
Impact of Biologically and Chemically Synthesized Zinc Oxide Nanoparticles on Seed Germination and Seedlings’ Growth
 , ,
, ,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Seed Germination Test
Physiological Parameters of Seedlings
2.2. Determination of Photosynthetic Pigments
2.3. Content of Bioactive Compounds
2.4. Enzymatic Activity
2.5. Statistical Analysis
3. Results and Discussion
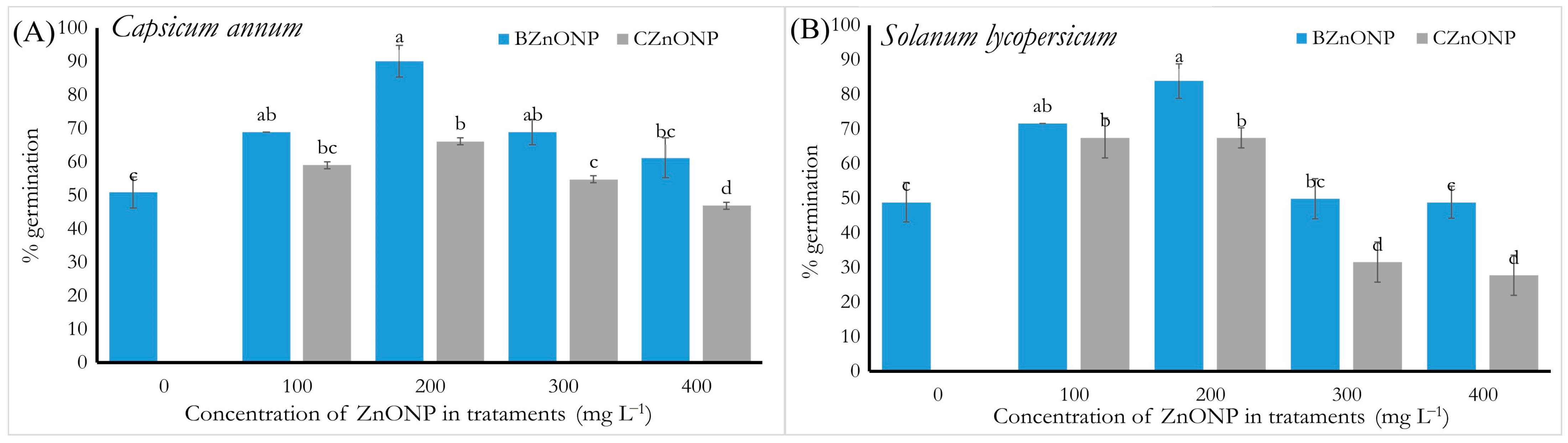
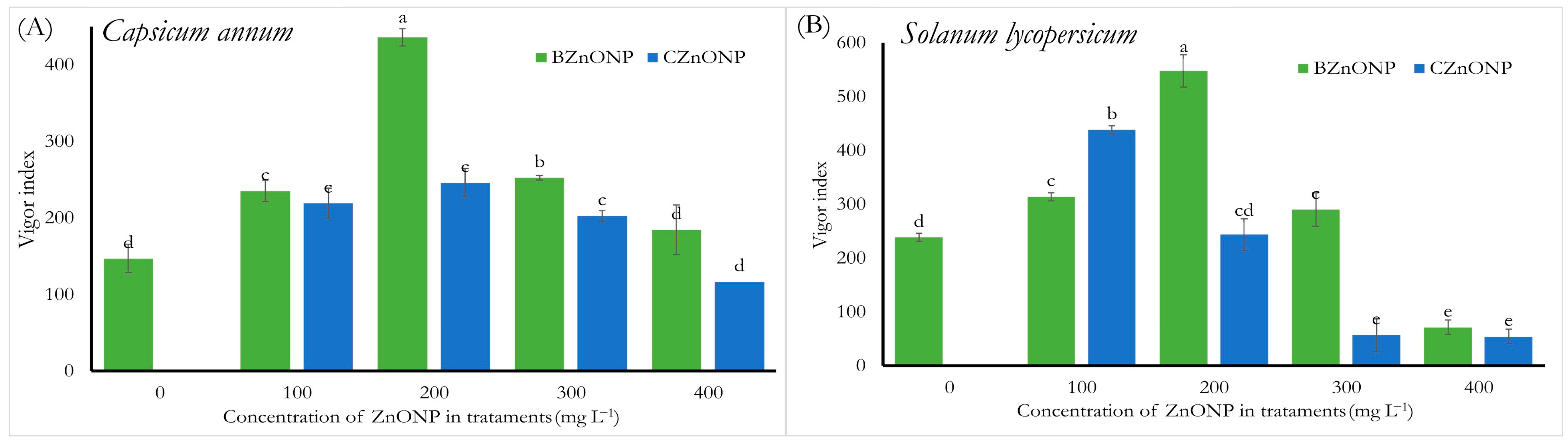
3.1. Physiological Parameters
3.2. Determination of Photosynthetic Pigments
3.2.1. Total Chlorophyll
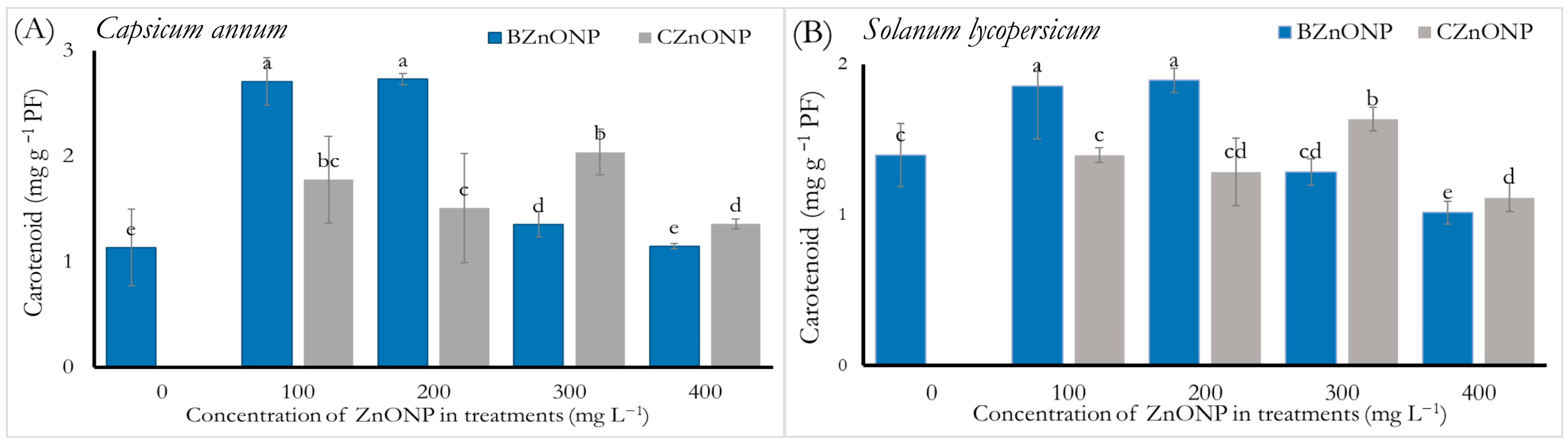
3.2.2. Carotenoids
3.3. Bioactive Compounds
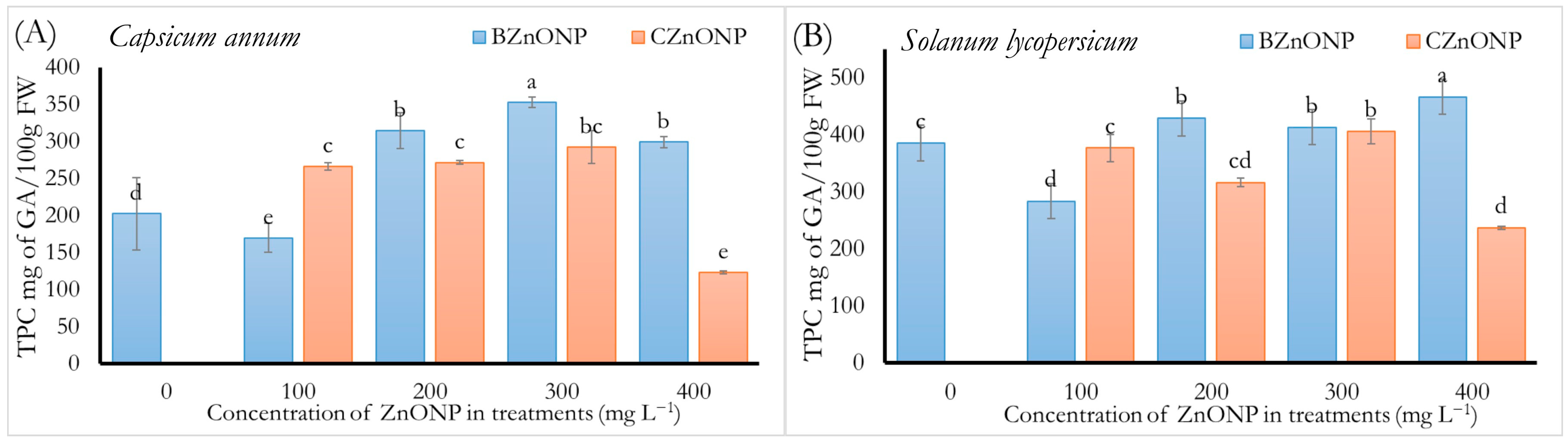
3.3.1. Total Phenols Content, TPC
3.3.2. Flavonoids
3.3.3. Antioxidant Activity
3.4. Enzymatic Activity
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Chhipa, H.; Joshi, P. Nanofertilisers, Nanopesticides and Nanosensors in Agriculture. Nanosci. Food Agric. 2016, 1, 247–282. [Google Scholar] [CrossRef]
- Fatima, F.; Hashim, A.; Anees, S. Efficacy of nanoparticles as nanofertilizer production: A review. Environ. Sci. Pollut. Res. 2021, 28, 1292–1303. [Google Scholar] [CrossRef] [PubMed]
- Faizan, M.; Hayat, S.; Pichtel, J. Effects of Zinc Oxide Nanoparticles on Crop Plants: A Perspective Analysis. In Sustainable Agriculture Reviews 41, 1st ed.; Hayat, S., Pichtel, J., Faizan, M., Fariduddin, Q., Eds.; Springer Nature AG: Cham, Switzerland, 2020; pp. 83–99. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Almutairi, K.F.; Alotaibi, M.; Shami, A.; Alhammad, B.A.; Battaglia, M.L. Nano-fertilization as an emerging fertilization technique: Why can modern agriculture benefit from its use? Plants 2021, 10, 2. [Google Scholar] [CrossRef] [PubMed]
- Ogunyemi, S.O.; Abdallah, Y.; Zhang, M.; Fouad, H.; Hong, X.; Ibrahim, E.; Masum, M.M.I.; Hossain, A.; Mo, J. Green synthesis of zinc oxide nanoparticles using different plant extracts and their antibacterial activity against Xanthomonas oryzae pv. oryzae. Artif. Cells Nanomed. Biotechnol. 2019, 47, 341–352. [Google Scholar] [CrossRef]
- Kumari, M.; Khan, S.S.; Pakrashi, S.; Mukherjee, A.; Chandrasekaran, N. Cytogenetic and genotoxic effects of zinc oxide nanoparticles on root cells of Allium cepa. J. Hazard. Mater. 2011, 190, 613–621. [Google Scholar] [CrossRef] [PubMed]
- Raliya, R.; Nair, R.; Chavalmane, S.; Wang, W.N.; Biswas, P. Mechanistic evaluation of translocation and physiological impact of titanium dioxide and zinc oxide nanoparticles on the tomato (Solanum lycopersicum L.) plant. Metallomics 2015, 7, 1584–1594. [Google Scholar] [CrossRef] [PubMed]
- Rizwan, M.; Ali, S.; Zia ur Rehman, M.; Adrees, M.; Arshad, M.; Qayyum, M.F.; Ali, L.; Hussain, A.; Chatha, S.A.S.; Imran, M. Alleviation of cadmium accumulation in maize (Zea mays L.) by foliar spray of zinc oxide nanoparticles and biochar to contaminated soil. Environ. Pollut. 2019, 248, 358–367. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.H.; Sobze, J.-M.; Pham, T.H.; Nadeem, M.; Liu, C.; Galagedara, L.; Cheema, M.; Thomas, R. Carbon nanoparticles functionalized with carboxylic acid improved the germination and seedling vigor in upland boreal forest species. Nanomaterials 2020, 10, 176. [Google Scholar] [CrossRef]
- El-Badri, A.M.; Batool, M.; Wang, C.; Hashem, A.M.; Tabl, K.M.; Nishawy, E.; Kuai, J.; Zhou, G.; Wang, B. Selenium and zinc oxide nanoparticles modulate the molecular and morpho-physiological processes during seed germination of Brassica napus under salt stress. Ecotoxicol. Environ. Saf. 2021, 225, 112695. [Google Scholar] [CrossRef]
