
Examining Potential Active Tempering of Adhesive Curing by Marine Mussels

Abstract
:1. Introduction
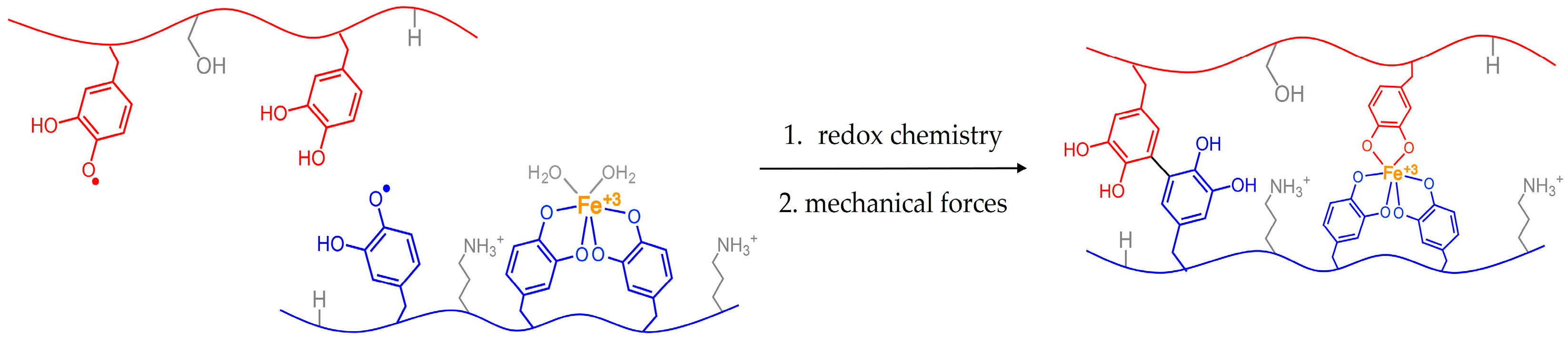
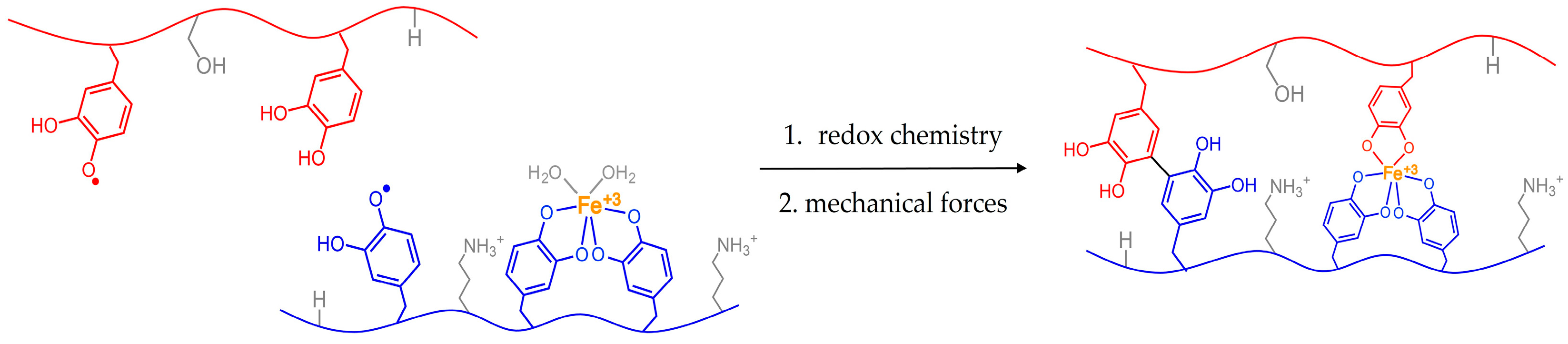
1.1 Catechols in the Sea
1.2 Animals Managing Their Glue with Mechanical Forces
1.3 Mussel Byssus and Mechanics
1.4 Animal Mechanics May Influence the Performance of Mimics
1.5 Testing the Influence of Forces from Mussels
2. Materials and Methods
2.1. Animal Handling
2.2. Adhesive Deposition
2.3. Attached versus Detached Adhesive
2.4. Adhesion Measurements
2.5. Statistics
3. Results and Discussion
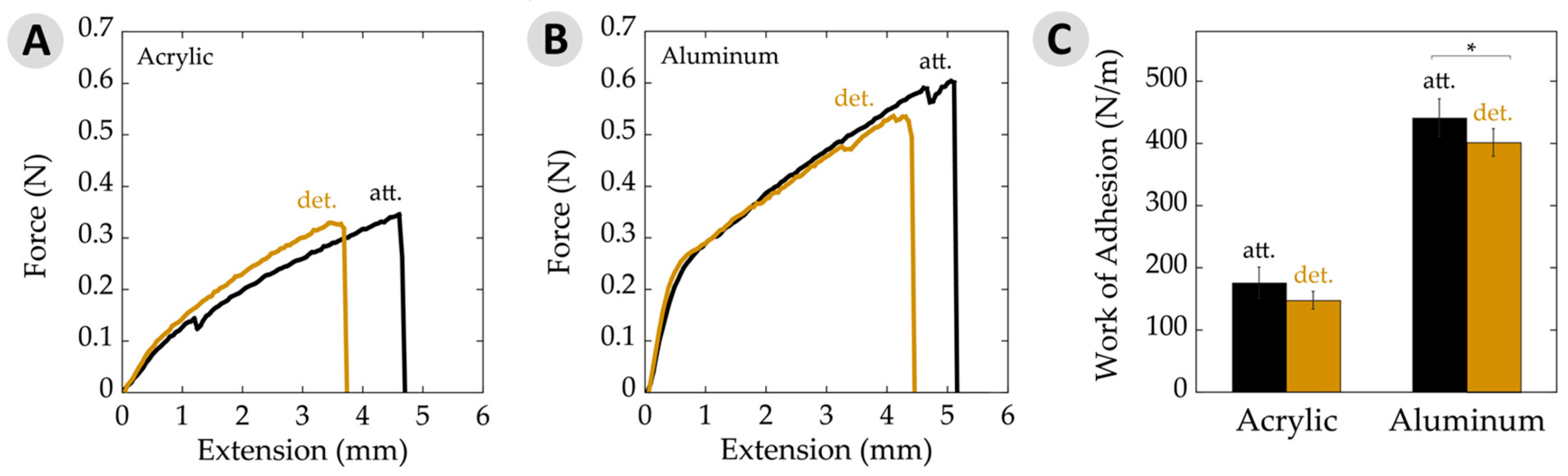
3.1. Adhesive Production and Failure Modes
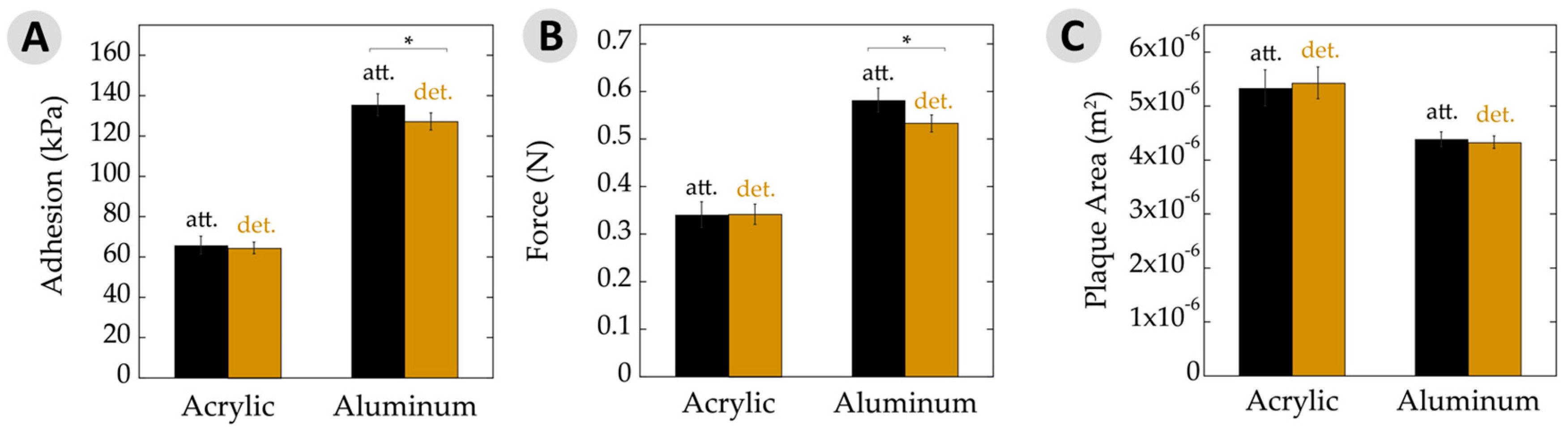
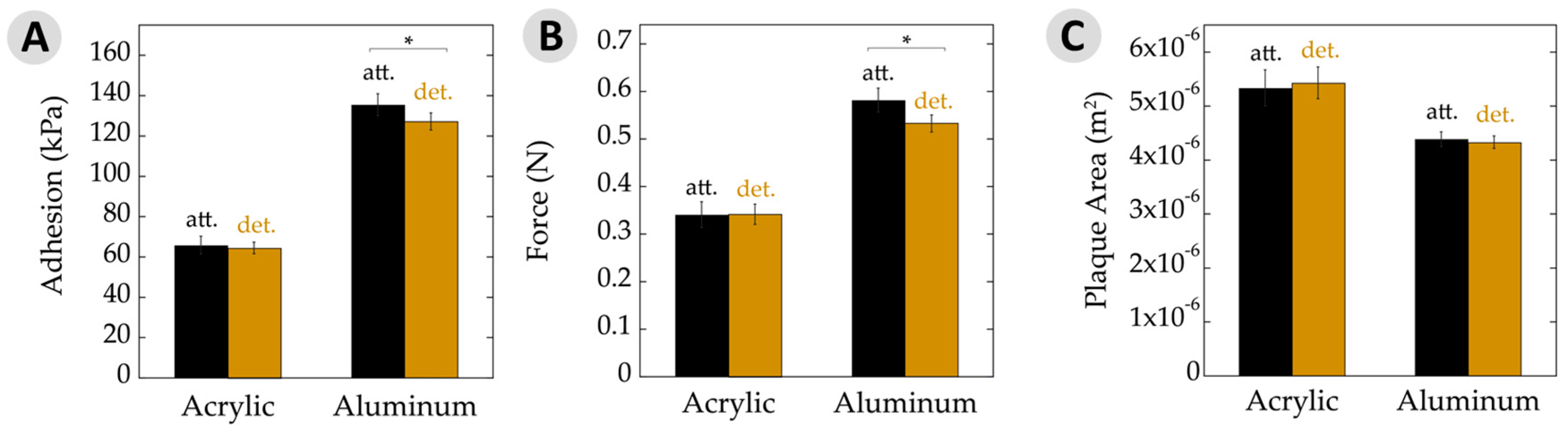
3.2. Overall Adhesion
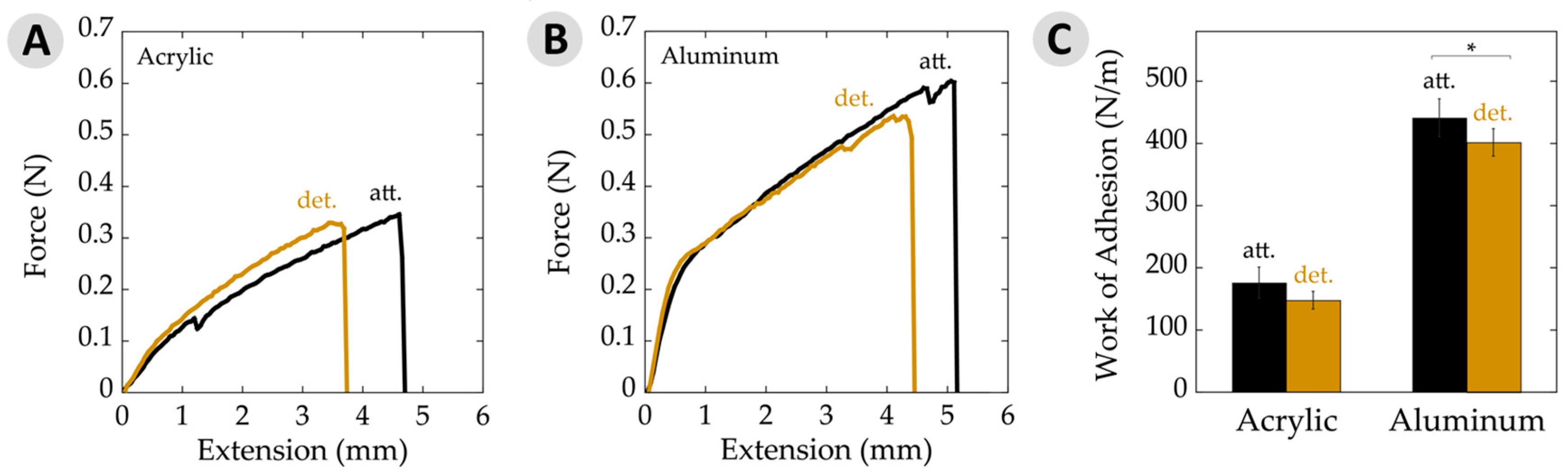
3.3. Work of Adhesion
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hagenau, A.; Suhre, M.H.; Scheibel, T.R. Nature as a blueprint for polymer material concepts: protein fiber-reinforced composites as holdfasts of mussels. Prog. Polym. Sci. 2014, 39, 1564–1583. [Google Scholar] [CrossRef]
- Lee, H.; Scherer, N.F.; Messersmith, P.B. Single-molecule mechanics of mussel adhesion. Proc. Natl. Acad. Sci. USA 2006, 103, 12999–13003. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Wei, W.; Danner, E.; Ashley, R.K.; Israelachvili, J.; Waite, J.W. Mussel protein adhesion depends on interprotein thiol-mediated redox modulation. Nat. Chem. Biol. 2011, 7, 588–590. [Google Scholar] [CrossRef] [PubMed]
- Leng, C.; Liu, Y.; Jenkins, C.; Meredith, H.; Wilker, J.J.; Chen, Z. Interfacial structure of a DOPA-inspired adhesive polymer studied by sum frequency generation vibrational spectroscopy. Langmuir 2013, 29, 6659–6664. [Google Scholar] [CrossRef] [PubMed]
