
Establishment of In Vitro Regeneration Protocol for Sabah’s Jewel Orchid, Macodes limii J.J. Wood & A.L. Lamb

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Establishment of In Vitro Plantlets
2.2. Effects of Node Position and PGRs
2.3. Rooting and Acclimatization of Plantlets
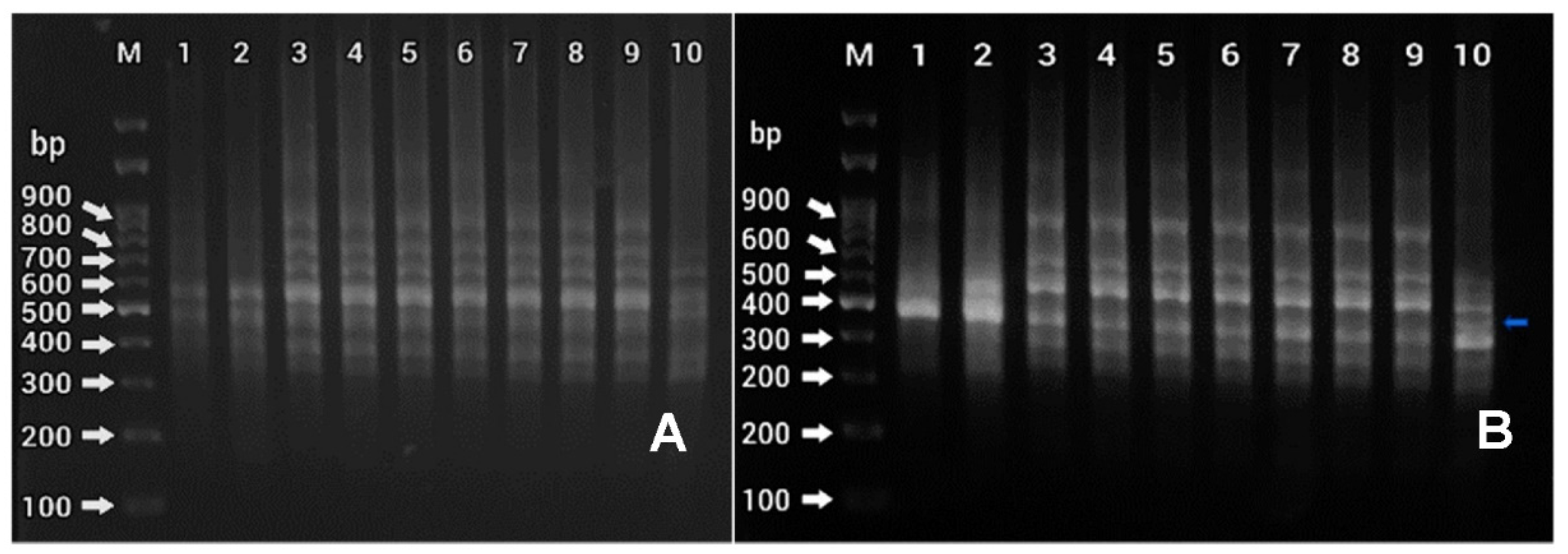
2.4. Analysis of Genetic Fidelity Using ISSR Primers
2.5. Histology and Scanning Electron Microscopy (SEM) of Axillary Shoot Induced from Nodal Explant
2.6. Statistical Analysis
3. Results
3.1. Effects of Node Position and PGRs on In Vitro Regeneration
3.2. In Vitro Rooting and Acclimatization of M. limii Plantlets
3.3. Determination of Genetic Homogeneity Using ISSR
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bhattacharjee, A.; Chowdhery, H.J. Two frequently confused species of ‘Jewel Orchid’ (Orchidaceae—Goodyerinae) from India. Taiwania 2013, 58, 213–216. [Google Scholar] [CrossRef]
- Du, X.M.; Irino, N.; Furusho, N.; Hayashi, J.; Shoyama, Y. Pharmacologically active compounds in the Anoectochilus and Goodyera species. J. Nat. Med. 2008, 62, 132–148. [Google Scholar] [CrossRef]
- He, C.N.; Wang, C.L.; Guo, S.X.; Yang, J.S.; Xiao, P.G. A novel flavonoid glucoside from Anoectochilus roxburghii (Wall.) Lindl. J. Integr. Plant Biol. 2006, 48, 359–363. [Google Scholar] [CrossRef]
- Wood, J.J.; Lamb, A.; Lun, C.C.; Beaman, J.H.; Beaman, T.E. The Orchids of Mount Kinabalu; Natural History Publication: Kota Kinabalu, Malaysia, 2011; Volume 2, p. 1184. [Google Scholar]
- Li, B.; Tang, M.J.; Tang, K.; Zhao, L.F.; Guo, S.X. Screening for differentially expressed genes in Anoectochilus roxburghii (Orchidaceae) during symbiosis with the mycorrhizal fungus Epulorhiza sp. Sci. China Life Sci. 2012, 55, 164–171. [Google Scholar] [CrossRef] [Green Version]
- Tomita, M. Symbiotic of seed germination and development of Goodyera schlechtendaliana in vitro. Plant Tissue Cult. Lett. 1995, 12, 151–155. [Google Scholar] [CrossRef]
- Shiau, Y.J.; Nalawade, S.M.; Hsai, C.N.; Tsay, H.S. Propagation of Haemaria discolor via in vitro seed germination. Biol. Plant. 2005, 49, 341–346. [Google Scholar] [CrossRef]
- Shiau, Y.J.; Sagare, A.P.; Chen, U.C.; Yang, S.R.; Tsay, H.S. Conservation of Anoectochilus formosanus Hayata by artificial cross-pollination and in vitro culture of seeds. Bot. Bull. Acad. Sin. 2002, 43, 123–130. [Google Scholar]
- Chou, L.C.; Chang, D.C.N. Asymbiotic and symbiotic seed germination of Anoectochilus formosanus and Haemaria discolor and their F1 hybrids. Bot. Bull. Acad. Sin. 2004, 45, 143–147. [Google Scholar] [CrossRef]
- Indan, H.; David, D.; Jawan, R.; Marbawi, H.; Gansau, J.A. Development and characterization of flower and capsule in Borneo jewel orchid Macodes limii J.J. Wood and A.L. Lamb (Orchidaceae: Asparagales). J. Asia-Pacific Biodivers. 2021, 14, 236–241. [Google Scholar] [CrossRef]
