
Chlorella vulgaris as Protein Source in the Diets of African Catfish Clarias gariepinus

Department of Fisheries and Aquatic Resources Management, Michael Okpara University of Agriculture, Umudike, Umuahia, Abia State, Nigeria
Fishes 2017, 2(4), 17; https://doi.org/10.3390/fishes2040017
Submission received: 12 September 2017
/
Revised: 24 September 2017
/
Accepted: 3 October 2017
/
Published: 16 October 2017
Abstract
:Plant proteins substitutes of fishmeal in aquafeed are usually lacking in some essential amino acids and fatty acids. The microalgae Chlorella vulgaris has good-quality protein with amino acids rich in methionine, lysine and alanine. Four novel diets having C. vulgaris as the main source of protein were produced for African catfish Clarias gariepinus with an initial average weight of 1.09 ± 0.05 g. The diets were labeled Feed 1 (F1) to feed 4 (F4). The treatment diets were included 25% (F1), 15% (F2), 5% (F3) and 0% (F4) green algae meal. The basal ingredients of the feed were corn (maize) included as F1, 40%, F2, 43%, F3, 53% and F4, 43%; and millet meal, which varied in F1 as 23%, F2, 30%, F3, 30% and F4, 30%. The ingredients were preconditioned at 110 °C and pelleted. Post-fingerling African catfish were stocked at 10 fish per aquarium. There were three replicate aquariums for each feed type and the fish were fed for 60 d. The specific growth rate was best for the catfish fed with 25% C. vulgaris diet 7.86 ± 0% day−1, and worst at 6.77 ± 0.07% day−1 for the control group F4, 0% algal meal. The food conversion ratio (FCR) was lowest (1.88 ± 0.02) for 25% algal meal diet (F1) and highest (2.98 ± 0.01) for the 0% algal meal diet F4. Similarly, catfish had average weight gain of 121.02 ± 0.04 g for those fed with F1 compared to 62.50 ± 0.0 g for those fed with 0% algae F4. Protein efficiency ratio was highest for the F1-fed fish (2.46 ± 0.22) and lowest for those fed with F4 (2.02 ± 0.09). The hepatosomatic index was lowest for F1-fed fish (1.48 ± 0.01) and highest for catfish fed with F4 (2.50 ± 0.59). Based on the results, C. vulgaris is a good protein source for African catfish and can also substitute fishmeal in the catfish diets.
1. Introduction
Plant proteins are currently the major substitute of fishmeal in aquafeed production. The substitution of fishmeal with plant protein meals is hampered by the presence of anti-nutritional factors [1,2,3]. Use of plant proteins is also affected by deficiencies in some essential amino acids such as lysine, methionine, threonine and tryptophan [4,5] and poor digestibility [4]. However, the use of protein concentrate meals has solved the problem of anti-nutritional factors (ANF) and low protein content, but ended up presenting problem of high cost [6]. Algae are simple aquatic plants, ranging from the microscopic (microalgae) Chlorella, Spirulina and Spirogyra to macroscopic algae like giant kelp. Algae are at the base of the aquatic food chain and food resources that fish eat are produced there [7]. Microalgae are source of protein and lipids depending on the species [8,9]. It may be possible to substitute fishmeal with algal protein meal in production of aquafeed. For microalgae to be used as a fish feed ingredient, it must not be toxic to the fish or humans, it should be easy to culture, and have high nutritional value with a digestible cell wall [10].
Chlorella vulgaris is a unicellular algae that grows fast. It does not compete as human food, and needs CO2 and nutrients to constantly produce biomass. Algae are normally found in the ponds, where it grows freely and can easily be harvested and re-grown. Algae can be grown in closed systems (vertical bubble column) or horizontal tubular photobioreactors, flat panels, biocoils, and bags. Closed systems favor efficient control of the growth of algae and, moreover, closed systems favor an efficient control of the algae growth by improving the monitoring of the growth parameters. Algae can also be grown in open ponds, and in other open systems (turf scrubber system, raceways, and tanks). Open system can give more yield than closed system. Chlorella has been incorporated into fish diets, and was fed to Korean rockfish [11]. It has been noted in previous studies that inclusion of (2.5–10% of the diet) of algae into fish diets enhanced growth performance, feed utilization efficiency, intestinal microbiota, carcass quality and physiological activity, [12,13]. C. vulgaris is unicellular green algae and has been widely used as human food supplements [14]. Analysis of amino acid content of numerous algae have found that, although there is significant variation in total protein content (8–50% dry weight) [8], they generally contain all the essential amino acids [15,16]; and studied algae have also been found to be rich in n-3 long-chain polyunsaturated fatty acids (LC-PUFA).