- Sagert, J.; Sun, C.; Waite, J.H. Chemical subtleties of mussel and polychaete holdfasts. In Biological Adhesives; Smith, A.M., Callow, J.A., Eds.; Springer: Berlin, Germany, 2006; pp. 125–143. [Google Scholar]
- Wilker, J.J. The iron-fortified adhesive system of marine mussels. Angew. Chem. Int. Ed. 2010, 49, 8076–8078. [Google Scholar] [CrossRef] [PubMed]
- Faure, E.; Falentin-Daudré, C.; Jérôme, C.; Lyskawa, J.; Fournier, D.; Woisel, P.; Detrembleur, C. Catechols as versatile platforms in polymer chemistry. Prog. Polym. Sci. 2013, 38, 236–270. [Google Scholar] [CrossRef]
- Sedó, J.; Saiz-Poseu, J.; Busqué, F.; Ruiz-Molina, D. Catechol-based biomimetic functional materials. Adv. Mater. 2013, 25, 653–701. [Google Scholar] [CrossRef] [PubMed]
- Ye, Q.; Zhou, F.; Liu, W. Bioinspired catecholic chemistry for surface modification. Chem. Soc. Rev. 2011, 40, 4244–4258. [Google Scholar] [CrossRef] [PubMed]
- Kord Forooshani, P.; Lee, B.P. Recent approaches in designing bioadhesive materials inspired by mussel adhesive protein. J. Polym. Sci. Part A Polym. Chem. 2017, 55, 9–33. [Google Scholar] [CrossRef] [PubMed]
- Moulay, S. Polymers with dihydroxy/dialkoxybenzene moieties. Comptes Rendus Chim. 2009, 12, 577–601. [Google Scholar] [CrossRef]
- Moulay, S. DOPA/Catechol-tethered polymers: bioadhesives and biomimetic adhesive materials. Polym. Rev. 2014, 54, 436–513. [Google Scholar] [CrossRef]
- Sever, M.J.; Weisser, J.T.; Monahan, J.; Srinivasan, S.; Wilker, J.J. Metal-mediated cross-linking in the generation of a marine mussel adhesive. Angew. Chem. Int. Ed. 2004, 43, 448–450. [Google Scholar] [CrossRef] [PubMed]
