
Metabolite Analysis of Lettuce in Response to Sulfur Nutrition

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Condition
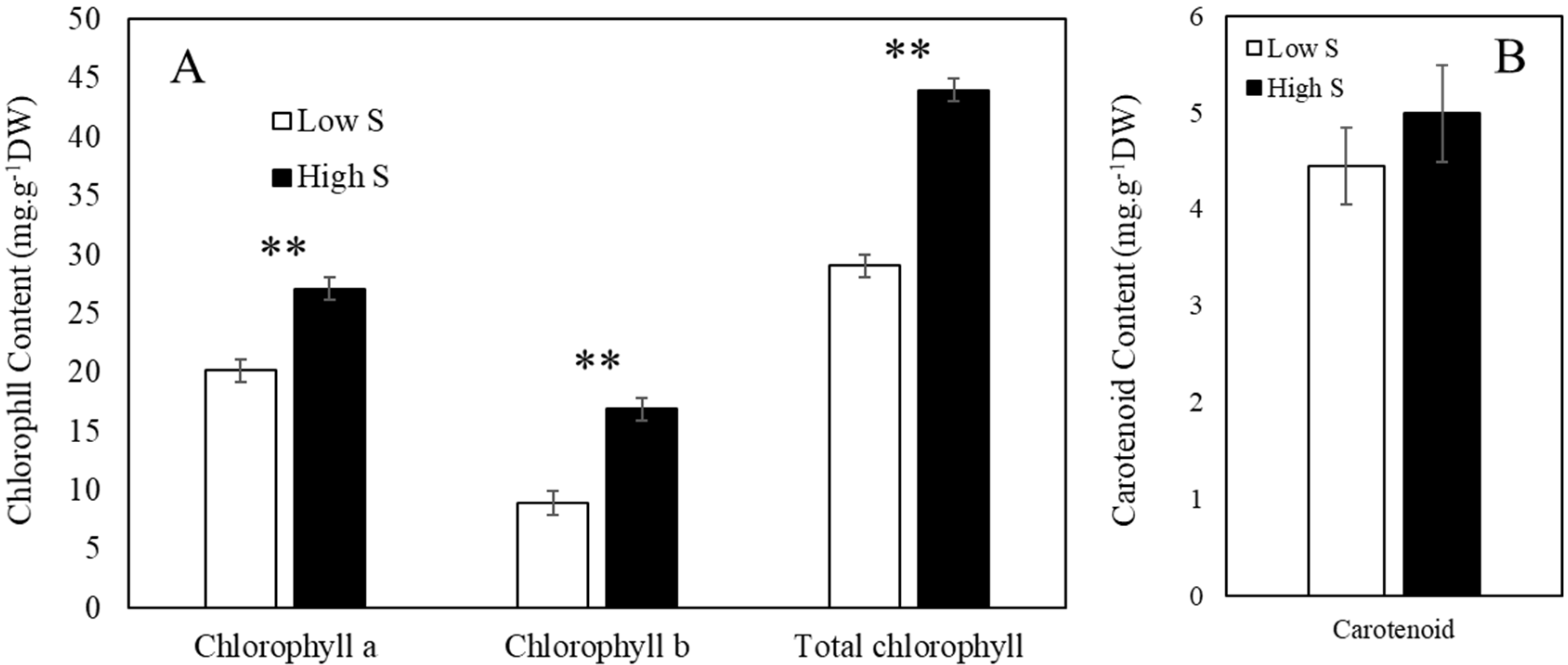
2.2. Analysis of Chlorophyll and Carotenoid Content
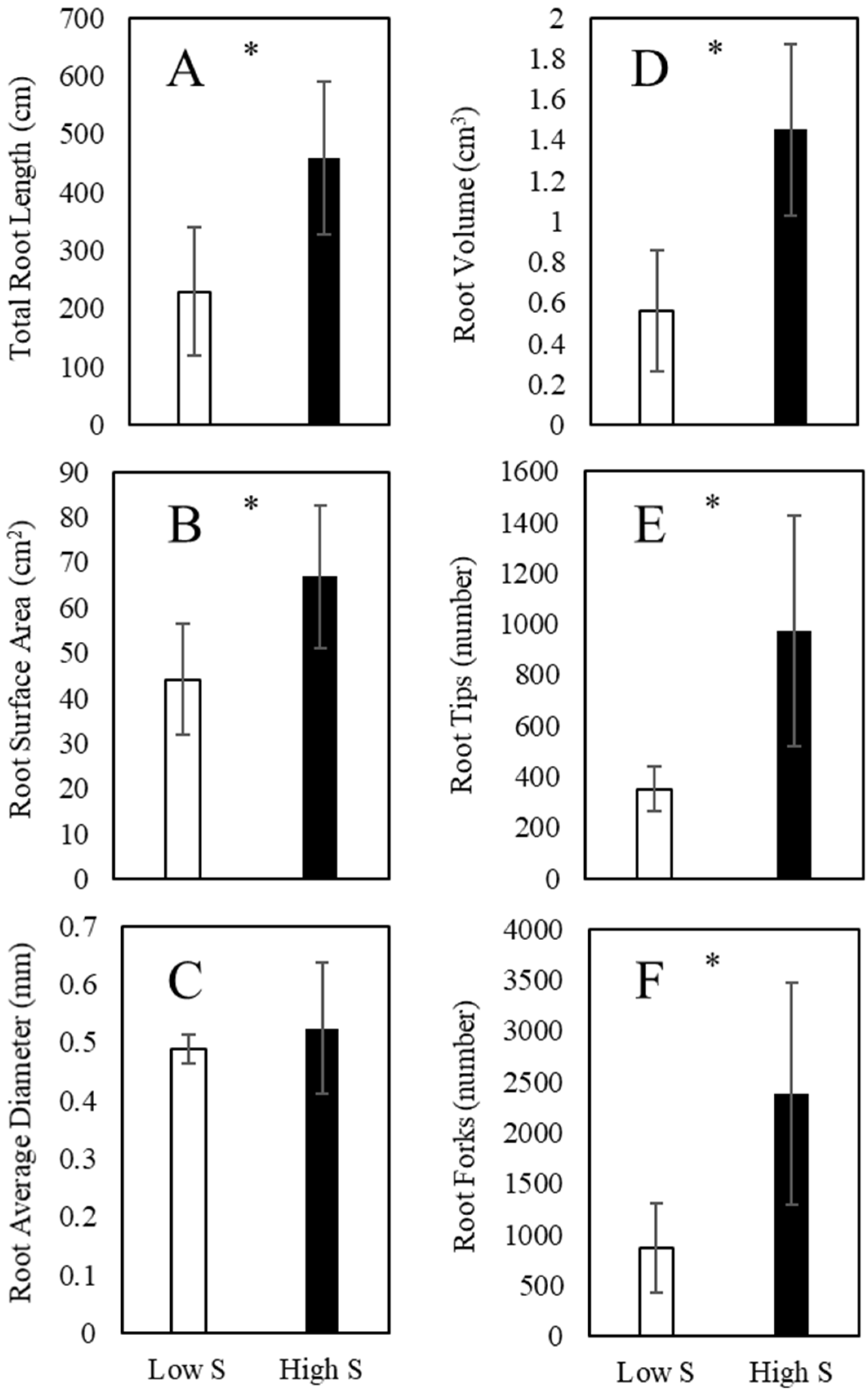
2.3. Root Morphology
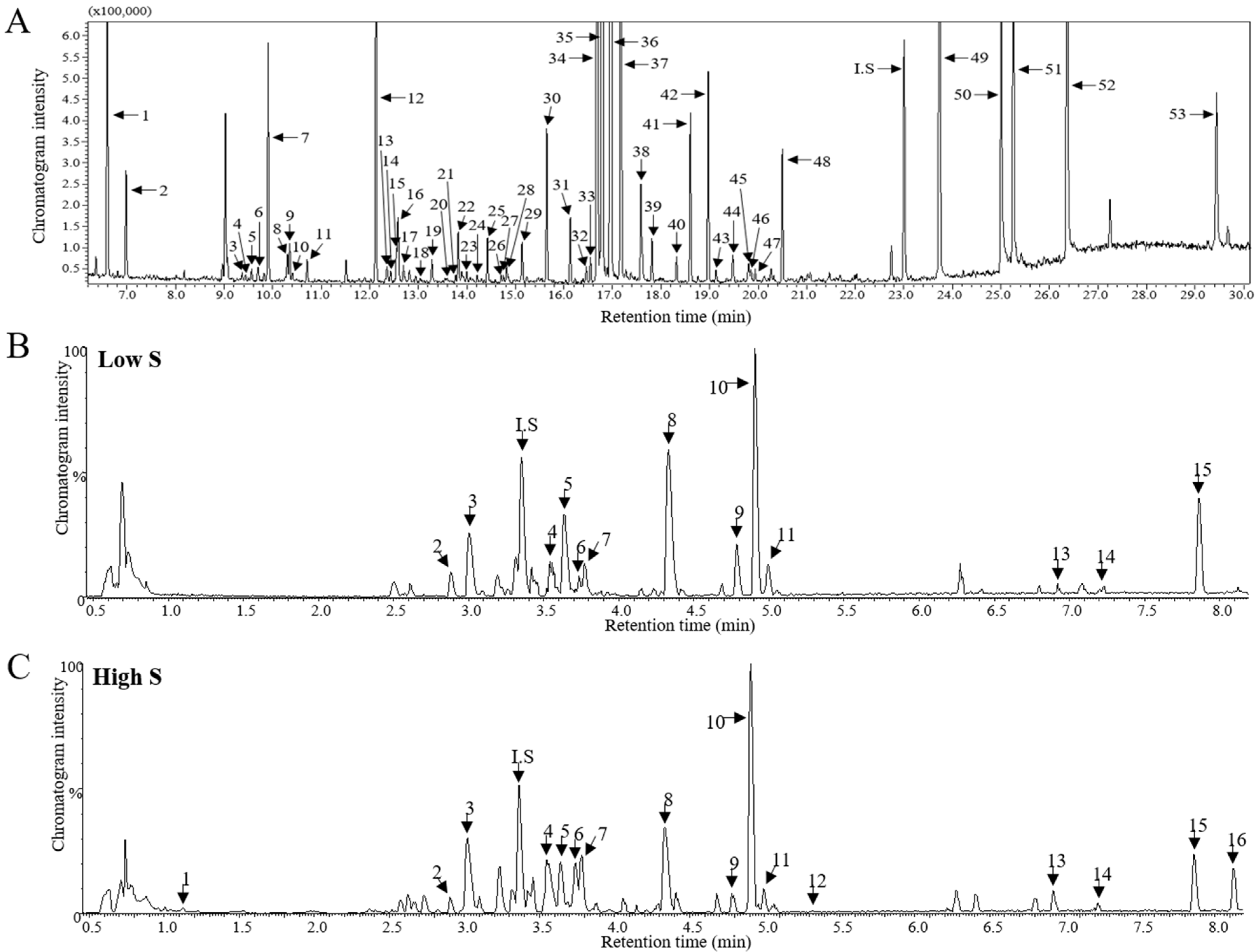
2.4. Analysis of Lettuce Metabolites Using LC-MS
2.5. Analysis of Lettuce Metabolites Using GC-MS
2.6. Data Processing
2.7. Statistical Analysis
3. Results and Discussion
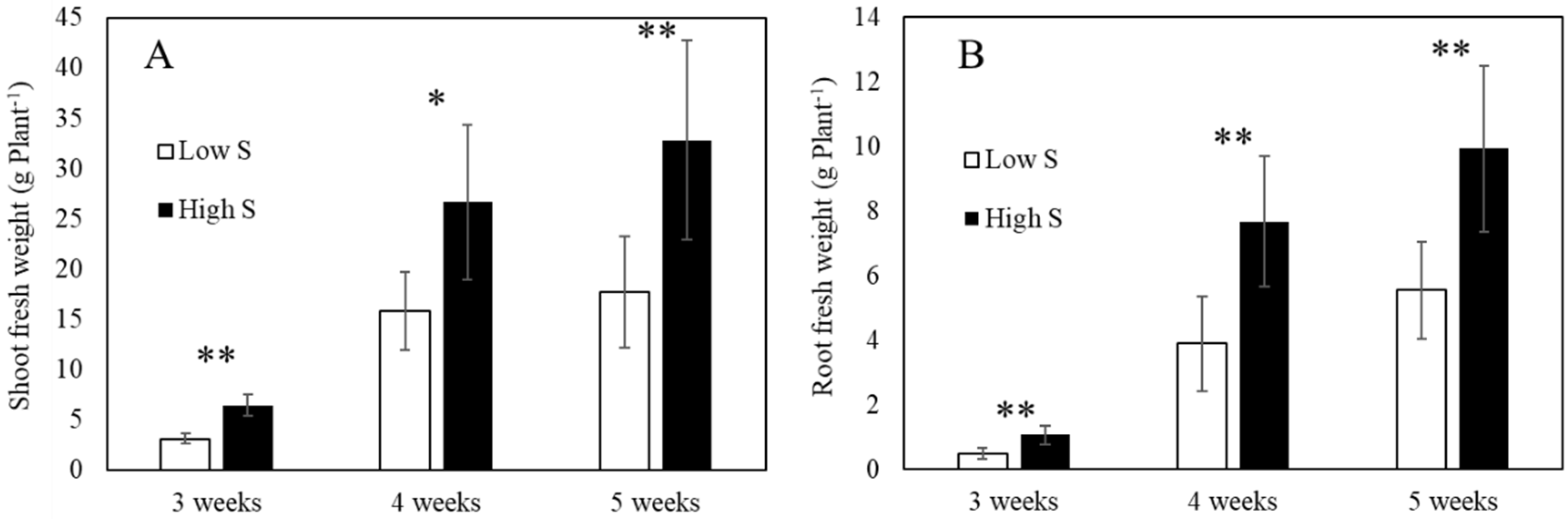
3.1. Lettuce Fresh Weight and Pigment Contents after Sulfur Treatment
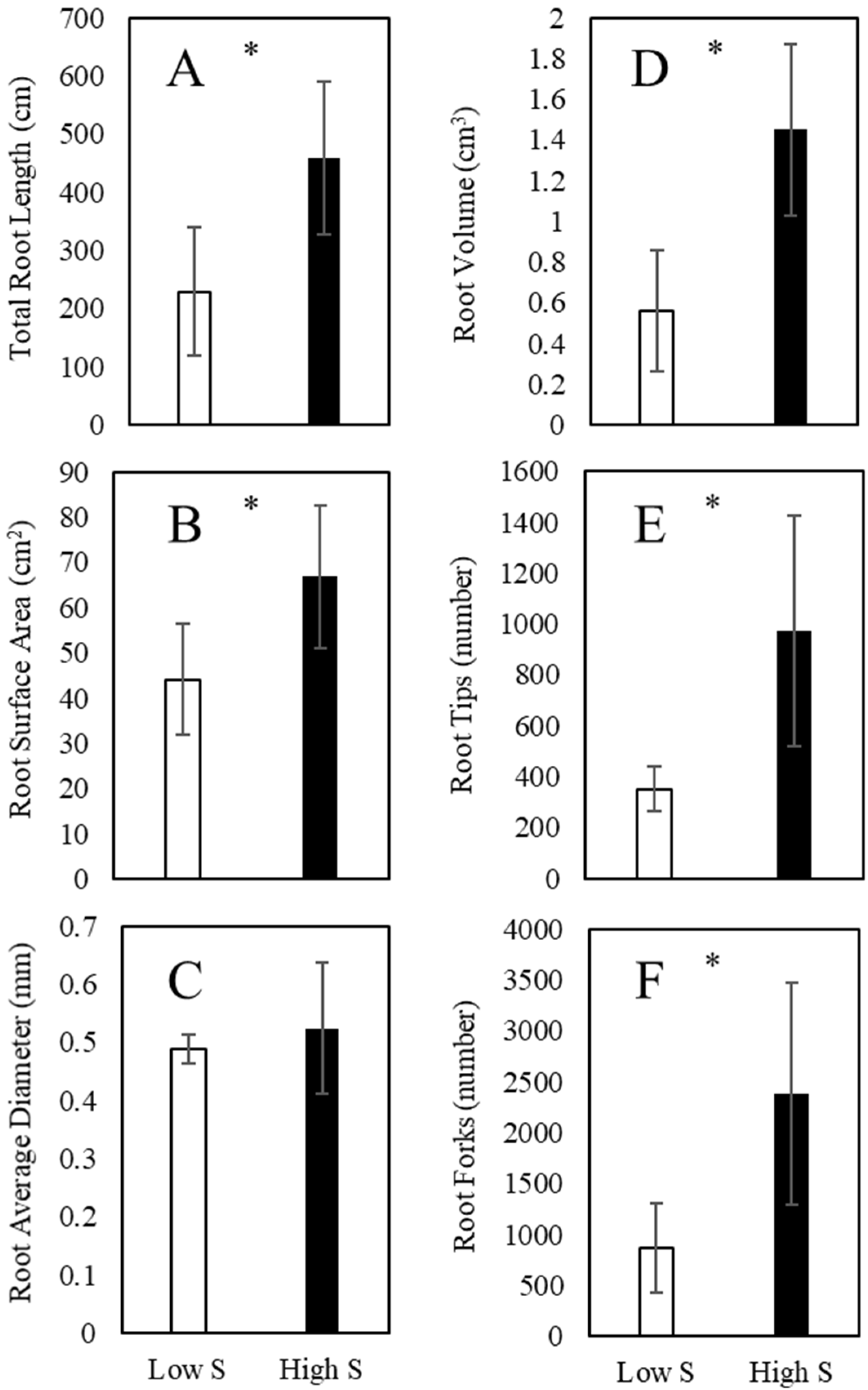
3.2. Root Morphology
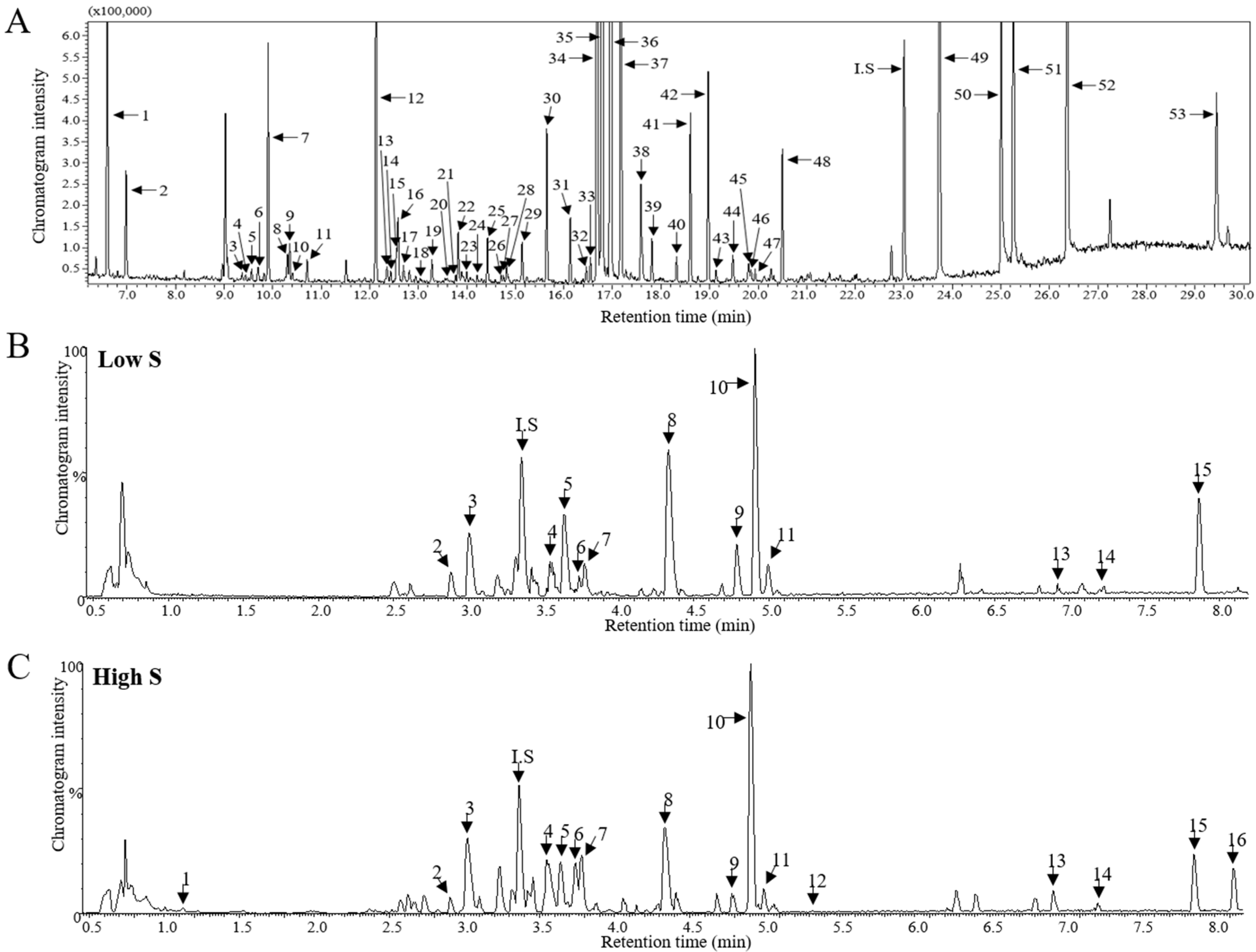
3.3. Metabolomic Analysis and PLS-DA Score Plots
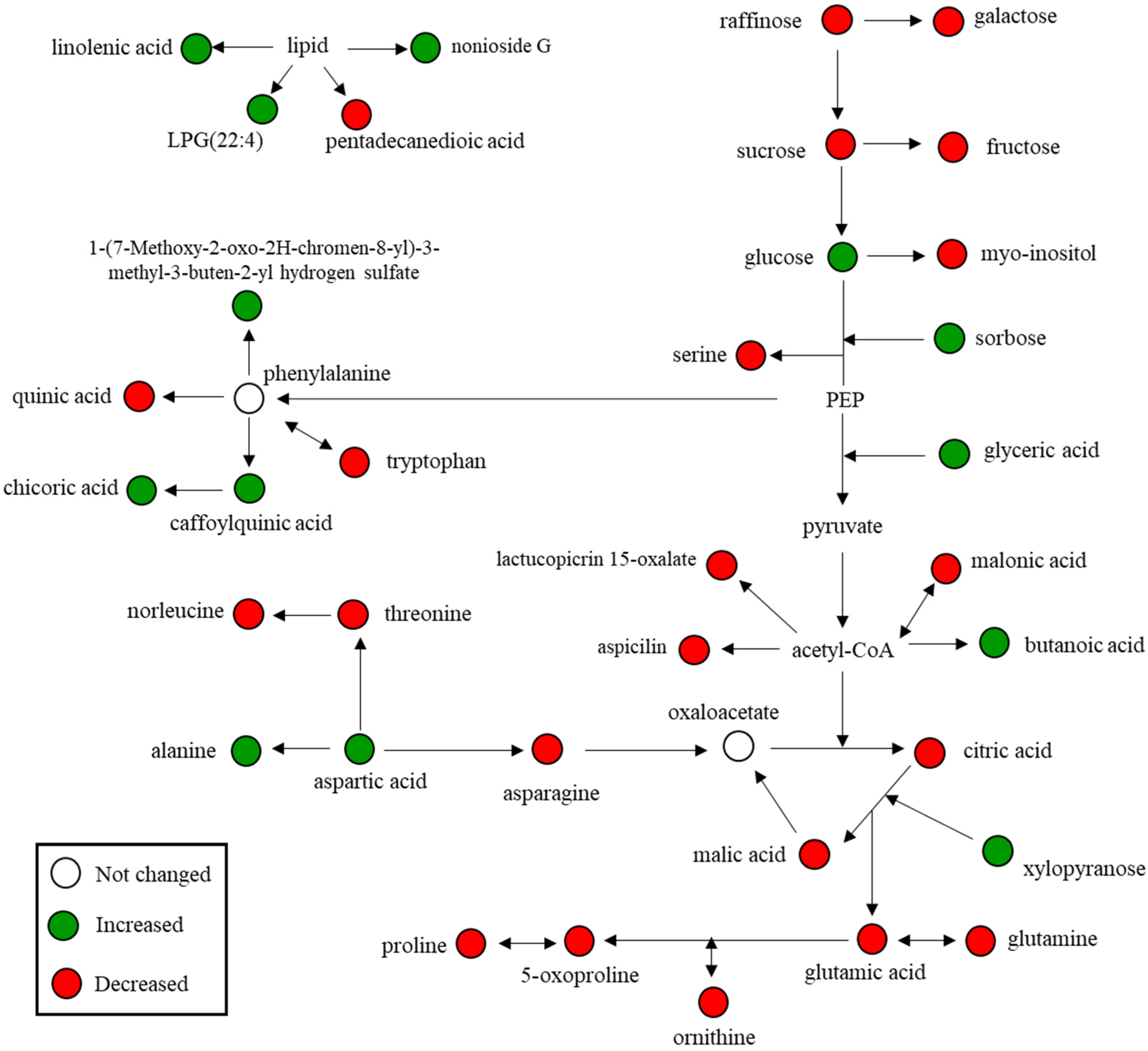
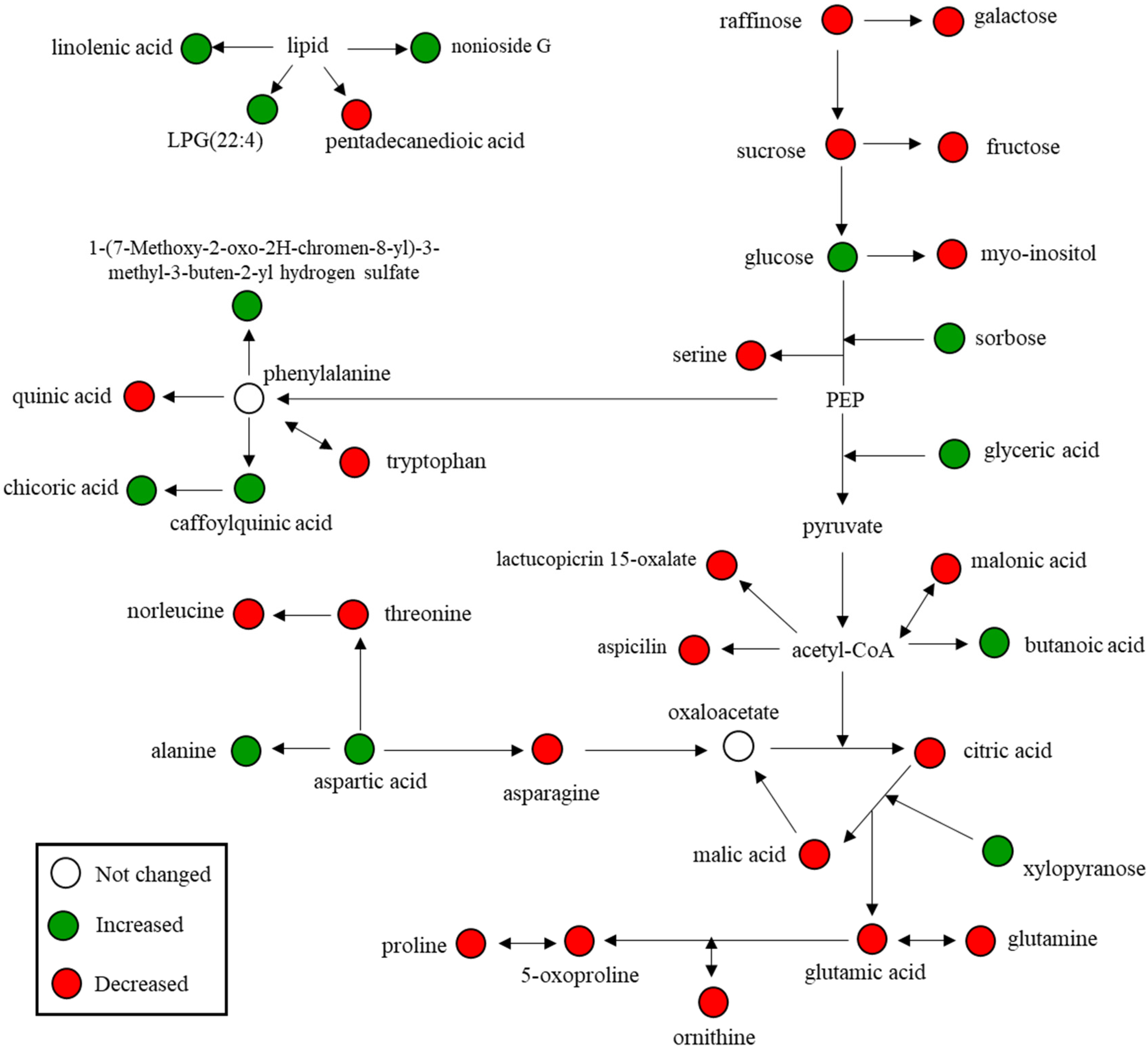
3.4. Relative Abundance of Identified Metabolites and Sulfur Treatment-Related Lettuce Metabolomic Pathway
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Takahashi, H.; Kopriva, S.; Giordano, M.; Saito, K.; Hell, R. Sulfur assimilation in photosynthetic organisms: Molecular functions and regulations of transporters and assimilatory enzymes. Annu. Rev. Plant Biol. 2011, 62, 157–184. [Google Scholar] [CrossRef]
- Abdalla, M.A.; Meschede, C.A.C.; Mühling, K.H. Selenium foliar application alters patterns of glucosinolate hydrolysis products of pak choi Brassica rapa L. var. Chinensis. Sci. Hortic. 2020, 273, 109614. [Google Scholar] [CrossRef]
- Meschede, C.A.C.; Abdalla, M.A.; Mühling, K.H. Sulfur but not nitrogen supply increases the ITC/Nitrile ratio in Pak Choi (Brassica rapa subsp. Chinensis (L.) Hanelt). J. Appl. Bot. Food Qual. 2020, 93, 95–104. [Google Scholar] [CrossRef]
- Kopriva, S.; Malagoli, M.; Takahashi, H. Sulfur nutrition: Impacts on plant development, metabolism, and stress responses. J. Exp. Bot. 2019, 70, 4069–4073. [Google Scholar] [CrossRef] [PubMed]