Essentially, algae derived products seem to be a very suitable raw material for use in aquafeed. A probable advantage of feeding algae to fish in aquaculture is that fish flesh will have what could be termed as natural river taste (for fresh water fish/shrimp). Wild fishes actually acquire this taste by consuming algae, from which they derive the nutrients. The ratio of omega-3:omega-6 in fish oil will be better when fish consumes algae compared to when plant proteins are used. Ordinarily, fish fed with plant proteins like soybean meal or corn meal would present low levels of omega-3 (ω3) with eicosapenatoic fatty acids and relatively high levels of omega-6 arachidonic fatty acids. Microalgae have been incorporated in aquafeed because of their high content of nutritionally essential polyunsaturated fatty acids [17]. Algae has more of ω3 LC-PUFA than plant, which has more of (short-chain fatty acids) SFA, monounsaturated fatty acids (MUFA) and ω6 PUFA [18]. Microalgae have been reported to improve growth when incorporated into diets of shrimp [19] and tilapia [20]. However, there was no significant improvement of growth in juvenile Atlantic salmon fed 2.5 to 10% commercial algae meal [21]. Spirogyra africana (Fritsch) Czurda is a filamentous alga found in freshwater habitats [22,23]. Spirogyra is among the commonest green algae abundant in freshwater, sewers and gutters. It comes as bright green free-floating masses on top of water bodies [24,25]. Spirogyra has arachidic acid, erucic acid and docosadienoic. The quantities of arachidic acid (C20:4), erucic (C22:1), and docosadienoic (C:22:2) to docosadienoic acid were 89.97 µg/mL for arachidic acid, 190.86 µg/mL for erucic acid, and 101.28 µg/mL for docosadienoic acid [26]. C. hlorella vulgaris is a microalgae and green algae that can thrive in both fresh and marine water and has wide application as a food supplement [27]. Chlorella is one of the most utilized microalgae in fish diets [28,29]. In previous research, Bakhtiyar et al. [30] noted that zooplanktons bioenriched with Chlorella and fed to Labeo rohita affected higher survival and body carcass composition of L. rohita. C. vulgaris have also been included in diets of shrimp and, after 90 d of a feeding experiment, the specific growth rate, feed conversion efficiency and weight gain and were significantly (p < 0.05) better for those fed 50% C. vulgaris inclusion diet than the rest [31].
The algae used in this experiment were identified in a private fish farm at Enugu, Nigeria. The identification and isolation of the algae followed a chance observation that the African catfish Clarias gariepinus stocked in a pond were constantly grazing on the algae formation at the perimeters of the concrete pond. The grazing of the algae created a line of ring round the pond, which attracted the researcher’s attention. The algae were identified as C. vulgaris and isolated and grown in a separate open pond all alone. The algae were fed with filtered effluent water from a nearby fish farm. The water was collected from the farms with a 20 L plastic can. The water was filtered with 0.001 mm sieve and fed into the pond where the isolated C. vulgaris were planted. The algae were allowed to grow for a week, after which it was constantly harvested twice daily until whole biomass used in this experiment was achieved.
This research seeks to examine the growth and feed utilization of African catfish C. gariepinus fed with diets incorporating freshwater microalgae Chlorella vulgaris for 60 d. African catfish C. gariepinus is the most popular aquaculture fish species in Africa. It grows very fast and tolerates harsh rearing conditions. C. gariepinus grows very fast and is easy to breed. The egg hatches within 24–42 h depending on water temperature.
2. Results
The catfish easily accepted feed made from C. vulgaris and grew with optimal specific growth rate (SGR) of 7.86 ± 0.45% day−1 achieved by those catfish fed with 25% algae diet, feed 1, (F1). There was no significant difference (p > 0.05) between the SGR of catfish fed with F1, F2 and F3 (Table 1). The SGR of the catfish fed with 5% chlorella diet (F3) was 7.36 ± 0.44% day−1, but was significantly different from those fed with (F4) 0% chlorella (p < 0.05). The lowest SGR was from the fish fed with F4 with 0% C. vulgaris 6.77 ± 0.07% day−1. However, the catfish SGR was higher for those fed with F2 compared to those fed with F3 (p < 0.05).