- Burkett, J.R.; Wojtas, J.L.; Cloud, J.L.; Wilker, J.J. A Method for measuring the adhesion strength of marine mussels. J. Adhes. 2009, 85, 601–615. [Google Scholar] [CrossRef]
- Del Grosso, C.A.; McCarthy, T.W.; Clark, C.L.; Cloud, J.L.; Wilker, J.J. Managing redox chemistry to deter marine biological adhesion. Chem. Mater. 2016, 28, 6791–6796. [Google Scholar] [CrossRef]
- North, M.A.; Del Grosso, C.A.; Wilker, J.J. High strength underwater bonding with polymer mimics of mussel adhesive proteins. ACS Appl. Mater. Interfaces 2017, 9, 7866–7872. [Google Scholar] [CrossRef] [PubMed]
- Waite, J.H. The Formation of Mussel Bysses: Anatomy of a natural manufacturing process. In Structure, Cellular Synthesis and Assembly of Biopolymers; Case, S.T., Ed.; Springer: Berlin, Germany, 1992; pp. 55–74. ISBN 3540555498. [Google Scholar]
- Carrington, E.; Waite, J.H.; Sara, G.; Sebens, K.P. Mussels as a model system for integrative ecomechanics. Ann. Rev. Mar. Sci. 2015, 7, 443–469. [Google Scholar] [CrossRef] [PubMed]
- Bell, E.C.; Gosline, J.M. Strategies for life in flow: Tenacity, morphometry, and probability of dislodgment of two Mytilus species. Mar. Ecol. Prog. Ser. 1997, 159, 197–208. [Google Scholar] [CrossRef]
- Dolmer, P.; Svane, I. Attachment and orientation of Mytilus edulis L. in flowing water. Ophelia 1994, 40, 63–74. [Google Scholar] [CrossRef]
- Qin, X.-X.; Coyne, K.J.; Waite, J.H. Tough tendons mussel byssus has collagen with silk-like domains. J. Biol. Chem. 1997, 272, 32623–32627. [Google Scholar] [CrossRef] [PubMed]
- Filippidi, E.; DeMartini, D.G.; de Molina, P.M.; Danner, E.W.; Kim, J.; Helgeson, M.E.; Waite, J.H.; Valentine, M.T. The microscopic network structure of mussel (Mytilus) adhesive plaques. J. R. Soc. Interface 2015, 12, 20150827. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, M.J.; George, M.N.; Carrington, E. Mussel byssus attachment weakened by ocean acidification. Nat. Clim. Chang. 2013, 3, 587–590. [Google Scholar] [CrossRef]