- Abdalla, M.A.; Mühling, K.H. Plant-derived sulfur containing natural products produced as a response to biotic and abiotic stresses: A review of their structural diversity and medicinal importance. J. Appl. Bot. Food Qual. 2019, 92, 204–215. [Google Scholar] [CrossRef]
- Hawkesford, M.J.; De Kok, L.J. Managing sulphur metabolism in plants. Plant Cell Environ. 2006, 29, 382–395. [Google Scholar] [CrossRef]
- Buchner, P. Plant Sulphate Transporters: Co-Ordination of Uptake, Intracellular, and Long-Distance Transport. J. Exp. Bot. 2004, 55, 1765–1773. [Google Scholar] [CrossRef]
- Buchner, P.; Stuiver, C.E.E.; Westerman, S.; Wirtz, M.; Hell, R.; Hawkesford, M.J.; De Kok, L.J. Regulation of Sulfate Uptake and Expression of Sulfate Transporter Genes in Brassica Oleracea as Affected by Atmospheric H(2)S and Pedospheric Sulfate Nutrition. Plant Physiol. 2004, 136, 3396–3408. [Google Scholar] [CrossRef]
- Etienne, P.; Sorin, E.; Maillard, A.; Gallardo, K.; Arkoun, M.; Guerrand, J.; Cruz, F.; Yvin, J.C.; Ourry, A. Assessment of sulfur deficiency under field conditions by single measurements of sulfur, chloride, and phosphorus in mature leaves. Plants 2018, 7, 37. [Google Scholar] [CrossRef]
- McGrath, S.P.; Zhao, F.J.; Withers, P.J.A. Development of Sulphur Deficiency in Crops and Its Treatment; No 379; Fertiliser Society: Petersborough, UK, 1996. [Google Scholar]
- Yu, Z.; She, M.; Zheng, T.; Diepeveen, D.; Islam, S.; Zhao, Y.; Zhang, Y.; Tang, G.; Zhang, Y.; Zhang, J. Impact and mechanism of sulphur-deficiency on modern wheat farming nitrogen-related sustainability and gliadin content. Commun. Biol. 2021, 4, 945. [Google Scholar] [CrossRef]
- Jobe, T.O.; Zenzen, I.; Rahimzadeh Karvansara, P.; Kopriva, S. Integration of sulfate assimilation with carbon and nitrogen metabolism in transition from C3 to C4 photosynthesis. J. Exp. Bot. 2019, 70, 4211–4221. [Google Scholar] [CrossRef] [PubMed]
- Houhou, M.; Joutei, K.A.; Louhalia, S. Biomass production, chlorophyll content, and morphological parameters are affected by sulfur deficiency in Eruca sativa L. Int. J. Ecol. Environ. Sci. 2018, 44, 67–75. [Google Scholar]