- Chugh, S.; Guha, S.; Rao, I.U. Micropropagation of orchids: A review on the potential of different explants. Sci. Hortic. 2009, 122, 507–520. [Google Scholar] [CrossRef]
- Yam, T.W.; Arditti, J. Micropropagation of Orchids, 3rd ed.; John Wiley & Sons, Inc.: Chischester, UK, 2017; p. 2225. [Google Scholar] [CrossRef]
- Pant, B.; Pradhan, S.; Paudel, M.R.; Shah, S.; Pandey, S.; Joshi, P.R. Various culture techniques for the mass propagation of medicinal orchids from Nepal. Acta Hortic. 2019, 1262, 109–124. [Google Scholar] [CrossRef]
- Sherif, N.A.; Benjamin, J.H.F.; Muthukrishnan, S.; Kumar, T.S.; Rao, M.V. Regeneration of plantlets from nodal and shoot tip explants of Anoectochilus elatus Lindley, an endangered terrestrial orchid. Afr. J. Biotechnol. 2012, 11, 7549–7553. [Google Scholar] [CrossRef]
- Zhang, A.; Wang, H.; Shao, Q.; Xu, M.; Zhang, W.; Li, M. Large scale in vitro propagation of Anoectochilus roxburghii for commercial application: Pharmaceutically important and ornamental plant. Ind. Crops Prod. 2015, 70, 158–162. [Google Scholar] [CrossRef]
- Winarto, B.; Samijan. Axillary shoots derived from shoot tips in in vitro mass propagation of Anoectochilus formosanus Hayata. Agraarteadus 2018, 29, 121–130. [Google Scholar] [CrossRef]
- Poobathy, R.; Zakaria, R.; Murugaiyah, V.; Subramaniam, S. Surface sterilization and micropropagation of Ludisia discolor. Biocatal. Agric. Biotechnol. 2019, 22, 101380. [Google Scholar] [CrossRef]
- Mien, P.; Hang, P.; Ket, N.; Anh, T.; Phe, P.; Thanh, N.T. In vitro Culture of Jewel orchids (Anoectochilus setaceus Blume). Annu. Reports. 1998, 33, 436–444. [Google Scholar]
- Thanh, N.T.; Hang, P.L.; Van Ket, N.; Anh, T.T.; Van Phe, P.; Gam, N.T.; Mien, P.T. The role of different medium and plant hormones on multiple shoots of Jewel orchids (Anoectochilus setaceus Blume). VNU J. Sci. Nat. Sci. Technol. 2012, 28, 47–53. [Google Scholar]
- Sherif, N.A.; Kumar, T.S.; Rao, M.V. In vitro regeneration by callus culture of Anoectochilus elatus Lindley, an endangered terrestrial jewel orchid. In Vitro Cell. Dev. Biol.-Plant 2016, 52, 72–80. [Google Scholar] [CrossRef]
- Krishna, H.; Alizadeh, M.; Singh, D. Somaclonal variations and their applications in horticultural crops improvement. 3 Biotech 2016, 6, 54. [Google Scholar] [CrossRef] [Green Version]
- Sherif, N.A.; Kumar, T.S.; Rao, M.V. DNA barcoding and genetic fidelity assessment of micropropagated Aenhenrya rotundifolia (Blatt.) C.S. Kumar and F.N. Rasm.: A critically endangered jewel orchid. Physiol. Mol. Biol. Plants 2020, 26, 2391–2405. [Google Scholar] [CrossRef]
- Tikendra, L.; Koijam, A.S.; Nongdam, P. Molecular markers based genetic fidelity assessment of micropropagated Dendrobium chrysotoxum Lindl. Meta Gene 2019, 20, 100562. [Google Scholar] [CrossRef]