The food conversion ratio (FCR) was lowest for those fed with diet F1, (1.92 ± 0.05) and F2 (1.88 ± 0.02). There was no significant difference between the FCR of catfish fed with (F2) and F1 (p > 0.05). There was significant difference (p < 0.05) between the FCR of catfish fed with F2 and F3 (2.22 ± 0.11). Similarly, the catfish fed with F3 had better FCR than those fed with F4 (2.98 ± 0.01). FCR seems to be increasing with decreasing inclusion of Chlorella meal in the diets.
The average weight of the catfish was positively affected by the amount of C. vulgaris in the feed. The catfish fed with 25% (F1) Chlorella diets achieved and average weight gain of 121.02 ± 0.04 g. The average weight gains of the catfish fed with F2, was 108.23 g and this was much lower than the average weight gain (AWG) of those fed with F1 (p < 0.05). Feeding the catfish with F3 reduced the average weight gain of the catfish to 89.25 ± 0.02 g. However, the catfish feed with F4 had the lowest AWG of 62.50 ± 0.01 g. The AWG seems to be increasing with the increasing inclusion of C. vulgaris meal in the catfish diets.
The protein efficiency ratio (PER) was highest for the fish fed with F1 (2.46 ± 0.22) followed by those fed with F2, (2.30 ± 0.13). There was no significant difference between the PER of F1 and F2 (p > 0.05). The catfish PER was 2.07 ± 0.03 for those that received F3 while it was 2.02 ± 0.09 for those on F4 (0% C. vulgaris). The catfish PER seems to be decreasing along with a reduction of algae in the diet (Table 1).
The protein conversion ratio (PCR) was lowest for catfish fed with F1 (0.61 ± 0.02) and F2 (0.60 ± 0.03). There was no statistical difference between the PCR of catfish fed with F1 and F2 (p > 0.05). However, the PCR was higher for catfish fed with F4 (0.95 ± 0.23) than those fed with F3 (Table 1).
The hepatosomatic index (HSI) of the fish was lowest for catfish fed with F1, 1.48 ± 0.01 followed by those fed with F2, HSI, 1.53 ± 0.1. There is no statistically significant difference (p > 0.05). The highest HSI of the catfish were obtained from those fish fed with 0% fishmeal (FM) diet.
The waste production ratio (WPR) of catfish was highest for those fed with F4 (2.50 ± 0.21).
That means that for each kg of fish flesh produced 1.91 kg of waste is produced. The WPR of catfish fed with F4 (2.50 ± 0.21) was the highest and significantly different from all other treatments (p < 0.05). The WPR of catfish fed with F1 (1.54 ± 0.10) was not significantly different (p > 0.05). The WPR of catfish fed with F2 was significantly different (p < 0.05) from those fed with F3 compared to the other groups. The catfish WPR does not seem to be following any pattern Table 2.
The dry matter ratio (DMR) measures the quantity of dry matter in feed needed to produce 1 kg of fish biomass on dry weight bases. The catfish fed with F1 had the least DMR of 7.95 ± 0.1. This means that 7.95 ± 0.1 of dry matter in the feed is needed to produce 1 kg of the fish. The DMR of the catfish fed with F2 was significantly was significantly different (p < 0.05) from the DMR of those fed with F3. The highest DMR was 12.34 ± 0.22 which was from the catfish fed with F4. The DMR of the catfish seems to be increasing with reducing inclusion of C. vulgaris meal in the catfish diets (Table 1).
The daily feed intake (DFI) of the catfish was similar for all treatment diets except F1. The catfish DFI of F1 was highest 3.88 ± 0.15 g day−1 and this was significantly higher (p < 0.05) than that of F2, which was nearest value (DFI, 3.39 ± 0.04 g day−1). The DFI of the catfish fed with F4 was lower in number (3.11 ± 0.08 g day−1) than those fed with F2, 3.39 ± 0.04 g day−1, but they were not significantly different (p > 0.05). Daily feed intake of the fish seems to be decreasing with decreasing inclusion of C. vulgaris in the diets.