- Henriet, C.; Aimé, D.; Térézol, M.; Kilandamoko, A.; Rossin, N.; Combes-Soia, L.; Labas, V.; Serre, R.F.; Prudent, M.; Kreplak, J.; et al. Water stress combined with sulfur deficiency in pea affects yield components but mitigates the effect of deficiency on seed globulin composition. J. Exp. Bot. 2019, 70, 4287–4304. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Huang, S.; Fan, Y.; Yu, Y.; Wang, Q.; Li, H.; Wan, Y. Sulfur reduces arsenic accumulation in rice shoot by enhancing root retention and altering arsenic metabolism. Chemosphere 2022, 305, 135080. [Google Scholar] [CrossRef]
- Zhang, N.; Huang, L.; Zhang, Y.; Liu, L.; Sun, C.; Lin, X. Sulfur deficiency exacerbates phytotoxicity and residues of imidacloprid through suppression of thiol-dependent detoxification in lettuce seedlings. Environ. Pollut. 2021, 291, 118221. [Google Scholar] [CrossRef]
- Filipek-Mazur, B.; Tabak, M.; Gorczyca, O.; Lisowska, A. Effect of sulfur-containing fertilizers on the quantity and quality of spring oilseed rape and winter wheat yield. J. Elementol. 2019, 24, 1383–1394. [Google Scholar] [CrossRef]
- Carciochi, W.D.; Divito, G.A.; Fernández, L.A.; Echeverría, H.E. Sulfur affects root growth and improves nitrogen recovery and internal efficiency in wheat. J. Plant Nutr. 2017, 40, 1231–1242. [Google Scholar] [CrossRef]
- Wilson, T.L.; Guttieri, M.J.; Nelson, N.O.; Fritz, A.; Tilley, M. Nitrogen and sulfur effects on hard winter wheat quality and asparagine concentration. J. Cereal Sci. 2020, 93, 102969. [Google Scholar] [CrossRef]
- Nakai, Y.; Maruyama-Nakashita, A. Biosynthesis of sulfur-containing small biomolecules in plants. Int. J. Mol. Sci. 2020, 21, 3470. [Google Scholar] [CrossRef]
- Yoshimoto, N.; Saito, K. S-Alk(en)ylcysteine sulfoxides in the genus Allium: Proposed biosynthesis, chemical conversion, and bioactivities. J. Exp. Bot. 2019, 70, 4123–4137. [Google Scholar] [CrossRef]
- Abdalla, M.A.; Lentz, C.; Mühling, K.H. Crosstalk between Selenium and Sulfur Is Associated with Changes in Primary Metabolism in Lettuce Plants Grown under Se and S Enrichment. Plants 2022, 11, 927. [Google Scholar] [CrossRef] [PubMed]
