
Optimization of In Vitro Regeneration Protocol of Tomato cv. MT1 for Genetic Transformation

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Plant Material
2.2. Culture Media for Shoot Induction
2.3. Culture and Subculture
2.4. Shoot Elongation, Rooting, and Acclimatization
2.5. Experimental Design, Data Collection, and Statistical Analysis
3. Results and Discussion
3.1. Regeneration of Shoot
3.1.1. Percentage of Explants Producing Direct Shoots
3.1.2. Percentage of Explants Producing Roots
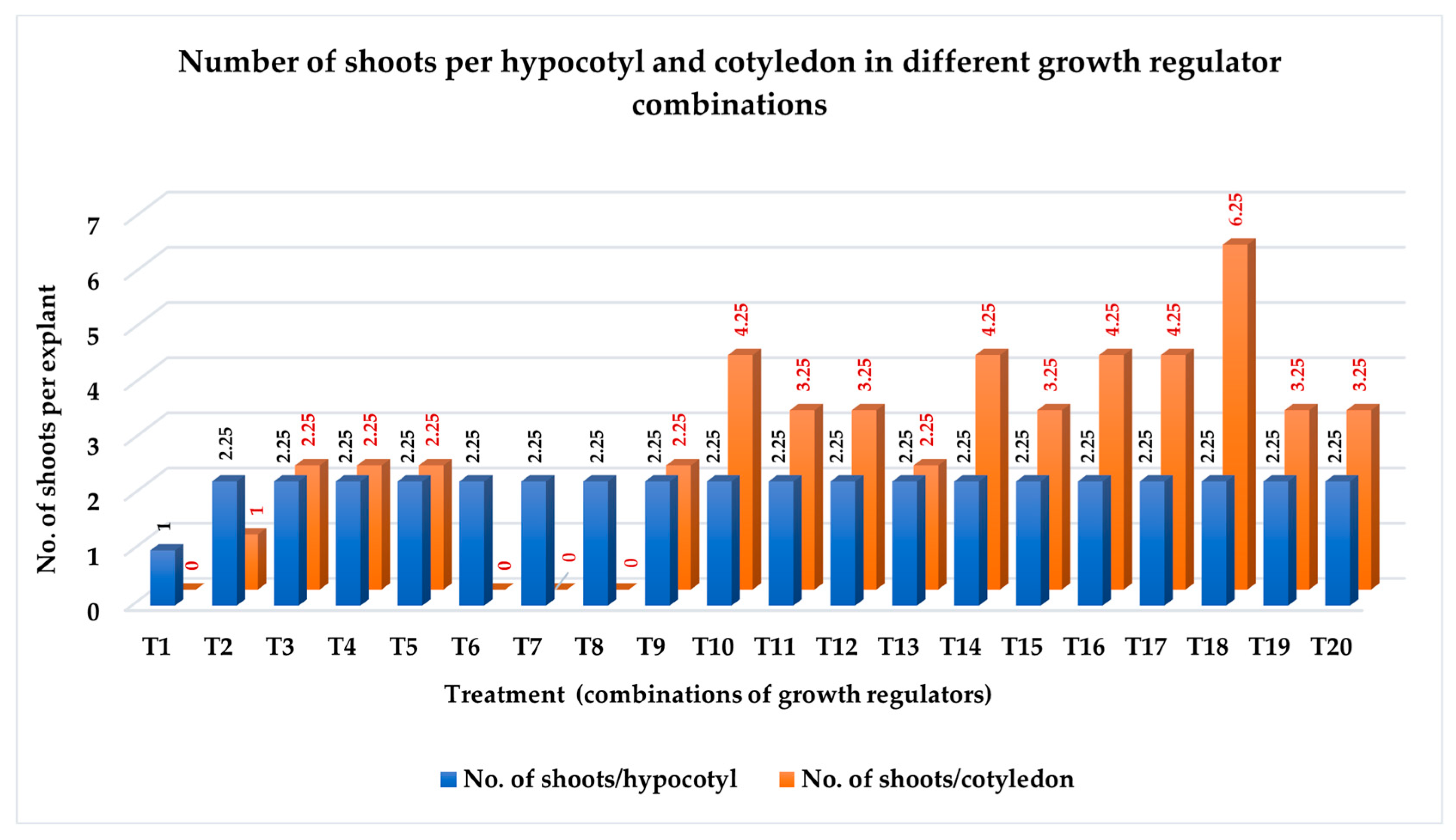
3.1.3. Number of Shoots per Explant
3.1.4. Type of Induced Organ
3.1.5. Shoot Elongation
3.2. Root Initiation
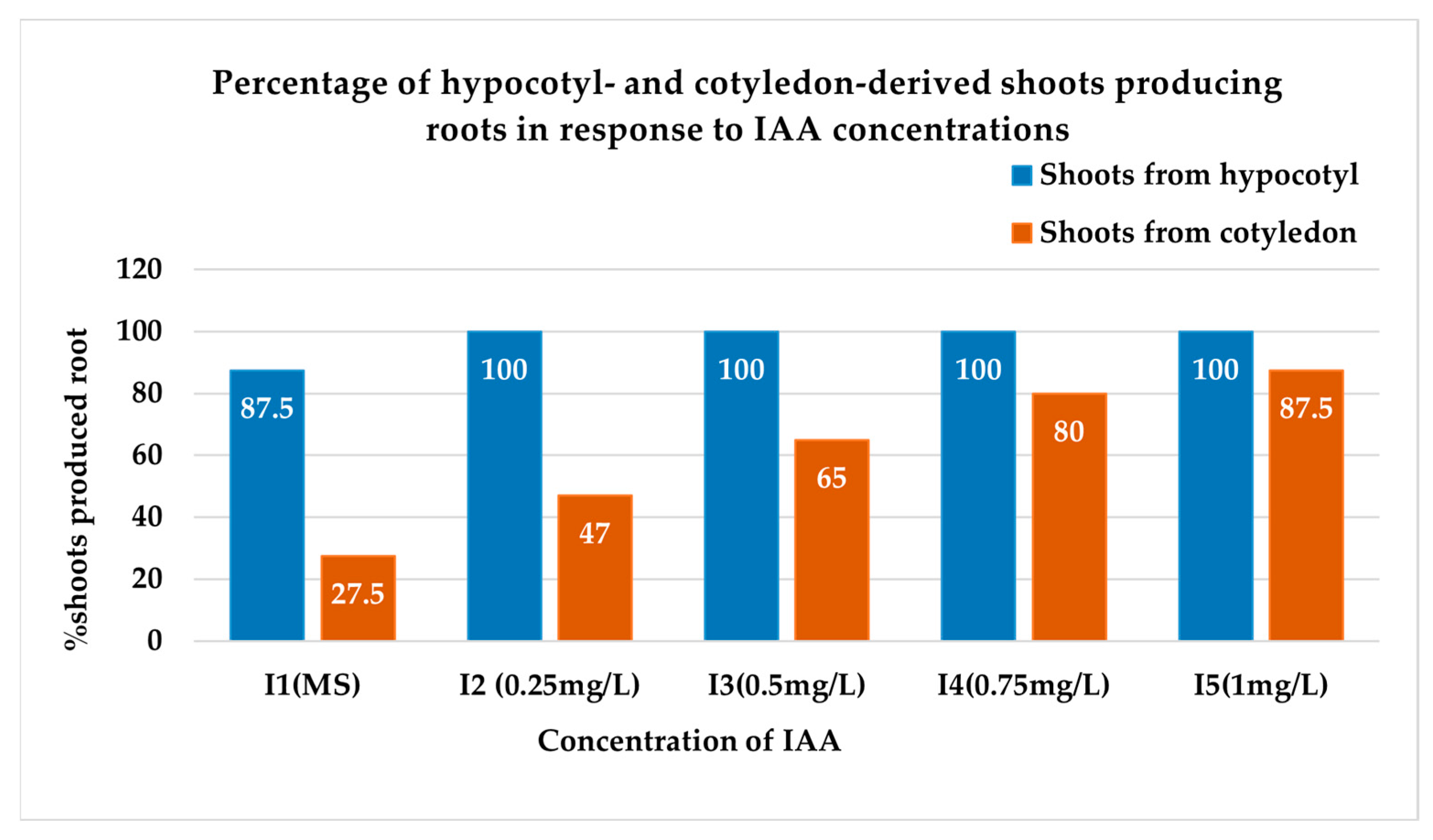
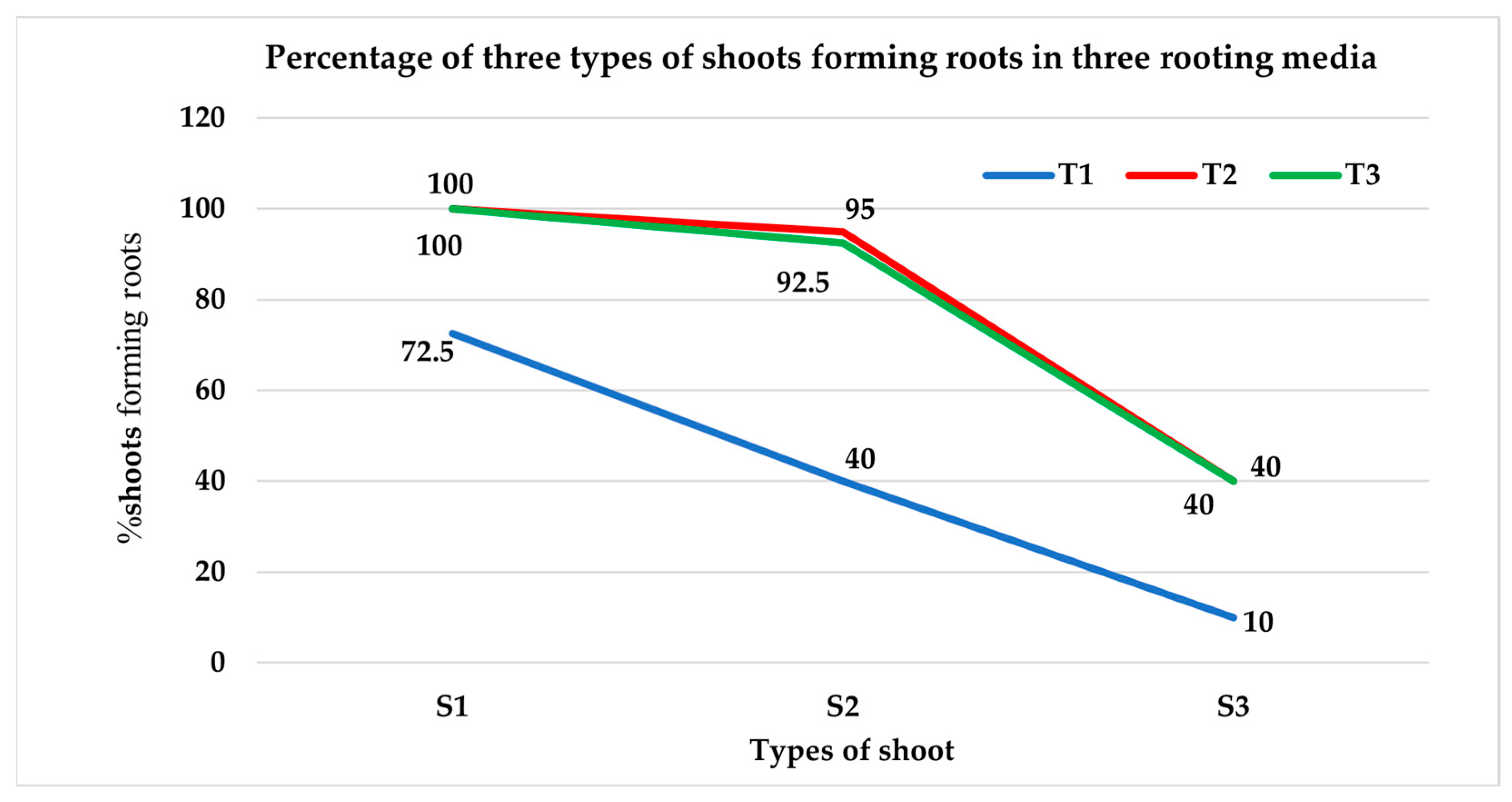