3. Discussion
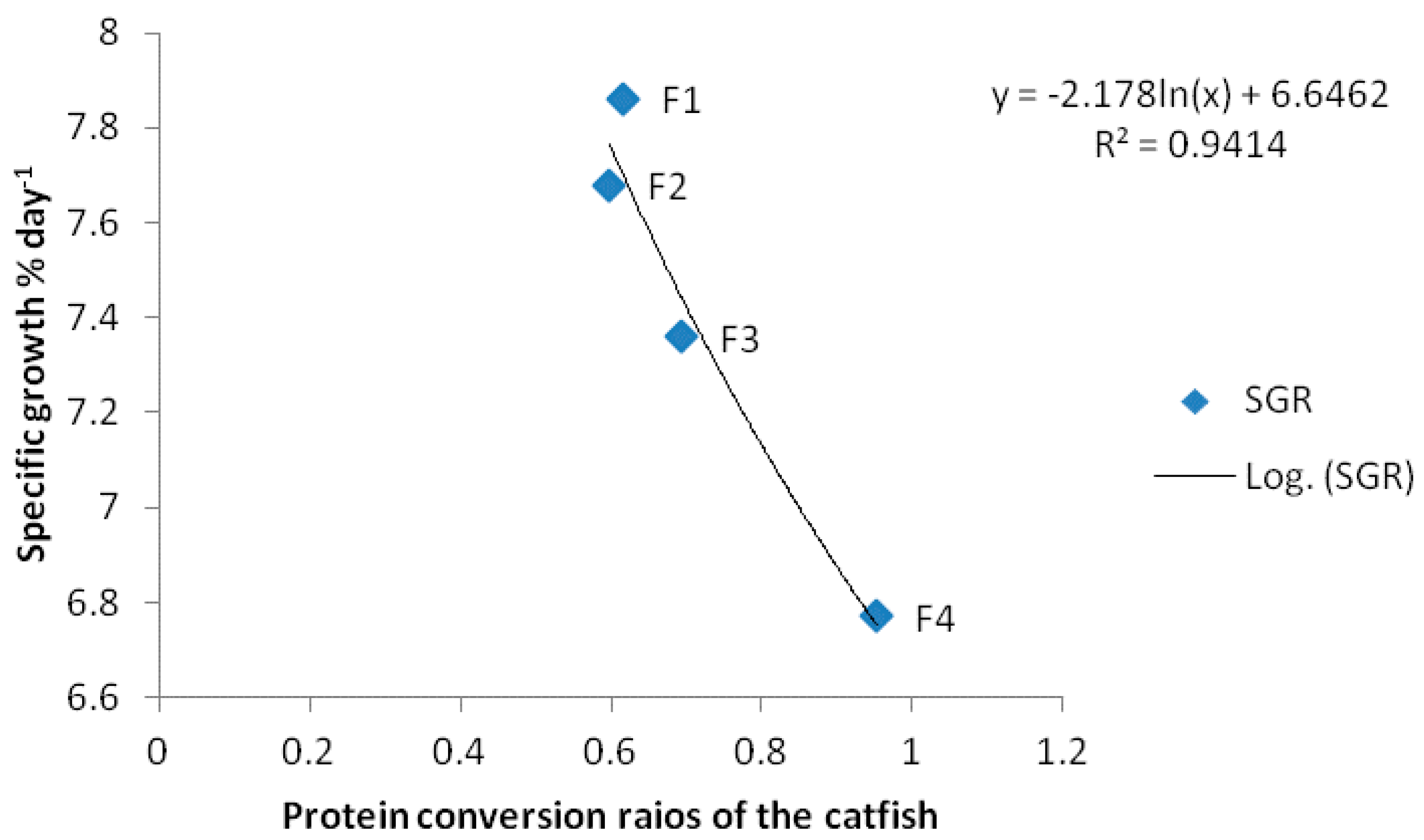
The acceptance did not come as a surprise because the catfish were accidentally observed to be grazing on the algae in a concrete pond. The specific growth rate of the fish fed with the experimental diets is indicative of the nutritional quality of the diets. The treatment diets were designed so that C. vulgaris would be the principal source of protein. Chlorella contains about 42–58% protein dry weight [32,33,34]. Similarly, Zeinhom [35] noted that inclusion of algae in fish diets improved growth. The high inclusion level of C. vulgaris (25%) could have contributed in the better growth rate of C. gariepinus more than the fishmeal diet. It is important to note that the C. gariepinus fed with F4 grew well at a SGR of 6.77 ± 0.07% day−1, but the algal diets even achieved a better growth performance. The growth rate of the C. gariepinus fed with the Chlorella based feeds could have also been because of the ease in digesting the ingredient. Although there was no digestibility test in the work, it seems the Chlorella being freshly harvested and processed into the feed could have been better utilized. It is also possible that fishmeal used in this work could have lost some amino acid due to long storage in the dealer’s shop before sales. More over the higher plant ingredients in F4 could have affected nutrient utilization at least more than the Chlorella meal. Nevertheless, the reason for the better performances of the Chlorella diets is not very clear. However the protein conversion ratio (PCR) of the catfish were lower for the Chlorella fed fish, suggesting that smaller quantity of the chlorella was needed to produce 1 g of the C. gariepinus flesh. There was negative correlation between the protein conversion ratio and specific growth rate of the catfish R2 = 0.9414 (Figure 1). Similarly, there was positive correlation between the protein conversion ratio and the food conversion ratio of the catfish (Figure 2). In a previous research, Radhakrishnan et al. [31] noted that postlarvae Macrobrachium rosenbergii fed with diets having