossRef] [PubMed]
- Li, X.G.; Ding, C.F.; Hua, K.; Zhang, T.L.; Zhang, Y.N.; Zhao, L.; Yang, Y.R.; Liu, J.G.; Wang, X.X. Soil sickness of peanuts is attributable to modifications in soil microbes induced by peanut root exudates rather than to direct allelopathy. Soil Biol. Biochem. 2014, 78, 149–159. [Google Scholar] [CrossRef]
- Zhao, Y.P.; Lin, S.; Chu, L.X.; Gao, J.T.; Azeem, S.; Lin, W.X. Insight into structure dynamics of soil microbiota mediated by the richness of replanted Pseudostellaria heterophylla. Sci. Rep. 2016, 6, 26175. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.J.; Chen, S.C.; Zhang, X.X.; Zhou, X.G.; Wu, F.Z. Rhizosphere bacterial community in watermelon-wheat intercropping was more stable than in watermelon monoculture system under Fusarium oxysporum f. sp. niveum invasion. Plant Soil Int. J. Plant Soil 2019, 445, 369–381. [Google Scholar] [CrossRef]
- Wu, H.M.; Lin, M.H.; Christopher, R.; Qin, X.J.; Zhang, S.K.; Chen, J.; Wu, L.K.; Zhao, Y.L.; Lin, S.; Lin, W.X. Plant-mediated rhizospheric interactions in intraspecific intercropping alleviate the replanting disease of Radix pseudostellariae. Plant Soil 2020, 454, 411–430. [Google Scholar] [CrossRef]
- Li, X.G.; Wietse, D.B.; Zhang, Y.N.; Ding, C.F.; Zhang, T.L.; Wang, X.X. Suppression of soil-borne Fusarium pathogens of peanut by intercropping with the medicinal herb Atractylodes lancea. Soil Biol. Biochem. 2018, 116, 120–130. [Google Scholar] [CrossRef]
- Yang, Y.; Wu, F.Z.; Liu, S.W. Allelopathic effects of root exudates of Chinese onion accessions on cucumber yield and Fusarium oxysporum f. sp. cucumerinum. Allelopath. J. 2011, 27, 75–85. [Google Scholar]
- Yuan, X.F.; Wang, B.B.; Hong, S.; Xiong, W.; Shen, Z.Z.; Ruan, Y.Z.; Li, R.; Shen, Q.R.; Dini, A.F. Promoting soil microbial-mediated suppressiveness against Fusarium wilt disease by the enrichment of specifc fungal taxa via crop rotation. Biol. Fertil. Soils 2021, 57, 1137–1153. [Google Scholar] [CrossRef]
- Zhou, X.G.; Liu, J.; Wu, F.Z. Soil microbial communities in cucumber monoculture and rotation systems and their feedback effects on cucumber seedling growth. Plant Soil 2017, 415, 507–520. [Google Scholar] [CrossRef]
- Ma, S.C.; Wang, T.C.; Guan, X.K.; Zhang, X. Effect of sowing time and seeding rate on yield components and water use efficiency of winter wheat by regulating the growth redundancy and physiological traits of root and shoot. Field Crop. Res. 2018, 221, 166–174. [Google Scholar] [CrossRef]
- Li, H.S.; Sun, Q.; Zhao, S.J.; Zhang, W.H. Principles and Techniques of Plant Physiological and Biochemical Experiments; Higher Education Press: Beijing, China, 2000. [Google Scholar]
- Liu, Q.W.; Li, K.; Guo, X.W.; Ma, L.; Guo, Y.S.; Liu, Z.D. Developmental characteristics of grapevine seedlings root border cells and their response to ρ-hydroxybenzoic acid. Plant Soil 2019, 443, 199–218. [Google Scholar] [CrossRef]