3.2.1. Percentage of Shoots Producing Roots
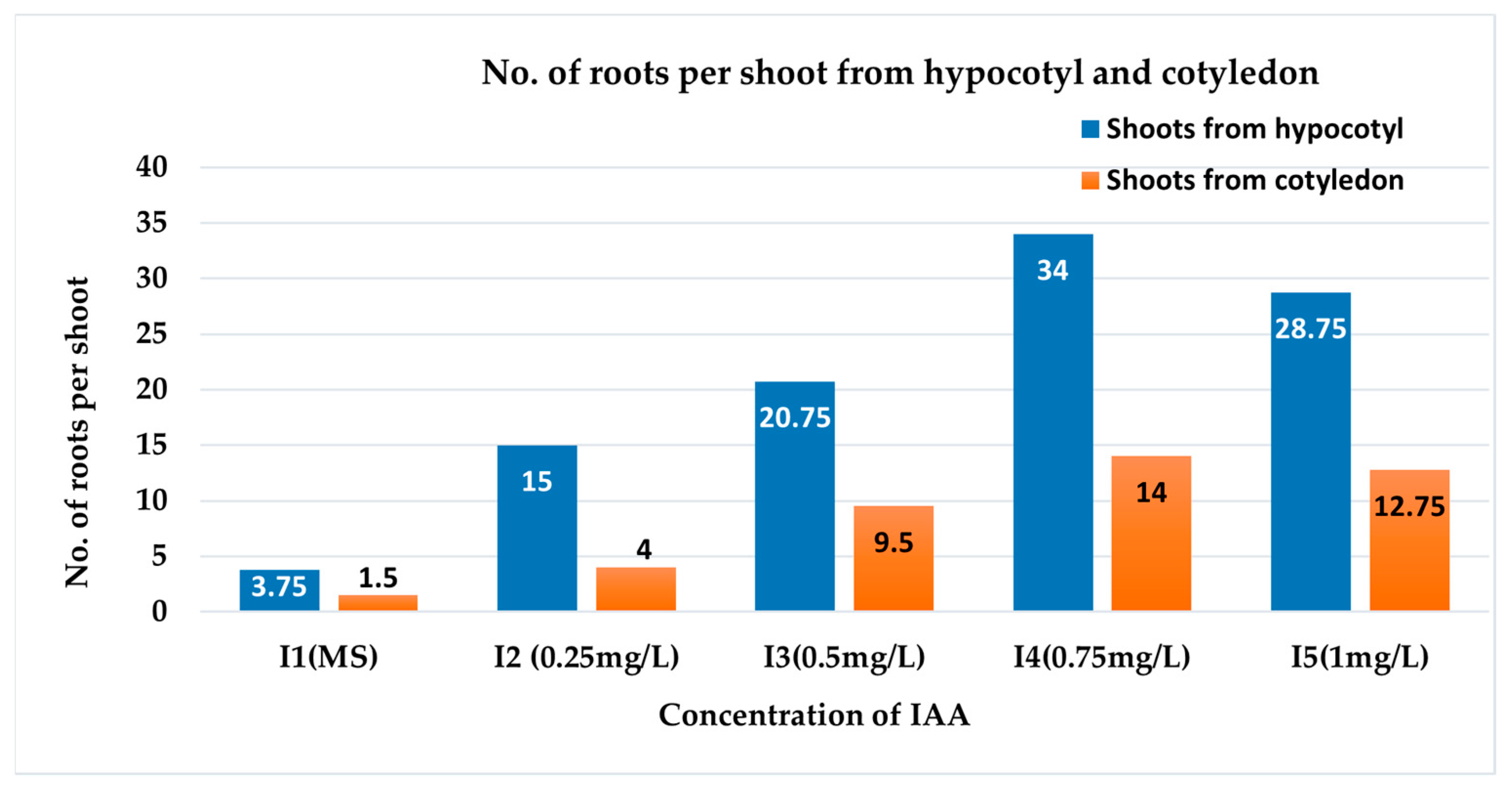
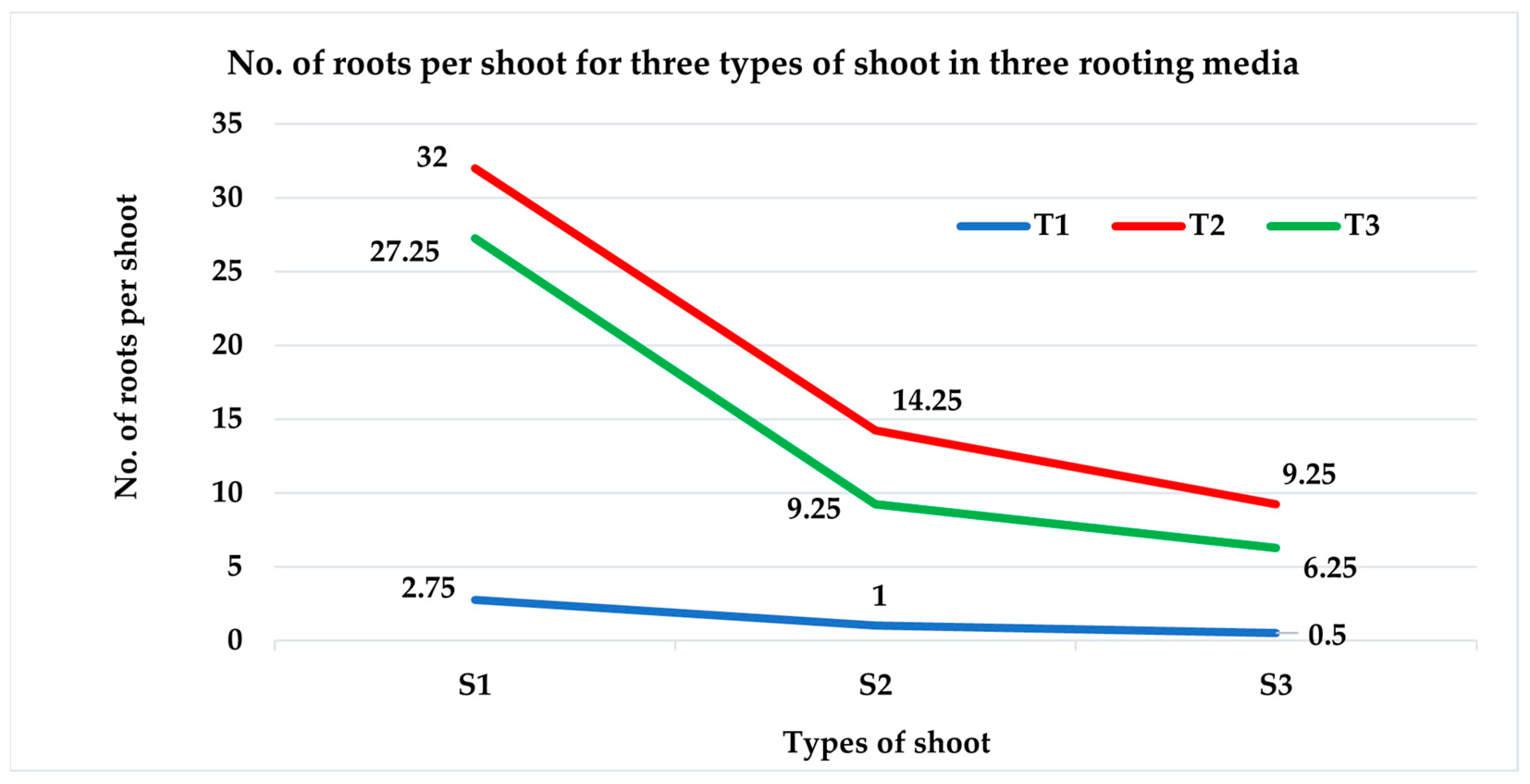
3.2.2. Number of Roots per Shoot
3.2.3. Root Length and Diameter (cm)
3.2.4. Survival Rates (%) of Rooted Shoots in Soil
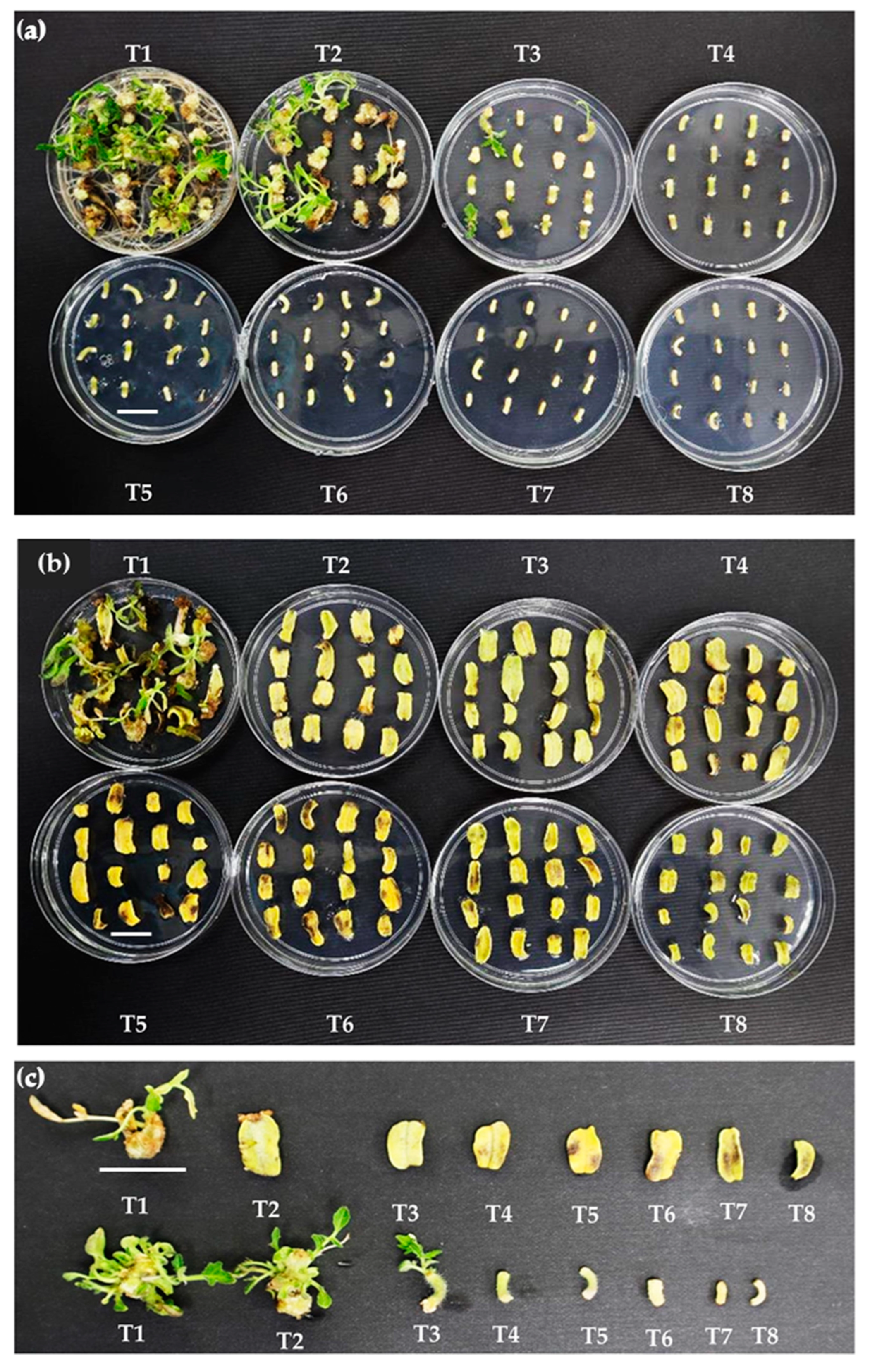
3.3. Minimal Inhibitory Concentration of Kanamycin on Hypocotyl and Cotyledon Explants
