
Proteomics Research Reveals the Molecular Mechanism by Which Grape Seed Oil Inhibits Tuber Sprouting in Potato

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Treatment of Potato Tuber
2.3. Measurement of POD Activity and Soluble Sugar
2.4. Proteomic Analysis
2.5. Bioinformatics Analysis of DAPs
2.6. qRT-PCR Analysis
2.7. Statistical Analysis
3. Results
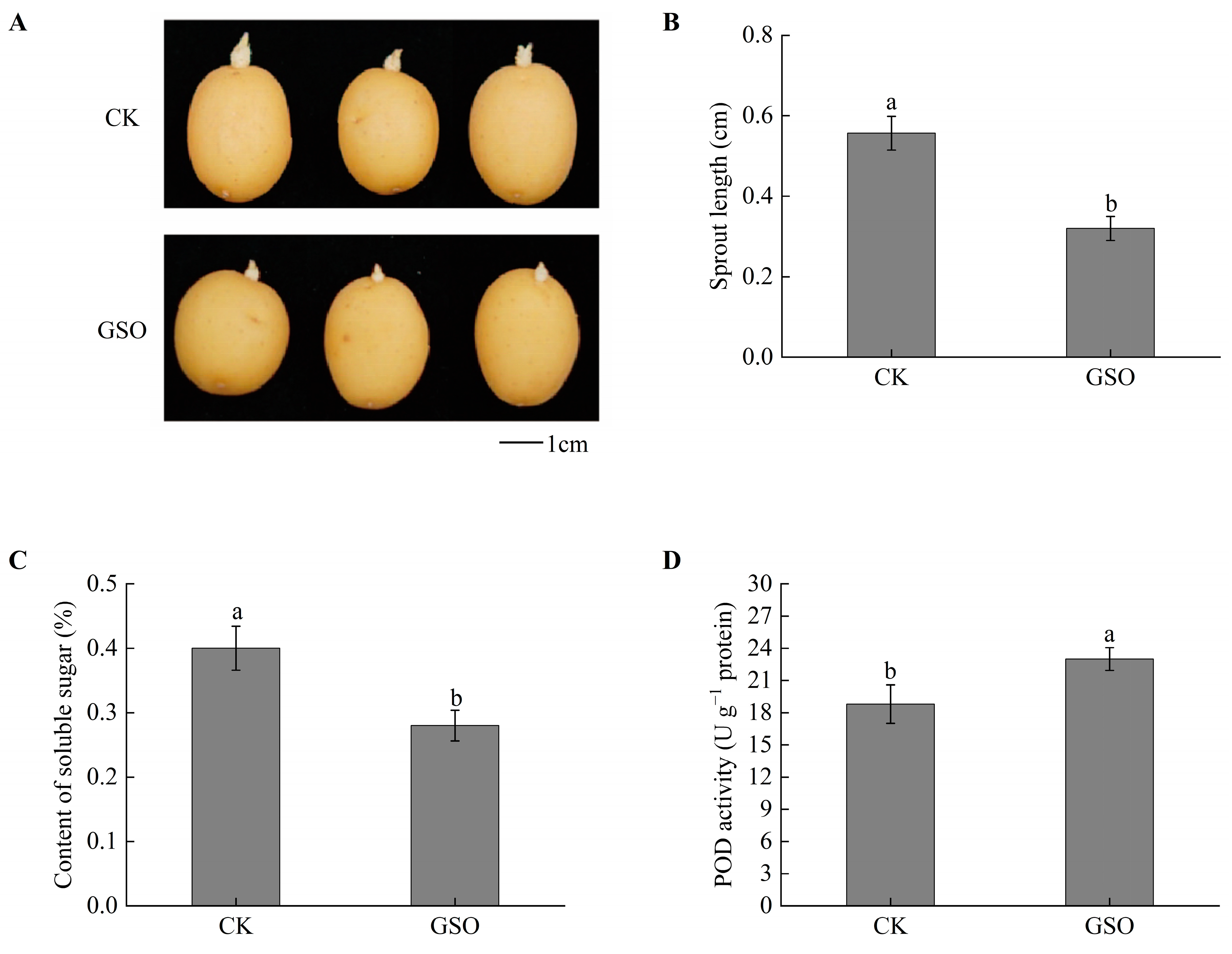
3.1. Effects of GSO Treatment on Potato Tuber Physiology
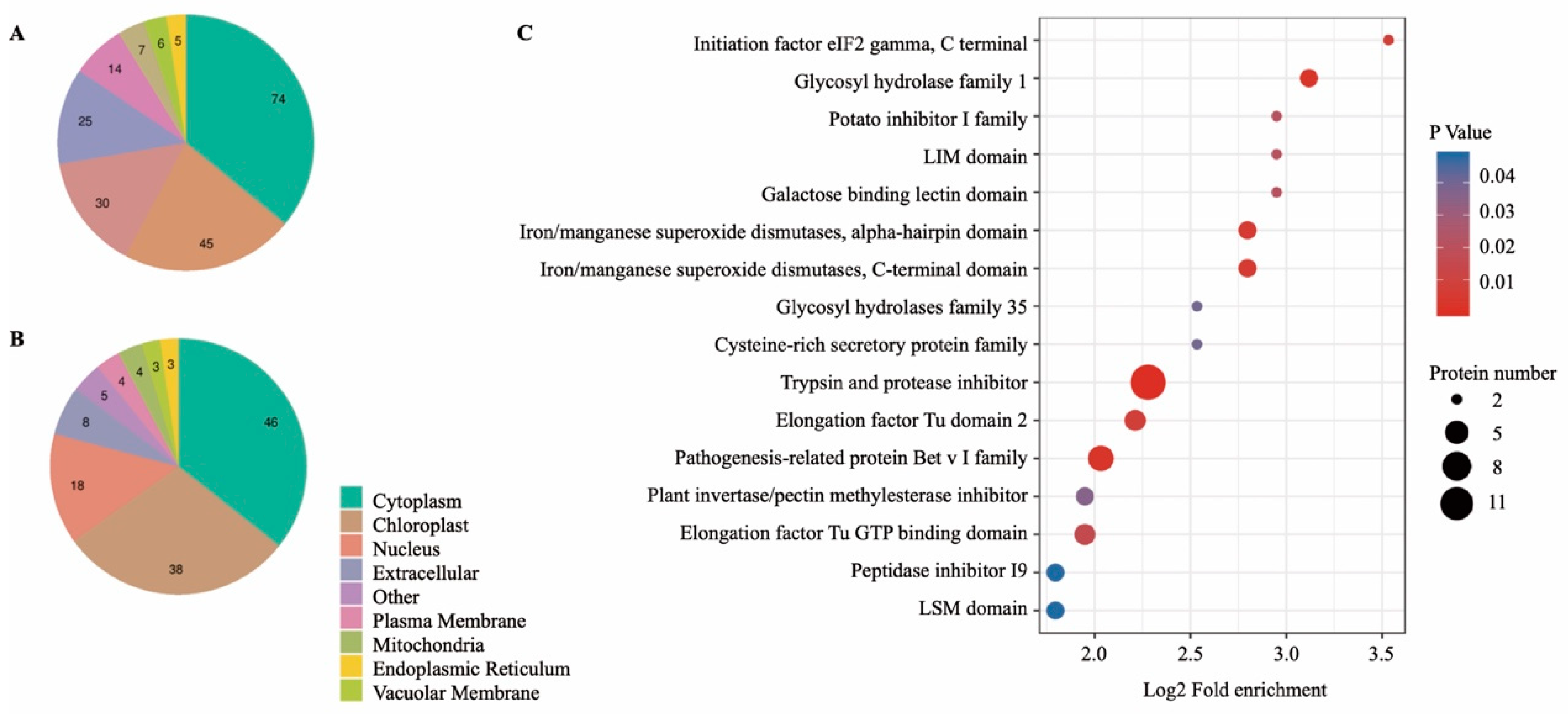
3.2. Subcellular Location and Protein Domain Analysis of DAPs
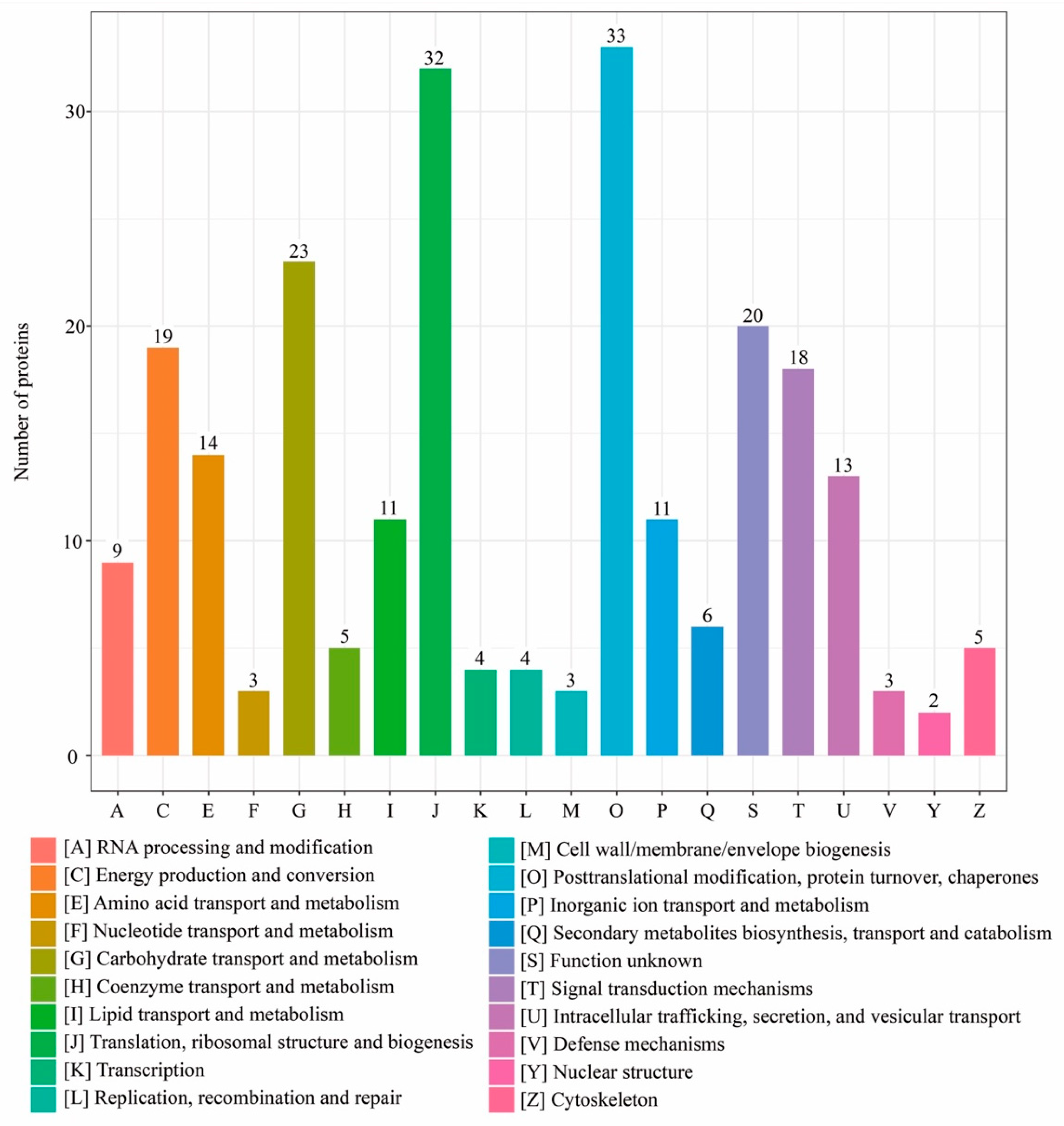
3.3. COG/KOG Functional Annotation
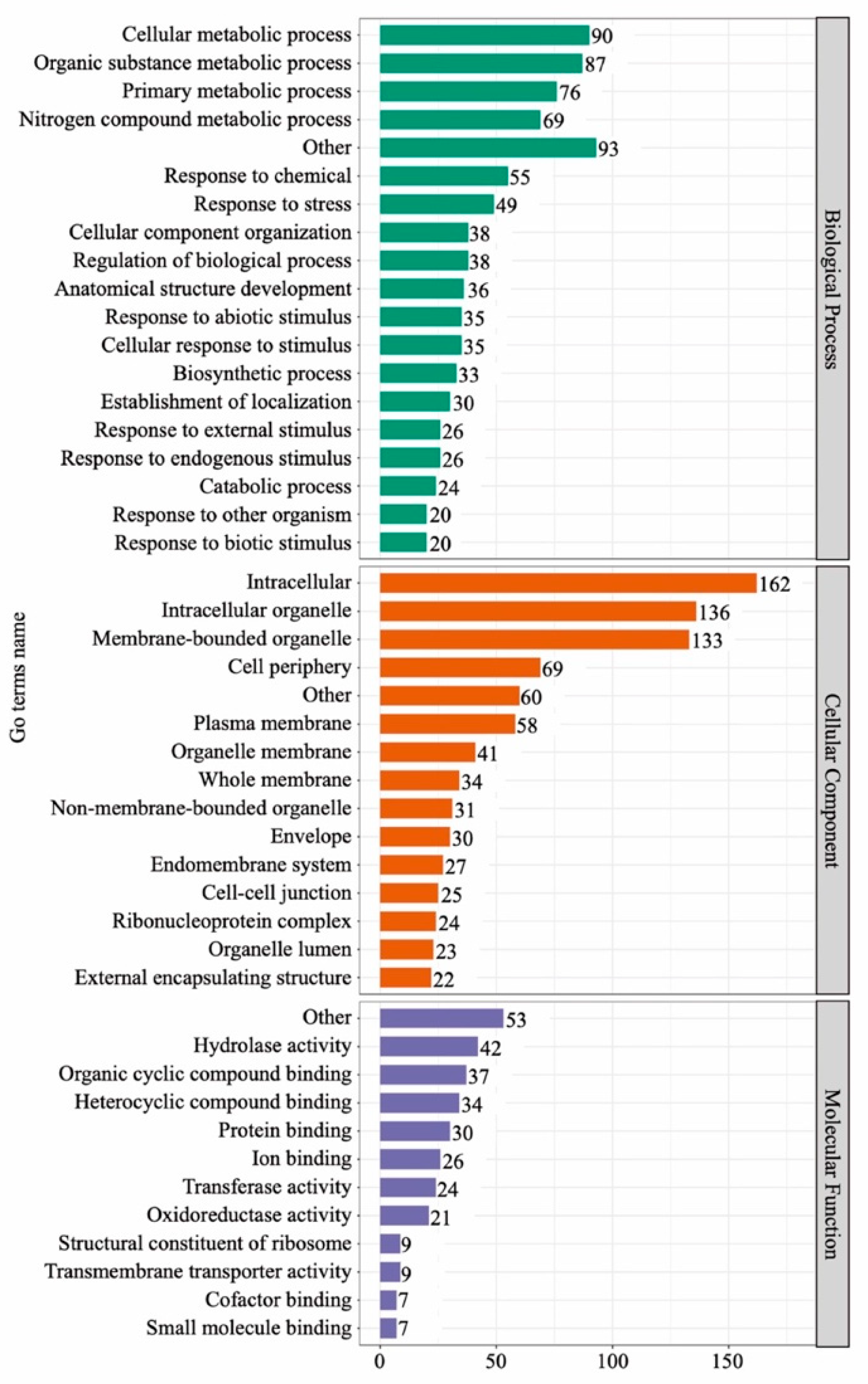
3.4. GO and KEGG Analysis of DAPs
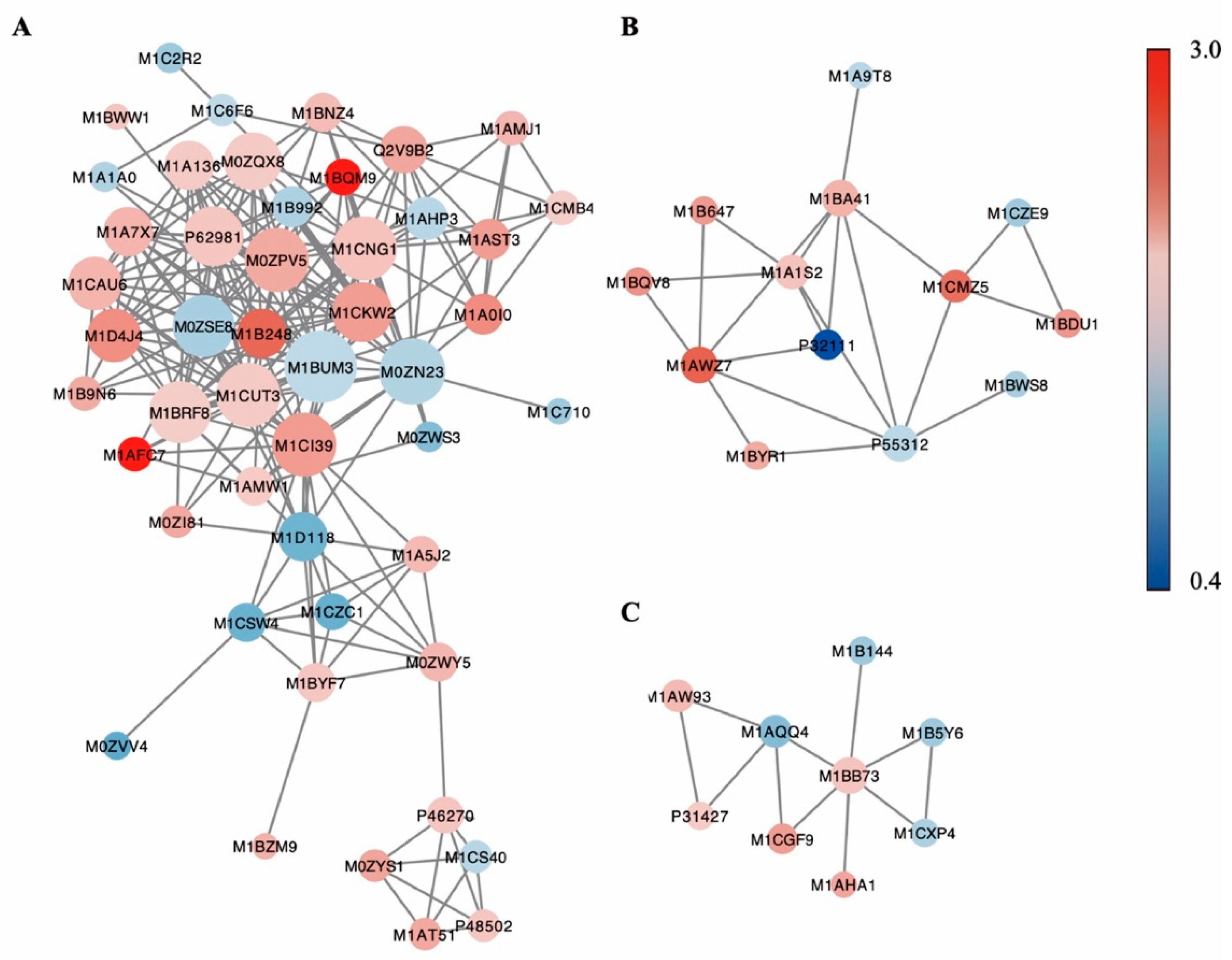
3.5. Protein-Protein Interaction Network Analysis
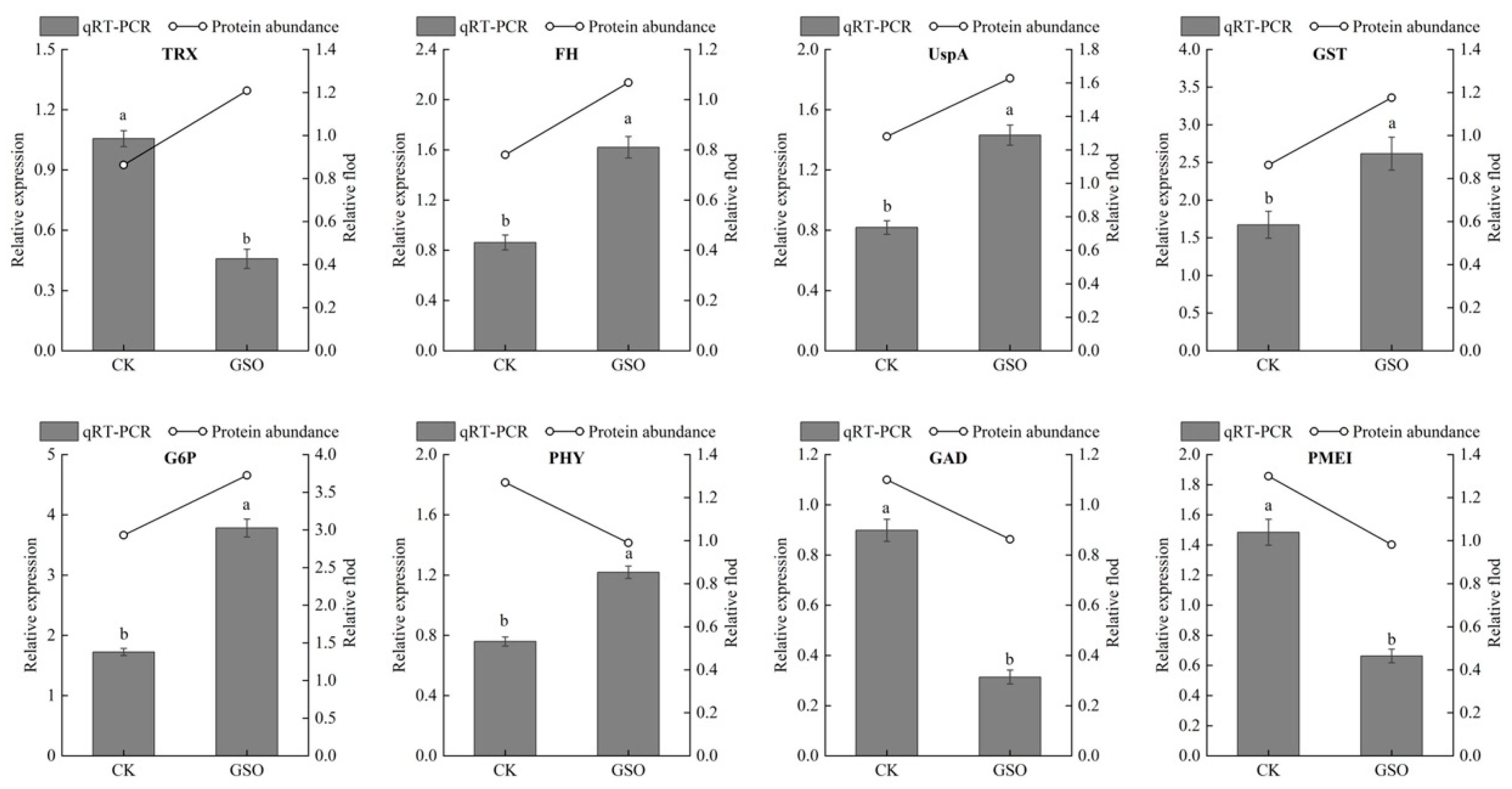
3.6. Complementation of the Proteomic Results via qRT-PCR
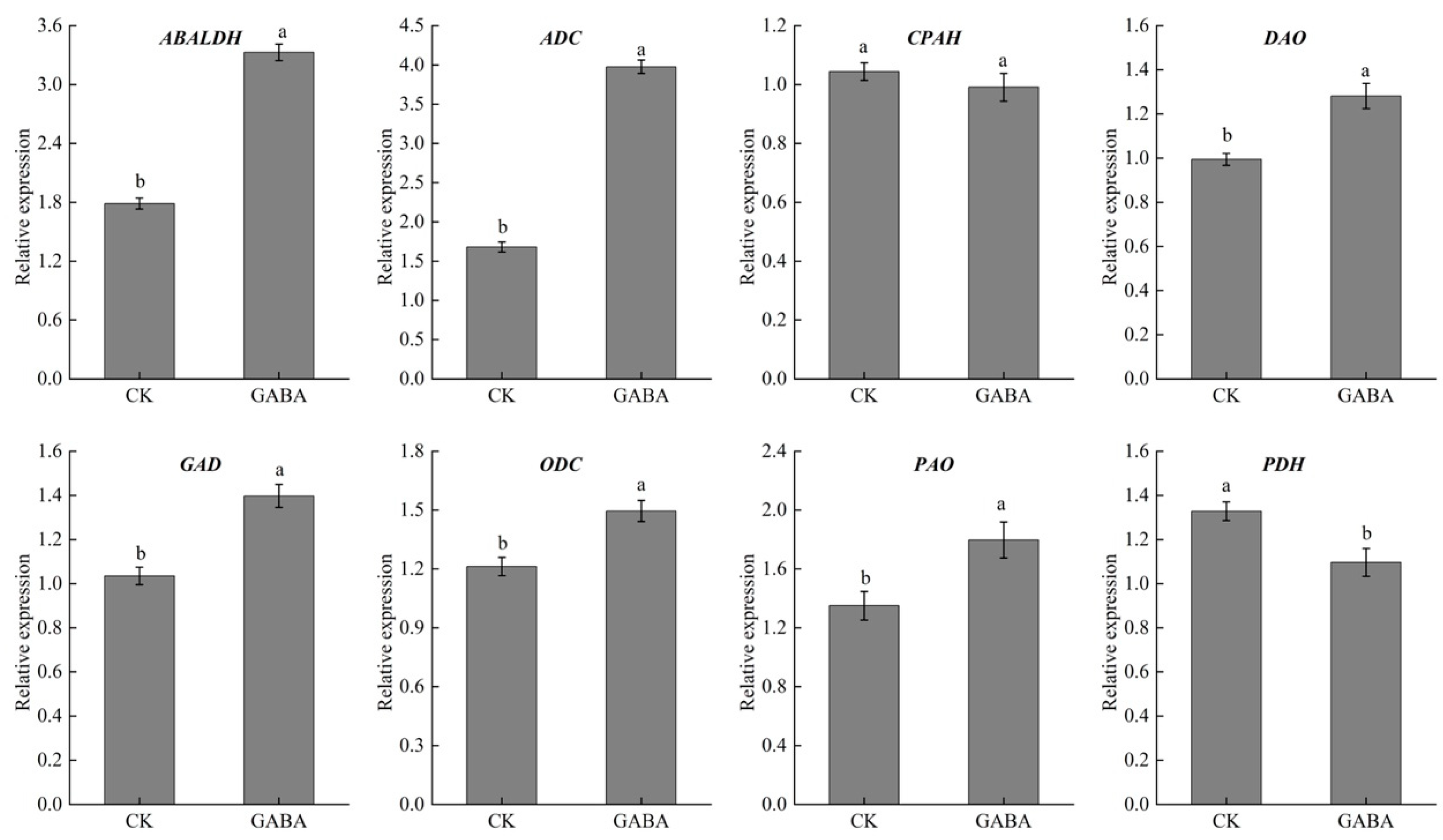
3.7. Analysis of Important DAPs in Tuber after GSO Treatment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Devaux, A.; Goffart, J.-P.; Petsakos, A.; Kromann, P.; Gatto, M.; Okello, J.; Suarez, V.; Hareau, G. Global food security, contributions from sustainable potato agri-food systems. In The Potato Crop; Springer: Cham, Switzerland, 2020; pp. 3–35. [Google Scholar]
- Gumbo, N.; Magwaza, L.S.; Ngobese, N.Z. Evaluating Ecologically Acceptable Sprout Suppressants for Enhancing Dormancy and Potato Storability: A Review. Plants 2021, 10, 2307. [Google Scholar] [CrossRef]
- Wanjiku, M.W.; Ombui, N.R.; Igosangwa, S.S. Effect of Storage Temperature and Postharvest Tuber Treatment with Chemical and Biorational Inhibitors on Suppression of Sprouts During Potato Storage. J. Hortic. Res. 2021, 29, 83–94. [Google Scholar]
- Etemadinasab, H.; Zahedi, M.; Ramin, A.-A.; Kadivar, M. Effects of electron beam irradiation on physicochemical, nutritional properties and storage life of five potato cultivars. Radiat. Phys. Chem. 2020, 177, 109093. [Google Scholar] [CrossRef]
- Paul, V.; Ezekiel, R.; Pandey, R. Sprout suppression on potato: Need to look beyond CIPC for more effective and safer alternatives. J. Food Sci. Technol. 2016, 53, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Vijay, P.; Ezekiel, R.; Pandey, R. Use of CIPC as a potato sprout suppressant: Health and environmental concerns and future options. Qual. Assur. Saf. Crops Foods 2018, 10, 17–24. [Google Scholar] [CrossRef]