- Oliya, B.K.; Chand, K.; Thakuri, L.S.; Baniya, M.K.; Sah, A.K.; Pant, B. Assessment of genetic stability of micropropagated plants of Rhynchostylis retusa (L.) using RAPD markers. Sci. Hortic. 2020, 281, 110008. [Google Scholar] [CrossRef]
- Bhattacharyya, P.; Ghosh, S.; Mandi, S.S.; Kumaria, S.; Tandon, P. Genetic variability and association of AFLP markers with some important biochemical traits in Dendrobium thyrsiflorum, a threatened medicinal orchid. S. Afr. J. Bot. 2017, 109, 214–222. [Google Scholar] [CrossRef]
- Lin, S.F.; Tsay, H.S.; Chou, T.W.; Yang, M.J.; Cheng, K.T. Genetic variation of Anoectochilus formosanus revealed by ISSR and AFLP analysis. J. Food Drug Anal. 2007, 15, 156–162. [Google Scholar] [CrossRef]
- Zhang, F.; Lv, Y.; Dong, H.; Guo, S. Analysis of genetic stability through Intersimple sequence repeats molecular markers in micropropagated plantlets of Anoectochilus formosanus H AYATA, a medicinal plant. Biol. Pharm. Bull. 2010, 33, 384–388. [Google Scholar] [CrossRef] [Green Version]
- Sherif, N.A.; Kumar, T.S.; Rao, M.V. In vitro propagation and genetic stability assessment of an endangered terrestrial jewel orchid Anoectochilus elatus Lindl. Indian J. Exp. Biol. 2017, 55, 853–863. [Google Scholar]
- Sherif, N.A.; Benjamin, J.F.; Kumar, T.S.; Rao, M.V. Somatic embryogenesis, acclimatization and genetic homogeneity assessment of regenerated plantlets of Anoectochilus elatus Lindl., an endangered terrestrial jewel orchid. Plant Cell. Tissue Organ Cult. 2018, 132, 303–316. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Otiende, M.A.; Fricke, K.; Nyabundi, J.O.; Ngamau, K.; Hajirezaei, M.R.; Druege, U. Involvement of the auxin–cytokinin homeostasis in adventitious root formation of rose cuttings as affected by their nodal position in the stock plant. Planta 2021, 254, 65. [Google Scholar] [CrossRef]
- Miller, S.A.; Dykes, D.D.; Polesky, H.F. A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Res. 1988, 16, 1215. [Google Scholar] [CrossRef] [Green Version]
- Stephin, S.; Gangaprasad, A.; Mathew, S.P.; Muthukrishnan, S. Enhanced in vitro shoot multiplication of Piper sarmentosum by suppression of apical dominance. Proc. Natl. Acad. Sci. India Sect. B-Biol. Sci. 2020, 90, 87–94. [Google Scholar] [CrossRef]
- Chabikwa, T.G.; Brewer, P.B.; Beveridge, C.A. Initial bud outgrowth occurs independent of Auxin flow from out of buds. Plant Physiol. 2019, 179, 55–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cline, M.G. Concepts and terminology of apical dominance. Am. J. Bot. 1997, 84, 1064–1069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kebrom, T.H. A Growing Stem Inhibits Bud Outgrowth—The Overlooked Theory of Apical Dominance. Front. Plant. Sci. 2017, 8, 1874. [Google Scholar] [CrossRef]