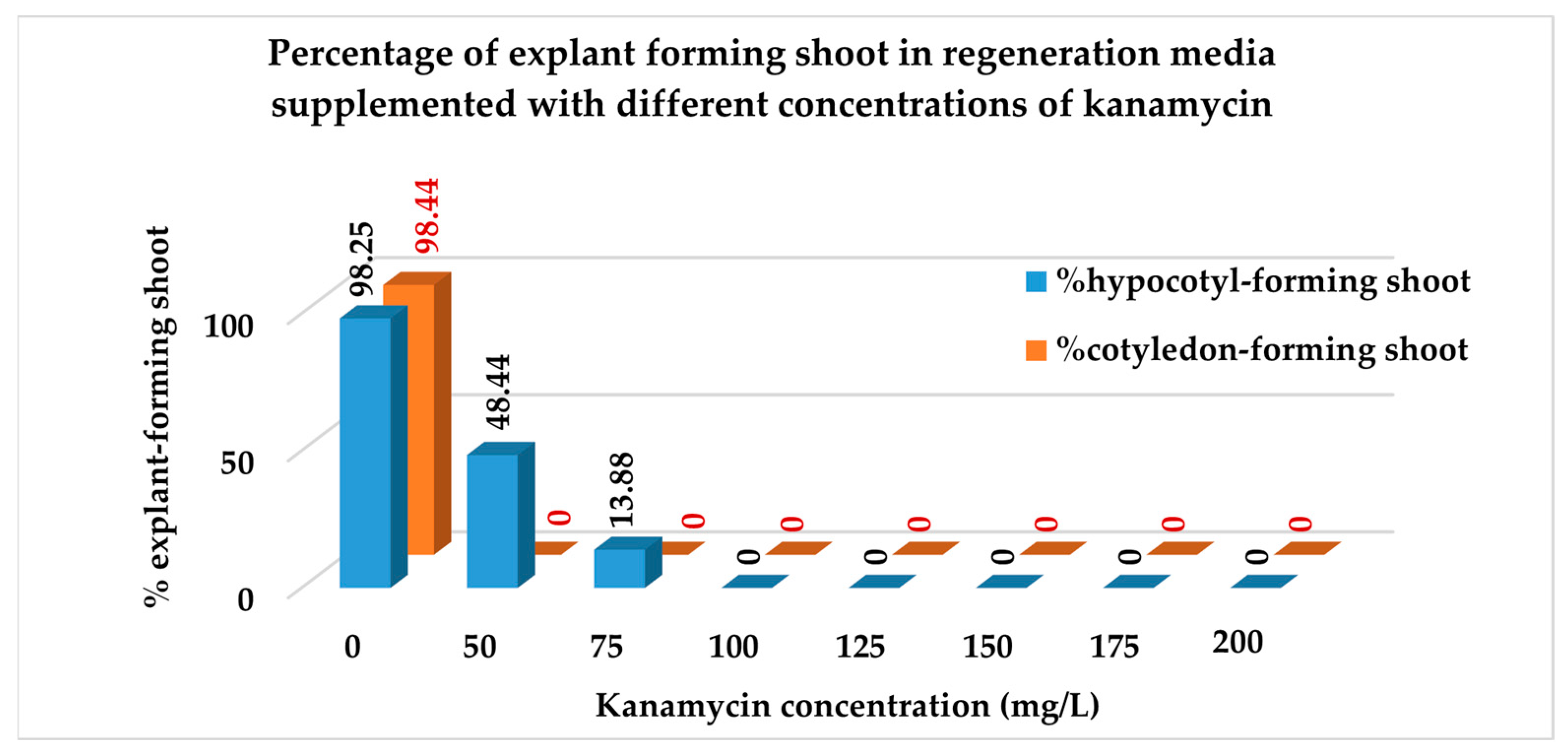
3.3.1. Percentage of Explants Forming Shoots
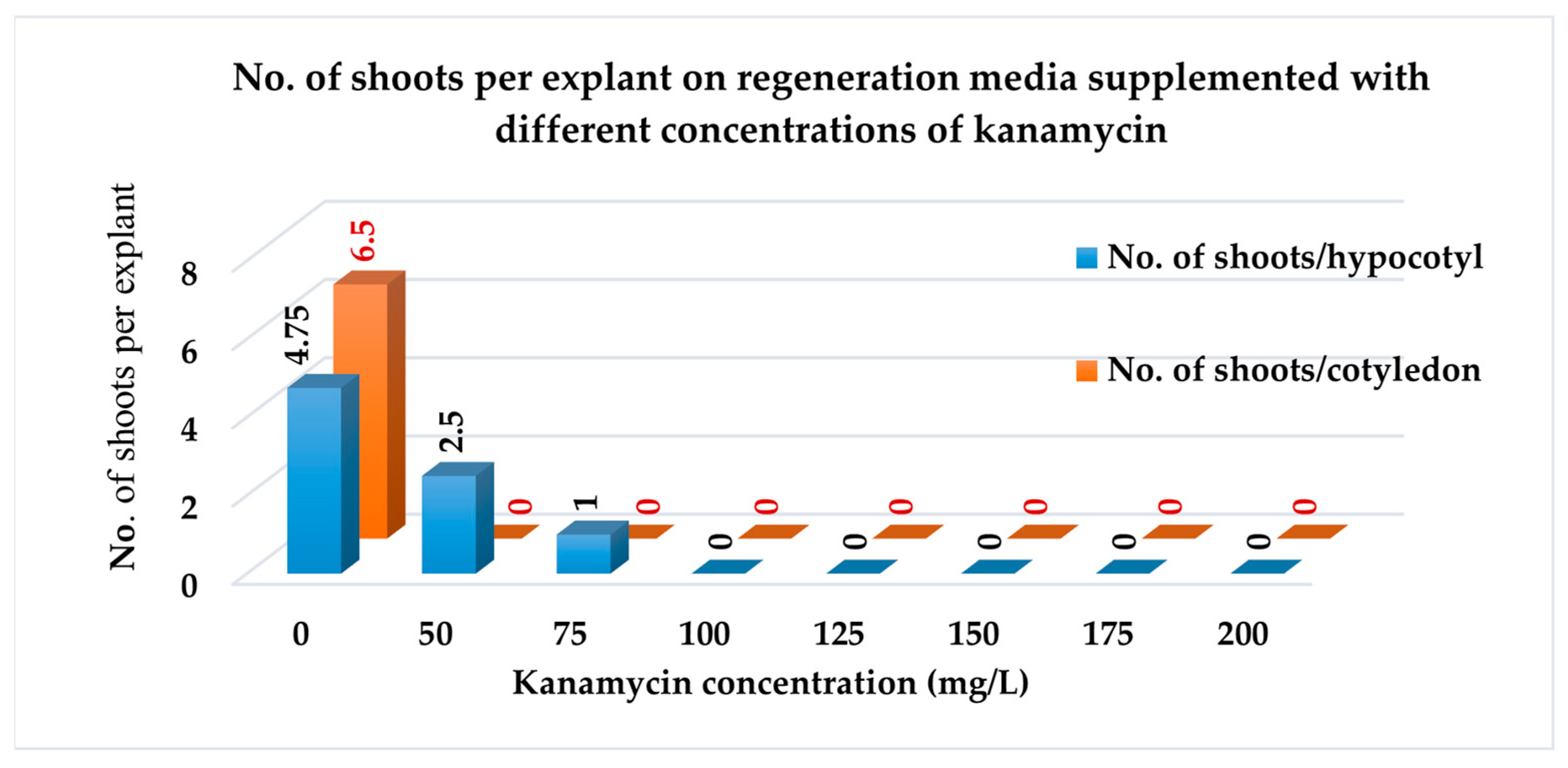
3.3.2. Number of Shoots per Explant
3.3.3. Rooting and Acclimatization
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT. Crops and Livestock Products. 2022. Available online: https://www.fao.org/faostat/en/#data/QCL/visualize (accessed on 14 May 2023).