- Yuan, L.; Wang, J.; Guan, Z.; Yue, F.; Wang, S.; Chen, Q.; Fu, M. Optimized Preparation of Methyl Salicylate Hydrogel and Its Inhibition Effect on Potato Tuber Sprouting. Horticulturae 2022, 8, 866. [Google Scholar] [CrossRef]
- Sivakumar, D.; Bautista-Baños, S. A review on the use of essential oils for postharvest decay control and maintenance of fruit quality during storage. Crop Prot. 2014, 64, 27–37. [Google Scholar] [CrossRef]
- Jia, B.; Xu, L.; Guan, W.; Lin, Q.; Brennan, C.; Yan, R.; Zhao, H. Effect of citronella essential oil fumigation on sprout suppression and quality of potato tubers during storage. Food Chem. 2019, 284, 254–258. [Google Scholar] [CrossRef]
- Gómez-Castillo, D.; Cruz, E.; Iguaz, A.; Arroqui, C.; Vírseda, P. Effects of essential oils on sprout suppression and quality of potato cultivars. Postharvest Biol. Technol. 2013, 82, 15–21. [Google Scholar] [CrossRef]
- Li, L.; Chen, J.; Li, Z.; Li, H.; Yang, S.; Ren, B.; Lu, Y.; Zheng, S.; Yu, L.; Wang, X. Proteomic analysis of garlic essential oil-treated potato reveals that StHSP26. 5 as a vital gene involving in tuber sprouting. Postharvest Biol. Technol. 2022, 183, 111725. [Google Scholar] [CrossRef]
- Martín, M.; Grao, E.; Millán, M.; Montserrat, S. Grape (Vitis vinifera L.) Seed Oil: A Functional Food from the Winemaking Industry. Foods 2020, 9, 1360. [Google Scholar] [CrossRef]
- Garavaglia, J.; Markoski, M.; Oliveira, A.; Marcadenti, A. Grape seed oil compounds: Biological and chemical actions for health. Nutr. Metab. Insights 2016, 9, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Ullah, F.; Zhou, D.; Yi, M.; Zhao, Y. Mechanisms of ROS regulation of plant development and stress responses. Front. Plant Sci. 2019, 10, 800. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Zhao, S.; Tan, F.; Zhao, H.; Wang, D.; Si, H.; Chen, Q. Changes in ROS production and antioxidant capacity during tuber sprouting in potato. Food Chem. 2017, 237, 205–213. [Google Scholar] [CrossRef]