- Muller, D.; Waldie, T.; Miyawaki, K.; To, J.P.; Melnyk, C.W.; Kieber, J.J.; Kakimoto, T.; Leyser, O. Cytokinin is required for escape but not release from auxin mediated apical dominance. Plant J. 2015, 82, 874–886. [Google Scholar] [CrossRef] [Green Version]
- Barbier, F.F.; Dun, E.A.; Kerr, S.C.; Chabikwa, T.G.; Beveridge, C.A. An Update on the Signals Controlling Shoot Branching. Trends Plant Sci. 2019, 24, 220–236. [Google Scholar] [CrossRef]
- Balla, J.; Medveďová, Z.; Kalousek, P.; Matiješčuková, N.; Friml, J. Auxin flow-mediated competition between axillary buds to restore apical dominance. Sci. Rep. 2016, 6, 35955. [Google Scholar] [CrossRef] [Green Version]
- Gangaprasad, A.; Latha, P.G.; Seeni, S. Micropropagation of terrestrial orchids, Anoectochilus sikkimensis and Anoectochilus regalis. Indian J. Exp. Biol. 2000, 38, 149–154. [Google Scholar]
- Vasudevan, R.; van Staden, J. Cytokinin and explant types influence in vitro plant regeneration of Leopard Orchid (Ansellia africana Lindl.). Plant Cell. Tissue Organ Cult. 2011, 107, 123–129. [Google Scholar] [CrossRef]
- Kieber, J.J.; Schaller, G.E. Cytokinin signaling in plant development. Development 2018, 145, dev149344. [Google Scholar] [CrossRef] [Green Version]
- Yuan, C.; Xi, L.; Kou, Y.; Zhao, Y.; Zhao, L. Current perspectives on shoot branching regulation. Front. Agric. Sci. Eng. 2015, 2, 38–52. [Google Scholar] [CrossRef] [Green Version]
- Deepa, A.V.; Anju, M.; Thomas, D.T. The Applications of TDZ in Medicinal Plant Tissue Culture. In Thidiazuron: From Urea Derivative to Plant Growth Regulator; Ahmad, N., Faisal, M., Eds.; Springer: Singapore, 2018; pp. 297–314. [Google Scholar] [CrossRef]
- Li, Y.; Hu, J.; Xiao, J.; Guo, G.; Jeong, B.R. Foliar thidiazuron promotes the growth of axillary buds in Strawberry. Agronomy 2021, 11, 594. [Google Scholar] [CrossRef]
- Ket, N.V.; Hahn, E.J.; Park, S.Y.; Chakrabarty, D.; Paek, K.Y. Micropropagation of an endangered orchid Anoectochilus formosanus. Biol. Plant. 2004, 48, 339–344. [Google Scholar] [CrossRef]
- Raja, D.H.D. Effect of cytokinins on micropropagation of Anoectochilus elatus Lindl. from shoot tip explants—An endangered medicinal orchid. IOSR J. Biotechnol. Biochem. 2017, 3, 73–76. [Google Scholar] [CrossRef]
- Balilashaki, K.; Vahedi, M.; Karimi, R. In vitro direct regeneration from node and leaf explants of Phalaenopsis cv. ‘Surabaya’. Plant Tissue Cult. Biotechnol. 2015, 25, 193–205. [Google Scholar] [CrossRef] [Green Version]
- Jainol, J.E.; Gansau, J.A. Embryogenic callus induction from leaf tip explants and protocorm-like body formation and shoot proliferation of Dimorphorchis lowii: Borneon endemic orchid. Agrivita 2017, 39, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Hazarika, B.N. Acclimatization of tissue-cultured plants. Curr. Sci. 2003, 85, 1704–1712. [Google Scholar]