- Carrington, E.; Moeser, G.M.; Thompson, S.B.; Coutts, L.C.; Craig, C.A. Mussel attachment on rocky shores: The effect of flow on byssus production. Integr. Comp. Biol. 2008, 48, 801–807. [Google Scholar] [CrossRef] [PubMed]
- Moeser, G.M.; Leba, H.; Carrington, E. Seasonal influence of wave action on thread production in Mytilus edulis. J. Exp. Biol. 2006, 209, 881–890. [Google Scholar] [CrossRef] [PubMed]
- Carrington, E. Seasonal variation in the attachment strength of blue mussels: Causes and consequences. Limnol. Oceanogr. 2002, 47, 1723–1733. [Google Scholar] [CrossRef]
- Sun, C.; Vaccaro, E.; Waite, J.H. Oxidative stress and the mechanical properties of naturally occurring chimeric collagen-containing fibers. Biophys. J. 2001, 81, 3590–3595. [Google Scholar] [CrossRef]
- Aldred, N.; Wills, T.; Williams, D.N.; Clare, A.S. Tensile and dynamic mechanical analysis of the distal portion of mussel (Mytilus edulis) byssal threads. J. R. Soc. Interface 2007, 4, 1159–1167. [Google Scholar] [CrossRef] [PubMed]
- Lazaris, A.; Arcidiacono, S.; Huang, Y.; Zhou, J.-F.; Duguay, F.; Chretien, N.; Welsh, E.A.; Soares, J.W.; Karatzas, C.N. Spider silk fibers spun from soluble recombinant silk produced in mammalian cells. Science 2002, 295, 472LP–476LP. [Google Scholar] [CrossRef] [PubMed]
- Rising, A.; Johansson, J. Toward spinning artificial spider silk. Nat. Chem. Biol. 2015, 11, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Rammensee, S.; Slotta, U.; Scheibel, T.; Bausch, A.R. Assembly mechanism of recombinant spider silk proteins. Proc. Natl. Acad. Sci. USA 2008, 105, 6590–6595. [Google Scholar] [CrossRef] [PubMed]