- Nile, S.H.; Thiruvengadam, M.; Wang, Y.; Samynathan, R.; Shariati, M.A.; Rebezov, M.; Nile, A.; Sun, M.; Venkidasamy, B.; Xiao, J.; et al. Nano-priming as emerging seed priming technology for sustainable agriculture—Recent developments and future perspectives. J. Nanobiotechnol. 2022, 20, 1–31. [Google Scholar] [CrossRef]
- Thangavelu, R.M.; Munisamy, B.; Krishnan, K. Effect of deoxycholate capped silver nanoparticles in seed dormancy breaking of Withania somnifera. Curr. Sci. 2019, 116, 952–958. [Google Scholar] [CrossRef]
- Amezcua, J.C.; Lara, M. El Zinc en las Plantas. Ciencia 2017, 68, 28–35. Available online: http://www.smart-fertilizer.com/es/articles/zinc-in-plants (accessed on 1 September 2023).
- Paramo, L.A.; Feregrino-Pérez, A.A.; Guevara, R.; Mendoza, S.; Esquivel, K. Nanoparticles in agroindustry: Applications, toxicity, challenges, and trends. Nanomaterials 2020, 10, 1654. [Google Scholar] [CrossRef] [PubMed]
- Bandeira, M.; Giovanela, M.; Roesch-Ely, M.; Devine, D.M.; da Silva Crespo, J. Green synthesis of zinc oxide nanoparticles: A review of the synthesis methodology and mechanism of formation. Sustain. Chem. Pharm. 2020, 15, 100223. [Google Scholar] [CrossRef]
- Haq, A.N.U.; Nadhman, A.; Ullah, I.; Mustafa, G.; Yasinzai, M.; Khan, I. Synthesis Approaches of Zinc Oxide Nanoparticles: The Dilemma of Ecotoxicity. J. Nanomater. 2017, 2017, 8510342. [Google Scholar] [CrossRef]
- Sur, U.K.; Ankamwar, B.; Karmakar, S.; Halder, A.; Das, P. Green synthesis of Silver nanoparticles using the plant extract of Shikakai and Reetha. Mater. Today Proc. 2018, 5, 2321–2329. [Google Scholar] [CrossRef]
- Jamkhande, P.G.; Ghule, N.W.; Bamer, A.H.; Mohan, G.; Ghule, N.W.; Bamer, A.H.; Kalaskar, M.G. Metal nanoparticles synthesis: An overview on methods of preparation, advantages and disadvantages, and applications. J. Drug Deliv. Sci. Technol. 2019, 53, 101174. [Google Scholar] [CrossRef]
- González-Fernández, J.V.; Pinzón-Moreno, D.D.; Neciosup-Puican, A.A.; Carranza-Oropeza, M.V. Green method, optical and structural characterization of ZnO nanoparticles synthesized using leaves extract of M. Oleifera. J. Renew. Mater. 2022, 10, 833–847. [Google Scholar] [CrossRef]
- Rada, S.S.; Sani, A.M.; Mohseni, S. Biosynthesis, characterization and antimicrobial activities of zinc oxide nanoparticles from leaf extract of Mentha pulegium (L.). Microb. Pathog. 2019, 131, 239–245. [Google Scholar] [CrossRef]
- Sánchez-Pérez, D.M.; Flores-Loyola, E.; Márquez-Guerrero, S.Y.; Galindo-Guzman, M.; Marszalek, J.E. Green Synthesis and Characterization of Zinc Oxide Nanoparticles Using Larrea tridentata Extract and Their Impact on the In-Vitro Germination and Seedling Growth of Capsicum annuum. Sustainability 2023, 15, 3080. [Google Scholar] [CrossRef]
- Pereira, T.M.; Polez, V.L.P.; Sousa, M.H.; Silva, L.P. Modulating physical, chemical, and biological properties of silver nanoparticles obtained by green synthesis using different parts of the tree Handroanthus heptaphyllus (Vell.) Mattos. Colloids Interface Sci. Commun. 2020, 34, 100224. [Google Scholar] [CrossRef]
- Gour, A.; Jain, N.K. Advances in green synthesis of nanoparticles. Artif. Cells Nanomed. Biotechnol. 2019, 47, 844–851. [Google Scholar] [CrossRef] [PubMed]
- Awasthi, A.; Bansal, S.; Jangir, L.K.; Awasthi, G.; Awasthi, K.K.; Awasthi, K. Effect of ZnO Nanoparticles on Germination of Triticum aestivum Seeds. Macromol. Symp. 2017, 376, 1–5. [Google Scholar] [CrossRef]