- Camejo, D.; Frutos, A.; Mestre, T.C.; Pinero, M.D.C.; Rivero, R.M.; Martinez, V. Artificial light impacts the physical and nutritional quality of lettuce. Hortic. Environ. Biotechnol. 2020, 61, 69–82. [Google Scholar] [CrossRef]
- Baslam, M.; Pascual, I.; Sanchez-Diaz, M.; Erro, J.; Garcia-Mina, J.M.; Goicoechea, N. Improvement of nutritional quality of greenhouse-grown lettuce by Arbuscular Mycorrhizal fungi is conditioned by the source of phosphorus nutrition. J. Agric. Food Chem. 2011, 59, 11129–11140. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Moon, Y.; Tou, J.C.; Mou, B.; Waterland, N.L.J. Nutritional value, bioactive compounds and health benefits of lettuce (Lactuca sativa L.). J. Food Compos. 2016, 49, 19–34. [Google Scholar] [CrossRef]
- Abdalla, M.A.; Sulieman, S.; Mühling, K.H. Regulation of selenium/sulfur interactions to enhance chemopreventive effects: Lessons to learn from Brassicaceae. Molecules 2020, 25, 5846. [Google Scholar] [CrossRef]
- Garg, M.; Sharma, N.; Sharma, S.; Kapoor, P.; Kumar, A.; Chunduri, V.; Arora, P. Biofortified crops generated by breeding, agronomy, and transgenic approaches are improving lives of millions of people around the world. Front. Nutr. 2018, 5, 12. [Google Scholar] [CrossRef]
- Kane, C.D.; Jasoni, R.L.; Peffley, E.P.; Thompson, L.D.; Green, C.J.; Pare, P. Nutrient solution and solution pH influences on onion growth and mineral content. J. Plant Nutr. 2006, 29, 375–390. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Buschmann, C. Chlorophylls and Carotenoids: Measurement and Characterization by UV-VIS Spectroscopy. In Current Protocols in Food Analytical Chemistry; Wrolstad, R.E., Acree, T.E., An, H., Decker, E.A., Penner, M.H., Reid, D.S., Schwartz, S.J., Shoemaker, C.F., Sporns, P., Eds.; John Wiley and Sons: New York, NY, USA, 2001; pp. F4.3.1–F4.3.8. [Google Scholar] [CrossRef]
