
Monitoring Red Sea Bream Scale Fluorescence as a Freshness Indicator

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Red Sea Bream
2.2. K Value Measurement
2.3. Scales Spectra Acquisition
2.3.1. Front-Face Fluorescence Spectra Measurement
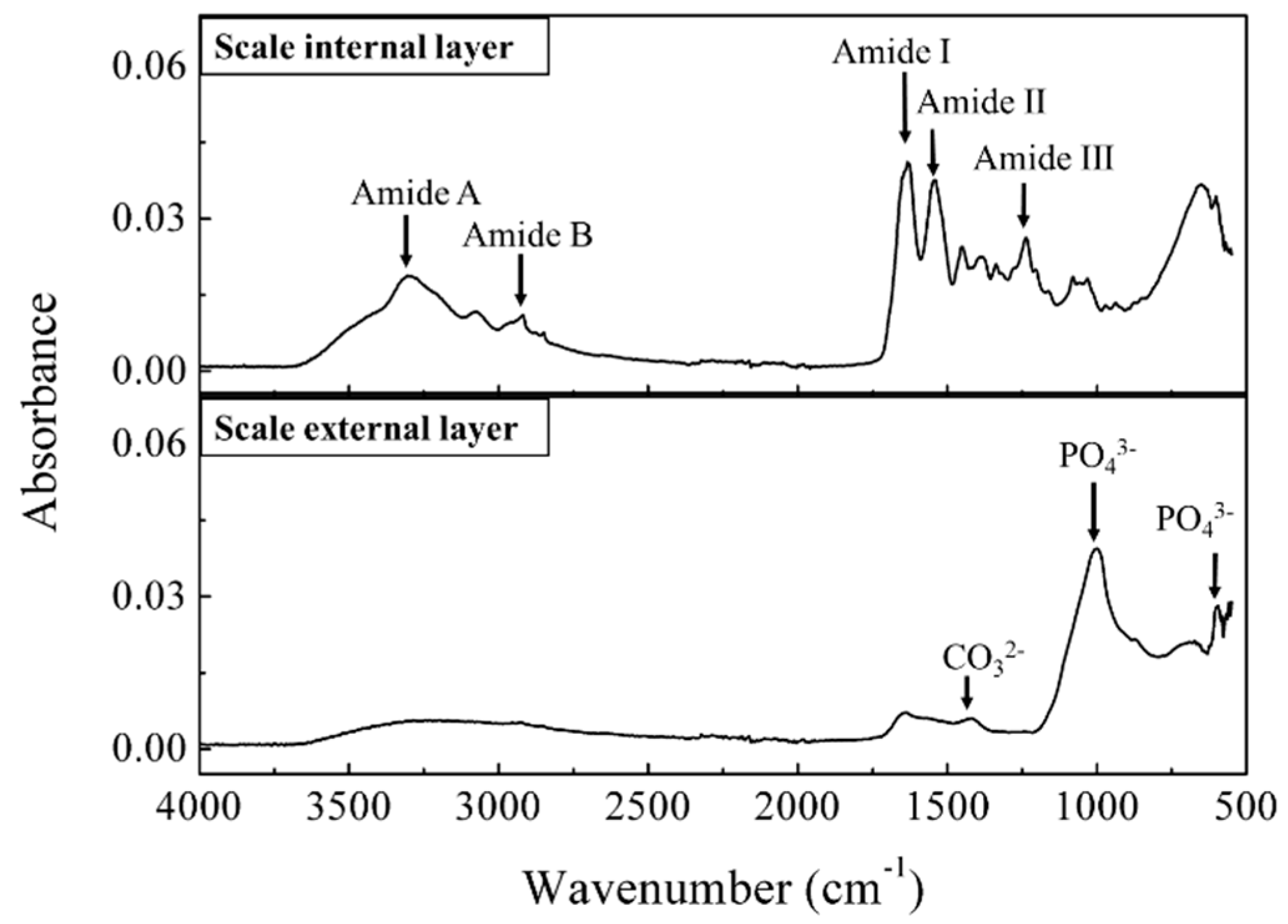
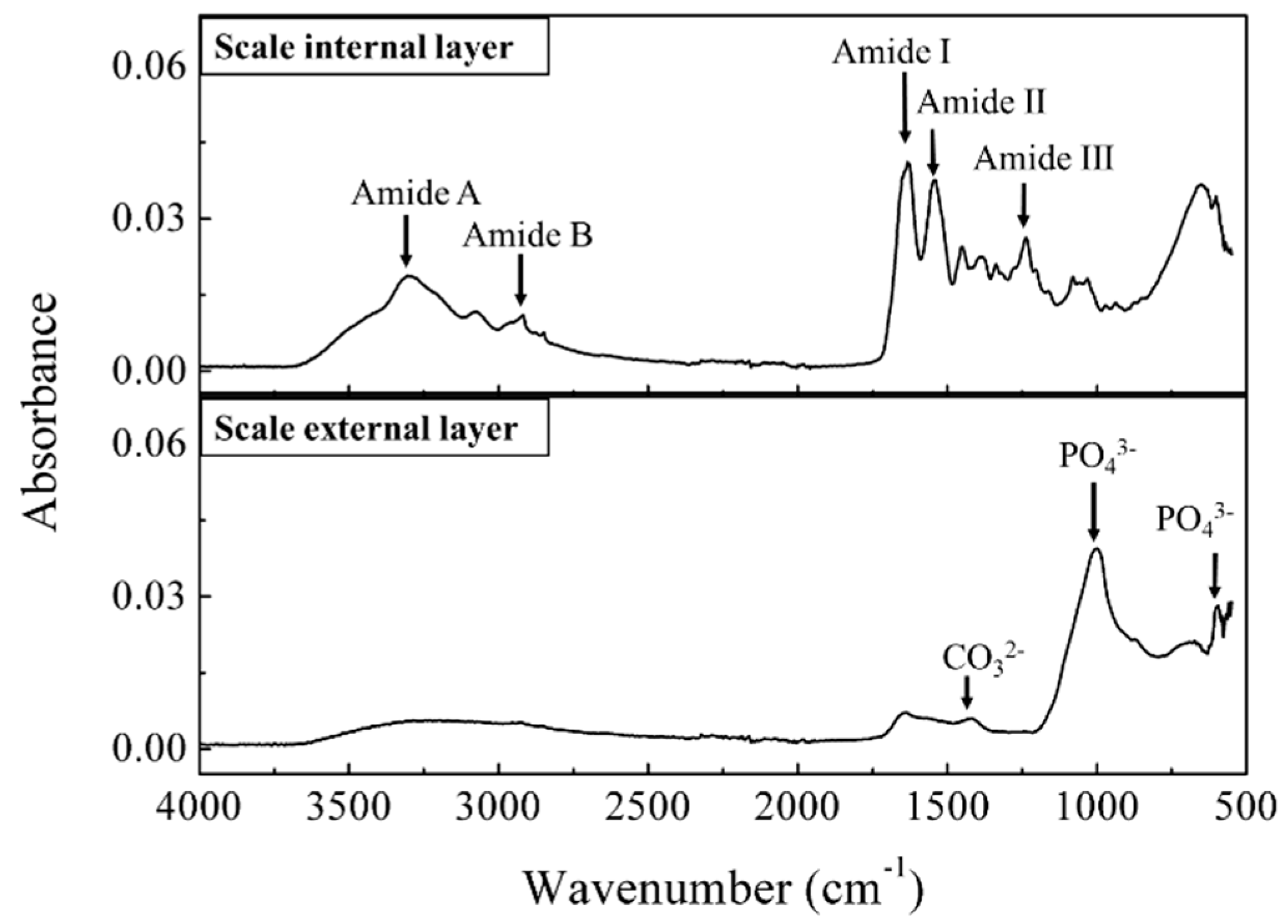
2.3.2. Infrared Spectra Measurement for Compound Identification
2.4. Scanning Electron Micrograph Image Acquisition
3. Results and Discussion
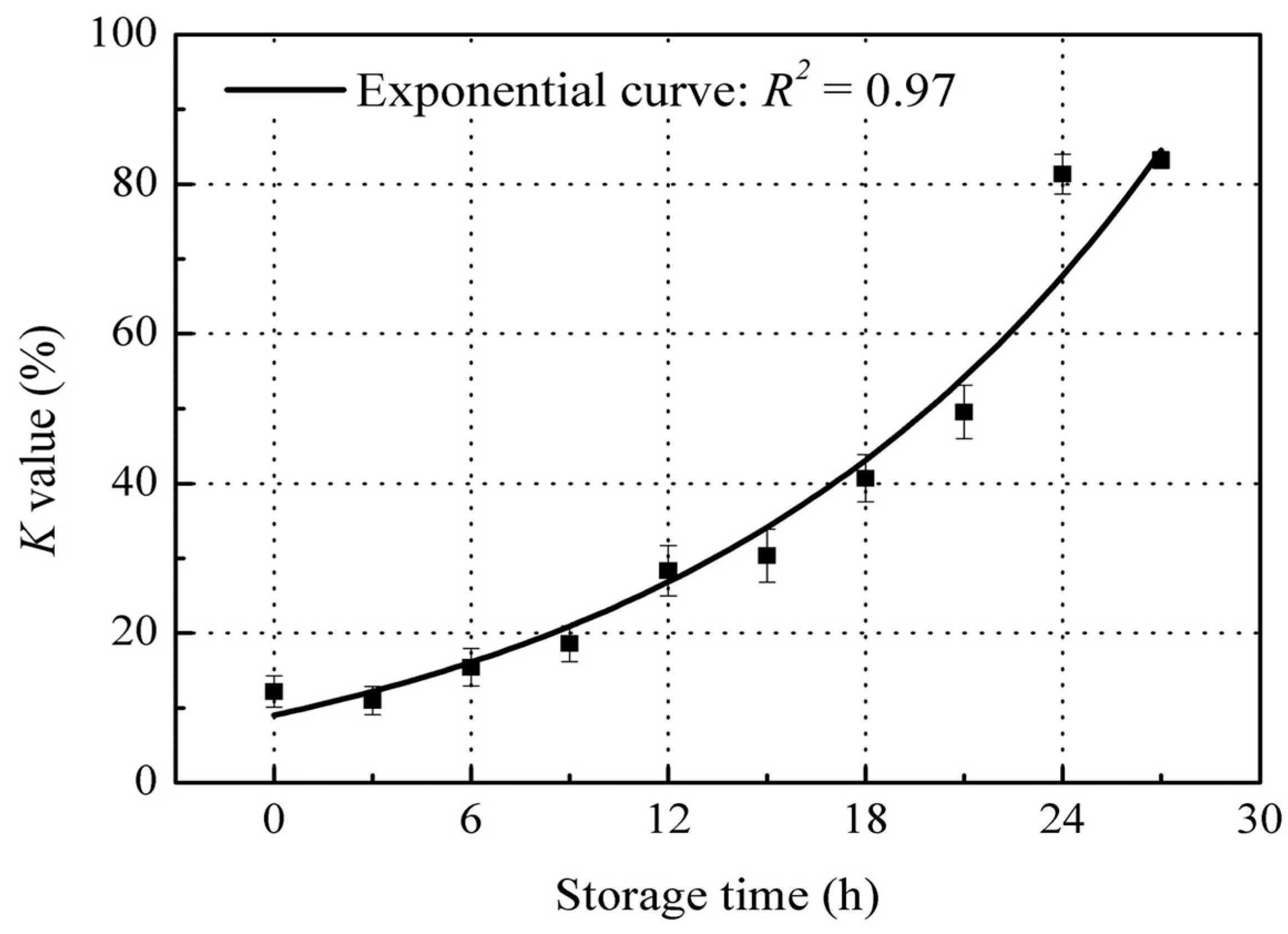
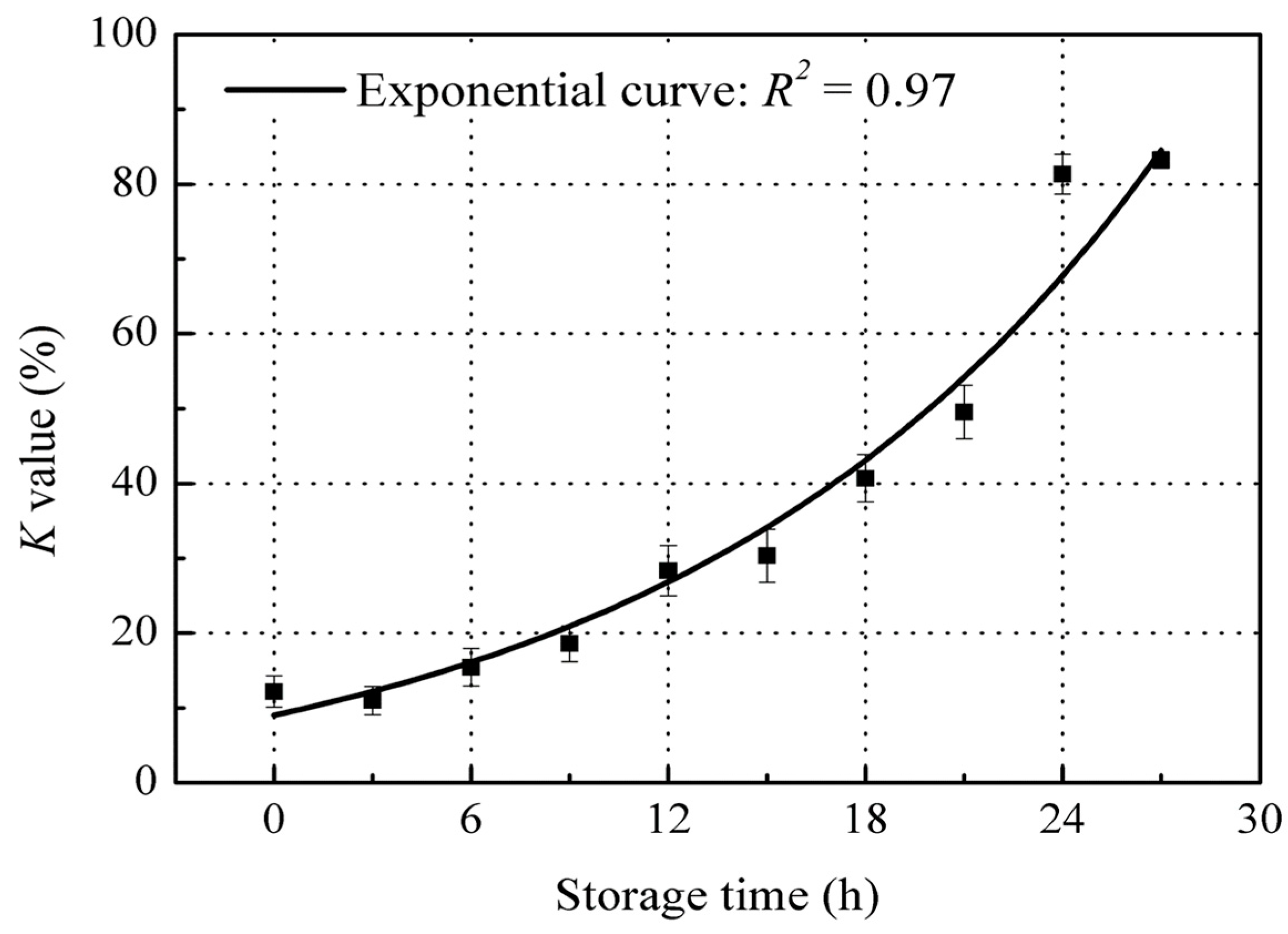
3.1. Time-Dependent Changes of K Value
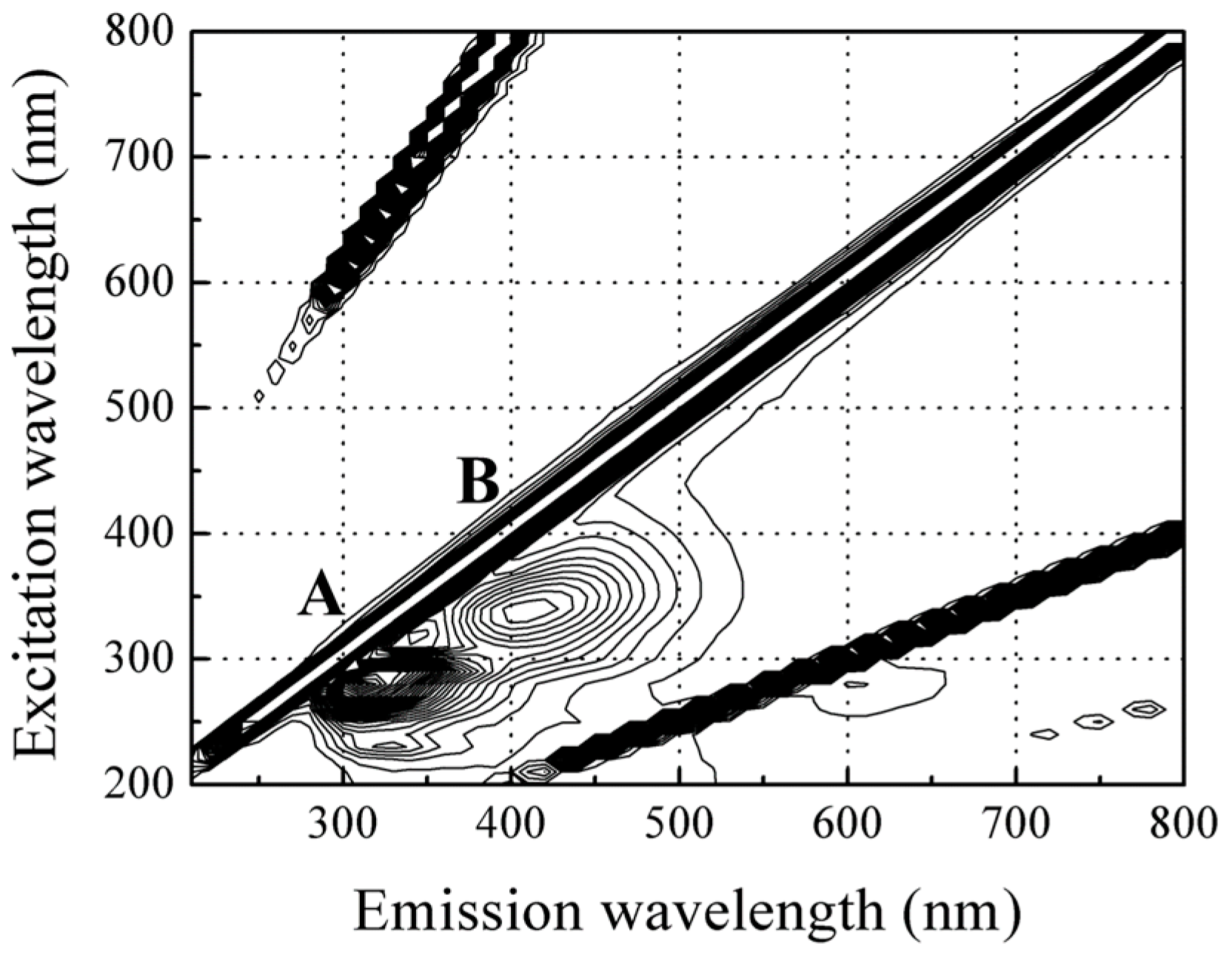
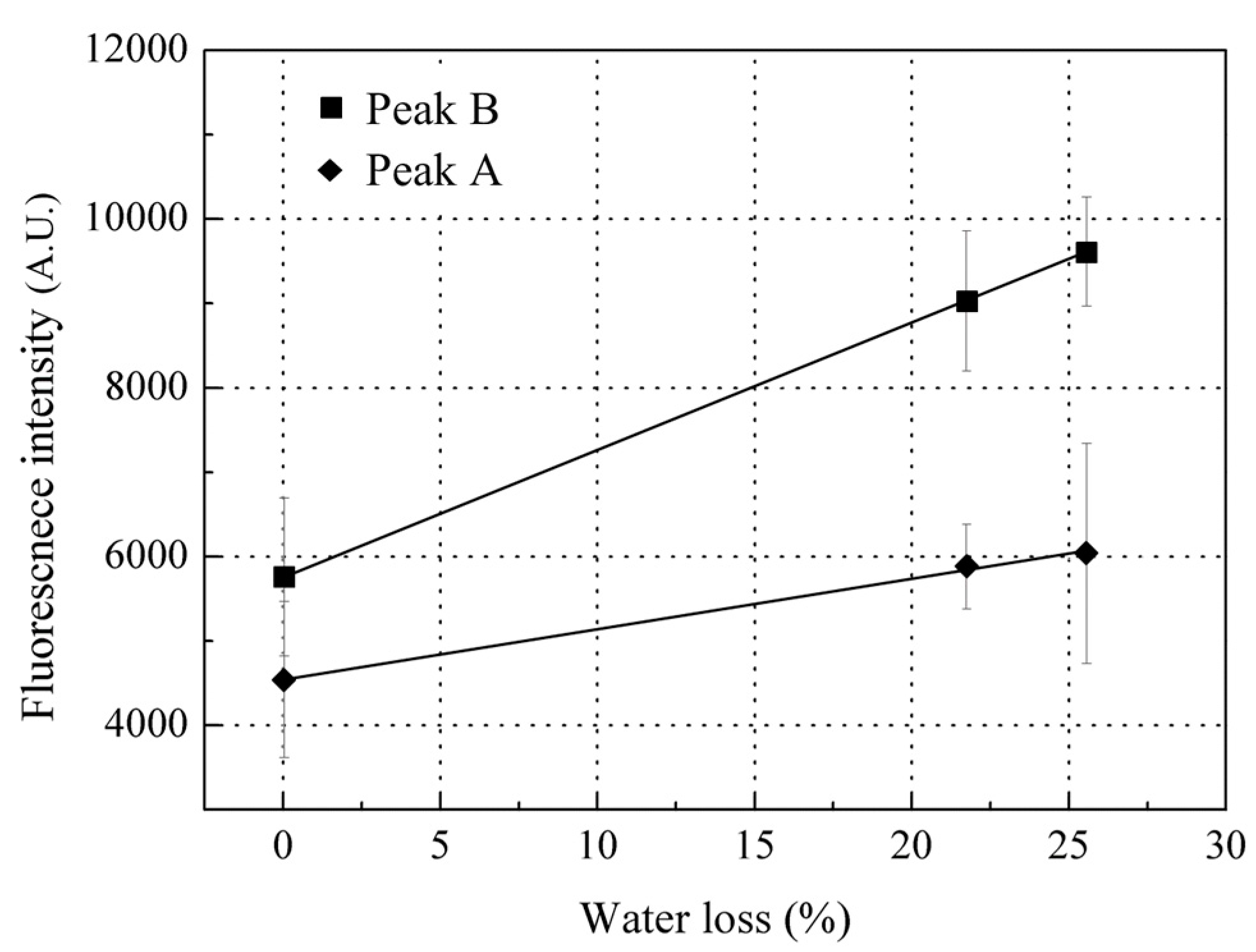
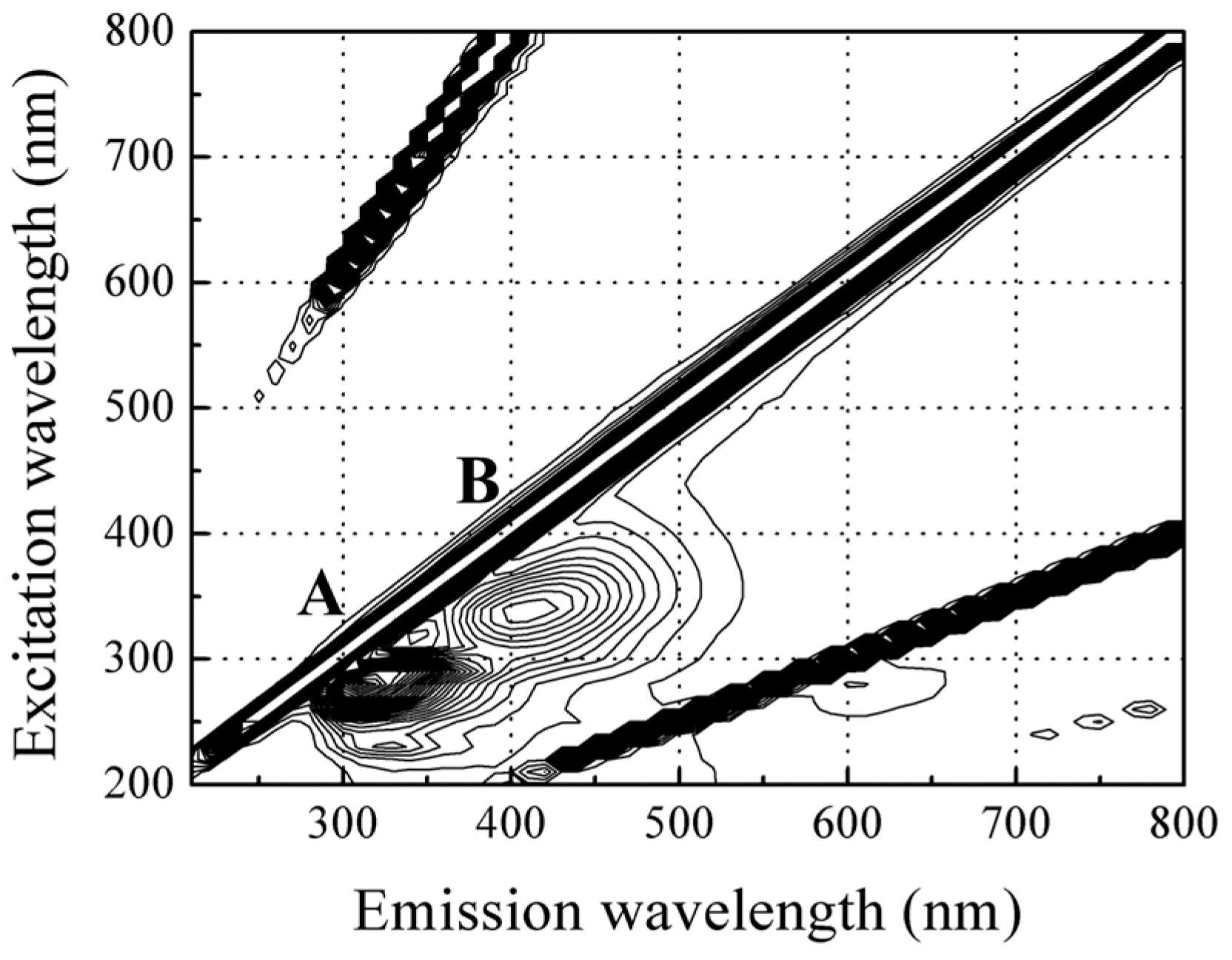
3.2. Fluorescence Spectroscopic Properties of Red Sea Bream Scales
3.2.1. Identification of the Major Compound Contributing to Peak A (Ex/Em: 280/310 nm)
3.2.2. Identification of the Major Compound Contributing to Peak B (Ex/Em: 340/420 nm)
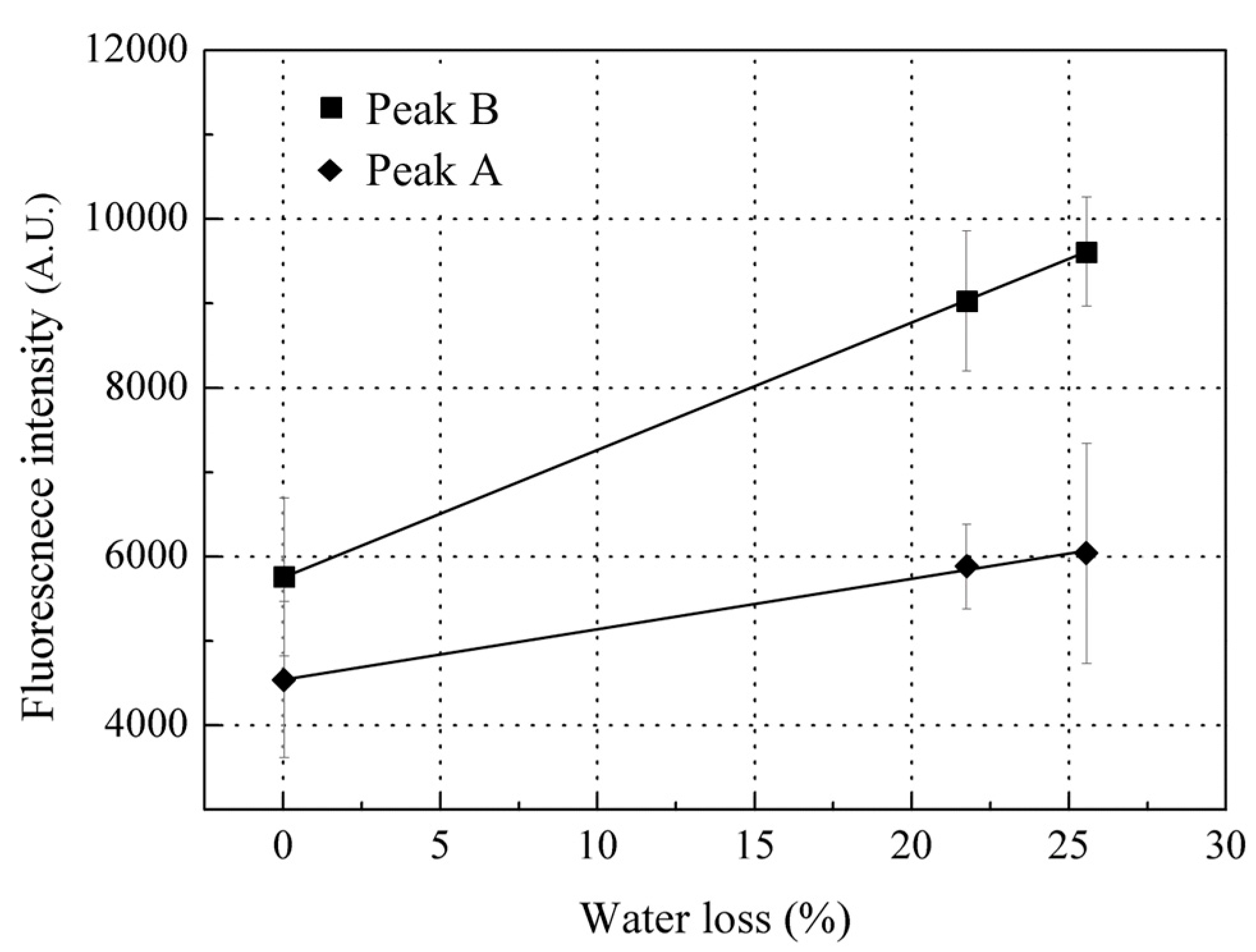
3.3. Time-Dependent Changes in Fluorescence Intensity of Scales
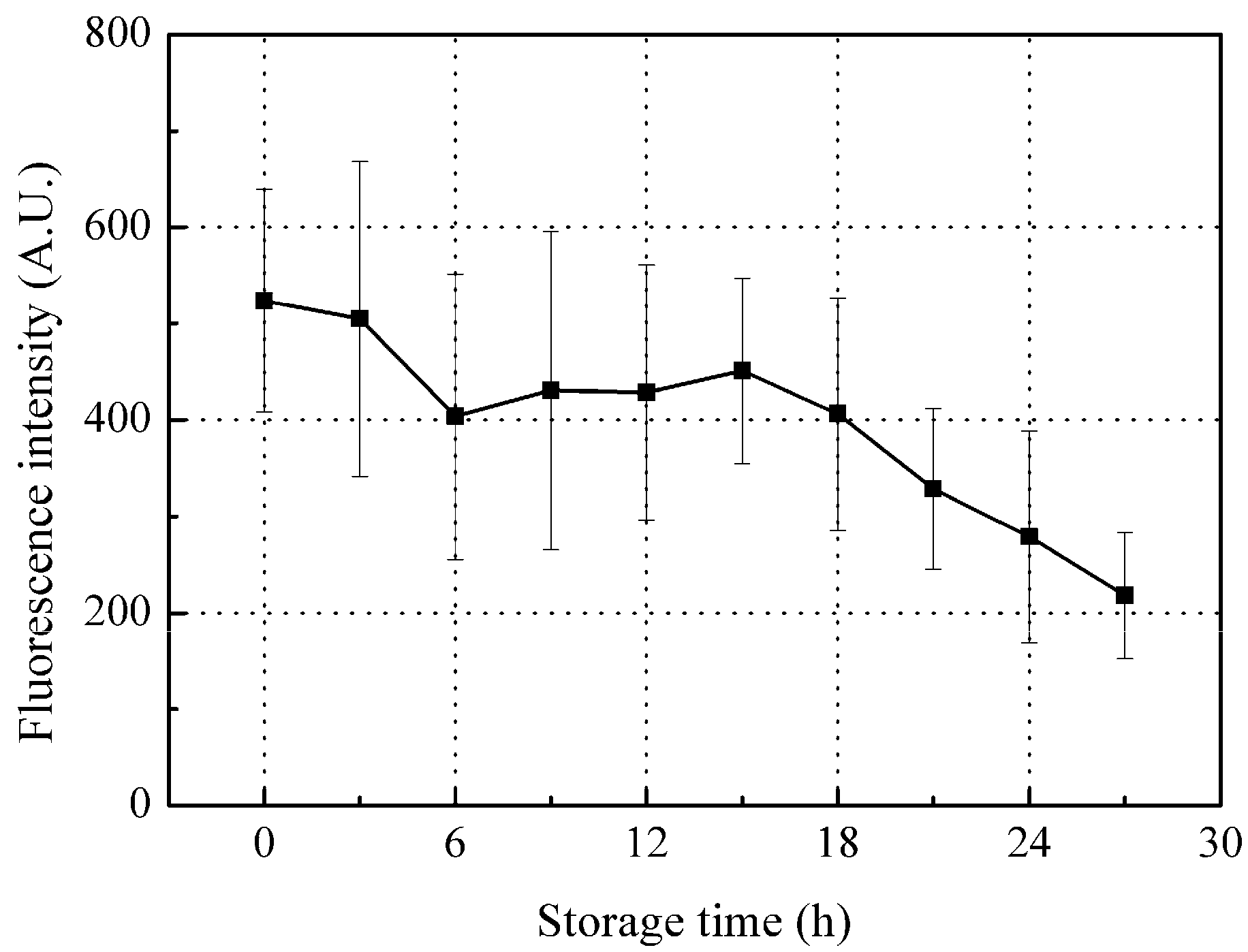
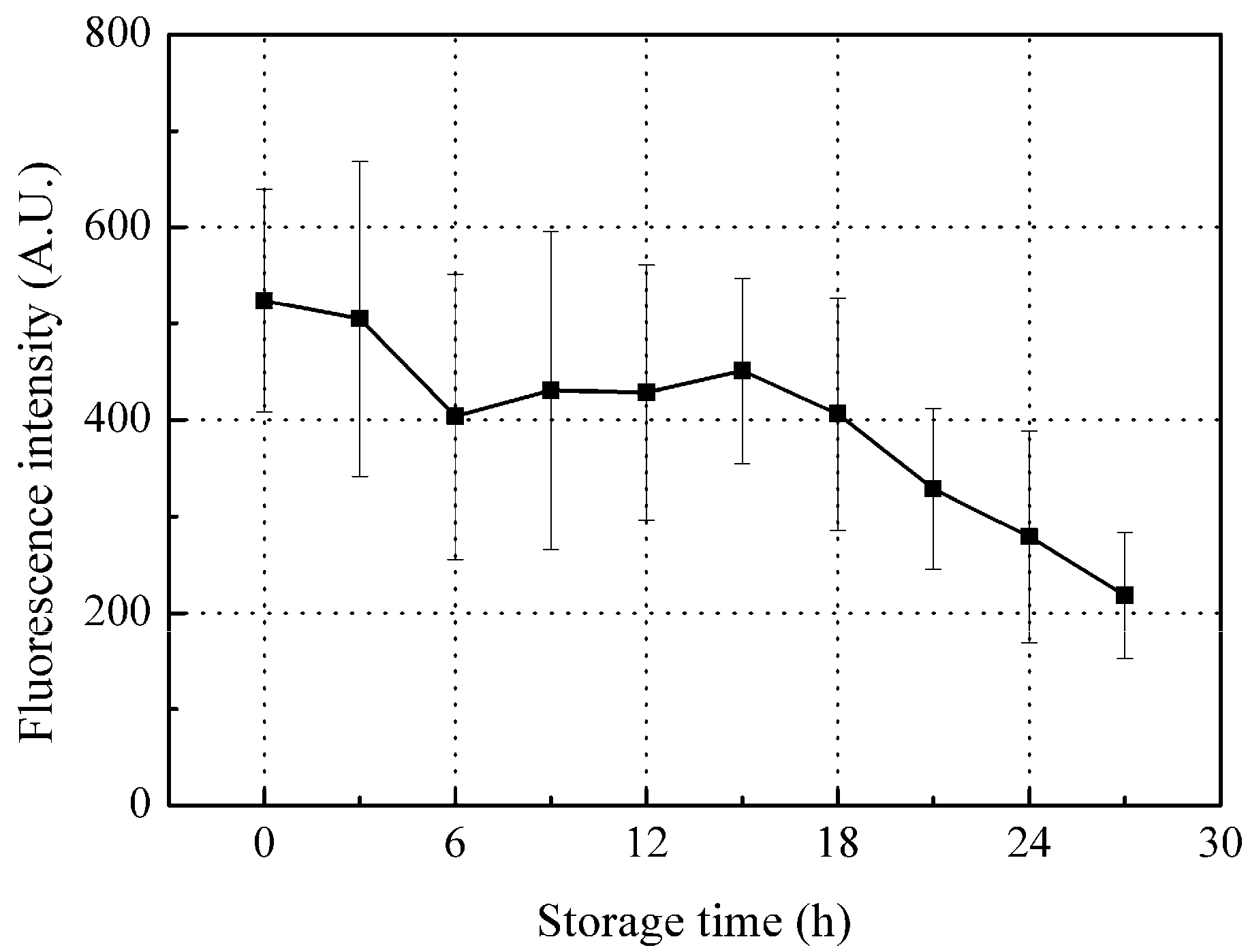
3.3.1. Time-Dependent Changes in Fluorescence Intensity of Peak A (Ex/Em: 280/310 nm)
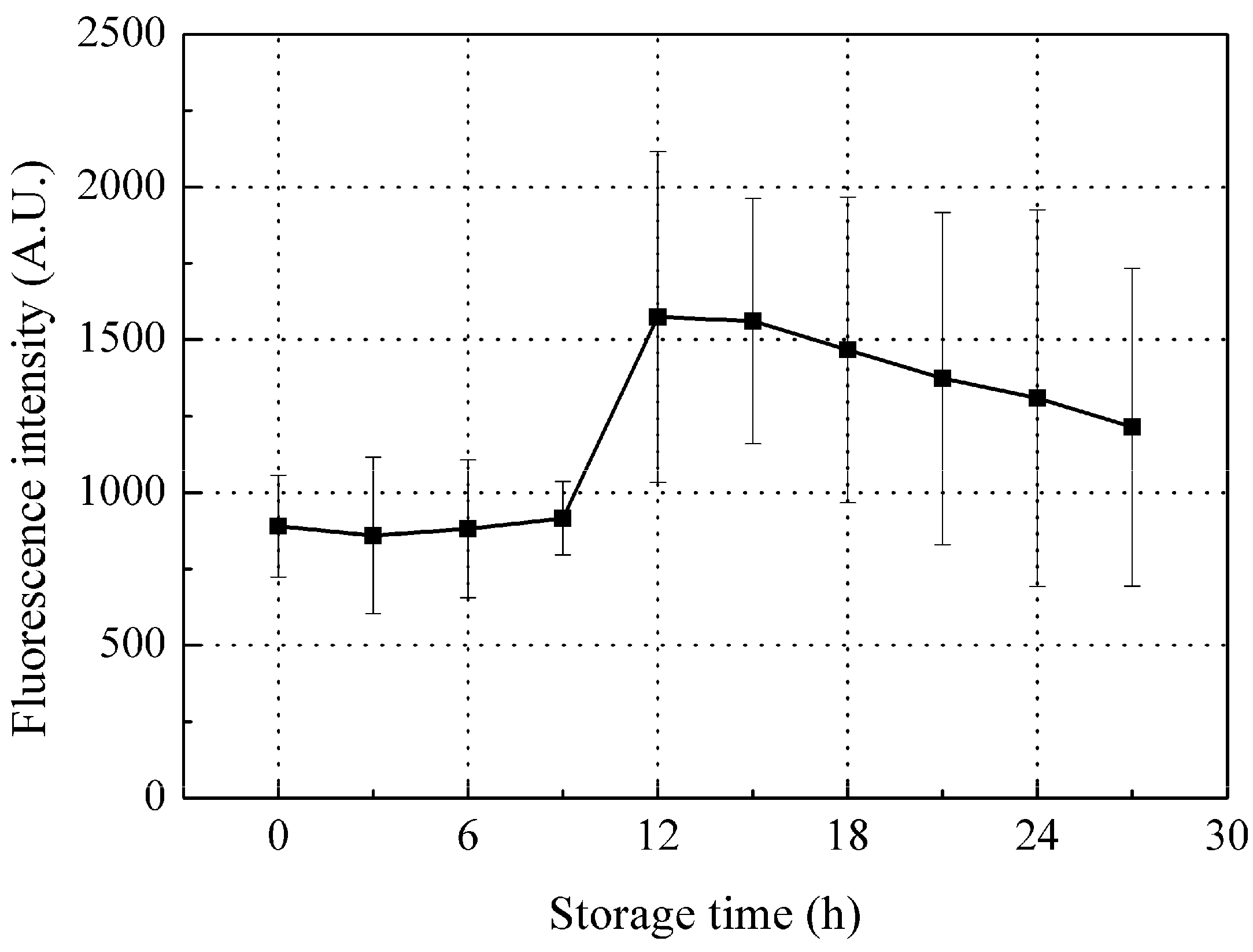
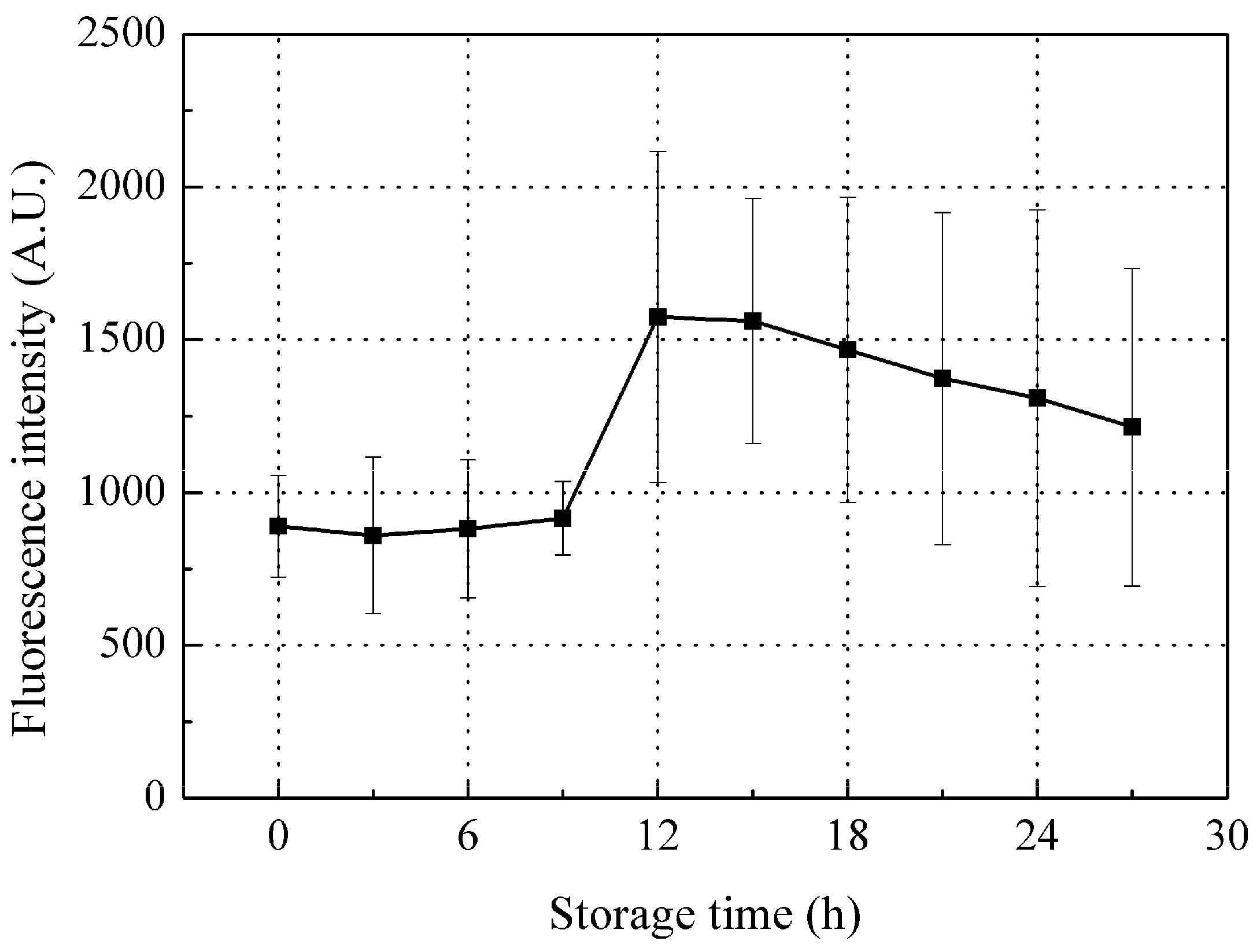
3.3.2. Time-Dependent Changes in Fluorescence Intensity of Peak B (Ex/Em: 340/420 nm)
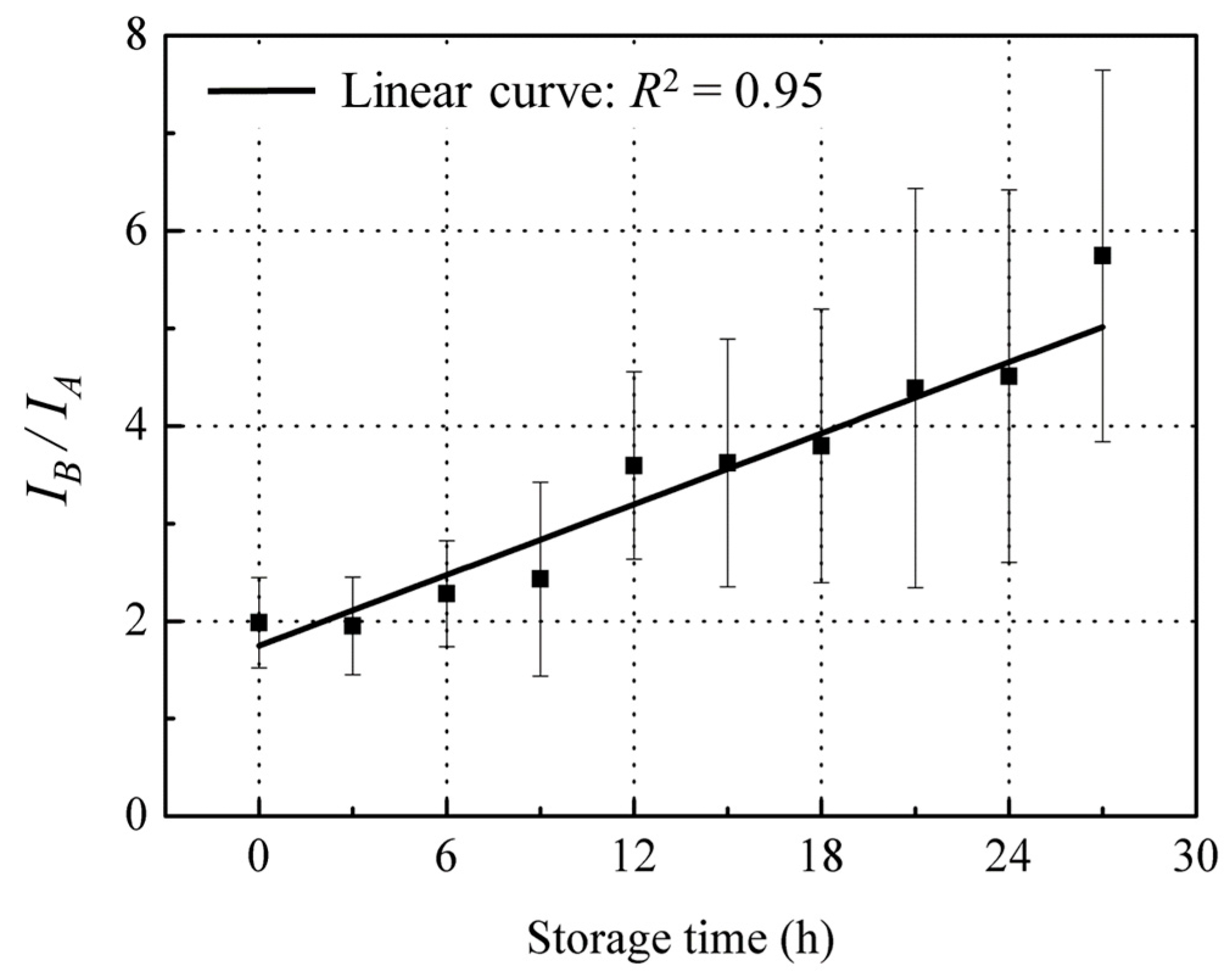
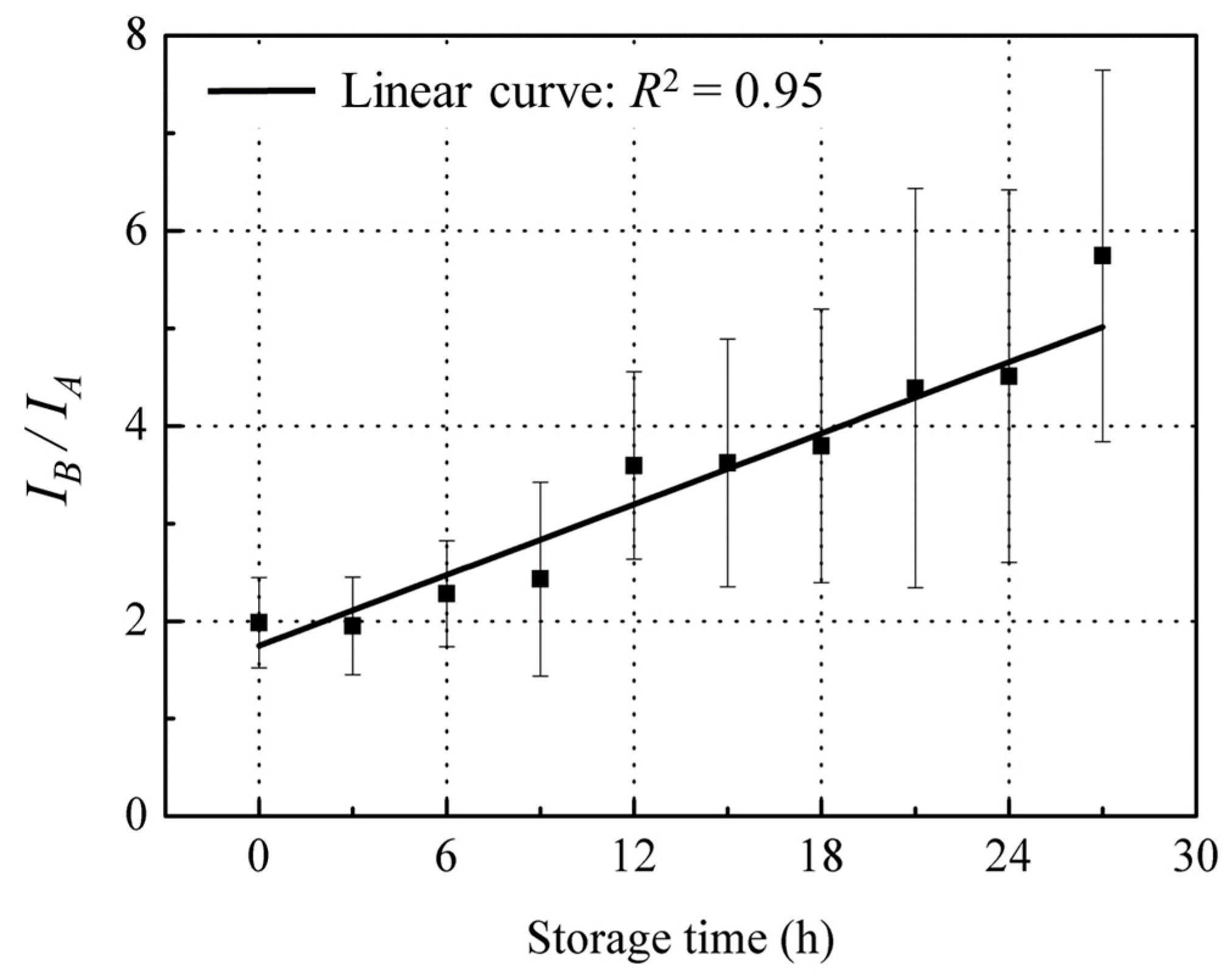
3.3.3. Ratio of IB/IA Related with Freshness