- Nguyen, D.T.C.; Le, H.T.; Nguyen, T.T.; Nguyen, T.T.T.; Bach, L.G.; Nguyen, T.D.; Van Tran, T. Multifunctional ZnO nanoparticles bio-fabricated from Canna indica L. flowers for seed germination, adsorption, and photocatalytic degradation of organic dyes. J. Hazard. Mater. 2021, 420, 126586. [Google Scholar] [CrossRef] [PubMed]
- Shaban, A.S.; Owda, M.E.; Basuoni, M.M.; Mousa, M.A.; Radwan, A.A.; Saleh, A.K. Punica granatum peel extract mediated green synthesis of zinc oxide nanoparticles: Structure and evaluation of their biological applications. Biomass Convers. Biorefinery 2022, 0123456789. [Google Scholar] [CrossRef]
- Asmat-Campos, D.; López-Medina, E.; de Oca-Vásquez, G.M.; Gil-Rivero, E.; Delfín-Narciso, D.; Juárez-Cortijo, L.; Villena-Zapata, L.; Gurreonero-Fernández, J.; Rafael-Amaya, R. ZnO Nanoparticles Obtained by Green Synthesis as an Alternative to Improve the Germination Characteristics of L. esculentum. Molecules 2022, 27, 2343. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Afzal, S.; Singh, N.K. Nanopriming with phytosynthesized zinc oxide nanoparticles for promoting germination and starch metabolism in rice seeds. J. Biotechnol. 2021, 336, 64–75. [Google Scholar] [CrossRef] [PubMed]
- Tymoszuk, A.; Wojnarowicz, J. Zinc oxide and zinc oxide nanoparticles impact on in vitro germination and seedling growth in Allium cepa L. Materials 2020, 13, 2784. [Google Scholar] [CrossRef]
- Pratap, V.; Durgesh, K.; Sheo, M.; Devendra, K.; Singh, S. Plant Responses to Nanomaterials, 1st ed.; Springer Nature AG: Cham, Switzerland, 2021; Available online: http://link.springer.com/10.1007/978-3-030-36740-4 (accessed on 1 September 2023).
- García-López, J.I.; Lira-Saldivar, R.H.; Zavala-García, F.; Olivares-Sáenz, E.; Niño-Medina, G.; Ruiz-Torres, N.A.; Méndez-Argüello, B.; Díaz-Barriga, E. Effects of zinc oxide nanoparticles on growth and antioxidant enzymes of Capsicum chinense. Toxicol. Environ. Chem. 2018, 100, 560–572. [Google Scholar] [CrossRef]
- Kujur, A.B.; Lal, G.M. Effect of hydropriming and osmopriming on germination behaviour and vigor of soybean (Glycine max L.) seeds. Agric. Sci. Dig. A Res. J. 2015, 35, 207–210. [Google Scholar] [CrossRef]
- García-López, J.I.; Zavala-García, F.; Olivares-Sáenz, E.; Lira-Saldívar, R.H.; Barriga-Castro, E.D.; Ruiz-Torres, N.A.; Ramos-Cortez, E.; Vázquez-Alvarado, R.; Niño-Medina, G. Zinc Oxide nanoparticles boosts phenolic compounds and antioxidant activity of Capsicum annuum L. during germination. Agronomy 2018, 8, 215. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and Carotenoids: Pigments of Photosynthetic Biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventos, R.M. Analysis of total phenols and Other oxidation substrates and antioxidants by means of Folin-Ciocalteu Reagent. Sci. Hortic. 1999, 213, 281–286. [Google Scholar] [CrossRef]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT-Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in Vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quiantities of protein utilizing the principle of protein-dye binding. Crop. J. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- GOnsa, H.; Saari, N.B.; Selamat, J.; Bakar, J. Purification and characterization of membrane-bound peroxidases from Metroxylon sagu. Food Chem. 2004, 85, 365–376. [Google Scholar] [CrossRef]
- Oktay, M.; Kufreviolu, I.; Kocacaliskan, I.; Şakirolu, H. Polyphenoloxidase from Amasya Apple. J. Food Sci. 1995, 60, 494–496. [Google Scholar] [CrossRef]
