1. Introduction
Recently, a growing awareness of the environmental impact of food production (in terms of pollution, greenhouse gas emissions, soil depletion, biodiversity loss, etc.), has led to the definition of “sustainable food quality” [
1]. A more sustainable food production system involves using resources at a pace that can be tolerated and ultimately, completely replenished by our environment. Consequently, a “sustainable food quality” should embrace various issues, including safety, affordability and nutritional and functional values, while controlling the use of chemical fertilizers, herbicides and pesticides by exploiting natural plant defenses and biodiversity [
2]. In this perspective, the use of biostimulants is gaining more and more popularity thanks to the possibility of application in more sustainable production systems, which at the same time allows for increasing the yield and quality of the product. Biostimulants can consist of organic substances (e.g., humic acids, algae or plant extracts, protein hydrolysates, chitosan, vitamins), inorganic compounds (e.g., cobalt, silica, selenium) or plant growth promoting microorganisms or their extracts (e.g., fungi and bacteria) [
3]. They are not classified as fertilizers or pesticides, but they can increase resource use efficiency, growth and yield and resilience and tolerance to abiotic stresses when applied to plants [
4]. Plant biostimulants act mainly on plants by inducing multiple direct and indirect physiological effects, which are linked, just to name a few, to the greater mobility and solubility of mineral nutrients in the soil, changes in the architecture of the root system, better efficiency in water use and ion uptake, mobilization and utilization [
5]. Regarding the composition and properties of plant foods, biostimulants can increase the synthesis and accumulation of primary and secondary metabolites, including important categories of antioxidants, such as carotenoids, polyphenols and ascorbic acid [
6], thus ultimately improving the nutritional and nutraceutical quality of edible products.
In this study, we focused on three types of biostimulants, two of which are of plant origin, (a tropical plant extract and a commercial legume derived-plant hydrolysate) and a microbial-based biostimulant containing
Trichoderma harzianum strain T22, an endophytic fungus, was applied to two varieties of tomato (San Marzano and Datterino) and spinach. Both tomato (
Solanum lycopersicum L.) and spinach (
Spinacia oleracea L.) are among the most widely grown vegetables, are globally consumed both fresh and in a variety of processed products, and are considered a powerhouse for nutrients and are low in calories [
7]. The San Marzano tomato is a traditional variety grown in the Campania region in southern Italy; indigenous tomato varieties may have interesting traits such as stress resistance and high-quality fruit [
8]. Spinach is a vegetable with a high biological value, but is prone to the accumulation of nitrates, which is considered potentially dangerous because they are related to methemoglobinemia in children (blue baby syndrome) and the synthesis of N-nitroso compounds (NOCs), classified as “probably carcinogenic to humans” [
9]. Biostimulants may also lower the nitrate content because they are able to improve their reductive assimilation, thus limiting their accumulation in the leaves. In many experimental tests, the use of biostimulants has allowed for keeping nitrate below the maximum residue levels (MRLs) [
10].
Many studies have focused their attention on the capacity of biostimulants to decrease the nitrate content in leafy vegetables or increase the content of beneficial such as K, Ca, and Mg; however, this study, for the first time, investigated the effects of different biostimulants on the concentration of essential macro- and micronutrients (P, K, Ca, Mg, Cu, Fe, Mn, Zn, Se) in San Marzano and Datterino tomatoes and spinach.
3. Results and Discussion
The macro- and micronutrients concentration in horticultural crops is influenced by genetic and environmental factors, whose interaction drives variation in plant traits [
20]. Recent studies have proven that plant biostimulants (PB), including plant-derived extracts or hydrolysates and microbial biostimulants, can improve nutrient use efficiency (NUE) in plants [
21,
22,
23,
24]. In particular, PB may boost macro- and micronutrients’ uptake and assimilation, mainly because of changes in root architecture, including increases in length, density, surface area, number of lateral roots and root exudation of low-molecular-weight metabolites able to promote the formation of micro-aggregates enhancing microbial activity and, in particular, nitrogen and phosphorus uptake [
25,
26]. In our previous studies, we focused our attention on the capacity of biostimulants to increase the content of beneficial ions such as K, Ca and Mg or decrease the content of the antinutrient nitrate in leafy vegetables [
2,
27].
In this study, for the first time, we show preliminary results on the effects of different biostimulants not only on the content of essential macronutrients (P, K, Ca and Mg) but also on micro-nutrients (Cu, Fe, Mn, Zn and Se) in tomato fruits and spinach.