- Subotin, I.; Druta, R.; Popovici, V.; Covaci, E.; Sturza, R. Kinetic of Forced Oxidation of Grape Seeds, Walnuts and Corn Germs Oils in the Presence of Antioxidants. Food Nutr. Sci. 2021, 12, 1326–1339. [Google Scholar] [CrossRef]
- Ramos, R.; Poirot, E.; Flores, M.; Yildiz, F. GABA, a non-protein amino acid ubiquitous in food matrices. Cogent Food Agric. 2018, 4, 1–89. [Google Scholar]
- Bouche, N.; Fromm, H. GABA in plants: Just a metabolite? [Review]. Trends Plant Sci. 2004, 9, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Shelp, B.J.; Bown, A.W.; Zarei, A. 4-Aminobutyrate (GABA): A metabolite and signal with practical significance. Botany 2017, 95, 1015–1032. [Google Scholar] [CrossRef]
- Fait, A.; Fromm, H.; Walter, D.; Galili, G.; Fernie, A.R. Highway or byway: The metabolic role of the GABA shunt in plants. Trends Plant Sci. 2008, 13, 14–19. [Google Scholar] [CrossRef]
- Wei, C.; Chen, M.; Jing, J.; Mai, L.; Xiaoman, Z.; Yanyan, W.; Tiantian, X.; Changjian, D.; Jiacheng, S.; Zeping, J. Exogenous GABA promotes adaptation and growth by altering the carbon and nitrogen metabolic flux in poplar seedlings under low nitrogen conditions. Tree Physiol. 2020, 40, 1744–1761. [Google Scholar]
- Xu, B.; Long, Y.; Feng, X.; Zhu, X.; Gilliham, M. GABA signalling modulates stomatal opening to enhance plant water use efficiency and drought resilience. Nat. Commun. 2021, 12, 1952. [Google Scholar] [CrossRef] [PubMed]
- Shi, J. Inhibition of α-ketoglutarate dehydrogenase activity affects adventitious root growth in poplar via changes in GABA shunt. Planta Int. J. Plant Biol. 2018, 248, 963–979. [Google Scholar]
- Xie, T.; Ji, J.; Chen, W.; Yue, J.; Du, C.; Sun, J.; Chen, L.; Jiang, Z.; Shi, S. GABA negatively regulates adventitious root development in poplar. J. Exp. Bot. 2020, 71, 1459–1474. [Google Scholar] [CrossRef]
- Takayama, M.; Matsukura, C.; Ariizumi, T.; Ezura, H. Activating glutamate decarboxylase activity by removing the autoinhibitory domain leads to hyper γ-aminobutyric acid (GABA) accumulation in tomato fruit. Plant Cell Rep. 2016, 36, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Nonaka, S.; Arai, C.; Takayama, M.; Matsukura, C.; Ezura, H. Efficient increase of ɣ-aminobutyric acid (GABA) content in tomato fruits by targeted mutagenesis. Sci. Rep. 2017, 7, 7057. [Google Scholar] [CrossRef]