- Thunugunta, T.; Reddy, A.C.; Seetharamaiah, S.K.; Hunashikatti, L.R.; Chandrappa, S.G.; Kalathil, N.C.; Reddy, L.R.D.C. Impact of Zinc oxide nanoparticles on eggplant (S. melongena): Studies on growth and the accumulation of nanoparticles. IET Nanobiotechnol. 2018, 12, 706–713. [Google Scholar] [CrossRef]
- Singh, A.; Singh, N.B.; Afzal, S.; Singh, T.; Hussain, I. Zinc oxide nanoparticles: A review of their biological synthesis, antimicrobial activity, uptake, translocation and biotransformation in plants. J. Mater. Sci. 2017, 53, 185–201. [Google Scholar] [CrossRef]
- Itroutwar, P.D.; Kasivelu, G.; Raguraman, V.; Malaichamy, K.; Sevathapandian, S.K. Effects of biogenic zinc oxide nanoparticles on seed germination and seedling vigor of maize (Zea mays). Biocatal. Agric. Biotechnol. 2020, 29, 101778. [Google Scholar] [CrossRef]
- Rai-Kalal, P.; Jajoo, A. Priming with zinc oxide nanoparticles improve germination and photosynthetic performance in wheat. Plant Physiol. Biochem. 2021, 160, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Joo, J.C.; Kim, D.; Lee, J.; Kim, J.Y. Estimation of the Seedling Vigor Index of Sunflowers Treated with Various Heavy Metals. J. Bioremediation Biodegrad. 2016, 7. [Google Scholar] [CrossRef]
- Rajjou, L.; Duval, M.; Gallardo, K.; Catusse, J.; Bally, J.; Job, C.; Job, D. Seed germination and vigor. Annu. Rev. Plant Biol. 2012, 63, 507–533. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Torres, N.; Flores-Naveda, A.; Barriga-Castro, E.D.; Camposeco-Montejo, N.; Ramírez-Barrón, S.; Borrego-Escalante, F.; Niño-Medina, G.; Hernández-Juárez, A.; Garza-Alonso, C.; Rodríguez-Salinas, P.; et al. Zinc oxide nanoparticles and zinc sulfate impact physiological parameters and boosts lipid peroxidation in soil grown coriander plants (Coriandrum sativum). Molecules 2021, 26, 1998. [Google Scholar] [CrossRef] [PubMed]
- Song, U.; Lee, S. Phytotoxicity and accumulation of zinc oxide nanoparticles on the aquatic plants Hydrilla verticillata and Phragmites Australis: Leaf-type-dependent responses. Environ. Sci. Pollut. Res. 2016, 23, 8539–8545. [Google Scholar] [CrossRef]
- Zhao, J.; He, Y.; Huang, S.; Wang, Z. Advances in the Identification of Quantitative Trait Loci and Genes Involved in Seed Vigor in Rice. Front. Plant Sci. 2021, 12, 659307. [Google Scholar] [CrossRef]
- Yang, Z.; Chen, J.; Dou, R.; Gao, X.; Mao, C.; Wang, L. Assessment of the phytotoxicity of metal oxide nanoparticles on two crop plants, maize (Zea mays L.) and rice (Oryza sativa L.). Int. J. Environ. Res. Public Health 2015, 12, 15100–15109. [Google Scholar] [CrossRef]
- Landi, M.; Zivcak, M.; Sytar, O.; Brestic, M.; Allakhverdiev, S.I. Plasticity of photosynthetic processes and the accumulation of secondary metabolites in plants in response to monochromatic light environments: A review. Biochim. Biophys. Acta Bioenerg. 2020, 1861, 148131. [Google Scholar] [CrossRef]
- Puccinelli, M.; Rosellini, I.; Malorgio, F.; Pardossi, A.; Pezzarossa, B. Hydroponic production of selenium-enriched baby leaves of Swiss chard and its wild ancestor sea beet. Horticulturae 2023, 9, 909. [Google Scholar] [CrossRef]
- Rajput, V.D.; Minkina, T.; Fedorenko, A.; Chernikova, N.; Hassan, T.; Mandzhieva, S.; Sushkova, S.; Lysenko, V.; Soldatov, M.A.; Burachevskaya, M. Effects of zinc oxide nanoparticles on physiological and anatomical indices in spring barley tissues. Nanomaterials 2021, 11, 1722. [Google Scholar] [CrossRef] [PubMed]