In particular, the analysis of microelements in Pixel cherry tomato fruits (
Table 1) showed that, regardless of treatment, Fe was the most abundant in micronutrients (122.30 µg 100g
−1 fw, on average), followed by Zn (16.93 µg 100g
−1 fw, on average), Cu (9.46 µg 100g
−1 fw, on average), Mn (8.51 µg 100g
−1 fw, on average) and Se (0.015 µg 100g
−1 fw, on average). The application of the T22 biostimulant significantly increased, compared to control, the concentration of all micronutrients analyzed by ICP-OES. Specifically, an increase of 49.66, 38.68, 129.79, 64.03 and 72.72% was recorded for Fe, Zn, Cu, Mn and Se, respectively. Similar results were also observed in
Lens culinaris [
28],
Triticum aestivum [
29],
Lupinus L [
30] and
Cucurbita pepo [
31] treated with
Trichoderma, which, as supported by [
32] through the mechanisms of chelation and reduction, would have allowed for better solubilization of these mineral compounds, including Fe
2O
3, MnO
2 and Zn.
T22 biostimulant treatment resulted in significant differences in EDI and NC for all micronutrients analyzed (
Table 2). Regardless of treatment, Fe showed the highest EDI and NC values, which averaged 244.6 µg die
−1 and 1.63%, respectively. Compared to control, biostimulant application increased the EDI values in all microelements. The NC values of Fe, Zn, Cu, Mn and Se increased by 49.61, 36.84, 131.57, 61.53 and 70%, respectively.
Regardless of the treatment, among the five micronutrients quantified in San Marzano tomato fruits (
Table 3), the most abundant were Fe, followed by Mn, Zn, Cu and Se (39.63 µg 100 g
−1 fw > 21.24 µg 100 g
−1 fw > 14.15 100 g
−1 fw, 1.39 µg 100 g
−1 fw, and 0.19 µg 100 g
−1 fw, on average, respectively), unlike what was observed in Tomato Pixel (
Table 2), highlighting the effect of genotypes on mineral bioaccumulation in fruits [
13]. For all micronutrients, the application of the TPE biostimulant resulted in a significant difference with
p ≤ 0.01. Specifically, the concentrations of Fe, Mn, Zn, Cu and Se in tomato fruits treated with the biostimulant were 128.87, 158.26, 181.91, 119.54 and 131, 30% higher than in control, similar to the findings of a previous study by [
33].
The same authors [
33] assessed that the positive effects of plant biostimulants on the mineral profile are not attributable to the micronutrients in commercial formulations (present in low concentrations) but to their role as direct promoters of root uptake. It is possible that biostimulants may influence active soil microorganisms [
34], thus changing the soil texture and structure, which influence the nutrients’ availability from soil to plant roots [
35]. Billard and coworkers [
36] suggest that such positive effects on the bioaccumulation of the mineral profile in leaves may also be attributable to the improved translocation of minerals from roots to shoots. This latter finding is confirmed by the fact that nutrient concentrations in plant tissues are known to fluctuate together [
37], therefore a biostimulation of translocation capacity may improve in general the use efficiency of most of the nutrients. In addition, since Zn has limited mobility in leaves [
38], the fact that it increased in tomato fruits means that translocation from leaves to fruit was also enhanced.
The results reported in
Table 4 show that the EDI and NC of all micronutrients were significantly affected by the biostimulant treatment. Compared with control, the application of biostimulant on tomato fruits increased EDI-Fe by 128.87%, EDI-Mn by 158.26%, EDI-Zn by 181.91%, EDI-Cu by 119.54% and EDI-Se by 131.30%. The same trend was observed for the NC of all micronutrients. Specifically, biostimulant treatment resulted in the highest values of NC-Mn (1.23%), NC-Se (0.97%), NC-Fe (0.74%), NC-Zn (0.28%) and NC-Cu (0.19%).
The results for the concentration of the five micronutrients in spinach leaves, reported in
Table 5, show that, regardless of biostimulant treatment, the most abundant microelement was Fe, followed by Zn, Mn, Cu and Se, with average values of 346.15, 194.3, 117.43, 21.22 and 0.176 µg 100 g
−1 fw, respectively. In line with other studies [
21,
39,
40] reporting that the VDPD improves the uptake of macro- and micronutrients by plants, compared with control, the foliar application of VDPH increased the concentration of all micronutrients which are all essential for human health [
41,
42]. Specifically, the highest percentage increase was recorded for Se (224.09%) followed by Zn (127.10%), Mn (125.90%), Fe (102.70%) and Cu (68.50%).
Remarkably, the latter element (Cu) plays a key role in the course of infectious disease caused by the SARS-CoV-2 virus (COVID-19) [
43].