- Becker, N.; Oroudjev, E.; Mutz, S.; Cleveland, J.P.; Hansma, P.K.; Hayashi, C.Y.; Makarov, D.E.; Hansma, H.G. Molecular nanosprings in spider capture-silk threads. Nat. Mater. 2003, 2, 278–283. [Google Scholar] [CrossRef] [PubMed]
- Römer, L.; Scheibel, T. The elaborate structure of spider silk: Structure and function of a natural high performance fiber. Prion 2008, 2, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Holland, G.P.; Jenkins, J.E.; Creager, M.S.; Lewis, R.V.; Yarger, J.L. Quantifying the fraction of glycine and alanine in β-sheet and helical conformations in spider dragline silk using solid-state NMR. Chem. Comm. 2008, 5568–5570. [Google Scholar] [CrossRef] [PubMed]
- Harrington, M.J.; Waite, J.H. Holdfast heroics: Comparing the molecular and mechanical properties of Mytilus californianus byssal threads. J. Exp. Biol. 2007, 210, 4307–4318. [Google Scholar] [CrossRef] [PubMed]
- Priemel, T.; Degtyar, E.; Dean, M.N.; Harrington, M.J. Rapid self-assembly of complex biomolecular architectures during mussel byssus biofabrication. Nat. Commun. 2017, 8, 14539. [Google Scholar] [CrossRef] [PubMed]
- Harrington, M.J.; Masic, A.; Holten-Andersen, N.; Waite, J.H.; Fratzl, P. Iron-clad fibers: A metal-based biological strategy for hard flexible coatings. Science 2010, 328, 216–220. [Google Scholar] [CrossRef] [PubMed]
- Kinloch, A. Adhesion and Adhesives: Science and Technology; Springer Science & Business Media: Berlin, Germany, 2012; ISBN 9401577641. [Google Scholar]
- Desmond, K.W.; Zacchia, N.A.; Waite, J.H.; Valentine, M.T. Dynamics of mussel plaque detachment. Soft Matter 2015, 11, 6832–6839. [Google Scholar] [CrossRef] [PubMed]