- Sathiyabama, M. Biopolymeric nanoparticles as a nanocide for crop protection. In Nanoscience for Sustainable Agriculture, 1st ed.; Pudake, R.N., Chauhan, N., Kole, C., Eds.; Springer Nature AG: Cham, Switzerland, 2019; pp. 139–152. [Google Scholar] [CrossRef]
- Alsuwayyid, A.A.; Alslimah, A.S.; Perveen, K.; Bukhari, N.A.; Al-Humaid, L.A. Effect of zinc oxide nanoparticles on Triticum aestivum L. and bioaccumulation assessment using ICP-MS and SEM analysis. J. King Saud Univ. Sci. 2022, 34, 101944. [Google Scholar] [CrossRef]
- Al Awadh, A.A.; Shet, A.R.; Patil, L.R.; Shaikh, I.A.; Alshahrani, M.M.; Nadaf, R.; Mahnashi, M.H.; Desai, S.V.; Muddapur, U.M.; Achappa, S.; et al. Sustainable Synthesis and Characterization of Zinc Oxide Nanoparticles Using Raphanus sativus Extract and Its Biomedical Applications. Crystals 2022, 12, 1142. [Google Scholar] [CrossRef]
- Balážová, Ľ.; Babula, P.; Baláž, M.; Bačkorová, M.; Bujňáková, Z.; Briančin, J.; Kurmanbayeva, A.; Sagi, M. Zinc oxide nanoparticles phytotoxicity on halophyte from genus Salicornia. Plant Physiol. Biochem. 2018, 130, 30–42. [Google Scholar] [CrossRef]
- Zoufan, P.; Baroonian, M.; Zargar, B. ZnO nanoparticles-induced oxidative stress in Chenopodium murale L, Zn uptake, and accumulation under hydroponic culture. Environ. Sci. Pollut. Res. 2020, 27, 11066–11078. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Guob, D.; Yina, X.; Dingc, S.; Shend, M.; Zhangd, R.; Wangc, Y.; Xuc, R. Zinc oxide nanoparticles induce human multiple myeloma cell death via reactive oxygen species and Cyt-C/Apaf-1/Caspase-9/Caspase-3 signaling pathway in vitro. Biomed. Pharmacother. 2020, 122, 109712. [Google Scholar] [CrossRef]
- Wang, X.; Li, Q.Q.; Pei, Z.M.; Wang, S.C. Zinc oxide nanoparticles affect biomass accumulation and photosynthesis in Arabidopsis. Front. Plant Sci. 2016, 6, 1243. [Google Scholar] [CrossRef]
- Wang, X.P.; Li, Q.Q.; Pei, Z.M.; Wang, S.C. Effects of zinc oxide nanoparticles on the growth, photosynthetic traits, and antioxidative enzymes in tomato plants. Biol. Plant. 2018, 62, 801–808. [Google Scholar] [CrossRef]
- Priyanka, N.; Venkatachalam, P. Biofabricated zinc oxide nanoparticles coated with phycomolecules as novel micronutrient catalysts for stimulating plant growth of cotton. Adv. Nat. Sci. Nanosci. Nanotechnol. 2016, 7, 045018. [Google Scholar] [CrossRef]
- Mogazy, A.M.; Hanafy, R.S. Foliar Spray of Biosynthesized Zinc Oxide Nanoparticles Alleviate Salinity Stress Effect on Vicia faba Plants. J. Soil Sci. Plant Nutr. 2022, 22, 2647–2662. [Google Scholar] [CrossRef]
- Chen, J.; Liua, X.; Wang, C.; Yin, S.; Lia, X.; Hua, W.; Simona, M.; Shena, Z.; Xiaod, Q.; Chue, C. Nitric oxide ameliorates zinc oxide nanoparticles-induced phytotoxicity in rice seedlings. J. Hazard. Mater. 2015, 297, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Numan, M.; Khan, A.L.; Lee, I.-J.; Imran, M.; Asaf, S.; Al-Harrasi, A. Melatonin: Awakening the defense mechanisms during plant oxidative stress. Plants 2020, 9, 407. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Bhardwaj, R.; Thukral, A.K.; Al-Huqail, A.A.; Siddiqui, M.H.; Ahmad, P. Oxidative stress mitigation and initiation of antioxidant and osmoprotectant responses mediated by ascorbic acid in Brassica juncea L. subjected to copper (II) stress. Ecotoxicol. Environ. Saf. 2019, 182, 109436. [Google Scholar] [CrossRef] [PubMed]