- Pospóšilová, J.; Tichá, I.; Kadleček, P.; Haisel, D.; Plzáková, Š. Acclimatization of micropropagated plants to ex-vitro conditions. Biol. Plant. 1999, 42, 481–497. [Google Scholar] [CrossRef]
- Konar, S.; Karmakar, J.; Ray, A.; Adhikari, S.; Bandyopadhyay, T.K. Regeneration of plantlets through somatic embryogenesis from root derived calli of Hibiscus sabdariffa L. (Roselle) and assessment of genetic stability by flow cytometry and ISSR analysis. PLoS ONE 2018, 13, e0202324. [Google Scholar] [CrossRef]
- Chin, C.K.; Lee, Z.H.; Mubbarakh, S.A.; James, A.J.J.; Chew, B.L.; Subramaniam, S. Effects of plant growth regulators and activated charcoal on somaclonal variations of protocorm-like bodies (PLBs) of Dendrobium Sabin Blue orchid. Biocatal. Agric. Biotechnol. 2019, 22, 101426. [Google Scholar] [CrossRef]
- Kang, L.; Zheng, K.; Xie, Y.; Deng, Y.; Yu, Y.; Zhu, M.; Xi, R.; Deng, X. Efficient tissue culture protocol for Magnolia lucida (Magnoliaceae) and confirmation of genetic stability of the regenerated plants. Plants 2020, 9, 997. [Google Scholar] [CrossRef] [PubMed]
- Dewir, Y.H.; Nurmansyah; Naidoo, Y.; Teixeira da Silva, J.A. Thidiazuron-induced abnormalities in plant tissue cultures. Plant Cell Rep. 2018, 37, 1451–1470. [Google Scholar] [CrossRef] [PubMed]
- Samarfard, S.; Kadir, M.A.; Kadzimin, S.B.; Saud, H.M.; Ravanfar, S.A.; Danaee, M. In vitro propagation and detection of somaclonal variation in Phalaenopsis gigantea as affected by chitosan and thidiazuron combinations. HortScience 2014, 49, 82–88. [Google Scholar] [CrossRef]
- Roy, A.R.; Sajeev, S.; Pattanayak, A.; Deka, B.C. TDZ induced micropropagation in Cymbidium giganteum Wall. Ex Lindl. and assessment of genetic variation in the regenerated plants. Plant Growth Regul. 2012, 68, 435–445. [Google Scholar] [CrossRef]
- Kumari, S.; Yadav, K.; Singh, N. Evaluation of genetic fidelity among micropropagated plants of Salvadora persica L. using DNA-based markers. Meta Gene 2017, 14, 129–133. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| PGRs | Node Position | % Shoot Formation | No. of Shoots | Shoot Length (cm) |
|---|---|---|---|---|
| Control | N1 | 35 | 0.7 ± 0.5 defgh | 1.6 ± 1.1 ab |
| N2 | 75 | 1.4 ± 0.5 bcdefgh | 1.6 ± 0.5 ab | |
| N3 | 85 | 1.9 ± 0.7 abcdefg | 1.4 ± 0.5 abc | |
| N4 | 60 | 2.1 ± 1.5 abcde | 1.2 ± 0.8 abcde | |
| 1.0 mg/L NAA | N1 | 0 | 0.0 h | 0.00 g |
| N2 | 35 | 0.9 ± 1.3 cdefgh | 0.2 ± 0.3 fg | |
| N3 | 20 | 0.4 ± 0.5 fgh | 0.3 ± 0.4 efg | |
| N4 | 15 | 0.3 ± 0.5 gh | 0.2 ± 0.3 fg | |
| 1.0 mg/L Picloram | N1 | 40 | 0.8 ± 0.4 defgh | 1.3 ± 0.8 abcd |
| N2 | 60 | 1.2 ± 0.8 cdefgh | 0.8 ± 0.4 bcdefg | |
| N3 | 60 | 1.2 ± 0.8 cdefgh | 0.8 ± 0.4 bcdefg | |
| N4 | 50 | 1.0 ± 0.9 cdefgh | 0.7 ± 0.6 bcdefg | |