- Francois-Xavier, B. Worldwide (Total Fresh) Tomato Production Exceeds 187 Million Tonnes in 2020, in Tomato News Online Conference. 2022. Available online: https://www.tomatonews.com/en/worldwide-total-fresh-tomato-production-exceeds-187-million-tonnes-in-2020_2_1565.html (accessed on 14 May 2023).
- Anwar, R.; Fatima, T.; Mattoo, A.K. Tomatoes: A Model Crop of Solanaceous Plants; Oxford University Press: Oxford, UK, 2019; pp. 1–50. [Google Scholar]
- Liu, W.; Liu, K.; Chen, D.; Zhang, Z.; Li, B.; El-Mogy, M.M.; Tian, S.; Chen, T. Solanum lycopersicum, a Model Plant for the Studies in Developmental Biology, Stress Biology and Food Science. Foods 2022, 11, 2402. [Google Scholar] [CrossRef] [PubMed]
- Van Eck, J.; Keen, P.; Tjahjadi, M. Agrobacterium tumefaciens-Mediated Transformation of Tomato. Methods Mol. Biol. 2019, 1864, 225–234. [Google Scholar] [PubMed]
- Park, S.H.; Morris, J.L.; Park, J.E.; Hirschi, K.D.; Smith, R.H. Efficient and genotype-independent Agrobacterium-mediated tomato transformation. J. Plant. Physiol. 2003, 160, 1253–1257. [Google Scholar] [CrossRef]
- Qiu, D.; Diretto, G.; Tavarza, R.; Giuliano, G. Improved protocol for Agrobacterium-mediated transformation of tomato and production of transgenic plants containing carotenoid biosynthetic gene CsZCD. Sci. Hortic. 2007, 112, 172–175. [Google Scholar] [CrossRef]
- El-Shafey, N.; Hassan, N.; Khodary, S.; Badr, A. Differential In vitro Direct Regeneration of Tomato Genotypes on Various Combinations of Growth Regulators. Biotechnology 2017, 16, 155–164. [Google Scholar] [CrossRef]
- Praveen, M.; Nanna, R.S. Effect of genotype, explant source and medium on in vitro regeneration of tomato. Int. J. Genet. Mol. Biol. 2011, 3, 45–50. [Google Scholar]
- Bhatia, P.; Ashwath, N.; Senaratna, T.; Midmore, D. Tissue Culture Studies of Tomato (Lycopersicon esculentum). Plant Cell Tissue Organ Cult. 2004, 78, 1–21. [Google Scholar] [CrossRef]
- Rahim, H.; Wahab, M.A.M.A.; Amin, M.Z.M.; Harun, A.; Haimid, M.T. Technological adoption evaluation of agricultural and food sectors towards modern agriculture: Tomato (Penilaian penggunaan teknologi bagi sektor pertanian dan makanan ke arah pertanian moden: Tomato. Econ. Technol. Manag. Rev. 2017, 12, 41–53. [Google Scholar]
- Kamaladini, H.; Siti Nor, A.A.; Maheran, A.A. In vitro shoot regeneration for cotyledonary leaf explant of tomato variety MT1. In Proceedings of the Biodiversity and Biotechnology Symposium 2008, Kuching, Sarawak, Malaysia, 19–21 November 2008. [Google Scholar]
- Behboodian, B.; Mohd Ali, Z.; Ismail, I.; Zainal, Z. Postharvest analysis of lowland transgenic tomato fruits harboring hpRNAi-ACO1 construct. Sci. World J. 2012, 2012, 439870. [Google Scholar] [CrossRef]
- Raj, S.K.; Singh, R.; Pandey, S.; Singh, B. Agrobacterium-mediated tomato transformation and regeneration of transgenic lines expressing Tomato leaf curl virus coat protein gene for resistance against TLCV infection. Curr. Sci. 2005, 88, 1674–1679. [Google Scholar]
- Plancarte-De la Torre, M.M.; Nunez-Palenius, H.G.; Gomez-Lim, M.A. Tomato Transformation with Genes Involved in Plant Immunity to Confer Broad Resistance Against Bacteria. Rev. Fitotec. Mex. 2016, 39, 349–358. [Google Scholar]
- Gay, P.B.; Gillespie, S.H. Antibiotic resistance markers in genetically modified plants: A risk to human health? Lancet Infect. Dis. 2005, 5, 637–646. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Ryu, D.; Houtkooper, R.H.; Auwerx, J. Antibiotic use and abuse: A threat to mitochondria and chloroplasts with impact on research, health, and environment. Bioessays 2015, 37, 1045–1053. [Google Scholar] [CrossRef] [PubMed]
- Farooq, N.; Nawaz, M.A.; Mukhtar, Z.; Ali, I.; Hundleby, P.; Ahmad, N. Investigating the In Vitro Regeneration Potential of Commercial Cultivars of Brassica. Plants 2019, 8, 558. [Google Scholar] [CrossRef]
- Joersbo, M.; Okkels, F.T. A novel principle for selection of transgenic plant cells: Positive selection. Plant Cell Rep. 1996, 16, 219–221. [Google Scholar] [CrossRef] [PubMed]
- Berry, M.; Gurung, A.; Easty, D.L. Toxicity of antibiotics and antifungals on cultured human corneal cells: Effect of mixing, exposure and concentration. Eye 1995, 9 Pt 1, 110–115. [Google Scholar] [CrossRef]
- Gerszberg, A.; Grzegorczyk-Karolak, I. Influence of Selected Antibiotics on the Tomato Regeneration in in vitro cultures. Not. Bot. Horti Agrobot. Cluj-Napoca 2019, 47, 558–564. [Google Scholar] [CrossRef]
- Andrews, J.M. Determination of minimum inhibitory concentrations. J. Antimicrob. Chemother. 2001, 48 (Suppl. S1), 5–16. [Google Scholar] [CrossRef]
- Saed Taha, R.; Ismail, I.; Zainal, Z.; Abdullah, S.N. The stearoyl-acyl-carrier-protein desaturase promoter (Des) from oil palm confers fruit-specific GUS expression in transgenic tomato. J. Plant. Physiol. 2012, 169, 1290–1300. [Google Scholar] [CrossRef]