- Anjali; Kumar, S.; Korra, T.; Thakur, R.; Arutselvan, R.; Kashyap, A.S.; Nehela, Y.; Chaplygin, V.; Minkina, T.; Keswani, C. Role of plant secondary metabolites in defence and transcriptional regulation in response to biotic stress. Plant Stress 2023, 8, 100154. [Google Scholar] [CrossRef]
- Dimkpa, C.O.; Singh, U.; Bindraban, P.S.; Elmer, W.H.; Gardea-Torresdey, J.L.; White, J.C. Zinc oxide nanoparticles alleviate drought-induced alterations in sorghum performance, nutrient acquisition, and grain fortification. Sci. Total Environ. 2019, 688, 926–934. [Google Scholar] [CrossRef] [PubMed]
- Páramo, L.; Feregrino-Pérez, A.A.; Vega-González, M.; Escobar-Alarcón, L.; Esquivel, K. Medicago sativa L. Plant Response against Possible Eustressors (Fe, Ag, Cu)-TiO2: Evaluation of Physiological Parameters, Total Phenol Content, and Flavonoid Quantification. Plants 2023, 12, 659. [Google Scholar] [CrossRef] [PubMed]
- Salih, A.M.; Al, F.; Khan, S.; Tarroum, M.; Nadeem, M.; Shaikhaldein, H.O.; Gaafar, A.R.Z.; Alfarraj, N.S. Biosynthesis of zinc oxide nanoparticles using Phoenix dactylifera and their effect on biomass and phytochemical compounds in Juniperus procera. Sci. Rep. 2021, 11, 19136. [Google Scholar] [CrossRef]
- Awan, S.; Shahzadi, K.; Javad, S.; Tariq, A.; Ahmad, A.; Ilyas, S. A preliminary study of influence of zinc oxide nanoparticles on growth parameters of Brassica oleracea var italic. J. Saudi Soc. Agric. Sci. 2020, 20, 18–24. [Google Scholar] [CrossRef]
- Iziy, E.; Majd, A.; Vaezi-Kakhki, M.R.; Nejadsattari, T.; Noureini, S.K. Effects of zinc oxide nanoparticles on enzymatic and nonenzymatic antioxidant content, germination, and biochemical and ultrastructural cell characteristics of Portulaca oleracea L. Acta Soc. Bot. Pol. 2019, 88, 1–14. [Google Scholar] [CrossRef]
- Gatoo, M.A.; Naseem, S.; Arfat, M.Y.; Dar, A.M.; Qasim, K.; Zubair, S. Physicochemical properties of nanomaterials: Implication in associated toxic manifestations. Biomed Res. Int. 2014, 2014, 498420. [Google Scholar] [CrossRef] [PubMed]
- Comandella, D.; Gottardo, S.; Rio-Echevarria, I.M.; Rauscher, H. Quality of physicochemical data on nanomaterials: An assessment of data completeness and variability. Nanoscale 2020, 12, 4695–4708. [Google Scholar] [CrossRef] [PubMed]
- Agati, G.; Brunetti, C.; Fini, A.; Gori, A.; Guidi, L.; Landi, M.; Sebastiani, F.; Tattini, M. Are flavonoids effective antioxidants in plants? Twenty years of our investigation. Antioxidants 2020, 9, 1098. [Google Scholar] [CrossRef] [PubMed]
- Javed, R.; Usman, M.; Yücesan, B.; Zia, M.; Gürel, E. Effect of zinc oxide (ZnO) nanoparticles on physiology and steviol glycosides production in micropropagated shoots of Stevia rebaudiana Bertoni. Plant Physiol. Biochem. 2017, 110, 94–99. [Google Scholar] [CrossRef] [PubMed]
- García-López, J.I.; Niño-Medina, G.; Olivares-Sáenz, E.; Lira-Saldivar, R.H.; Barriga-Castro, E.D.; Vázquez-Alvarado, R.; Rodríguez-Salinas, P.A.; Zavala-García, F. Foliar application of zinc oxide nanoparticles and zinc sulfate boosts the content of bioactive compounds in habanero peppers. Plants 2019, 8, 254. [Google Scholar] [CrossRef] [PubMed]
- Alhujaily, M.; Albukhaty, S.; Yusuf, M.; Mohammed, M.K.A.; Sulaiman, G.M.; Al-Karagoly, H.; Alyamani, A.A.; Albaqami, J.; AlMalki, F.A. Recent Advances in Plant-Mediated Zinc Oxide Nanoparticles with Their Significant Biomedical Properties. Bioengineering 2022, 9, 541. [Google Scholar] [CrossRef] [PubMed]
- Ushahra, J.; Bhati-Kushwaha, H.; Malik, C.P. Biogenic nanoparticle-mediated augmentation of seed germination, growth, and antioxidant level of Eruca sativa Mill. varieties. Appl. Biochem. Biotechnol. 2014, 174, 729–738. [Google Scholar] [CrossRef] [PubMed]
