
Impact of Plant Growth-Promoting Rhizobacteria Inoculation on the Physiological Response and Productivity Traits of Field-Grown Tomatoes in Hungary

 ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Research Materials and Planning
2.2. Rhizobacteria Inoculation
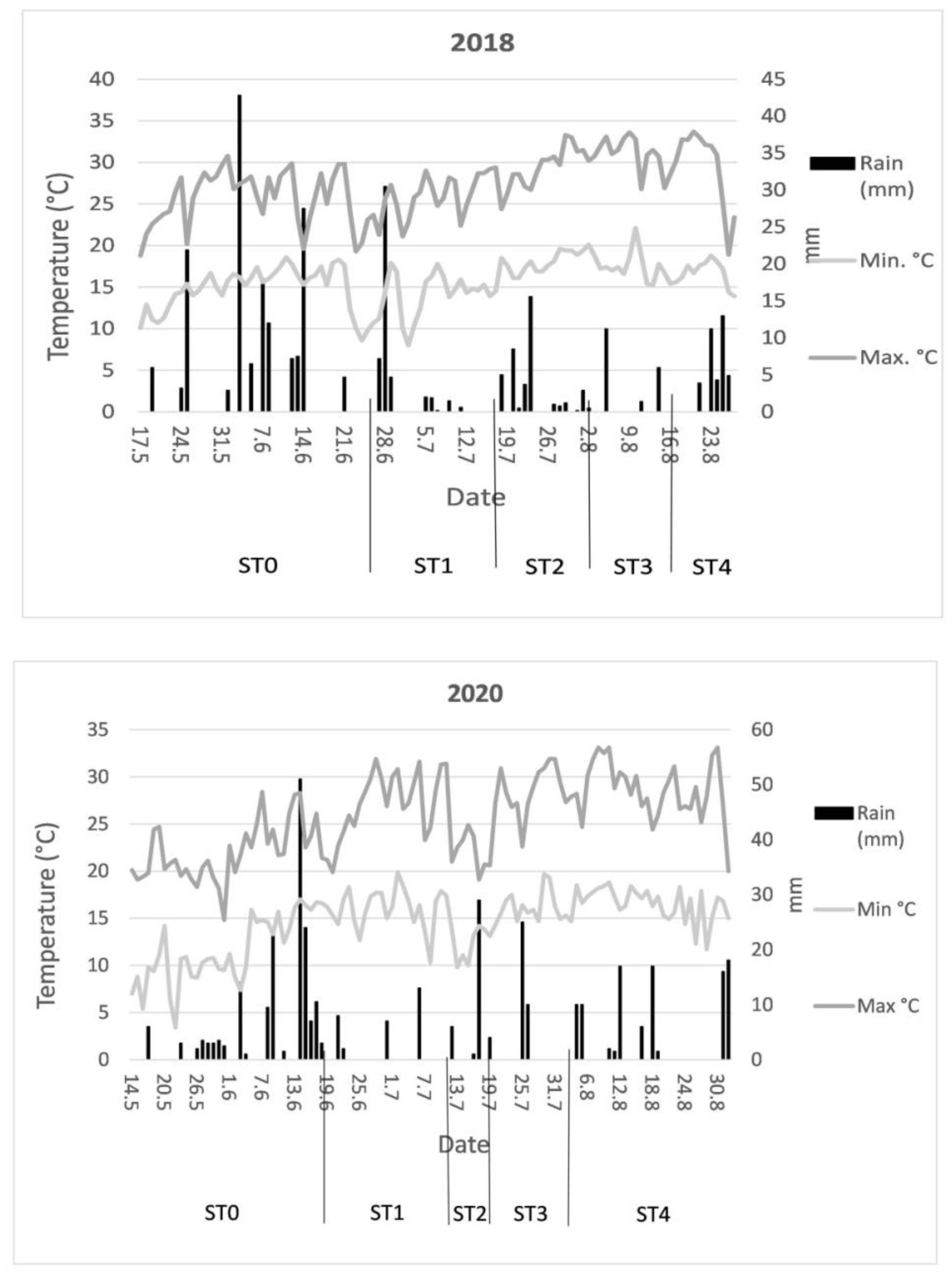
2.3. Climatic Data and Irrigation Treatments during Tomato Development
2.4. Field Measurements
2.5. Yield Analysis
2.6. Statistical Analysis
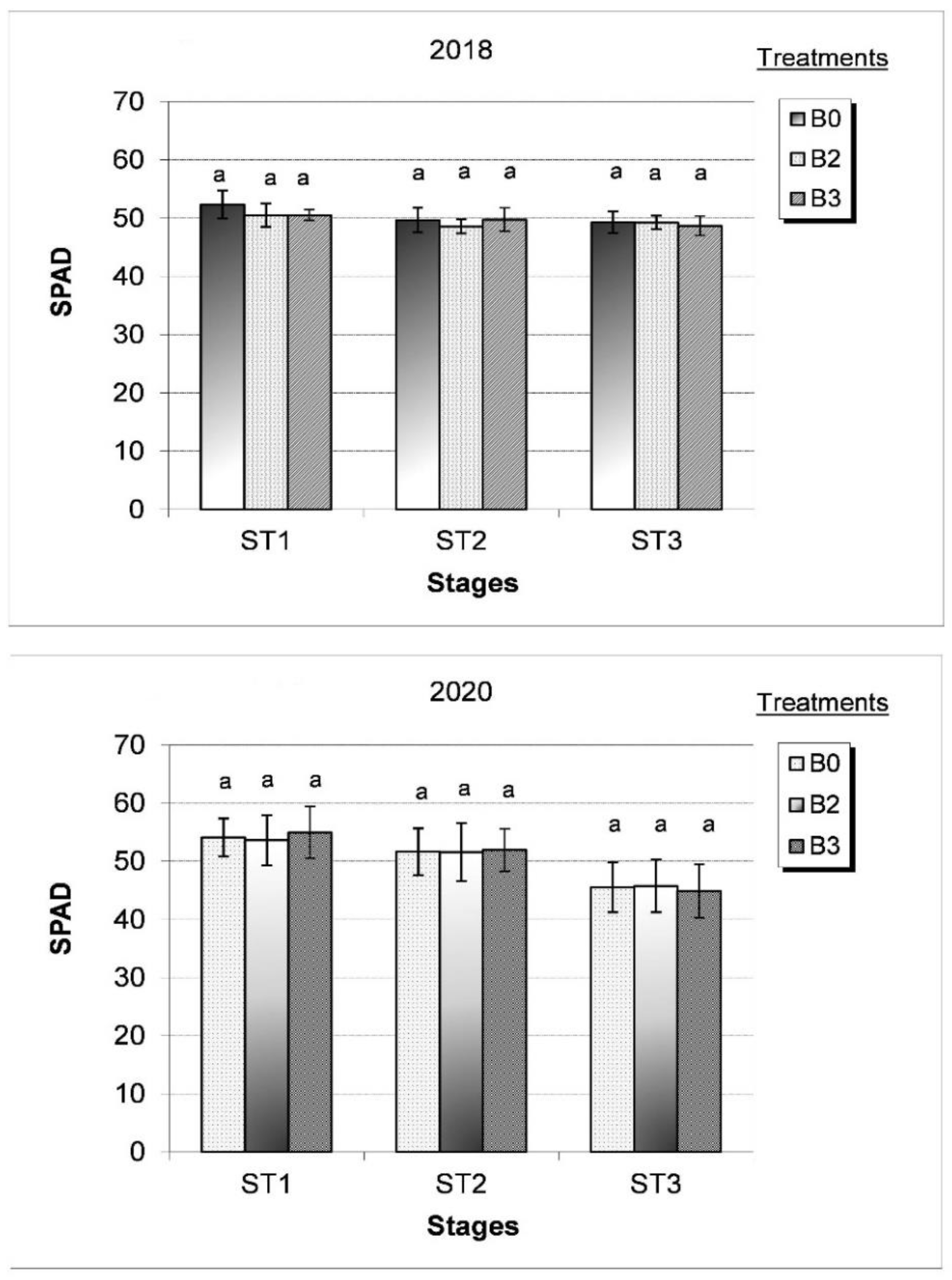
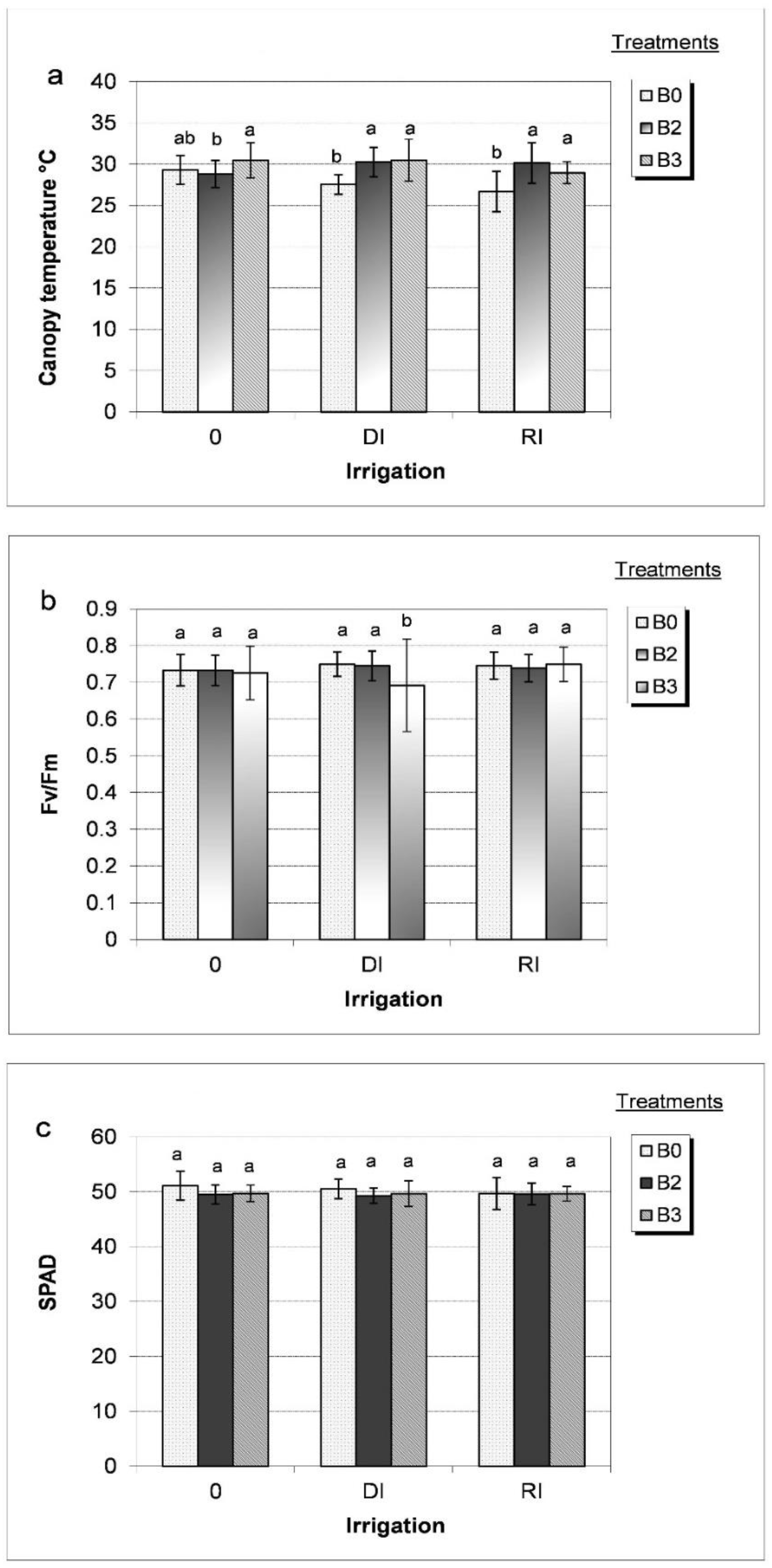
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Agbna, G.H.D.; Donglia, S.; Zhipeng, L.; Elshaikha, N.A.; Guangchenga, S.; Timm, L.C. Effects of deficit irrigation and biochar addition on the growth, yield, and quality of tomato. Sci. Hortic. 2017, 222, 90–101. [Google Scholar] [CrossRef]
- Du, Y.D.; Cao, H.-X.; Liu, S.-Q.; Gu, X.-B.; Cao, Y.-X. Response of yield, quality, water and nitrogen use efficiency of tomato to different levels of water and nitrogen under drip irrigation in Northwestern China. J. Integr. Agric. 2017, 16, 1153–1161. [Google Scholar] [CrossRef] [Green Version]
- Lorenzi, F.; Alfieri, S.M.; Monaco, E.; Bonfante, A.; Basile, A.; Patanè, C.; Menenti, M. Adaptability to future climate of irrigated crops: The interplay of water management and cultivars responses. A case study on tomato. Biosyst. Eng. 2017, 157, 45–62. [Google Scholar] [CrossRef]
- Takács, S.; Csengeri, E.; Pék, Z.; Bíró, T.; Szuvandzisev, P.; Palotás, G.; Helyes, L. Performance evaluation of AquaCrop model in processing tomato biomass, fruit yield and water stress indicator modelling. Water 2021, 13, 3587. [Google Scholar] [CrossRef]
- Sing, S.K.; Reddy, K.R. Regulation of photosynthesis, fluorescence, stomatal conductance and water-use efficiency of cowpea (Vigna unguiculata [L.] Walp.) under drought. J. Photoch. Photobiol. B Biol. 2011, 105, 40–50. [Google Scholar] [CrossRef]
- Nemeskéri, E.; Molnár, K.; Pék, Z.; Helyes, L. Effect of water supply on water use related physiological traits and yield of snap beans in dry seasons. Irrig. Sci. 2018, 36, 143–158. [Google Scholar] [CrossRef]
- Vadez, V. Root hydraulics: The forgotten side of roots in drought adaptation. Field Crops Res. 2014, 165, 15–24. [Google Scholar] [CrossRef] [Green Version]