- Kim, B.M.; Lee, H.J.; Song, Y.H.; Kim, H.J. Effect of salt stress on the growth, mineral contents, and metabolite profiles of spinach. J. Sci. Food Agric. 2021, 101, 3787–3794. [Google Scholar] [CrossRef]
- Hesse, H.; Nikiforova, V.; Gakiere, B.; Hoefgen, R. Molecular analysis and control of cysteine biosynthesis: Integration of nitrogen and sulphur metabolism. J. Exp. Bot. 2004, 55, 1283–1292. [Google Scholar] [CrossRef]
- Prosser, I.M.; Purves, J.V.; Saker, L.R.; Clarkson, D.T. Rapid disruption of nitrogen metabolism and nitrate transport in spinach plants deprived of sulphate. J. Exp. Bot. 2001, 52, 113–121. [Google Scholar] [CrossRef]
- Satbhai, S.B.; Ristova, D.; Busch, W. Underground tuning: Quantitative regulation of root growth. J. Exp. Bot. 2015, 66, 1099–1112. [Google Scholar] [CrossRef] [PubMed]
- Bates, T.R.; Lynch, J.P. Stimulation of root hair elongation in Arabidopsis thaliana by low phosphorus availability. Plant Cell Environ. 1996, 19, 529–538. [Google Scholar] [CrossRef]
- Teranishi, R.; Buttery, R.G.; Shahidi, F. Role of Free Amino Acids and Peptides in Food Taste; ACS Symposium Series; American Chemical Society: Washington, DC, USA, 1989; Flavor Chemistry Chapter 13; pp. 158–174. [Google Scholar]
- Galili, G. The aspartate-family pathway of plants: Linking production of essential amino acids with energy and stress regulation. Plant Signal. Behav. 2011, 6, 192–195. [Google Scholar] [CrossRef] [PubMed]
- Cuppettz, S.L.; McCluskey, M.M.; Paparozzi, E.T.; Parkhursp, A. Nitrogen and sulfur effects on leaf lettuce quality. J. Food Qual. 1999, 22, 363–373. [Google Scholar] [CrossRef]
- Long, S.R.; Kahn, M.; Seefeldt, L.; Tsay, Y.F.; Kopriva, S. Chapter 16 nitrogen and sulfur. In Biochemistry & Molecular Biology of Plants; Buchana, B.B., Gruissem, W., Jones, R.L., Eds.; Wiley Blackwell: Hoboken, NJ, USA, 2015; pp. 746–768. [Google Scholar]
- Maruyama-Nakashita, A. Metabolic changes sustain the plant life in low-sulfur environments. Curr. Opin. Plant Biol. 2017, 39, 144–151. [Google Scholar] [CrossRef]