- Guo, X.W.; Li, K.; Xie, H.G.; Sun, Y.N.; Hu, X.X.; Zhang, L.H. Effect of sterilized replant soil on grape growth and root exudation characteristics. J. Fruit Sci. 2010, 27, 29–33. (In Chinese) [Google Scholar]
- Zhao, F.Y.; Zhang, Y.Y.; Dong, W.G.; Zhang, Y.Q.; Zhang, G.X.; Sun, Z.P.; Yang, L.J. Vermicompost can suppress Fusarium oxysporum f. sp. lycopersici via generation of beneficial bacteria in a long-term tomato monoculture soil. Plant Soil 2019, 440, 491–505. [Google Scholar] [CrossRef]
- Ding, Y.R.; Chen, Y.L.; Lin, Z.Q.; Tuo, Y.Y.; Li, H.L.; Wang, Y. Differences in soil microbial community composition between suppressive and root rot-conducive in tobacco fields. Curr. Microbiol. 2021, 78, 624–633. [Google Scholar] [CrossRef]
- Kloepper, J.W.; Ryu, C.M.; Zhang, S.A. Induced systemic resistance and promotion of plant growth by Bacillus spp. Phytopathology 2004, 94, 1259–1266. [Google Scholar] [CrossRef]
- Zhang, M.Y.; Muhammad, R.; Zhang, L.; El-Desouki, Z.; Jiang, C.C. Biochar induces changes to basic soil properties and bacterial communities of different soils to varying degrees at 25 mm rainfall: More effective on acidic soils. Front. Microbiol. 2019, 10, 1321. [Google Scholar] [CrossRef]
- Zheng, B.X.; Bi, Q.F.; Hao, X.L.; Zhou, G.W.; Yang, X.R. Massilia phosphatilytica sp. nov., a phosphate solubilizing bacteria isolated from a long-term fertilized soil. Int. J. Syst. Evol. Microbiol. 2017, 67, 2514–2519. [Google Scholar] [CrossRef]
- Lou, J.; Gu, H.P.; Wang, H.Z.; An, Q.L.; Xu, J.M. Complete genome sequence of Massilia sp. WG5, an efficient phenanthrene-degrading bacterium from soil. J. Biotechnol. 2016, 218, 49–50. [Google Scholar] [CrossRef]
- Jara, E.; Morel, M.A.; Lamolle, G.; Castro-Sowinski, S.; Simón, D.; Iriarte, A.; Musto, H. The complex pattern of codon usage evolution in the family Comamonadaceae. Ecol. Genet. Genom. 2018, 6, 1–8. [Google Scholar] [CrossRef]
- Yu, J.Y.; Liu, Y.H.; Wang, Z.Y.; Huang, X.H.; Chai, D.; Gu, Y.F.; Zhao, K.; Yu, X.M.; Shuai, Z.B.; Liu, H.J.; et al. The cropping obstacle of garlic was associated with changes in soil physicochemical properties, Enzymatic activities and bacterial and fungal communities. Front. Microbiol. 2022, 13, 828196. [Google Scholar] [CrossRef] [PubMed]
- Han, P.P.; Zhang, X.P.; Xu, D.; Zhang, B.W.; Lai, D.W.; Zhou, L.G. Metabolites from Clonostachys Fungi and their biological activities. J. Fungi 2020, 6, 229. [Google Scholar] [CrossRef] [PubMed]
- Gräfenhan, T.; Schroers, H.J.; Nirenberg, H.I.; Seifert, K.A. An overview of the taxonomy, phylogeny, and typifcation of nectriaceous fungi in Cosmospora, Acremonium, Fusarium, Stilbella, and Volutella. Stud. Mycol. 2011, 68, 79–113. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.J.; David, M.G.; Robert, H.P.; Alejandro, P.R.; Kerry, O.D.; Frances, T.; Donald, M.G.; John, M.M.; Kemal, K. Fusarium pathogenomics. Annu. Rev. Microbiol. 2013, 67, 399–416. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.W.; Wang, S.X.; Li, K.; Qiao, J.; Guo, Y.S.; Liu, Z.D.; Guo, X.W. Responses of soil bacterial and fungal communities to the long-term monoculture of grapevine. Appl. Microbiol. Biotechnol. 2021, 105, 7035–7050. [Google Scholar] [CrossRef]