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Özogul, Y.; Özyurt, G.; Özogul, F.; Kuley, E.; Polat, A. Freshness assessment of European eel (Anguilla anguilla) by sensory, chemical and microbiological methods. Food Chem. 2005, 92, 745–751. [Google Scholar] [CrossRef]
- Arulkumar, A.; Paramasivam, S. Sensory quality and biochemical changes in deep queen fish (Scomberoides tala) during ice storage. Asian J. Microbiol. Biotechnol. Environ. Sci. 2015, 17, 117–125. [Google Scholar]
- Chytiri, S.; Chouliara, I.; Savvaidis, I.N.; Kontominas, M.G. Microbiological, chemical and sensory assessment of iced whole and filleted aquacultured rainbow trout. Food Microbiol. 2004, 21, 157–165. [Google Scholar] [CrossRef]
- Ocaño-Higuera, V.M.; Maeda-Martínez, A.N.; Marquez-Ríos, E.; Canizales-Rodríguez, D.F.; Castillo-Yáñez, F.J.; Ruíz-Bustos, E.; Graciano-Erdugo, A.Z.; Plascencia-Jatomea, M. Freshness assessment of ray fish stored in ice by biochemical, chemical and physical methods. Food Chem. 2011, 125, 49–54. [Google Scholar] [CrossRef]
- Itoh, D.; Koyachi, E.; Yokokawa, M.; Murata, Y.; Murata, M.; Suzuki, H. Microdevice for on-site fish freshness checking based on K-value measurement. Anal. Chem. 2013, 85, 10962–10968. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.H.; Dai, Q.; Sun, D.W.; Zeng, X.A.; Liu, D.; Pu, H.B. Applications of non-destructive spectroscopic techniques for fish quality and safety evaluation and inspection. Trends Food Sci. Technol. 2013, 34, 18–31. [Google Scholar] [CrossRef]