- Azzeme, A.M.; Abdullah, S.N.A.; Aziz, M.A.; Wahab, P.E.M. Oil palm drought inducible DREB1 induced expression of DRE/CRT- and non-DRE/CRT-containing genes in lowland transgenic tomato under cold and PEG treatments. Plant. Physiol. Biochem. 2017, 112, 129–151. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F.K. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Gamborg, O.L.; Miller, R.A.; Ojima, K. Nutrient requirements of suspension cultures of soybean root cells. Exp. Cell Res. 1968, 50, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Mendívil, A.; López, J.; Germán Báez, L.; Lopez-meyer, M.; Hernández-Verdugo, S.; López-Valenzuela, J.; Reyes Moreno, C.; Valdez, A. A Simple and Efficient Protocol for Plant Regeneration and Genetic Transformation of Tomato cv. Micro-Tom from Leaf Explants. HortScience 2011, 46, 1655–1660. [Google Scholar] [CrossRef]
- Billah, M.; Banu, T.A.; Islam, M.; Banu, N.A.; Khan, S.; Akter, S.; Habib, A. In vitro regeneration and molecular characterization of some varieties of Lycopersicon esculentum Mill. in Bangladesh. Bangladesh J. Sci. Ind. Res. 2019, 54, 117–124. [Google Scholar] [CrossRef]
- Zaman, M.R.U.; Bhuiyan, M.S.U.; Ahmed, T.; Haque, M.K.; Uddin, M. In Vitro Plant Regeneration of Tomato (Lycopersicon esculentum). SF J. Agri Crop. Manag. 2020, 1, 1001. [Google Scholar]
- Sarker, R.; Islam, K.; Hoque, I. In vitro Regeneration and Agrobacterium-mediated Genetic Transformation of Tomato (Lycopersicon esculentum Mill.). Plant Tissue Cult. Biotechnol. 2010, 19, 101–111. [Google Scholar] [CrossRef]
- Jamous, F.; Abu-Qaoud, H. In vitro regeneration of tomato (Lycopersicon esculentum Mill). Plant Cell Biotechnol. Mol. Biol. 2015, 16, 181–190. [Google Scholar]
- Jawad, A.U.; Ay, A.-M.; Aj, K.; Al-Sadi, A. Enhanced somatic embryogenesis and Agrobacterium-mediated transformation of three cultivars of tomato by exogeneous application of putrescine. Int. J. Agric. Biol. 2014, 16, 81–88. [Google Scholar]
- Ashakiran, K.; Sivankalyani, V.; Malaiy; Jayanthi, I.; Govindasamy, V.; Girija, S. Genotype specific shoots regeneration from different explants of tomato (Solanum lycopersicum L.) using TDZ. Asian J. Plant. Sci. Res. 2011, 1, 107–113. [Google Scholar]
- Jehan, S.; Hassanein, A. Hormonal Requirements Trigger Different Organogenic Pathways on Tomato Nodal Explants. Am. J. Plant Sci. 2013, 4, 2118–2125. [Google Scholar] [CrossRef]
- Osman, M.; Elhadi, E.; Khalafalla, M. Callus formation and organogenesis of tomato (Lycopersicon esculentum Mill, CV Omdurman) induced by thidiazuron. Afr. J. Biotechnol. 2010, 9, 4407–4413. [Google Scholar]
- Gubis, J.; Lajchová, Z.; Faragó, J.; Jureková, Z. Effect of growth regulators on shoot induction and plant regeneration in tomato (Lycopersicon esculentum Mill.). Biol.-Sect. Cell. Mol. Biol. 2004, 59, 405–408. [Google Scholar]
- Baye, E.; Matewos, T.; Belew, D. Effect of 6-Benzyl Amino Purine on In Vitro Multiplication of Tomato (Lycopersicon esculentum Mill.) Varieties using Shoot Explant. J. Plant. Sci. Agric. Res. 2020, 4, 32. [Google Scholar]
- Mohamed, N.; Ismail, M.; Rahman, M. In vitro response from cotyledon and hypocotyls explants in tomato by inducing 6-benzylaminopurine. Afr. J. Biotechnol. 2010, 9, 4802–4807. [Google Scholar]
- Jawad, Z.A.; Turker, M.; Ozdemir, F.A. Effect of different plant growth regulator on in vitro propagation of endangered plant; yellow tomato (Lycopersicon esculentum Mill.). Int. J. Agric. For. Life Sci. 2020, 4, 92–98. [Google Scholar]
- Hanur, V.S.; Krishnareddy, B. In-Vitro Organogenesis in Tomato (Solanum Lycopersicum) Using Kinetin. Adv. Plants Agric. Res. 2016, 4, 397–401. [Google Scholar] [CrossRef]
- Brassard, N.; Brissette, L.; Lord, D.; Laliberté, S. Elongation, rooting and acclimatization of micropropagated shoots from mature material of hybrid larch. Plant Cell Tissue Organ Cult. 1996, 44, 37–44. [Google Scholar] [CrossRef]
- Arulananthu, G.; Bhat, S.G.; Ramesh, N. Callus induction and in vitro regeneration of tomato (Lycopersicon esculentum L.). Res. J. Life Sci. Bioinform. Pharm. Chem. Sci. 2019, 5, 491–503. [Google Scholar]
- Yesmin, S.; Mollika, S.R.; Islam, M.N.; Nasrin, S. In vitro Regeneration of two BINA Tomato (Lycopersicon esculentum Mill.) Varieties of Bangladesh. Plant Tissue Cult. Biotech. 2022, 32, 43–51. [Google Scholar] [CrossRef]
- Siti Suhaila, A.R.; Saleh, N.M. Inhibitory effect of Kanamycin on in vitro culture of Lycopersicon esculentum Mill cv. MT11. J. Agrobiotech. 2010, 1, 79–86. [Google Scholar]
- Stavridou, E.; Tzioutziou, N.; Madesis, P.; Labrou, N.; Nianiou, I. Effect of different factors on regeneration and transformation efficiency of tomato (Lycopersicum esculentum) hybrids. Czech J. Genet. Plant Breed. 2019, 55, 120–127. [Google Scholar] [CrossRef]
- Velcheva, M.; Faltin, Z.; Flaishman, M.; Eshdat, Y.; Perl, A. A liquid culture system for Agrobacterium-mediated transformation of tomato (Lycopersicon esculentum L. Mill.). Plant Sci. 2005, 168, 121–130. [Google Scholar] [CrossRef]