- Chen, Q.; Jiang, J.R.; Zhang, G.Z.; Cai, L.; Crous, P.W. Resolving the Phoma enigma. Stud. Mycol. 2015, 82, 137–217. [Google Scholar] [CrossRef]
- Margesin, R.; Ludwikowski, T.M.; Kutzner, A.; Wagner, A.O. Low-temperature biodegradation of lignin-derived aromatic model monomers by the cold-adapted yeast Rhodosporidiobolus colostri isolated from alpine forest soil. Microorganisms 2022, 10, 515. [Google Scholar] [CrossRef]
- Ozimek, E.; Hanaka, A. Mortierella species as the plant growth-promoting fungi present in the agricultural soils. Agriculture 2021, 11, 7. [Google Scholar] [CrossRef]
- Eroshin, V.; Dedyukhina, E. Effect of lipids from Mortierella hygrophila on plant resistance to phytopathogens. World J. Microbiol. Biotechnol. 2002, 18, 165–167. [Google Scholar] [CrossRef]
- Zhou, D.B.; Tao, J.; Chen, Y.F.; Wang, F.; Qi, D.F.; Feng, R.J.; Xie, J.H.; Li, H.P. Deciphering microbial diversity associated with Fusarium wilt-diseased and disease-free banana rhizosphere soil. BMC Microbiol. 2019, 19, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Xiong, W.; Li, R.; Ren, Y.; Liu, C.; Zhao, Q.Y.; Wu, H.S.; Jousset, A.; Shen, Q.R. Distinct roles for soil fungal and bacterial communities associated with the suppression of vanilla Fusarium wilt disease. Soil Biol. Biochem. 2017, 107, 198–207. [Google Scholar] [CrossRef]
- Sobrino-Plata, J.; Martinez-Arias, C.; Ormeno-Moncalvillo, S.; Fernandez, I.; Collada, C.; Gil, L.; Pieterse, C.M.J.; Martin, J. No priming, just fighting–endophytic yeast attenuates the defense response and the stress induced by Dutch elm disease in Ulmus minor Mill. Tree Physiol. 2022, tpac062. [Google Scholar] [CrossRef]
- Shi, S.J.; Nuccio, E.E.; Shi, Z.J.; He, Z.L.; Zhou, J.Z.; Firestone, M.K. The interconnected rhizosphere: High network complexity dominates rhizosphere assemblages. Ecol. Lett. 2016, 19, 926–936. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.H.; Tariq, A.; Zeng, F.J.; Graciano, C.; Sun, F.; Chai, X.T.; Ahmed, Z. Nitrogen and water addition regulate fungal community and microbial co-occurrence network complexity in the rhizosphere of Alhagi sparsifolia seedlings. Appl. Soil Ecol. 2021, 164, 103940. [Google Scholar] [CrossRef]
- Liu, X.; Yang, T.; Shi, Y.; Zhu, Y.C.; He, M.L.; Zhao, Y.K.; Adams, J.M.; Chu, H.Y. Strong partitioning of soil bacterial community composition and co-occurrence networks along a small-scale elevational gradient on Zijin Mountain. Soil Ecol. Lett. 2021, 3, 290–302. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Sample | pH | Organic Matter (%) | Available N (mg/kg) | Available P (mg/kg) | Available K (mg/kg) |
|---|---|---|---|---|---|
| Replant soil | 6.50 ± 0.0176 | 1.39 ± 0.0353 | 35.53 ± 0.3351 | 138.69 ± 2.3902 | 11.03 ± 0.3283 |
| Treatment | Shannon | Simpson | Ace | Chao 1 |
|---|---|---|---|---|
| E | 5.359 ± 0.0904 b | 0.024 ± 0.0026 a | 2323.497 ± 40.0125 a | 2295.178 ± 33.8677 a |