- Ratnakumar, P.; Vadez, V. Tolerant groundnut (A. hypogaea L.) genotypes to intermittent drought maintains high harvest index and has small leaf canopy under stress. Funct. Plant Biol. 2011, 38, 1016–1023. [Google Scholar] [CrossRef] [Green Version]
- Pasala, R.; ·Pandey, B.B.; Gandi, S.L.; Kulasekaran, R.; Guhey, A.; Reddy, A.V. An insight into the mechanisms of intermittent drought adaptation in sesame (Sesamum indicum L.): Linking transpiration efficiency and root architecture to seed yield. Acta Physiol. Plant 2021, 43, 148. [Google Scholar] [CrossRef]
- Helyes, L.; Bőcs, A.; Pék, Z. Effect of water supply on canopy temperature, stomatal conductance and yield quantity of processing tomato (Lycopersicon esculentum Mill.). Int. J. Hortic. Sci. 2010, 16, 13–15. [Google Scholar] [CrossRef]
- Nemeskéri, E.; Neményi, A.; Bőcs, A.; Pék, Z.; Helyes, L. Physiological factors and their relationship with the productivity of processing tomato under different water supplies. Water 2019, 11, 586. [Google Scholar] [CrossRef] [Green Version]
- Lobos, G.; Retamales, J.B.; Hancock, J.F.; Flore, J.A.; Cobo, N.; del Pozo, A. Spectral irradiance, gas exchange characteristics and leaf traits of Vaccinium corymbosum L. ‘Elliott’ grown under photo-selective nets. Environ. Exp. Bot. 2012, 75, 142–149. [Google Scholar] [CrossRef]
- Ogweno, J.O.; Song, X.S.; Hu, W.H.; Shi, K.; Zhou, Y.H.; Yu, J.Q. Detached leaves of tomato differ in their photosynthetic physiological response to moderate high and low temperature stress. Sci. Hortic. 2009, 123, 17–22. [Google Scholar] [CrossRef]
- Mishra, K.B.; Iannacone, R.; Petrozza, A.; Mishra, A.; Armentano, N.; La Vecchia, G.; Trtilek, M.; Cellini, F.; Nedbal, L. Engineered drought tolerance in tomato plants is reflected in chlorophyll fluorescence emission. Plant Sci. 2012, 182, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Huang, G.; Jia, D.; Wang, J.; Mota, M.; Pereira, L.S.; Huang, Q.; Xu, X.; Liu, H. Responses of drip irrigated tomato (Solanum lycopersicum L.) yield, quality and water productivity to various soil matric potential thresholds in an arid region of Northwest China. Agric. Water Manag. 2013, 129, 181–193. [Google Scholar] [CrossRef]
- Bahadur, A.; Chatterjee, A.; Kumar, R.; Singh, M.; Naik, P.S. Physiological and biochemical basis of drought tolerance in vegetables. Veg. Sci. 2011, 38, 1–16. [Google Scholar]