- Falk, K.L.; Tokuhisa, J.G.; Gershenzon, J. The effect of sulfur nutrition on plant glucosinolate content: Physiology and molecular mechanisms. Plant Biol. (Stuttg) 2007, 9, 573–581. [Google Scholar] [CrossRef]
- Maruyama-Nakashita, A.; Nakamura, Y.; Tohge, T.; Saito, K.; Takahashi, H. Arabidopsis SLIM1 is a central transcriptional regulator of plant sulfur response and metabolism. Plant Cell. 2006, 18, 3235–3251. [Google Scholar] [CrossRef]
- Noctor, G.; Queval, G.; Mhamdi, A.; Chaouch, S.; Foyer, C.H. Glutathione. In The Arabidopsis Book; BioOne: Washington, DC, USA, 2011; Volume 9, p. e0142. [Google Scholar] [CrossRef]
- Maruyama-Nakashita, A.; Nakamura, Y.; Watanabe-Takahashi, A.; Inoue, E.; Yamaya, T.; Takahashi, H. Identification of a novel cis-acting element conferring sulfur deficiency response in Arabidopsis roots. Plant J. 2005, 42, 305–314. [Google Scholar] [CrossRef]
- Aarabi, F.; Kusajima, M.; Tohge, T.; Konishi, T.; Gigolashvili, T.; Takamune, M.; Sasazaki, Y.; Watanabe, M.; Nakashita, H.; Fernie, A.R.; et al. Sulfur deficiency-induced repressor proteins optimize glucosinolate biosynthesis in plants. Sci. Adv. 2016, 2, e1601087. [Google Scholar] [CrossRef]
- Abdalla, M.A.; Li, F.; Wenzel-Storjohann, A.; Sulieman, S.; Tasdemir, D.; Mühling, K.H. Comparative metabolite profile, biological activity and overall quality of three lettuce (Lactuca sativa L., Asteraceae) cultivars in response to sulfur nutrition. Pharmaceutics 2021, 13, 713. [Google Scholar] [CrossRef]
- Chadwick, M.; Gawthrop, F.; Michelmore, R.W.; Wagstaff, C.; Methven, L.L. Perception of bitterness, sweetness and liking of different genotypes of lettuce. Food Chem. 2016, 197, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Nikiforova, V.; Freitag, J.; Kempa, S.; Adamik, M.; Hesse, H.; Hoefgen, R. Transcriptome analysis of sulfur depletion in Arabidopsis thaliana: Interlacing of biosynthetic pathways provides response specificity. Plant J. 2003, 33, 633–650. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.; Smith, D.L. Flavonoids in agriculture: Chemistry and roles in, biotic and abiotic stress responses, and microbial associations. Agronomy 2020, 10, 1209. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