- Ying, C.Y.; Huang, X.Q.; Guo, Y.Q.; Zhong, L.L.; Liu, Y.; Li, S.L.; Gu, X.M.; Zhou, X.H. Optimization of tomato genetic transformation, kanamycin-resistant screening and seed selection. J. South. Med. Univ. 2008, 28, 1117–1122. [Google Scholar]
- Chen, C.; Xiaoyan, F.; Peng, R.; Tian, Y.; Yao, Q. Detoxifying processes during kanamycin-induced stress to Arabidopsis thaliana seedling growth. Biotechnol. Biotechnol. Equip. 2020, 34, 673–679. [Google Scholar] [CrossRef]
- Duan, H.; Ding, X.; Song, J.; Zhikun, D.; Zhou, Y.; Zhou, C. Effects of kanamycin on growth and development of Arabidopsis thaliana seedling, cotyledon and leaf. Pak. J. Bot. 2009, 41, 1611–1618. [Google Scholar]
- Hung, C.-Y.; Xie, J. Development of an Efficient Plant Regeneration System for the Selenium-hyperaccumulator Astragalus racemosus and the Nonaccumulator Astragalus canadensis. Hortscience 2008, 43, 2138–2142. [Google Scholar] [CrossRef]
- Sharma, C.; Aggarwal, G.; Srivastava, D.K. Effect of antibiotic kanamycin on cultured cotyledon and hypocotyl tissues of Tomato (Solanum lycopersicon cv. Solan vajr). Int. J. Agric. Environ. Biotechnol. 2012, 5, 77–82. [Google Scholar]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | BAP + IAA (mg/L) | Hypocotyls (E1) on Corresponding Treatment | Cotyledons (E2) on Corresponding Treatment |
|---|---|---|---|
| T1 | 0 + 0 | HT1 | CT1 |
| T2 | 1 + 0 | HT2 | CT2 |
| T3 | 2 + 0 | HT3 | CT3 |
| T4 | 3 + 0 | HT4 | CT4 |
| T5 | 4 + 0 | HT5 | CT5 |
| T6 | 0 + 0.05 | HT6 | CT6 |
| T7 | 0 + 0.1 | HT7 | CT7 |
| T8 | 0 + 0.5 | HT8 | CT8 |
| T9 | 1 + 0.05 | HT9 | CT9 |
| T10 | 2 + 0.05 | HT10 | CT10 |
| T11 | 3 + 0.05 | HT11 | CT11 |
| T12 | 4 + 0.05 | HT12 | CT12 |
| T13 | 1 + 0.1 | HT13 | CT13 |
| T14 | 2 + 0.1 | HT14 | CT14 |
| T15 | 3 + 0.1 | HT15 | CT15 |
| T16 | 4 + 0.1 | HT16 | CT16 |
| T17 | 1 + 0.5 | HT17 | CT17 |
| T18 | 2 + 0.5 | HT18 | CT18 |
| T19 | 3 + 0.5 | HT19 | CT19 |
| T20 | 4 + 0.5 | HT20 | CT20 |
| Treatment | % Explants Producing Direct Shoots | % Explants Producing Root | No. of Shoots per Explant | Type of Induced Organ |
|---|---|---|---|---|
| H = Hypocotyl | 79.63 a ± 2.73 | 22.19 a ± 3.97 | 2.73 a ± 0.13 | - |
| C = Cotyledon | 54.28 b ± 3.95 | 17.11 b ± 3.94 | 2.59 a ± 0.19 | - |
| Level of significance | ** | ** | ns | - |
| MSD value | 1.06 | 0.6 | 0.14 | - |
| CV (%) | 5.04 | 9.74 | 17.08 | - |
| HT1 | 31.00 k ± 2.45 | 62.50 b ± 2.55 | 1.00 ef ± 0 | RS |
| HT2 | 83.50 d–g ± 1.50 | 7.81 cd ± 1.56 | 2.25 de ± 0.25 | SR |
| HT3 | 90.50 a–e ± 2.02 | 0 e ± 0 | 2.25 de ± 0.25 | S |
| HT4 | 92.25 a–d ± 1.75 | 0 e ± 0 | 3.25 cd ± 0.25 | SCL |
| HT5 | 92.25 a–d ± 1.75 | 0 e ± 0 | 3.25 cd ± 0.25 | SCL |
| HT6 | 48.50 j ± 1.50 | 100 a ± 0 | 2.25 de ± 0.25 | RS |
| HT7 | 82.50 e-h ± 1.50 | 100 a ± 50 | 3.25 cd ± 0.25 | RS |
| HT8 | 0 m ± 0 | 100 a ± 0 | 0.00 f ± 0 | R |
| HT9 | 90.50 a–e ± 2.02 | 6.25 d ± 0 | 3.25 cd ± 0.25 | SR |
| HT10 | 92.25 a–d ± 1.75 | 0 e ± 0 | 2.25 de ± 0.25 | S |
| HT11 | 82.50 e–h ± 1.50 | 0 eh ± 0 | 2.25 de ± 0.25 | LCS |
| HT12 | 82.50 e–h ± 1.50 | 0 e ± 0 | 2.25 de ± 0.25 | LCS |
| HT13 | 92.25 a–d ± 1.75 | 9.38 cd ± 1.80 | 2.25 de ± 0.25 | SRL |
| HT14 | 98.50 a ± 1.50 | 10.94 cd ± 1.56 | 5.25 ab ± 0.25 | SRL |
| HT15 | 82.50 e–h ± 1.50 | 6.25 d ± 0 | 2.25 de ± 0.25 | SRL |
| HT16 | 88.75 b–f ± 1.75 | 6.25 d ± 0 | 3.25 cd ± 0.25 | SRL |
| HT17 | 90.50 a–e ± 2.02 | 12.50 c ± 2.55 | 3.25 cd ± 0.25 | SRL |
| HT18 | 97.00 ab ± 1.73 | 7.81 cd ± 1.56 | 4.25 bc ± 0.25 | SRL |
| HT19 | 88.75 b–f ± 1.75 | 7.81 cd ± 1.56 | 3.25 cd ± 0.25 | SRLC |
| HT20 | 86.00 efg ± 1.00 | 6.25 d ± 0 | 3.25 cd ± 0.25 | SRLC |
| CT1 | 0 m ± 0 | 20.31 c ± 0 | 00 f ± 0 | R |
| CT2 | 13.50 l ± 1.50 | 0 e ± 0 | 1.00 ef ± 0 | S |
| CT3 | 49.75 j ± 2.66 | 0 e ± 0 | 2.25 de ± 0.25 | S |
| CT4 | 26.50 k ± 1.50 | 0 e ± 0 | 2.25 de ± 0.25 | LSC |
| CT5 | 31.00 k ± 2.45 | 0 e ± 0 | 2.25 de ± 0.25 | LSC |
| CT6 | 0 m ± 0 | 100 a ± 0 | 00 f ± 0 | R |
| CT7 | 0 m ± 0 | 100 a ± 0 | 00 f ± 0 | R |
| CT8 | 0 m ± 0 | 100 a ± 0 | 00 f ± 0 | R |
| CT9 | 54.50 j ± 2.87 | 0 e ± 0 | 2.25 de ± 0.25 | SL |
| CT10 | 79.50 f–i ± 1.50 | 0 e ± 0 | 4.25 bc ± 0.25 | SL |
| CT11 | 73.25 hi ± 1.75 | 0 e ± 0 | 3.25 cd ± 0.25 | LS |
| CT12 | 71.50 i ± 2.02 | 0 e ± 0 | 3.25 cd ± 0.25 | LS |
| CT13 | 82.50 e–h ± 1.50 | 6.25 d ± 0 | 2.25 de ± 0.25 | SRL |
| CT14 | 95.50 abc ± 1.50 | 0 e ± 0 | 4.25 bc ± 0.25 | SL |
| CT15 | 76.50 ghi ± 1.50 | 0 e ± 0 | 3.25 cd ± 0.25 | LCS |
| CT16 | 76.50 ghi ± 1.50 | 0 ei ± 0 | 4.25 bc ± 0.25 | LCS |
| CT17 | 85.50 d–g ± 1.50 | 10.94 cd ± 1.56 | 4.25 bc ± 0.25 | SRL |
| CT18 | 98.50 a ± 1.50 | 9.38 cd ± 1.80 | 6.25 a ± 0.25 | SRL |
| CT19 | 87.00 c–f ± 2.44 | 7.81 cd ± 1.56 | 3.25 cd ± 0.25 | LSRC |
| CT20 | 84.00 d–g ± 1.73 | 7.81 cd ± 1.56 | 3.25 cd ± 0.25 | LSRC |
| MSD value | 9.28 | 5.42 | 1.28 | - |
| CV (%) | 4.91 | 9.77 | 17.01 | - |
| Level of (E × T) significance | ** | ** | ** | - |
| Treatment | % Shoot Induced Root | No. of Roots per Shoot | Root Length (cm) | Root Diameter (cm) | Survival Rate in Soil (%) |