- Pék, Z.; Szuvandzsiev, P.; Neményi, A.; Helyes, L. Effect of season and irrigation on yield parameters and soluble solids content of processing cherry tomato. Acta Hortic. 2015, 1081, 197–202. [Google Scholar] [CrossRef]
- Shao, H.B.; Chu, L.Y.; Jaleel, A.; Zhao, C.X. Water-deficit stress-induced anatomical changes in higher plants. CR Biol. 2008, 331, 215–225. [Google Scholar] [CrossRef]
- Zhang, X.; Lu, G.; Long, W.; Zou, X.; Li, F.; Nishio, T. Recent progress in drought and salt tolerance studies in Brassica crops. Breed. Sci. 2014, 64, 60–73. [Google Scholar] [CrossRef] [Green Version]
- Barriuso, J.; Solano, B.R.; Mañero, F.J.G. Protection against pathogen and salt stress by four plant growth-promoting rhizobacteria isolated from Pinus sp. on Arabidopsis thaliana. Phytopathology 2008, 98, 666–672. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Yang, X.G.; Batchelor, W.D.; Liu, Z.J.; Zhang, Z.T.; Wan, N.H.; Sun, S.; He, B.; Gao, J.Q.; Bai, F.; et al. A case study of climate-smart management in foxtail millet (Setaria italica) production under future climate change in Lishu county of Jilin, China. Agric. For. Meteorol. 2020, 292–293, 108131. [Google Scholar] [CrossRef]
- Sauter, A.; Dietz, K.J.; Hartung, W. A possible stress physiological role of abscisic acid conjugates in root-to-shoot signalling. Plant Cell Environ. 2002, 25, 223–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, A.C.; Travaglia, C.N.; Bottini, R.; Piccoli, P.N. Participation of abscisic acid and gibberellins produced by endophytic Azospirillum in the alleviation of drought effects in maize. Botany 2009, 87, 455–462. [Google Scholar] [CrossRef]
- Sharifi, R.; Ryu, C.M. Chatting with a Tiny Belowground Member of Holobiome: Communication Between Plants and Gowth-Promoting Rhizobacteria. Adv. Bot. Res. 2017, 82, 135–160. [Google Scholar]
- Kamilova, F.; Kravchenko, L.V.; Shaposhnikov, A.I.; Azarova, T.; Makarova, N.; Lugtenberg, B. Organic acids, sugars and L-tryptophane in exudates of vegetables growing on stonewool and their effects on activities of rhizosphere bacteria. Mol. Plant Microbe Interact. 2006, 9, 250–256. [Google Scholar] [CrossRef] [Green Version]