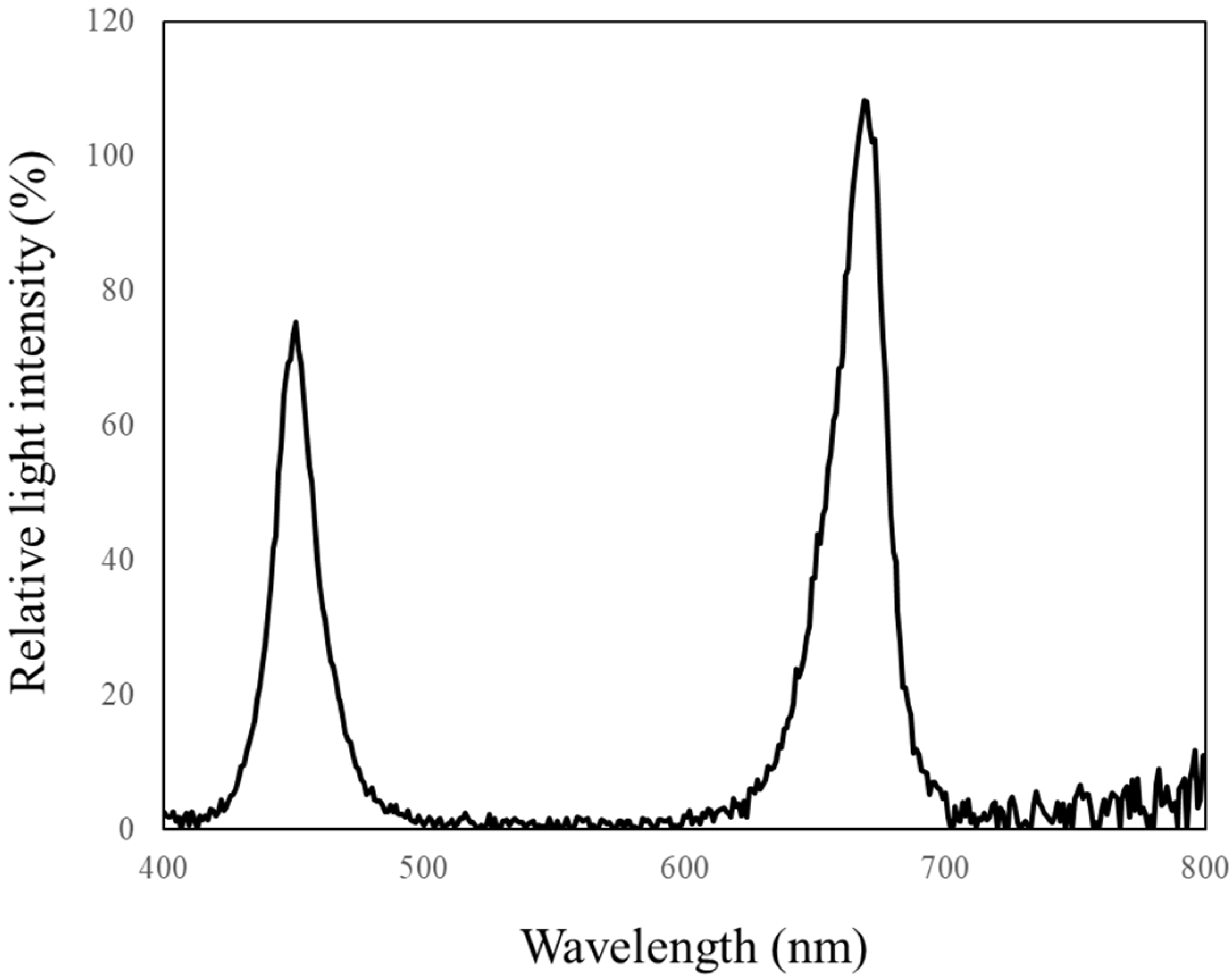
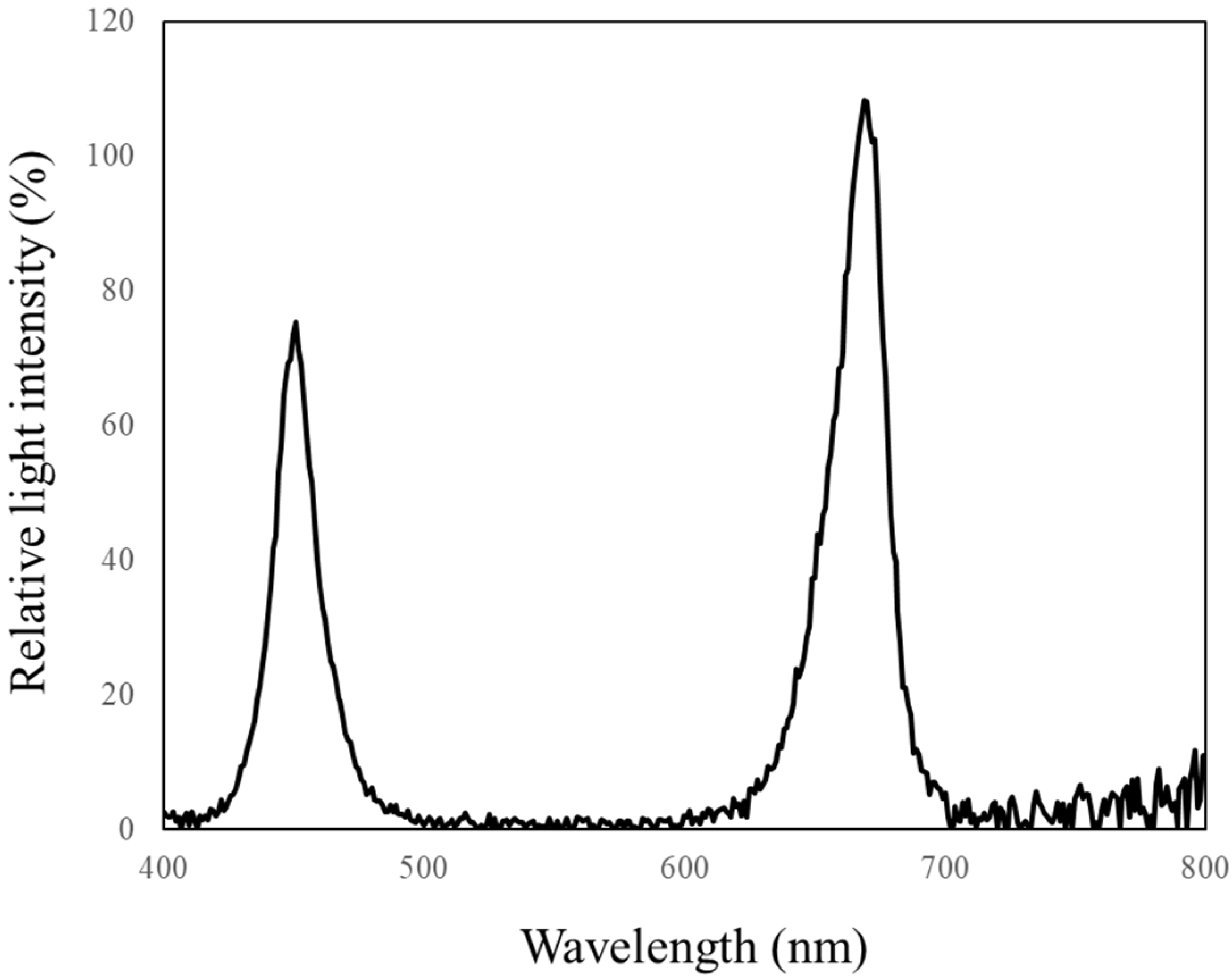
| Light | PPFD * | Blue (400–499 nm) | Green (500–599 nm) | Red (600–699 nm) | Far-Red (700–799 nm) |
|---|---|---|---|---|---|
| Red and Blue | 232 | 70.9 (33.3%) | 3.6 (1.7%) | 119.9 (56.3%) | 13.1 (6.1%) |
| Metabolites | RT a (min) | RI b | VIP c | p-Value d | Fold Change (vs. Low Sulfur) |
|---|---|---|---|---|---|
| Alanine | 6.36 | 1099 | 1.07 | 3.42 × 10−2 | 1.36 |
| Norleucine | 9.38 | 1288 | 1.11 | 2.34 × 10−2 | −1.68 |
| Proline | 9.45 | 1293 | 1.19 | 5.20 × 10−3 | −2.66 |
| Glyceric acid | 9.91 | 1323 | 1.26 | 5.84 × 10−5 | 2.83 |
| Serine | 10.36 | 1353 | 1.26 | 4.42 × 10−5 | −2.47 |
| Threonine | 10.72 | 1376 | 1.26 | 4.30 × 10−5 | −3.21 |
| Malic acid | 12.13 | 1481 | 1.03 | 4.86 × 10−2 | −1.05 |
| Aspartic acid | 12.55 | 1514 | 1.26 | 3.01 × 10−3 | + e |
| 5-Oxoproline | 12.59 | 1516 | 1.27 | 2.62 × 10−6 | −11.10 |
| Butanoic acid | 12.70 | 1525 | 1.20 | 3.89 × 10−3 | 4.33 |
| Ornithine | 13.77 | 1609 | 1.26 | 2.13 × 10−3 | - f |
| Glutamic acid | 13.83 | 1614 | 1.11 | 2.13 × 10−2 | −1.28 |
| Asparagine | 14.43 | 1663 | 1.26 | 8.76 × 10−4 | −5.37 |
| Malonic acid | 14.76 | 1690 | 1.27 | 2.42 × 10−7 | - |
| Glutamine | 15.66 | 1767 | 1.26 | 9.88 × 10−4 | - |
| Citric acid | 16.14 | 1809 | 1.26 | 1.69 × 10−5 | −2.97 |
| Quinic acid | 16.56 | 1847 | 1.05 | 3.98 × 10−2 | −1.32 |
| Fructose | 16.69 | 1860 | 1.26 | 2.12 × 10−5 | 3.92 |
| Sorbose | 16.79 | 1869 | 1.26 | 1.44 × 10−3 | 3.84 |
| Glucose | 16.97 | 1885 | 1.20 | 4.55 × 10−3 | 1.45 |
| Galactose | 17.18 | 1904 | 1.25 | 1.37 × 10−4 | 1.42 |
| Inositol | 17.59 | 1944 | 1.26 | 1.98 × 10−5 | −1.35 |
| Myo-inositol | 18.98 | 2080 | 1.26 | 1.08 × 10−4 | −1.37 |
| Xylopyranose | 19.86 | 2171 | 1.09 | 2.76 × 10−2 | 3.09 |
| Phosphoric acid | 19.95 | 2180 | 1.15 | 1.18 × 10−2 | −1.22 |
| Sucrose | 23.75 | 2620 | 1.25 | 2.60 × 10−4 | −1.67 |