- Çekiç, F.Ö.; Ekinci, S.; İnal, M.; Ünal, D. Silver nanoparticles induced genotoxicity and oxidative stress in tomato plants. Turk. J. Biol. 2017, 41, 700–707. [Google Scholar] [CrossRef]
- Elsheery, N.I.; Helaly, M.N.; El-Hoseiny, H.M.; Alam-Eldein, S.M. Zinc oxide and silicone nanoparticles to improve the resistance mechanism and annual productivity of salt-stressed mango trees. Agronomy 2020, 10, 558. [Google Scholar] [CrossRef]
- Taie, H.A.A.; El-Yazal, M.A.S.; Ahmed, S.M.A.; Rady, M.M. Polyamines modulate growth, antioxidant activity, and genomic DNA in heavy metal–stressed wheat plant. Environ. Sci. Pollut. Res. 2019, 26, 22338–22350. [Google Scholar] [CrossRef]
- D’Alonzo, D.; De Fenza, M.; Pavone, V.; Lombardi, A.; Nastri, F. Selective Oxidation of Halophenols Catalyzed by an Artificial Miniaturized Peroxidase. Int. J. Mol. Sci. 2023, 24, 8058. [Google Scholar] [CrossRef]
- Pejam, F.; Ardebili, Z.O.; Ladan-Moghadam, A.; Danaee, E. Zinc oxide nanoparticles mediated substantial physiological and molecular changes in tomato. PLoS ONE 2021, 16, e0248778. [Google Scholar] [CrossRef] [PubMed]
- El-Zohri, M.; Al-Wadaani, N.A.; Bafeel, S.O. Foliar Sprayed Green Zinc Oxide Nanoparticles Mitigate. Plants 2021, 10, 2400. [Google Scholar] [CrossRef]
- Latef, A.A.H.A.; Alhmad, M.F.A.; Abdelfattah, K.E. The Possible Roles of Priming with ZnO Nanoparticles in Mitigation of Salinity Stress in Lupine (Lupinus termis) Plants. J. Plant Growth Regul. 2017, 36, 60–70. [Google Scholar] [CrossRef]
- Hafizi, Z.; Nasr, N. The Effect of Zinc Oxide Nanoparticles on Safflower Plant Growth and Physiology. Eng. Technol. Appl. Sci. Res. 2018, 8, 2508–2513. [Google Scholar] [CrossRef]
- García-Gómez, C.; Obrador, A.; González, D.; Babín, M.; Fernández, M.D. Comparative effect of ZnO NPs, ZnO bulk and ZnSO4 in the antioxidant defences of two plant species growing in two agricultural soils under greenhouse conditions. Sci. Total Environ. 2017, 589, 11–24. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentration of ZnONP | Fresh Weight of Plumule | Fresh Weight of Root | Lenght of the Plumule | Length of the Roots | ||||
|---|---|---|---|---|---|---|---|---|
| (mg L−1) | (mg) | (mg) | (mm) | (mm) | ||||
| 100B | 11.24 ± 0.29 | b | 1.831 ± 0.046 | bc | 22.60 ± 0.15 | b | 11.61 ± 0.46 | d |
| 200B | 13.41 ± 0.51 | a | 1.651 ± 0.048 | c | 25.71 ± 0.47 | a | 22.71 ± 0.30 | a |
| 300B | 11.77 ± 0.15 | b | 2.171 ± 0.082 | a | 20.60 ± 1.10 | c | 19.22 ± 0.20 | b |
| 400B | 9.54 ± 0.33 | c | 1.912 ± 0.024 | b | 19.72 ± 1.10 | c | 10.38 ± 0.20 | de |
| control | 9.85 ± 0.41 | c | 1.840 ± 0.067 | bc | 19.37 ± 0.17 | c | 9.52 ± 0.43 | e |
| 100C | 11.20 ± 0.40 | b | 1.982 ± 0.064 | b | 20.38 ± 0.90 | d | 16.79 ± 1.05 | c |
| 200C | 11.33 ± 0.42 | b | 1.294± 0.045 | d | 21.20 ± 1.54 | bc | 15.89 ± 0.88 | c |
| 300C | 9.20 ± 0.13 | c | 1.512 ± 0.055 | c | 24.22 ± 1.97 | ab | 12.74 ± 0.63 | d |
| 400C | 9.44 ± 0.30 | c | 1.296 ± 0.058 | d | 14.87 ± 0.38 | e | 9.043 ± 0.25 | e |
| Concentration of ZnONP | Fresh Weight of Plumule | Fresh Weight of Roots | Length of Plumule | Length of Roots | ||||
|---|---|---|---|---|---|---|---|---|
| (mg L−1) | (mg) | (mg) | (mm) | (mm) | ||||
| 100B | 23.26 ± 0.29 | b | 8.49 ± 0.05 | b | 26.96 ± 0.15 | b | 8.60 ± 0.46 | d |
| 200B | 24.43 ± 0.85 | a | 10.64 ± 0.62 | a | 30.07 ± 0.47 | a | 27.01 ± 1.61 | a |