- Oku, S.; Komatsu, A.; Tajima, T.; Nakashimada, Y.; Kato, J. Identification of chemotaxis sensory proteins for amino acids in Pseudomonas fluorescens Pf0-1 and their involvement in chemotaxis to tomato root exudate and root colonization. Microbes Environ. 2012, 27, 462–469. [Google Scholar] [CrossRef] [Green Version]
- Dodd, I.C.; Zinovkina, N.Y.; Safronova, V.I.; Belimov, A.A. Rhizobacterial mediation of plant hormone status. Ann. Appl. Biol. 2010, 157, 361–379. [Google Scholar] [CrossRef]
- Zhang, Y.; Cui, G.; Zhang, W.; Lang, D.; Zhixian, L.; Zhang, X. Bacillus sp. G2 improved the growth of Glycyrrhiza uralensis Fisch. related to antioxidant metabolism and osmotic adjustment. Acta Physiol. Plant 2021, 43, 152. [Google Scholar] [CrossRef]
- Nadeem, S.M.; Ahmad, M.; Zahir, Z.A.; Javaid, A.; Ashraf, M. The role of mycorrhizae and plant growth promoting rhizobacteria (PGPR) in improving crop productivity under stressful environments. Biotechnol. Adv. 2014, 32, 429–448. [Google Scholar] [CrossRef]
- Singh, V.K.; Singh, A.K.; Singh, P.P.; Kumar, A. Interaction of plant growth promoting bacteria with tomato under abiotic stress: A review. Agric. Ecosyst. Environ. 2018, 267, 129–140. [Google Scholar] [CrossRef]
- Berg, G. Plant-microbe interactions promoting plant growth and health: Perspectives for controlled use of microorganisms in agriculture. Appl. Microbiol. Biotechnol. 2009, 84, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Khanghahi, M.Y.; Leoni, B.; Crecchio, C. Photosynthetic responses of durum wheat to chemical/microbiological fertilization management under salt and drought stresses. Acta Physiol. Plant 2021, 43, 123. [Google Scholar] [CrossRef]
- Le, A.T.; Pék, Z.; Takács, S.; Neményi, A.; Helyes, L. The effect of plant growth-promoting rhizobacteria on yield, water use efficiency and Brix Degree of processing tomato. Plant Soil Environ. 2018, 64, 523–529. [Google Scholar] [CrossRef] [Green Version]
- Andryei, B.; Horváth, K.Z.; Agyemang Duah, S.; Takács, S.; Égei, M.; Szuvandzsiev, P.; Neményi, A. Use of plant growth promoting rhizobacteria (PGPRs) in the mitigation of water deficiency of tomato plants (Solanum lycopersicum L.). J. Cent. Eur. Agric. 2021, 22, 167–177. [Google Scholar]