| Oleamide | 25.27 | 2831 | 1.15 | 1.17 × 10−2 | −1.74 |
| Raffinose | 29.45 | 2988 | 1.27 | 1.87 × 10−6 | −12.99 |
| Compound | RT a | Exact Mass (M-H) | MS Fragments | VIP b | p-Value c | Fold Change (vs. Low Sulfur) |
|---|---|---|---|---|---|---|
| Tryptophan | 2.90 | 203.0879 | 186, 142, 116, 74 | 1.46 | 1.15 × 10−2 | −1.52 |
| Caffeoylquinic acid | 3.01 | 353.0919 | 191, 179, 133 | 1.51 | 5.23 × 10−3 | 1.53 |
| Chicoric acid | 3.56 | 473.0719 | 427, 311, 293, 179 | 1.58 | 2.28 × 10−4 | 8.32 |
| 1-(7-Methoxy-2-oxo-2H-chromen-8-yl)-3-methyl-3-buten-2-yl hydrogen sulfate | 3.77 | 339.0563 | 175, 96 | 1.58 | 2.42 × 10−4 | 2.84 |
| Lactucopicrin 15-oxalate | 4.33 | 481.1132 | 409, 257, 151 | 1.39 | 2.57 × 10−2 | −1.49 |
| Aspicilin | 4.79 | 327.2213 | 211, 171 | 1.55 | 1.67 × 10−3 | −2.59 |
| Nonioside G | 4.90 | 755.3664 | 681, 561, 161, 159 | 1.49 | 7.83 × 10−3 | 1.42 |
| LPG(22:4) | 6.89 | 559.3089 | 277, 116 | 1.52 | 2.39 × 10−2 | 4.70 |
| Pentadecanedioic acid | 7.86 | 271.1976 | 199 | 1.56 | 1.19 × 10−3 | −1.43 |
| Linolenic acid | 8.12 | 277.2211 | 144, 116 | 1.58 | 3.44 × 10−4 | 12.97 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chung, J.-S.; Kim, H.-C.; Yun, S.-M.; Kim, H.-J.; Kim, C.-S.; Lee, J.-J. Metabolite Analysis of Lettuce in Response to Sulfur Nutrition. Horticulturae 2022, 8, 734. https://doi.org/10.3390/horticulturae8080734
Chung J-S, Kim H-C, Yun S-M, Kim H-J, Kim C-S, Lee J-J. Metabolite Analysis of Lettuce in Response to Sulfur Nutrition. Horticulturae. 2022; 8(8):734. https://doi.org/10.3390/horticulturae8080734
Chicago/Turabian StyleChung, Jung-Sung, Hyeong-Cheol Kim, Su-Min Yun, Hyun-Jin Kim, Cheol-Soo Kim, and Jeung-Joo Lee. 2022. "Metabolite Analysis of Lettuce in Response to Sulfur Nutrition" Horticulturae 8, no. 8: 734. https://doi.org/10.3390/horticulturae8080734
APA StyleChung, J.-S., Kim, H.-C., Yun, S.-M., Kim, H.-J., Kim, C.-S., & Lee, J.-J. (2022). Metabolite Analysis of Lettuce in Response to Sulfur Nutrition. Horticulturae, 8(8), 734. https://doi.org/10.3390/horticulturae8080734