| 300B | 23.79 ± 0.15 | b | 8.83 ± 0.08 | b | 24.96 ± 1.10 | c | 24.81 ± 1.07 | bc |
| 400B | 11.79 ± 0.76 | e | 7.90 ± 0.02 | c | 8.45 ± 0.98 | e | 9.61 ± 0.50 | d |
| control | 20.10 ± 1.01 | c | 7.83 ± 0.07 | c | 25.06 ± 0.55 | c | 8.20 ± 0.41 | d |
| 100C | 23.22 ± 0.40 | b | 8.64 ± 0.06 | b | 24.74 ± 0.90 | c | 26.83 ± 1.40 | b |
| 200C | 23.35 ± 0.42 | b | 7.95 ± 0.04 | c | 25.56 ± 0.46 | bc | 12.96 ± 0.54 | c |
| 300C | 15.89 ± 0.55 | d | 8.17 ± 0.05 | b | 11.91 ± 0.34 | d | 9.07 ± 1.21 | d |
| 400C | 19.86 ± 0.15 | c | 4.89 ± 0.31 | d | 19.23 ± 0.38 | cd | 6.033 ± 0.25 | e |
| Concentration of ZnONP | Catalase | Peroxidase | Polyphenol Oxidase | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Units of CAT mg−1 of Protein | Units of POD mg−1 of Protein | Units of PPO mg−1 of Protein | ||||||||||
| (mg L−1) | Capsicum annuum | Solanum lycopersicum | Capsicum annuum | Solanum lycopersicum | Capsicum annuum | Solanum lycopersicum | ||||||
| 100B | 6.67 ± 0.66 | a | 7.02 ± 0.93 | b | 10.19 ± 0.41 | cd | 13.06 ± 0.58 | a | 0.26 ± 0.01 | a | 0.24 ± 0.07 | ab |
| 200B | 5.11 ± 0.16 | c | 7.79 ± 0.17 | a | 10.32 ± 0.57 | bc | 11.21 ± 0.68 | b | 0.26 ± 0.01 | a | 0.22 ± 0.03 | b |
| 300B | 6.14 ± 0.31 | b | 5.13 ± 0.37 | d | 10.01 ± 0.80 | cd | 10.79 ± 0.92 | bc | 0.21 ± 0.06 | cd | 0.22 ± 0.02 | b |
| 400B | 4.31 ± 0.29 | d | 5.22 ± 0.32 | d | 11.33 ± 1.12 | a | 12.27 ± 1.00 | ab | 0.22 ± 0.01 | e | 0.25 ± 0.02 | a |
| control | 3.32 ± 0.10 | e | 4.57 ± 0.09 | e | 9.47 ± 0.29 | cd | 10.35 ± 0.30 | c | 0.16 ± 0.01 | e | 0.15 ± 0.01 | d |
| 100C | 4.73 ± 0.28 | d | 5.93 ± 0.32 | c | 8.26 ± 0.33 | d | 8.96 ± 0.46 | d | 0.23 ± 0.02 | bc | 0.23 ± 0.02 | b |
| 200C | 4.94 ± 0.61 | cd | 5.48 ± 0.33 | cd | 8.97 ± 1.15 | d | 9.68 ± 1.13 | d | 0.19 ± 0.07 | d | 0.21 ± 0.04 | c |
| 300C | 5.83 ± 0.82 | b | 6.93 ± 0.96 | b | 10.73 ± 0.45 | b | 11.63 ± 0.51 | b | 0.25 ± 0.06 | a | 0.22 ± 0.02 | b |
| 400C | 4.90 ± 0.43 | c | 5.59 ± 0.15 | c | 10.39 ± 0.69 | bc | 11.38 ± 0.77 | b | 0.21 ± 0.03 | c | 0.21 ± 0.02 | c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Pérez, D.M.; Márquez-Guerrero, S.Y.; Ramírez-Moreno, A.; Rodríguez-Sifuentes, L.; Galindo-Guzmán, M.; Flores-Loyola, E.; Marszalek, J.E. Impact of Biologically and Chemically Synthesized Zinc Oxide Nanoparticles on Seed Germination and Seedlings’ Growth. Horticulturae 2023, 9, 1201. https://doi.org/10.3390/horticulturae9111201
Sánchez-Pérez DM, Márquez-Guerrero SY, Ramírez-Moreno A, Rodríguez-Sifuentes L, Galindo-Guzmán M, Flores-Loyola E, Marszalek JE. Impact of Biologically and Chemically Synthesized Zinc Oxide Nanoparticles on Seed Germination and Seedlings’ Growth. Horticulturae. 2023; 9(11):1201. https://doi.org/10.3390/horticulturae9111201
Chicago/Turabian StyleSánchez-Pérez, Daniela Monserrat, Selenne Yuridia Márquez-Guerrero, Agustina Ramírez-Moreno, Lucio Rodríguez-Sifuentes, Magdalena Galindo-Guzmán, Erika Flores-Loyola, and Jolanta E. Marszalek. 2023. "Impact of Biologically and Chemically Synthesized Zinc Oxide Nanoparticles on Seed Germination and Seedlings’ Growth" Horticulturae 9, no. 11: 1201. https://doi.org/10.3390/horticulturae9111201
APA StyleSánchez-Pérez, D. M., Márquez-Guerrero, S. Y., Ramírez-Moreno, A., Rodríguez-Sifuentes, L., Galindo-Guzmán, M., Flores-Loyola, E., & Marszalek, J. E. (2023). Impact of Biologically and Chemically Synthesized Zinc Oxide Nanoparticles on Seed Germination and Seedlings’ Growth. Horticulturae, 9(11), 1201. https://doi.org/10.3390/horticulturae9111201