- Zhou, Y.; Sang, T.; Tian, M.; Jahan, M.S.; Wang, J.; Li, X.; Guo, S.; Liu, H.; Wang, Y.; Shu, S. Effects of Bacillus cereus on photosynthesis and antioxidant metabolism of cucumber seedlings under salt stress. Horticulturae 2022, 8, 463. [Google Scholar] [CrossRef]
- Papoui, E.; Bantis, F.; Kapoulas, N.; Ipsilantis, I.; Koukounaras, A. A sustainable intercropping system for organically produced lettuce and green onion with the use of arbuscular mycorrhizal inocula. Horticulturae 2022, 8, 466. [Google Scholar] [CrossRef]
- Sneha, S.; Anitha, B.; Sahair, R.A.; Raghu, N.; Gopenath, T.S.; Chandrashekrappa, G.K.; Basalingappa, K.M. Biofertilizer for crop production and soil fertility. Acad. J. Agric. Res. 2018, 6, 299–306. [Google Scholar]
- Kaur, R.; Kaur, S.; Kaur, G. Molecular and physiological manipulations in rhizospheric bacteria. Acta Physiol. Plant 2021, 43, 77. [Google Scholar] [CrossRef]
- Vejan, P.; Abdullah, R.; Khadiran, T.; Ismail, S.; Boyce, A.N. Role of plant growth promoting rhizobacteria in agricultural sustainability: A review. Molecules 2016, 21, 573. [Google Scholar] [CrossRef]
- Aslam, M.M.; Okal, E.J.; Idris, A.L.; Qian, Z.; Xu, W.; Karanja, J.K.; Wani, S.H.; Yuan, W. Rhizosphere microbiomes can regulate plant drought tolerance. Pedosphere 2022, 32, 61–74. [Google Scholar] [CrossRef]
- Yang, J.; Kloepper, J.W.; Ryu, C.M. Rhizosphere bacteria help plants tolerate abiotic stress. Trends Plant Sci. 2009, 14, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Forni, C.; Duca, D.; Glick, B.R. Mechanisms of plant response to salt and drought stress and their alteration by rhizobacteria. Plant Soil 2017, 410, 335–356. [Google Scholar] [CrossRef]
- Davies, W.J.; Zhang, J. Root signals and the regulation of growth and development of plants in drying soil. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1991, 42, 55–76. [Google Scholar] [CrossRef]
- Bresson, J.; Varoquaux, F.; Bontpart, T.; Touraine, B.; Vile, D. The PGPR strain Phyllobacterium brassicacearum STM196 induces a reproductive delay and physiological changes that result in improved drought tolerance in Arabidopsis. N. Phytol. 2013, 200, 558–569. [Google Scholar] [CrossRef]
- Nemeskéri, E.; Horváth, K.; Pék, Z.; Helyes, L. Effect of mycorrhizal and bacterial products on the traits related to photosynthesis and fruit quality of tomato under water deficiency conditions. Acta Hortic. 2019, 1233, 61–66. [Google Scholar] [CrossRef]