a 50% inclusion of C. vulgaris did better in terms of SGR, weight gain, feed efficiency and survival compared to other inclusion levels below and above. The performances of fish fed with C. vulgaris could have been related with the amino acid balance. C. vulgaris has array of essential amino acids better than soybean and even egg [36]. In previous work, Shi et al. [37] noted that Chlorella mixed with rapeseed meal can completely replace fishmeal in diets of crussian carp. Similarly, Pakravan et al. [38] noted that Chlorella can completely replace fishmeal in the diets of Litopenaeus vannamei. Chlorella is rich in amino acids like, aspartic acid, serine, alanine, leucine and glycine, and these have been reported to be collectively responsible for 50% or more of the total dry matter content of Chlorella spp. [39]. This research is also in line with Dawah et al. [40], who noted that inclusion of Scenedesmus Bijuga (Chlorophyceae) in the diets of monosex O. niloticus improved growth performances.
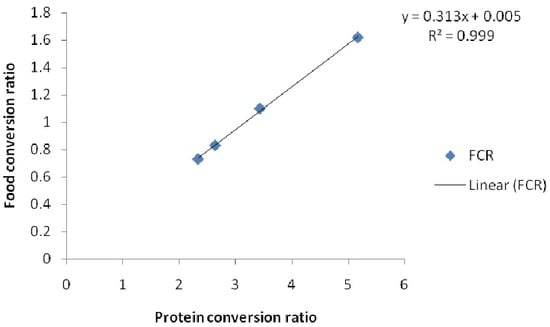
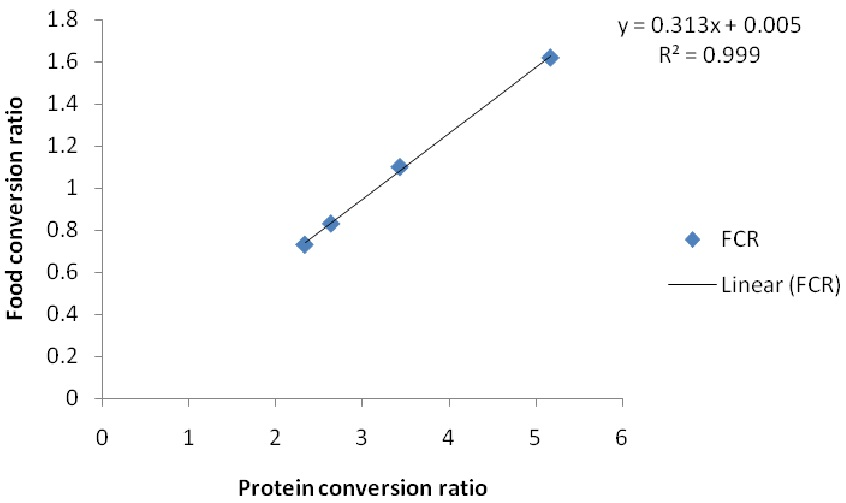
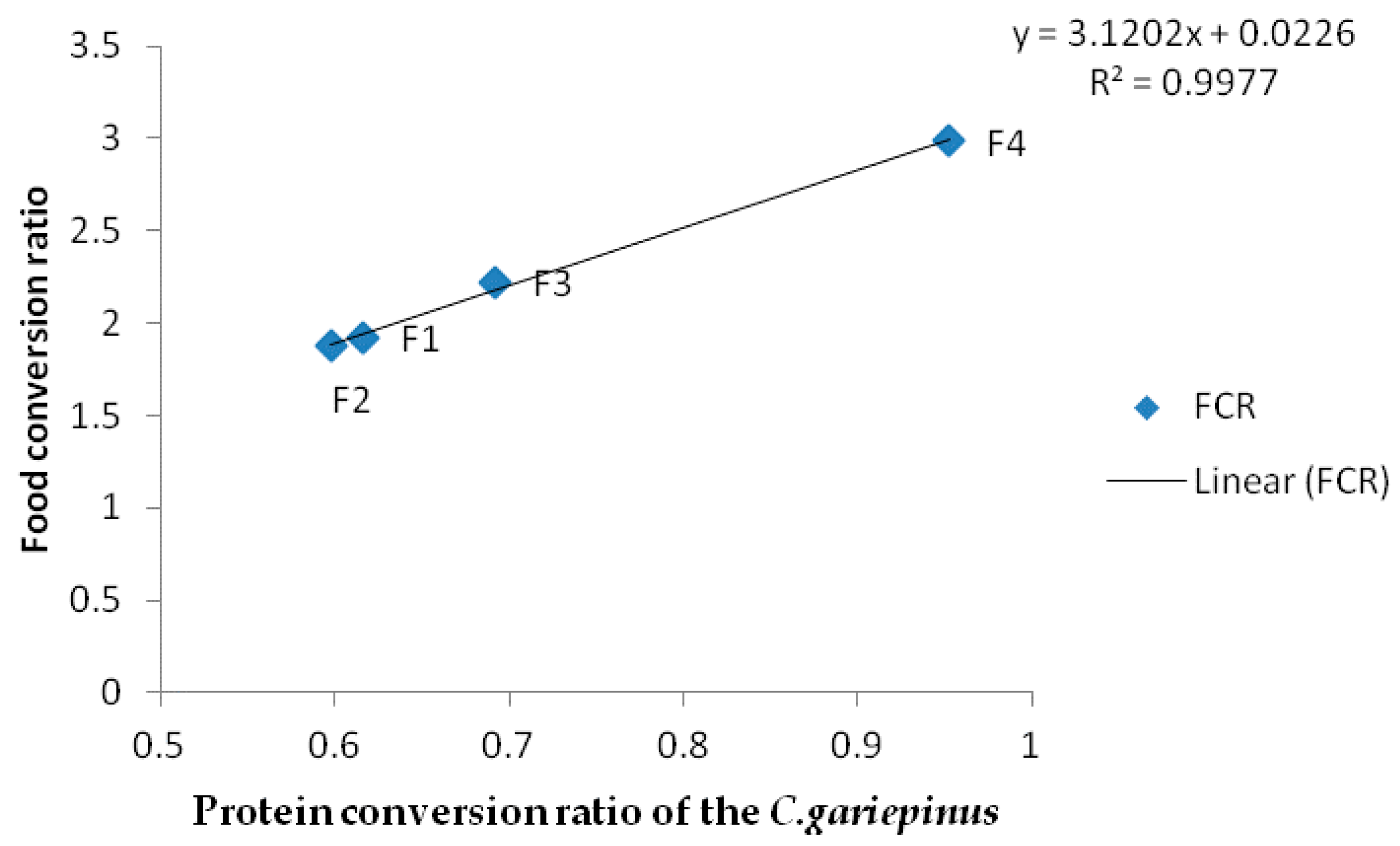
The FCR of the fish reduced with increasing PCR and the inclusion of the algal meal R2 0.9973 (Figure 2). This could have been due to combined effects of the fatty acid and amino acids on the nutrient digestibility. Zeinhom [35] observed that inclusion of algae improved FCR of fish. Also, Dawah et al. [40] noted that food conversion ratio was lower when the fish were fed on artificial diets having 10% and 20% algal meal. The effects of algae on feed utilization of fish had been noted by previous workers. Green algae enhanced the nutritional value of zooplanktons in rearing of Atlantic cod larvae [41]. Similarly, Wang et al. [42] noted that inclusion of daphnia and Chlorella improved the growth of larval loach. Cahu et al. [43] had earlier noted that microalgae inclusion in the diets of sea bass (Dicentrarchus labrax) improved digestive functions. These reports point to the fact that inclusion of algae-enhanced nutrient utilization and the digestibility of larval forms. These