- Murakoshi, T.; Masuda, T.; Utsumi, K.; Tsubota, K.; Wada, Y. Glossiness and Perishable Food Quality: Visual Freshness Judgment of Fish Eyes Based on Luminance Distribution. PLoS ONE 2013, 8, e58994. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Xin, J.; Zhao, J. A novel technique for rapid evaluation of fish freshness using colorimetric sensor array. J. Food Eng. 2011, 105, 632–637. [Google Scholar] [CrossRef]
- Han, F.; Huang, X.; Teye, E.; Gu, H.; Dai, H.; Yao, L. A nondestructive method for fish freshness determination with electronic tongue combined with linear and non-linear multivariate algorithms. Czech J. Food Sci. 2014, 32, 532–537. [Google Scholar]
- Karoui, R.; Blecker, C. Fluorescence spectroscopy measurement for quality assessment of food systems—A review. Food Bioprocess Technol. 2011, 4, 364–386. [Google Scholar] [CrossRef]
- Airado-Rodríguez, D.; Skaret, J.; Wold, J.P. Assessment of the quality attributes of cod caviar paste by means of front-face fluorescence spectroscopy. J. Agric. Food Chem. 2010, 58, 5276–5285. [Google Scholar] [CrossRef] [PubMed]
- Lakhal, L.; Acha, V.; Aussenac, T. PARAFAC analysis of front-face fluorescence data: Absorption and scattering effects assessed by means of Monte Carlo simulations. Chemom. Intell. Lab. Syst. 2012, 116, 112–122. [Google Scholar] [CrossRef]
- Christensen, J.; Nørgaard, L.; Bro, R.; Engelsen, S.B. Multivariate autofluorescence of intact food systems. Chem. Rev. 2006, 106, 1979–1994. [Google Scholar] [CrossRef] [PubMed]