- Horvath, K.Z.; Andryei, B.; Helyes, L.; Pék, Z.; Neményi, A.; Nemeskéri, E. Effect of mycorrhizal inoculations on physiological traits and bioactive compounds of tomato under water scarcity in field conditions. Not. Bot. Hortic. Agrobot Cluj Napoca 2020, 48, 1233–1247. [Google Scholar] [CrossRef]
- Costa-Santos, M.; Mariz-Ponte, N.; Dias, M.C.; Moura, L.; Marques, G.; Santos, C. Effect of Bacillus spp. and Brevibacillus sp. on the Photosynthesis and Redox Status of Solanum lycopersicum. Horticulturae 2021, 7, 24. [Google Scholar] [CrossRef]
- Tahir, M.; Khalid, U.; Khan, M.B.; Shahid, M.; Ahmad, I.; Akram, M.; Ijaz, M.; Hussain, M.; Farooq, A.B.; Naeem, M.A.; et al. Auxin and 1-aminocyclopropane-1-carboxylate deaminase activity exhibiting rhizobacteria enhanced maize quality and productivity under water deficit conditions. Int. J. Agric. Biol. 2019, 21, 943–954. [Google Scholar]
- Gagné-Bourque, F.; Bertrand, A.; Claessens, A.; Aliferis, K.A.; Jabaji, S. Alleviation of drought stress and metabolic changes in timothy (Phleum pratense L.) colonized with Bacillus subtilis B26. Front. Plant Sci. 2016, 7, 584. [Google Scholar] [CrossRef] [Green Version]
- Zhou, C.; Ma, Z.; Zhu, L.; Xiao, X.; Xie, Y.; Zhu, J.; Wang, J. Rhizobacterial strain Bacillus megaterium BOFC15 induces cellular polyamine changes that improve plant growth and drought resistance. Int. J. Mol. Sci. 2016, 17, 976. [Google Scholar] [CrossRef]
- Chen, L.; Liu, Y.; Wu, G.; Veronican, N.; Jeri, K.; Shen, Q.; Zhang, N.; Zhang, R. Induced maize salt tolerance by rhizosphere inoculation of Bacillus amyloliquefaciens SQR9. Physiol. Plant 2016, 158, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Kloepper, J.W.; Lifshitz, R.; Zablotowich, R.K. Free-living bacterial inocula for enhancing crop productivity. Trends Biotechnol. 1989, 7, 39–43. [Google Scholar] [CrossRef]
- Bona, E.; Cantamessa, S.; Massa, N.; Manassero, P.; Marsano, F.; Copetta, A.; Lingua, G.; D’Agostino, G.; Gamalero, E.; Berta, G. Arbuscular mycorrhizal fungi and plant growth-promoting pseudomonads improve yield, quality and nutritional value of tomato: A field study. Mycorrhiza 2017, 27, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Mondani, F.; Khani, K.; Honarmand, S.L.; Saeidi, M. Evaluating effects of plant growth-promoting rhizobacteria on the radiation use efficiency and yield of soybean (Glycine max) under water deficit stress condition. Agric. Water Manag. 2019, 213, 707–713. [Google Scholar] [CrossRef]