| W | 5.384 ± 0.0684 b | 0.019 ± 0.0013 a | 2539.307 ± 42.0133 a | 2274.975 ± 32.9005 a |
| EP | 6.071 ± 0.0309 a | 0.007 ± 0.0004 b | 2287.683 ± 38.9391 a | 2302.916 ± 41.6874 a |
| WP | 6.066 ± 0.0116 a | 0.008 ± 0.0004 b | 2338.919 ± 43.2459 a | 2381.215 ± 42.2063 a |
| Treatment | Shannon | Simpson | Ace | Chao 1 |
|---|---|---|---|---|
| E | 3.627 ± 0.0184 a* | 0.052 ± 0.0015 c | 667.250 ± 45.5907 a | 628.231 ± 36.7562 a |
| W | 3.379 ± 0.0746 a | 0.077 ± 0.0077 bc* | 612.745 ± 83.0068 a | 575.227 ± 51.2110 a |
| EP | 2.996 ± 0.0383 b | 0.149 ± 0.0012 a | 599.619 ± 20.8751 a | 583.743 ± 16.4071 a |
| WP | 3.347 ± 0.0951 a* | 0.104 ± 0.0136 b | 604.929 ± 2.6914 a | 593.883 ± 6.3941 a |
| Network Metrics | E | W | EP | WP | |
|---|---|---|---|---|---|
| Bacteria | Number of nodes | 152 | 151 | 199 | 189 |
| Total number of edges | 3449 | 3184 | 5768 | 5262 | |
| Number of positive correlations | 1712 | 1602 | 2877 | 2599 | |
| Number of negative correlations | 1737 | 1582 | 2891 | 2663 | |
| Ratio of positive to negative correlations | 0.986 | 1.013 | 0.995 | 0.976 | |
| Network density | 0.301 | 0.281 | 0.293 | 0.296 | |
| Network diameter | 1 | 1 | 1 | 1 | |
| Connected components | 6 | 8 | 7 | 7 | |
| Clustering coefficient | 0.987 | 0.960 | 1 | 0.989 | |
| Average number of neighbors | 45.382 | 42.172 | 57.970 | 55.683 | |
| Fungi | Number of nodes | 79 | 63 | 64 | 75 |
| Total number of edges | 1038 | 658 | 665 | 937 | |
| Number of positive correlations | 530 | 382 | 416 | 454 | |
| Number of negative correlations | 508 | 276 | 249 | 483 | |
| Ratio of positive to negative correlations | 1.04 | 1.38 | 1.67 | 0.94 | |
| Network density | 0.337 | 0.337 | 0.330 | 0.338 | |
| Network diameter | 1 | 1 | 1 | 1 | |
| Connected components | 3 | 3 | 3 | 3 | |
| Clustering coefficient | 1 | 1 | 1 | 1 | |
| Average number of neighbors | 26.278 | 20.889 | 20.781 | 24.987 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Liu, Q.; Li, K.; Ma, L. Peanut Root Exudates Suppress Fusarium solani and Modulate the Microbial Community Structure of Rhizosphere in Grape Replant Soil. Horticulturae 2022, 8, 892. https://doi.org/10.3390/horticulturae8100892
Zhang J, Liu Q, Li K, Ma L. Peanut Root Exudates Suppress Fusarium solani and Modulate the Microbial Community Structure of Rhizosphere in Grape Replant Soil. Horticulturae. 2022; 8(10):892. https://doi.org/10.3390/horticulturae8100892
Chicago/Turabian StyleZhang, Jiale, Qianwen Liu, Kun Li, and Li Ma. 2022. "Peanut Root Exudates Suppress Fusarium solani and Modulate the Microbial Community Structure of Rhizosphere in Grape Replant Soil" Horticulturae 8, no. 10: 892. https://doi.org/10.3390/horticulturae8100892
APA StyleZhang, J., Liu, Q., Li, K., & Ma, L. (2022). Peanut Root Exudates Suppress Fusarium solani and Modulate the Microbial Community Structure of Rhizosphere in Grape Replant Soil. Horticulturae, 8(10), 892. https://doi.org/10.3390/horticulturae8100892