- Mondal, S.; Mahata, S.; Kundu, S.; Mondal, B. Processing of natural resourced hydroxyapatite ceramics from fish scale. Adv. Appl. Ceram. 2010, 109, 234–239. [Google Scholar] [CrossRef]
- Ghozlen, N.B.; Cerovic, Z.G.; Germain, C.; Toutain, S.; Latouche, G. Non-destructive optical monitoring of grape maturation by proximal sensing. Sensors 2010, 10, 10040–10068. [Google Scholar] [CrossRef] [PubMed]
- Nishino, K.; Nakamura, K.; Tsuta, M.; Yoshimura, M.; Sugiyama, J.; Nakauchi, S. Optimization of excitation–emission band-pass filter for visualization of viable bacteria distribution on the surface of pork meat. Opt. Express 2013, 21, 12579–12591. [Google Scholar] [CrossRef] [PubMed]
- Johal, M.S.; Esmaeili, H.R.; Sharma, M.L. Scale structure of a cobitid fish, Cobitis linea (Heckel, 1849) using different modes of SEM. Curr. Sci. 2006, 91, 1464–1466. [Google Scholar]
- Schamberger, G.P.; Labuza, T.P. Evaluation of Front-face Fluorescence for Assessing Thermal Processing of Milk. J. Food Sci. 2006, 71, 69–74. [Google Scholar] [CrossRef]
- Ikoma, T.; Kobayashi, H.; Tanaka, J.; Walsh, D.; Mann, S. Microstructure, mechanical, and biomimetic properties of fish scales from Pagrus major. J. Struct. Biol. 2003, 142, 327–333. [Google Scholar] [CrossRef]
- Kato, N.; Kunimoto, M.; Koseki, S.; Kitakami, S.; Arai, K. Freshness and Quality of Fish and Shellfish (Supplementary Edition). J. Sch. Mar. Sci. Technol. 2009, 7, 87–99. [Google Scholar]