- Vaikuntapu, P.R.; Dutta, S.; Samudrala, R.B.; Rao, V.R.V.N.; Kalam, S.; Podile, A.R. Preferential promotion of Lycopersicon esculentum (tomato) growth by plant growth promoting bacteria associated with tomato. Indian J. Microbiol. 2014, 54, 403–412. [Google Scholar] [CrossRef] [Green Version]
- Zhou, D.; Huang, X.F.; Chaparro, J.M.; Badri, D.V.; Manter, D.K.; Vivanco, J.M.; Guo, J. Root and bacterial secretions regulate the interaction between plants and PGPR leading to distinct plant growth promotion effects. Plant Soil 2016, 401, 259–272. [Google Scholar] [CrossRef]
- Kalam, S.; Basu, A.; Podile, A.R. Functional and molecular characterization of plant growth promoting Bacillus isolates from tomato rhizosphere. Heliyon 2020, 6, e04734. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Tmax Average °C | Tmin Average °C | Precipitation (mm) | Irrigation (mm) | Total Water (mm) | |||
|---|---|---|---|---|---|---|---|---|
| DI | RI | I0 | DI | RI | ||||
| 2018 | 27.5 | 15.7 | 304.6 | 80.2 | 160.3 | 349.9 | 384.8 | 464.9 |
| 2020 | 25.7 | 14.5 | 375.1 | 54.8 | 102.7 | 380.1 | 429.9 | 477.8 |
| 2018 | 2020 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Water Supply (WS) a | PGPR b | TY tha−1 | MY ha−1 | GY tha−1 | DY tha−1 | GBm tha−1 | Soluble Solids (°Brix) | TY tha−1 | MY ha−1 | GY tha−1 | DY tha−1 | GBm tha−1 | Soluble Solids (°Brix) |
| I0 | B0 | 50.71 b | 45.45 b | 4.40 c | 0.86 a | 66.3 b | 4.25 a | 49.78 b | 40.13 a | 2.52 a | 7.08 c | 60.3 b | 4.64 a |
| B2 | 68.6 a | 57.46 a | 11.05 a | 1.70 a | 84.7 a | 4.36 a | 56.78 a | 39.92 a | 4.43 a | 12.40 a | 70.0 a | 4.55 a | |
| B3 | 68.83 a | 58.27 a | 8.32 b | 2.25 a | 88.6 a | 4.39 a | 51.48 a | 38.71 a | 3.51 a | 9.28 b | 61.4 b | 4.20 b | |
| I0 | 62.72 B | 53.72 B | 7.92 A | 1.60 A | 79.87 B | 4.33 A | 52.68 A | 39.59 A | 3.49 A | 9.59 B | 63.9 A | 4.46 A | |
| DI | B0 | 64.97 b | 59.26 c | 4.65 d | 1.06 a | 81.1 b | 3.65 a | 51.10 b | 40.33 a | 2.05 a | 8.71 b | 62.7 ab | 4.00 a |
| B2 | 83.54 a | 65.30 b | 15.36 a | 2.89 a | 100.4 a | 3.61 a | 55.75 a | 39.11 a | 2.57 a | 14.03 a | 66.9 a | 3.80 a | |