reports seem to suggest that diets made with algae are easily digestible even in the larval gut. The high content of essential amino acids like lysine and methionine in C. vulgaris may have also contributed to enhance feed utilization. Microalgae have been reported to enhance larval growth and survival [41,42]. This could be due to the high and amino acid content which are reported well for lowering FCR in bigger fish. Nevertheless, some previous researchers noted that inclusion of algae in diets of carnivorous fish like salmon did not have any effect except on FCR [21]. Others, like Güroy et al. [44], noted no visible growth effect in rainbow trout, and in European sea bass (Dicentrarchus labrax), as did Valente et al. [13]. The acceptance and performance of fish fed with Chlorella species meal could depend on the food preference, feeding habits and distribution of the fish species.
The increase in catfish body weight as C. vulgaris inclusion increased is in line with previous reports. Nandeesha et al. [45] noted that feeding of Spirulina platensis below 20% improved the growth, proximate composition and that body weight gain of Nile tilapia (O. niloticus) had linear increased. Similarly, Tartiel et al. [46] reported increase in body weight of O. niloticus fed with dried meals of Chlorella spp. and Scenedesmus spp. The effects of algae on weight increase can be linked to the increase in fatty acids contents of the fish. The fatty acids in fish, especially eicosapentaenoic acid (EPA), docosahexaenoic acid (DHA), and arachidonic acid (ARA), are essentially passed from the food chain to the fish and algae seem to be one of the major sources [47]. In this present research we noted linear weight increase as inclusion level of C. vulgaris increases. This may have been due to increase in lipids of the fish. Moreover, our diets had high inclusion levels of carbohydrate like millet and corn meal. Carbohydrates are well utilized by African catfish [48], but are converted to fat and stored in the fish body.