- Lakowicz, J.R. Principles of Fluorescence Spectroscopy: Protein Fluorescence; Springer Science + Business Media LLC: New York, NY, US, 2006; pp. 63–95. [Google Scholar]
- Masood, Z.; Yasmeen, R.; Haider, M.S.; Tarar, O.M. Evaluations of crude protein and amino acid contents from the scales of four mullet species (Mugilidae) collected from Karachi fish harbor, Pakistan. Indian J. Geo-Mar. Sci. 2015, 44, 724–731. [Google Scholar]
- Tsuchiya, Y. Chemical of Supporting Tissue on Fishery Animals. Nippon Suisan Gakkaishi 1959, 24, 865–874. [Google Scholar] [CrossRef]
- Pati, F.; Adhikari, B.; Dhara, S. Isolation and characterization of fish scale collagen of higher thermal stability. Bioresour. Technol. 2010, 101, 3737–3742. [Google Scholar] [CrossRef] [PubMed]
- Wainwright, D.K.; Lauder, G.V. Three-dimensional analysis of scale morphology in bluegill sunfish, Lepomis macrochirus. Zoology 2016, 119, 182–195. [Google Scholar] [CrossRef] [PubMed]
- Ángeles, E.M. An Overview of the Immunological Defenses in Fish Skin. ISRN Immunol. 2012, 2012, 1–29. [Google Scholar] [CrossRef]
- Park, S.B.; Jang, H.B.; Fagutao, F.F.; Kim, Y.K.; Nho, S.W.; Cha, I.S.; Jung, T.S. Combination treatment against scuticociliatosis by reducing the inhibitor effect of mucus in olive flounder, Paralichthys olivaceus. Fish Shellfish Immunol. 2014, 38, 282–286. [Google Scholar] [CrossRef] [PubMed]
- Oda, Y.; Ichida, S.; Mimura, T.; Maeda, K.; Tsujikawa, K.; Aonuma, S. Purification and characterization of a fish lectin from the external mucous of Ophidiidae, Genypterus blacodes. J. Pharmacobio-Dyn. 1984, 7, 614–623. [Google Scholar] [CrossRef] [PubMed]