- Abràmoff, M.D.; Magalhães, P.J.; Ram, S.J. Image processing with ImageJ. Biophotonics Int. 2004, 11, 36–42. [Google Scholar]
- Gutowski, W. Thermodynamics of Adhesion. In Fundamentals of Adhesion; Lee, L.-H., Ed.; Springer: Boston, MA, USA, 1991; pp. 87–135. ISBN 978-1-4899-2073-7. [Google Scholar]
- Pocius, A.V.; Dillard, D.A. Adhesion Science and Engineering: Surfaces, Chemistry and Applications; Elsevier: Amsterdam, The Netherlands, 2002; ISBN 0080525989. [Google Scholar]
- Young, G.A.; Crisp, D.J. Marine animals and adhesion. Adhesion 1982, 6, 19–39. [Google Scholar]
- Crisp, D.J.; Walker, G.; Young, G.A.; Yule, A.B. Adhesion and substrate choice in mussels and barnacles. J. Colloid Interface Sci. 1985, 104, 40–50. [Google Scholar] [CrossRef]
- Aldred, N.; Ista, L.K.; Callow, M.E.; Callow, J.A.; Lopez, G.P.; Clare, A.S. Mussel (Mytilus edulis) byssus deposition in response to variations in surface wettability. J. R. Soc. Interface 2006, 3, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Yang, H.; Liu, F.; Chen, T.; Hu, G.; Guo, D.; Hou, Q.; Wu, X.; Su, Y.; Wang, J. Molecular interactions between DOPA and surfaces with different functional groups: A chemical force microscopy study. RSC Adv. 2017, 7, 32518–32527. [Google Scholar] [CrossRef]
- Qin, Z.; Buehler, M.J. Molecular mechanics of mussel adhesion proteins. J. Mech. Phys. Solids 2014, 62, 19–30. [Google Scholar] [CrossRef]
- Vaccaro, E.; Waite, J.H. Yield and post-yield behavior of mussel byssal thread: A self-healing biomolecular materials. Biomacromolecules 2001, 2, 906–911. [Google Scholar] [CrossRef] [PubMed]