|---|---|---|---|---|---|
| SH = Shoot from hypocotyl | 97.50 a ± 0.02 | 20.45 a ± 2.50 | 6.70 a ± 0.056 | 0.38 a ± 0.03 | 60.00 a ± 6.28 |
| SC = Shoot from cotyledon | 61.50 b ± 0.02 | 8.35 b ± 1.16 | 5.41 b ± 0.27 | 0.34 b ± 0.03 | 55.00 b ± 6.75 |
| Level of significance | ** | ** | ** | * | * |
| MSD value | 2.89 | 1.58 | 0.9 | 0.03 | 3.99 |
| CV (%) | 5.62 | 16.96 | 23.11 | 15.94 | 10.77 |
| SH I1 | 87.50 b ± 2.50 | 3.75 e ± 1.11 | 5.75 b ± 0.85 | 0.20 e ± 0.04 | 12.50 c ± 2.50 |
| SH I2 | 100 a ± 00 | 15.00 c ± 1.47 | 5.88 b ± 0.43 | 0.35 cd ± 0.03 | 47.50 b ± 2.50 |
| SH I3 | 100 a ± 00 | 20.75 b ± 1.75 | 10.50 a ± 1.04 | 0.35 cd ± 0.03 | 75.00 a ± 2.88 |
| SH I4 | 100 a ± 00 | 34.00 a ± 1.68 | 5.13 b ± 1.01 | 0.43 bc ± 0.03 | 82.50 a ± 2.50 |
| SH I5 | 100 a ± 00 | 28.75 a ± 1.49 | 6.25 b ± 0.85 | 0.58 a ± 0.05 | 82.50 a ± 2.50 |
| SC I1 | 27.50 e ± 2.50 | 1.50 e ± 0.29 | 5.25 b ± 0.32 | 0.18 e ± 0.03 | 5.00 c ± 2.88 |
| SC I2 | 47.00 d ± 2.50 | 4.00 e ± 0.91 | 5.13 b ± 0.43 | 0.28 de ± 0.03 | 40.00 b ± 4.08 |
| SC I3 | 65.00 c ± 2.88 | 9.50 d ± 0.96 | 7.06 ab ± 0.41 | 0.30 cde ± 0 | 75.00 a ± 2.88 |
| SC I4 | 80.00 b ± 4.08 | 14.00 cd ± 0.91 | 4.88 b ± 0.69 | 0.40 bcd ± 0 | 77.50 a ± 2.50 |
| SC I5 | 87.50 b ± 2.50 | 12.75 cd ± 0.85 | 4.75 b ± 0.48 | 0.53 ab ± 0.03 | 77.50 a ± 4.78 |
| MSD value | 11.18 | 5.47 | 3.53 | 0.14 | 14.35 |
| CV (%) | 5.78 | 15.6 | 23.95 | 16.21 | 10.26 |
| Interaction level (S × I) of significance | ** | ** | ** | ** | ** |
| Treatment | % Explants Forming Shoots | No. of Shoots per Explant |
|---|---|---|
| E1 = Hypocotyl | 20.07 a ± 6.03 | 1.03 a ± 0.30 |
| E2 = Cotyledon | 12.30 b ± 5.85 | 0.81 b ± 0.39 |
| Level of significance | ** | ** |
| MSD value | 1.02 | 0.21 |
| CV (%) | 12.48 | 46.32 |
| T1 | 98.34 a ± 1.09 | 5.63 a ± 0.50 |
| T2 | 24.22 b ± 9.26 | 1.25 b ± 0.50 |
| T3 | 6.94 c ± 2.70 | 0.50 c ± 0.19 |
| T4 | 0 d ± 0 | 0 c ± 0 |
| T5 | 0 d ±0 | 0 c ± 0 |
| T6 | 0 d ±0 | 0 c ± 0 |
| T7 | 0 d ±0 | 0 c ± 0 |
| T8 | 0 d ±0 | 0 c ± 0 |
| Level of significance | ** | ** |
| MSD value | 3.2 | 0.67 |
| CV (%) | 12.48 | 46.32 |
| E1T1 | 98.25 a ± 1.75 | 4.75 b ± 0.48 |
| E1T2 | 48.44 b ± 2.99 | 2.50 c ± 0.29 |
| E1T3 | 13.88 c ± 1.38 | 1.00 d ± 0 |
| E1T4 | 0 d ± 0 | 0 d ± 0 |
| E1T5 | 0 d ± 0 | 0 d ± 0 |
| E1T6 | 0 d ± 0 | 0 d ± 0 |
| E1T7 | 0 d ± 0 | 0 d ± 0 |
| E1T8 | 0 d ± 0 | 0 d ± 0 |
| E2T1 | 98.44 a ± 1.56 | 6.50 a ± 0.65 |
| E2T2 | 0 d ± 0 | 0 d ± 0 |
| E2T3 | 0 d ± 0 | 0 d ± 0 |
| E2T4 | 0 d ± 0 | 0 d ± 0 |
| E2T5 | 0 d ± 0 | 0 d ± 0 |
| E2T6 | 0 d ± 0 | 0 d ± 0 |
| E2T7 | 0 d ± 0 | 0 d ± 0 |
| E2T8 | 0 d ± 0 | 0 d ± 0 |
| MSD value | 5.24 | 1.09 |
| CV (%) | 12.63 | 46.32 |
| Interaction level (E × T) of significance | ** | ** |
| Treatment | % Shoots Forming Roots | No. of Shoots per Explant |
|---|---|---|
| S1 | 90.83 a ± 4.17 | 20.67 a ± 3.92 |
| S2 | 75.83 b ± 7.93 | 8.17 b ± 1.70 |
| S3 | 30.00 c ± 4.77 | 5.33 c ± 1.16 |
| Level of significance | ** | ** |
| MSD value | 7.42 | 1.98 |
| CV (%) | 11.18 | 17.19 |
| T1 | 40.83 b ± 8.02 | 1.42 c ± 0.36 |
| T2 | 78.33 a ± 8.33 | 18.50 a ± 3.02 |
| T3 | 77.50 a ± 8.27 | 14.25 b ± 2.84 |
| Level of significance | ** | ** |
| MSD value | 7.42 | 1.98 |
| CV (%) | 11.18 | 17.19 |
| S1T1 | 72.50 b ± 4.79 | 2.75 ef ± 0.48 |
| S1T2 | 100 a ± 0 | 32.00 a ± 1.83 |
| S1T3 | 100 a ± 0 | 27.25 b ± 1.11 |
| S2T1 | 40.00 c ± 4.08 | 1.00 f ± 0.41 |
| S2T2 | 95.00 a ± 2.89 | 14.25 c ± 0.85 |
| S2T3 | 92.50 a ± 4.79 | 9.25 d ± 1.11 |
| S3T1 | 10.00 d ± 4.08 | 0.50 f ± 0.29 |
| S3T2 | 40.00 c ± 4.08 | 9.25 d ± 1.11 |
| S3T3 | 40.00 c ± 4.08 | 6.25 de ± 0.63 |
| MSD value | 18.39 | 4.39 |
| CV (%) | 11.67 | 16.03 |
| Interaction level (E × G) significance | ** | ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmed, S.; Wan Azizan, W.A.S.; Akhond, M.A.Y.; Juraimi, A.S.; Ismail, S.I.; Ahmed, R.; Md Hatta, M.A. Optimization of In Vitro Regeneration Protocol of Tomato cv. MT1 for Genetic Transformation. Horticulturae 2023, 9, 800. https://doi.org/10.3390/horticulturae9070800
Ahmed S, Wan Azizan WAS, Akhond MAY, Juraimi AS, Ismail SI, Ahmed R, Md Hatta MA. Optimization of In Vitro Regeneration Protocol of Tomato cv. MT1 for Genetic Transformation. Horticulturae. 2023; 9(7):800. https://doi.org/10.3390/horticulturae9070800
Chicago/Turabian StyleAhmed, Shiuli, Wan Aina Sakeenah Wan Azizan, Md. Abdullah Yousuf Akhond, Abdul Shukor Juraimi, Siti Izera Ismail, Razu Ahmed, and Muhammad Asyraf Md Hatta. 2023. "Optimization of In Vitro Regeneration Protocol of Tomato cv. MT1 for Genetic Transformation" Horticulturae 9, no. 7: 800. https://doi.org/10.3390/horticulturae9070800
APA StyleAhmed, S., Wan Azizan, W. A. S., Akhond, M. A. Y., Juraimi, A. S., Ismail, S. I., Ahmed, R., & Md Hatta, M. A. (2023). Optimization of In Vitro Regeneration Protocol of Tomato cv. MT1 for Genetic Transformation. Horticulturae, 9(7), 800. https://doi.org/10.3390/horticulturae9070800