- Sheng, Y.; Xiao, H.; Guo, C.; Wu, H.; Wang, X. Effects of exogenous gamma-aminobutyric acid on α-amylase activity in the aleurone of barley seeds. Plant Physiol. Biochem. Ppb 2018, 127, 39. [Google Scholar] [CrossRef]
- Du, C.; Chen, W.; Wu, Y.; Wang, G.; Shi, S. Effects of GABA and Vigabatrin on the Germination of Chinese Chestnut Recalcitrant Seeds and Its Implications for Seed Dormancy and Storage. Plants 2020, 9, 449. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Zeng, Q.; Ren, Z.; Li, P.; Xu, X. Effect of exogenous gamma-aminobutyric acid treatment on the enzymatic browning of fresh-cut potato during storage. J. Food Sci. Technol. 2018, 55, 5035–5044. [Google Scholar] [CrossRef]
- Palma, F.; Carvajal, F.; Jiménez-Muñoz, R.; Pulido, A.; Jamilena, M.; Garrido, D. Exogenous gamma-aminobutyric acid treatment improves the cold tolerance of zucchini fruit during postharvest storage. Plant Physiol. Biochem. 2019, 136, 188–195. [Google Scholar] [CrossRef]
- Baranzelli, J.; Kringel, D.H.; Colussi, R.; Paiva, F.F.; Aranha, B.C.; Miranda, M.Z.D.; Zavareze, E.D.R.; Dias, A.R.G. Changes in enzymatic activity, technological quality and gamma-aminobutyric acid (GABA) content of wheat flour as affected by germination. LWT 2018, 90, 483–490. [Google Scholar] [CrossRef]
- Yang, R.; Han, Y.; Han, Z.; Ackah, S.; Li, Z.; Bi, Y.; Yang, Q.; Prusky, D. Hot water dipping stimulated wound healing of potato tubers. Postharvest Biol. Technol. 2020, 167, 111245. [Google Scholar] [CrossRef]
- Li, L.; Zou, X.; Deng, M.; Peng, J.; Huang, X.; Lu, X.; Fang, C.; Wang, X.-Y. Comparative Morphology, Transcription, and Proteomics Study Revealing the Key Molecular Mechanism of Camphor on the Potato Tuber Sprouting Effect. Int. J. Mol. Sci. 2017, 18, 2280. [Google Scholar] [CrossRef]
- Viola, R.; Pelloux, J.; Van Der Ploeg, A.; Gillespie, T.; Marquis, N.; Roberts, A.G.; Hancock, R.D. Symplastic connection is required for bud outgrowth following dormancy in potato (Solanum tuberosum L.) tubers. Plant Cell Environ. 2007, 30, 973–983. [Google Scholar] [CrossRef]
- Deng, M.; Peng, J.; Zhang, J.; Ran, S.; Cai, C.; Yu, L.; Ni, S.; Huang, X.; Li, L.; Wang, X. The cysteine-rich peptide snakin-2 negatively regulates tubers sprouting through modulating lignin biosynthesis and H2O2 accumulation in potato. Int. J. Mol. Sci. 2021, 22, 2287. [Google Scholar] [CrossRef] [PubMed]