- Chong, K.; Tham, S.Y.; Foo, J.; Lam, T.J.; Chong, A. Characterisation of proteins in epidermal mucus of discus fish (Symphysodon spp.) during parental phase. Aqua 2005, 249, 469–476. [Google Scholar] [CrossRef]
- Stella, L.; Capodilupo, A.L.; Bietti, M. A reassessment of the association between azulene and fullerene. Possible pitfalls in the determination of binding constants through fluorescence spectroscopy. Chem. Commun. 2008, 39, 4744–4746. [Google Scholar] [CrossRef] [PubMed]
- Rouissi, H.; Dridi, S.; Kammoun, M.; De, B.J.; Karoui, R. Front face fluorescence spectroscopy: A rapid tool for determining the effect of replacing soybean meal with scotch bean in the ration on the quality of Sicilo-Sarde ewe’s milk during lactation period. Eur. Food Res. Technol. 2008, 226, 1021–1030. [Google Scholar] [CrossRef]
- Georgakoudi, I.; Jacobson, B.C.; Müller, M.G.; Sheets, E.E.; Badizadegan, K.; Carr-Locke, D.L.; Feld, M.S. NADPH and collagen as in vivo quantitative fluorescent biomarkers of epithelial precancerous changes. Cancer Res. 2002, 62, 682–687. [Google Scholar] [PubMed]
- Liu, W.T.; Zhang, Y.; Li, G.Y.; Miao, Y.Q.; Wu, X.H. Structure and composition of teleost scales from snakehead Channa argus (Cantor) (Perciformes: Channidae). J. Fish Biol. 2008, 72, 1055–1067. [Google Scholar] [CrossRef]