| B3 | 93.28 a | 82.56 a | 10.28 b | 2.36 a | 116.3 a | 3.69 a | 47.05 c | 31.71 b | 1.70 a | 13.62 a | 56.2 b | 3.88 a | |
| DI | 80.57 A | 69.04 A | 10.10 A | 2.10 A | 99.3 A | 3.65 B | 51.30 A | 37.05 A | 2.11 B | 12.12 A | 61.9 A | 3.89 B | |
| RI | B0 | 72.43 b | 59.75 b | 9.92 b | 2.71 a | 87.5 b | 3.38 a | 48.40 b | 34.51 a | 1.71 a | 12.21 b | 64.5 a | 3.40 a |
| B2 | 76.37 b | 62.99 a | 10.41 b | 14.44 a | 89.8 b | 3.34 a | 56.18 a | 35.71 a | 2.23 a | 18.22 a | 67.1 a | 3.42 a | |
| B3 | 85.01 a | 69.72 a | 13.16 a | 2.14 a | 98.1 b | 3.22 a | 43.58 c | 29.36 b | 2.83 a | 11.34 b | 54.7 b | 3.04 b | |
| RI | 77.94 A | 64.15 A | 11.16 A | 2.61 A | 91.8 A | 3.31 C | 49.38 A | 33.19 B | 2.26 B | 13.93 A | 62.1 A | 3.29 C | |
| Significance | WS | * | * | ns | ns | * | *** | ns | * | * | ** | ns | *** |
| PGPR | * | * | * | ns | * | ns | *** | * | Ns | *** | ** | *** | |
| WS × PGPR | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nemeskéri, E.; Horváth, K.Z.; Andryei, B.; Ilahy, R.; Takács, S.; Neményi, A.; Pék, Z.; Helyes, L. Impact of Plant Growth-Promoting Rhizobacteria Inoculation on the Physiological Response and Productivity Traits of Field-Grown Tomatoes in Hungary. Horticulturae 2022, 8, 641. https://doi.org/10.3390/horticulturae8070641
Nemeskéri E, Horváth KZ, Andryei B, Ilahy R, Takács S, Neményi A, Pék Z, Helyes L. Impact of Plant Growth-Promoting Rhizobacteria Inoculation on the Physiological Response and Productivity Traits of Field-Grown Tomatoes in Hungary. Horticulturae. 2022; 8(7):641. https://doi.org/10.3390/horticulturae8070641
Chicago/Turabian StyleNemeskéri, Eszter, Kitti Zsuzsanna Horváth, Bulgan Andryei, Riadh Ilahy, Sándor Takács, András Neményi, Zoltán Pék, and Lajos Helyes. 2022. "Impact of Plant Growth-Promoting Rhizobacteria Inoculation on the Physiological Response and Productivity Traits of Field-Grown Tomatoes in Hungary" Horticulturae 8, no. 7: 641. https://doi.org/10.3390/horticulturae8070641
APA StyleNemeskéri, E., Horváth, K. Z., Andryei, B., Ilahy, R., Takács, S., Neményi, A., Pék, Z., & Helyes, L. (2022). Impact of Plant Growth-Promoting Rhizobacteria Inoculation on the Physiological Response and Productivity Traits of Field-Grown Tomatoes in Hungary. Horticulturae, 8(7), 641. https://doi.org/10.3390/horticulturae8070641