- Sapkota, S.; Liu, J.; Islam, M.T.; Sherif, S.M. Changes in reactive oxygen species, antioxidants and carbohydrate metabolism in relation to dormancy transition and bud break in apple (Malus× domestica borkh) cultivars. Antioxidants 2021, 10, 1549. [Google Scholar] [CrossRef]
- Liu, B.; Zhang, N.; Wen, Y.; Jin, X.; Yang, J.; Si, H.; Wang, D. Transcriptomic changes during tuber dormancy release process revealed by RNA sequencing in potato. J. Biotechnol. 2015, 198, 17–30. [Google Scholar] [CrossRef]
- Vaughn, S.F.; Spencer, G.F. Volatile monoterpenes inhibit potato tuber sprouting. Am. Potato J. 1991, 68, 821–831. [Google Scholar] [CrossRef]
- Job, C.; Rajjou, L.; Lovigny, Y.; Belghazi, M.; Job, D. Patterns of protein oxidation in Arabidopsis seeds and during germination. Plant Physiol. 2005, 138, 790–802. [Google Scholar] [CrossRef]
- Oracz, K.; Bouteau, H.; Farrant, J.; Cooper, K.; Belghazi, M.; Job, C.; Job, D.; Corbineau, F.; Bailly, C. ROS production and protein oxidation as a novel mechanism for seed dormancy alleviation. Plant J. 2007, 50, 452–465. [Google Scholar] [CrossRef]
- Li, L.; Chen, J.; Lu, Y.; Ren, B.; Huang, X.; Yu, L.; Zeng, F.; Wang, Q.; Wang, X.; Lu, L. Physiological and proteomic analyses of γ-aminobutyric acid (GABA)-treated tubers reveals that StPOD42 promotes sprouting in potato. J. Plant Physiol. 2022, 278, 153826. [Google Scholar] [CrossRef]
- Sarath, G.; Hou, G.; Baird, L.M.; Mitchell, R.B. Reactive oxygen species, ABA and nitric oxide interactions on the germination of warm-season C4-grasses. Planta 2007, 226, 697–708. [Google Scholar] [CrossRef]
- Lv, J.; Bai, L.; Han, X.; Xu, D.; Ding, S.; Li, C.; Ge, Y.; Li, J. Effects of 1-MCP treatment on sprouting and preservation of ginger rhizomes during storage at room temperature. Food Chem. 2021, 349, 129004. [Google Scholar] [CrossRef] [PubMed]
- Buchanan, B.B.; Balmer, Y. Redox regulation: A broadening horizon. Annu. Rev. Plant Biol. 2005, 56, 187. [Google Scholar] [CrossRef] [PubMed]
- Silva, C.; Galhano, C.; Moreira, A. A new sprout inhibitor of potato tuber based on carvone/β-cyclodextrin inclusion compound. J. Incl. Phenom. Macrocycl. Chem. 2007, 57, 121–124. [Google Scholar] [CrossRef][Green Version]