As shown in
Table 6, VDPH significantly influenced spinach’s EDI and NC of micronutrients. Compared to control, spinach plants treated with the biostimulant increased EDI-Fe, EDI-Zn, EDI-Mn, EDI-Cu and EDI-Se by 98.98, 127.09, 125.93, 68.52 and 230.76%, respectively. Biostimulant treatment resulted in the highest values in NC-Fe (5.11%), NC-Zn (2.88%), NC-Mn (5.21%), NC-Cu (2.13%) and NC-Se (0.784%).
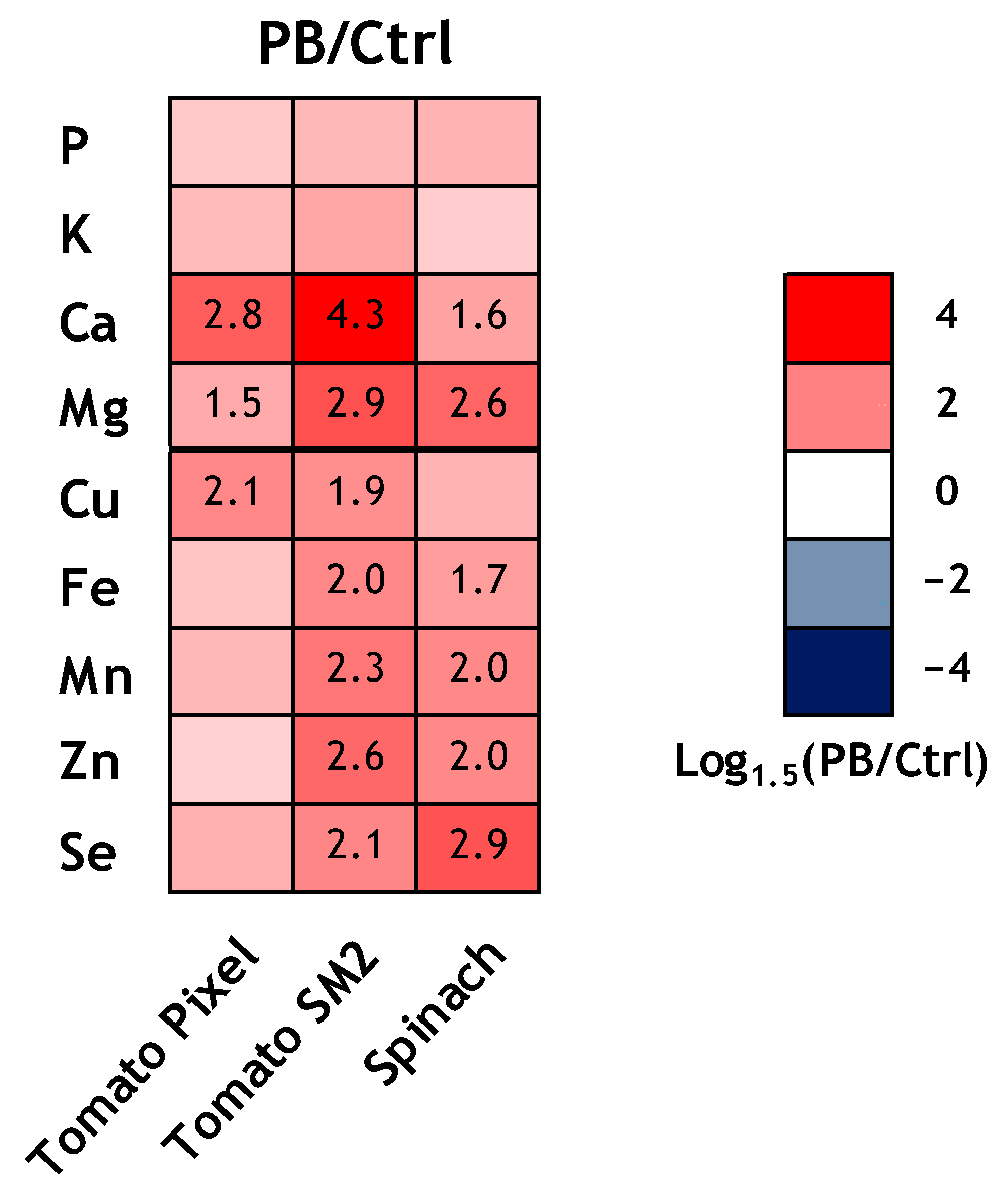
As shown by the heat map analysis (
Figure 1), all the biostimulant treatments induced an increase of the content of the macro- and micronutrients. In particular, San Marzano under TPE and spinach under VDPH treatment underwent the most noteworthy increase of micronutrients, except for Cu, in addition to Mg, whereas Ca and Cu highly increased under T22 and TPE treatments.

 ,
, 

{kind=link}