- Hamada, N.A.; Roman, V.A.; Howell, S.M.; Wilker, J.J. Purdue University, Lafayette, IN, USA. Potential differences in adhesive properties as a function of failure modes. 2017. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Acrylic | Aluminum | |||
|---|---|---|---|---|
| Detached | Attached | Detached | Attached | |
| Number of plaques | 231 | 134 | 314 | 244 |
| Average adhesion (kPa) | 65 ± 3 | 66 ± 4 | 127 ± 4 | 135 ± 5 |
| Force of detachment (N) | 0.34 ± 0.02 | 0.34 ± 0.02 | 0.53 ± 0.02 | 0.58 ± 0.03 |
| Plaque area (×10−6 m2) | 5.4 ± 0.3 | 5.3 ± 0.3 | 4.3 ± 0.1 | 4.4 ± 0.2 |
| Work of adhesion (N/m) | 148 ± 14 | 176 ± 25 | 401 ± 22 | 441 ± 30 |
| Adhesive failure % | 90.0 | 84.2 | 26.8 | 23.4 |
| Cohesive failure % | 4.8 | 7.5 | 57.3 | 58.6 |
| Thread break failure % | 3.5 | 3.0 | 10.8 | 8.6 |
| Thread–plaque interface failure % | 1.7 | 5.3 | 5.1 | 9.4 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hamada, N.A.; Roman, V.A.; Howell, S.M.; Wilker, J.J. Examining Potential Active Tempering of Adhesive Curing by Marine Mussels. Biomimetics 2017, 2, 16. https://doi.org/10.3390/biomimetics2030016
Hamada NA, Roman VA, Howell SM, Wilker JJ. Examining Potential Active Tempering of Adhesive Curing by Marine Mussels. Biomimetics. 2017; 2(3):16. https://doi.org/10.3390/biomimetics2030016
Chicago/Turabian StyleHamada, Natalie A., Victor A. Roman, Steven M. Howell, and Jonathan J. Wilker. 2017. "Examining Potential Active Tempering of Adhesive Curing by Marine Mussels" Biomimetics 2, no. 3: 16. https://doi.org/10.3390/biomimetics2030016
APA StyleHamada, N. A., Roman, V. A., Howell, S. M., & Wilker, J. J. (2017). Examining Potential Active Tempering of Adhesive Curing by Marine Mussels. Biomimetics, 2(3), 16. https://doi.org/10.3390/biomimetics2030016