- Knowles, L.O.; Knowles, N.R. Toxicity and metabolism of exogenous α, β-unsaturated carbonyls in potato (Solanum tuberosum L.) tubers. J. Agric. Food Chem. 2012, 60, 11173–11181. [Google Scholar] [CrossRef]
- Yang, L.; Liu, S.; Lin, R. The role of light in regulating seed dormancy and germination. J. Integr. Plant Biol. 2020, 62, 1310–1326. [Google Scholar] [CrossRef]
- Shu, K.; Liu, X.; Xie, Q.; He, Z. Two faces of one seed: Hormonal regulation of dormancy and germination. Mol. Plant 2016, 9, 34–45. [Google Scholar] [CrossRef]
- Legris, M.; Ince, Y.; Fankhauser, C. Molecular mechanisms underlying phytochrome-controlled morphogenesis in plants. Nat. Commun. 2019, 10, 5219. [Google Scholar] [CrossRef] [PubMed]
- Oh, E.; Kim, J.; Park, E.; Kim, J.; Kang, C.; Choi, G. PIL5, a phytochrome-interacting basic helix-loop-helix protein, is a key negative regulator of seed germination in Arabidopsis thaliana. Plant Cell 2004, 16, 3045–3058. [Google Scholar] [CrossRef]
- Gorshkova, D.; Pojidaeva, E. Members of the universal stress protein family are indirectly involved in gibberellin-dependent regulation of germination and post-germination growth. Russ. J. Plant Physiol. 2021, 68, 451–462. [Google Scholar] [CrossRef]
- Gorshkova, D.; Getman, I.; Voronkov, A.; Chizhova, S.; Kuznetsov, V.V.; Pojidaeva, E. The gene encoding the universal stress protein AtUSP is regulated by phytohormones and involved in seed germination of Arabidopsis thaliana. In Doklady Biochemistry and Biophysics; Springer: Berlin/Heidelberg, Germany, 2018; pp. 105–107. [Google Scholar]
- Wu, Z.; Wang, Z.; Zhang, K. Isolation and functional characterization of a glucose-6-phosphate/phosphate translocator (IbG6PPT1) from sweet potato (Ipomoea batatas (L.) Lam.). BMC Plant Biol. 2021, 21, 595. [Google Scholar] [CrossRef] [PubMed]
- Barrera-Gavira, J.; Pont, S.; Morris, J.; Hedley, P.; Stewart, D.; Taylor, M.; Hancock, R. Senescent sweetening in potato (Solanum tuberosum) tubers is associated with a reduction in plastidial glucose-6-phosphate/phosphate translocator transcripts. Postharvest Biol. Technol. 2021, 181, 111637. [Google Scholar] [CrossRef]