- Heu, M.S.; Lee, J.H.; Kim, H.J.; Jee, S.J.; Lee, J.S.; Jeon, Y.; Kim, J.S. Characterization of acid- and pepsin-soluble collagens from flatfish skin. Food Sci. Biotechnol. 2010, 19, 27–33. [Google Scholar] [CrossRef]
- Muyonga, J.H.; Cole, C.G.B.; Duodu, K.G. Fourier transform infrared (FTIR) spectroscopic study of acid soluble collagen and gelatin from skins and bones of young and adult Nile perch (Lates niloticus). Food Chem. 2004, 86, 325–332. [Google Scholar] [CrossRef]
- Mori, H.; Tone, Y.; Shimizu, K.; Zikihara, K.; Tokutomi, S.; Ida, T.; Hara, M. Studies on fish scale collagen of Pacific saury (Cololabis saira). Mater. Sci. Eng. C 2013, 33, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Santana, C.Á.; Lima, D.M.V.; da Cunha Andrade, L.H.; Súarez, Y.R.; Lima, S.M. Laser-induced fluorescence in fish scales to evaluate the environmental integrity of ecosystems. J. Photochem. Photobiol. B Biol. 2016, 165, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, H.H.; Fidel, G.; López, G. Protein-Based Nanoparticles. In Food Nanoscience and Nanotechnology; Springer: New York, NY, USA, 2015; pp. 69–79. [Google Scholar]
- Guardiola, F.A.; Cuesta, A.; Arizcun, M.; Meseguer, J.; Esteban, M.A. Comparative skin mucus and serum humoral defense mechanisms in the teleost gilthead sea bream (Sparus aurata). Fish Shellfish Immunol. 2014, 36, 545–551. [Google Scholar] [CrossRef] [PubMed]
- Lakshmanan, P.T. Fish spoilage and quality assessment. In Quality assurance in seafood processing; Lyre, T.S.G., Kandoran, M.K., Thomas, M., Mathew, P.T., Eds.; Society of Fisheries Technologists: Cochin, India, 2000; pp. 26–40. [Google Scholar]