| 1.0 mg/L 2,4-D | N1 | 20 | 0.4 ± 0.5 fgh | 0.6 ± 0.8 cdefg |
| N2 | 60 | 1.2 ± 1.0 cdefgh | 0.9 ± 0.8 abcdefg | |
| N3 | 80 | 2.2 ± 1.2 abcde | 1.2 ± 0.5 abcde | |
| N4 | 30 | 1.0 ± 1.6 cdefgh | 0.4 ± 0.5 defg | |
| 1.0 mg/L BAP | N1 | 40 | 0.8 ± 0.4 defgh | 0.8 ± 0.7 bcdefg |
| N2 | 80 | 1.8 ± 0.8 abcdefg | 0.7 ± 0.3 bcdefg | |
| N3 | 80 | 2.0 ± 1.1 abcdef | 0.9 ± 0.7 abcdefg | |
| N4 | 70 | 2.2 ± 1.4 abcde | 0.5 ± 0.3 cdefg | |
| 1.0 mg/L Kinetin | N1 | 30 | 0.6 ± 0.5 efgh | 0.7 ± 0.7 bcdefg |
| N2 | 80 | 1.8 ± 0.8 abcdefg | 0.7 ± 0.4 bcdefg | |
| N3 | 80 | 2.5 ± 1.4 abc | 0.6 ± 0.4 cdefg | |
| N4 | 80 | 2.2 ± 1.2 abcde | 0.7 ± 0.4 bcdefg | |
| 1.0 mg/L TDZ | N1 | 40 | 0.8 ± 0.4 defgh | 1.0 ± 0.7 abcdef |
| N2 | 85 | 2.3 ± 1.1 abcd | 1.4 ± 0.2 abc | |
| N3 | 95 | 3.1 ± 1.0 a | 1.6 ± 0.5 ab | |
| N4 | 95 | 3.0 ± 1.1 ab | 1.8 ± 0.4 a |
| Source of Variation | SS | df | Mean Square | F | p Value |
|---|---|---|---|---|---|
| No. of shoots | |||||
| Node position (N) | 70.414 | 3 | 23.471 | 25.539 | <0.001 ** |
| PGRs (P) | 88.621 | 6 | 14.770 | 16.071 | <0.001 ** |
| N × P | 29.636 | 18 | 1.646 | 1.791 | 0.027 * |
| Shoot length | |||||
| Node position (N) | 1.270 | 3 | 0.423 | 1.362 | 0.255 NS |
| PGRs (P) | 49.240 | 6 | 8.207 | 26.397 | <0.001 ** |
| N × P | 9.757 | 18 | 0.542 | 1.744 | 0.033 * |
| Primer Code | Primer Sequence (5′-3′) | Nucleotide Fragment Size (bp) | Total Bands Amplified | Monomorphic Band | Polymorphic Band | Monomorphism (%) | Polymorphism (%) |
|---|---|---|---|---|---|---|---|
| UBC808 | AGAGAGAGAGAGAGAGC | 350–950 | 70 | 70 | 0 | 100 | 0 |
| UBC835 | AGAGAGAGAGAGAGAGYC | 325–1200 | 69 | 68 | 1 | 98.6 | 1.5 |
| UBC836 | AGAGAGAGAGAGAGAGYC | 600–1000 | 16 | 16 | 0 | 100 | 0 |
| UBC841 | GAGAGAGAGAGAGAGAYC | 320–900 | 51 | 49 | 2 | 96.1 | 3.9 |
| UBC842 | GAGAGAGAGAGAGAGAYG | 310–800 | 54 | 52 | 2 | 96.3 | 3.7 |
| Total | 260 | 255 | 5 | 98.1 | 1.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
David, D.; Rusdi, N.A.; Mohd Mokhtar, R.A.; Mohd Faik, A.A.; Gansau, J.A. Establishment of In Vitro Regeneration Protocol for Sabah’s Jewel Orchid, Macodes limii J.J. Wood & A.L. Lamb. Horticulturae 2022, 8, 155. https://doi.org/10.3390/horticulturae8020155
David D, Rusdi NA, Mohd Mokhtar RA, Mohd Faik AA, Gansau JA. Establishment of In Vitro Regeneration Protocol for Sabah’s Jewel Orchid, Macodes limii J.J. Wood & A.L. Lamb. Horticulturae. 2022; 8(2):155. https://doi.org/10.3390/horticulturae8020155
Chicago/Turabian StyleDavid, Devina, Nor Azizun Rusdi, Ruzaidi Azli Mohd Mokhtar, Ainol Azifa Mohd Faik, and Jualang Azlan Gansau. 2022. "Establishment of In Vitro Regeneration Protocol for Sabah’s Jewel Orchid, Macodes limii J.J. Wood & A.L. Lamb" Horticulturae 8, no. 2: 155. https://doi.org/10.3390/horticulturae8020155