- Kammerer, B.; Fischer, K.; Hilpert, B.; Schubert, S.; Gutensohn, M.; Weber, A.; Flügge, U.-I. Molecular characterization of a carbon transporter in plastids from heterotrophic tissues: The glucose 6-phosphate/phosphate antiporter. Plant Cell 1998, 10, 105–117. [Google Scholar] [CrossRef]
- Mazzaferro, L.; Breccia, J. Functional and biotechnological insights into diglycosidases. Biocatal. Biotransform. 2011, 29, 103–112. [Google Scholar] [CrossRef]
- Yogeswara, I.B.A.; Maneerat, S.; Haltrich, D. Glutamate Decarboxylase from Lactic Acid Bacteria—A Key Enzyme in GABA Synthesis. Microorganisms 2020, 8, 1923. [Google Scholar] [CrossRef] [PubMed]
- Seiler, N. Chapter 33 Polyamine oxidase, properties and functions. Prog. Brain Res. 1995, 106, 333–344. [Google Scholar]
- Somani; Rakesh, R.; Rai; Priyanshu, R.; Kandpile; Pooja, S. Ornithine Decarboxylase Inhibition: A Strategy to Combat Various Diseases. Mini Rev. Med. Chem. 2018, 18, 1008–1021. [Google Scholar] [CrossRef]
- Sharma, S.; Saxena, D.; Riar, C. Analysing the effect of germination on phenolics, dietary fibres, minerals and γ-amino butyric acid contents of barnyard millet (Echinochloa frumentaceae). Food Biosci. 2016, 13, 60–68. [Google Scholar] [CrossRef]
- Kim, M.; Kwak, H.; Kim, S. Effects of germination on protein, γ-aminobutyric acid, phenolic acids, and antioxidant capacity in wheat. Molecules 2018, 23, 2244. [Google Scholar] [CrossRef]
- Boutsika, A.; Michailidis, M.; Ganopoulou, M.; Dalakouras, A.; Skodra, C.; Xanthopoulou, A.; Stamatakis, G.; Samiotaki, M.; Tanou, G.; Moysiadis, T. A wide foodomics approach coupled with metagenomics elucidates the environmental signature of potatoes. Iscience 2023, 26, 511727. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Accession | Protein Annotation | Fold | p-Value |
|---|---|---|---|
| M0ZIL5 | Peroxidase | 1.398 | 0.037185 |
| M1A1S2 | Glutathione peroxidase | 1.257 | 0.0127168 |
| M1A251 | Peroxidase | 1.236 | 0.00067654 |
| M1A2Y4 | Peroxidase | 1.300 | 0.0020564 |
| M1A7Q6 | Perakine reductase | 1.765 | 0.029841 |
| M0ZIL5 | Peroxidase | 1.398 | 0.037185 |
| M1AWZ7 | Glutathione peroxidase | 1.963 | 0.027001 |
| M1BDU1 | Superoxide dismutase | 1.533 | 4.6877 × 10−6 |
| M1BQV8 | Microsomal glutathione S-transferase 3 | 1.566 | 0.00078101 |
| M1BWS8 | Peroxisomal (S)-2-hydroxy-acid oxidase GLO1 | 0.771 | 0.00079654 |