- Rørå, A.M.B.; Regost, C.; Lampe, J. Liquid holding capacity, texture and fatty acid profile of smoked fillets of Atlantic salmon fed diets containing fish oil or soybean oil. Food Res. Int. 2003, 36, 231–239. [Google Scholar] [CrossRef]
- Yi, E.; Sato, S.; Takahashi, A.; Parameswaran, H.; Blute, T.A.; Bartolák-Suki, E.; Suki, B. Mechanical forces accelerate collagen digestion by bacterial collagenase in lung tissue strips. Front. Physiol. 2016, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]


© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liao, Q.; Suzuki, T.; Yasushi, K.; Al Riza, D.F.; Kuramoto, M.; Kondo, N. Monitoring Red Sea Bream Scale Fluorescence as a Freshness Indicator. Fishes 2017, 2, 10. https://doi.org/10.3390/fishes2030010
Liao Q, Suzuki T, Yasushi K, Al Riza DF, Kuramoto M, Kondo N. Monitoring Red Sea Bream Scale Fluorescence as a Freshness Indicator. Fishes. 2017; 2(3):10. https://doi.org/10.3390/fishes2030010
Chicago/Turabian StyleLiao, Qiuhong, Tetsuhito Suzuki, Kohno Yasushi, Dimas Firmanda Al Riza, Makoto Kuramoto, and Naoshi Kondo. 2017. "Monitoring Red Sea Bream Scale Fluorescence as a Freshness Indicator" Fishes 2, no. 3: 10. https://doi.org/10.3390/fishes2030010
APA StyleLiao, Q., Suzuki, T., Yasushi, K., Al Riza, D. F., Kuramoto, M., & Kondo, N. (2017). Monitoring Red Sea Bream Scale Fluorescence as a Freshness Indicator. Fishes, 2(3), 10. https://doi.org/10.3390/fishes2030010