| M0ZIL5 | Peroxidase | 1.398 | 0.037185 |
| M1CMY9 | Superoxide dismutase | 1.89 | 0.000004788 |
| M1CMY9 | Superoxide dismutase | 1.89 | 0.000004788 |
| M1D5G4 | Peroxidase | 0.797 | 0.0186962 |
| P55312 | Catalase isozyme 2 | 0.827 | 0.00132391 |
| K9MBD0 | Beta-1,3-glucanase 2 | 0.781 | 0.0183778 |
| K9MBL3 | Beta-1,3-glucanase 11 | 0.831 | 0.0153825 |
| M0ZKH6 | Glucose-6-phosphate/phosphate translocator 1 | 1.93 | 0.0095205 |
| M1AAD9 | Epidermis-specific secreted glycoprotein EP1- | 1.22 | 0.025897 |
| M1AUM5 | Gamma aminobutyrate transaminase 1 | 0.784 | 0.00056108 |
| M1B144 | Glutamate decarboxylase | 0.748 | 0.000080024 |
| P07745 | Patatin | 1.211 | 0.0187844 |
| P15478 | Patatin-T5 | 1.32 | 0.00065612 |
| P25083 | ADP, ATP carrier protein | 0.609 | 0.029918 |
| P80595 | Apyrase | 1.454 | 0.0082 |
| Q00081 | Glucose-1-phosphate adenylyl transferase | 1.204 | 0.0075754 |
| Q00775 | Granule-bound starch synthase 1 | 1.427 | 1.7086 × 10−6 |
| Q2MY36 | Patatin-15 | 1.213 | 0.0078564 |
| Q2MY44 | Patatin-07 | 1.319 | 0.0076968 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lyu, C.; Zhang, X.; Li, X.; Lu, Y.; Yuan, J.; Lu, L.; Wang, Q.; Wang, X.; Li, L. Proteomics Research Reveals the Molecular Mechanism by Which Grape Seed Oil Inhibits Tuber Sprouting in Potato. Horticulturae 2023, 9, 890. https://doi.org/10.3390/horticulturae9080890
Lyu C, Zhang X, Li X, Lu Y, Yuan J, Lu L, Wang Q, Wang X, Li L. Proteomics Research Reveals the Molecular Mechanism by Which Grape Seed Oil Inhibits Tuber Sprouting in Potato. Horticulturae. 2023; 9(8):890. https://doi.org/10.3390/horticulturae9080890
Chicago/Turabian StyleLyu, Chengcheng, Xing Zhang, Xiang Li, Yifei Lu, Jichao Yuan, Liming Lu, Qiang Wang, Xiyao Wang, and Liqin Li. 2023. "Proteomics Research Reveals the Molecular Mechanism by Which Grape Seed Oil Inhibits Tuber Sprouting in Potato" Horticulturae 9, no. 8: 890. https://doi.org/10.3390/horticulturae9080890
APA StyleLyu, C., Zhang, X., Li, X., Lu, Y., Yuan, J., Lu, L., Wang, Q., Wang, X., & Li, L. (2023). Proteomics Research Reveals the Molecular Mechanism by Which Grape Seed Oil Inhibits Tuber Sprouting in Potato. Horticulturae, 9(8), 890. https://doi.org/10.3390/horticulturae9080890