The hepatosomatic index of the fish reduced with increasing inclusion of C. vulgaris meal in the catfish diet. It seems that inclusion of microalgae in the C. gariepinus diet enhanced the utilization of the carbohydrate. High carbohydrate inclusion in the diets of catfish usually reflects an increased HSI of the fish. Elevated HSI had been associated with high carbohydrate diets in hybrid striped bass [49], African catfish [48,50], Chinese longsnout catfish [51], southern catfish [52]. The utilization of carbohydrates by the catfish fed with algae suggests enhancement of the carbohydrate digestion by nutrients from the algae. It seems that algae inclusion increased the natural enzymatic digestion in the small intestine, which is optimal for nutrient utilization. This is because virtually the same quantity of carbohydrate ingredients was included in the diets.
The inclusion of algae in the catfish diet did not have any definite effect on the waste production ratio. This could be because algal inclusion may have some other physiological functions that may not be quantified by growth. This was noted by Sato et al. [53] for red sea bream, enhancing predation on zooplankton [54] and for rainbow trout [44]. The dry matter ratio of the catfish was reduced with increasing inclusion of C. vulgaris. This suggests that lesser dry matter in the feed is needed to produce 1 kg of dry matter in the fish. Consequently, the diets made from algal meal are more efficient. This result is in line with findings of Shi et al. [37], who noted that FM can be completely replaced by mixture of Chlorella and rapeseed meal. The daily feed intake did not show much difference except for the 25% C. vulgaris. The reason could be organoleptic, since fish were observed prior to the experiment to be grazing on the algae on their own. There could have been some organoleptic attractant that made the fish attracted to the algae. This goes to suggests that some fish that may be obligate carnivores may not accept the algae meal inclusion. However, this is subject to further research as our findings here cannot support such a claim.
4. Materials and Methods
4.1. Isolation and Cultivation of Algae
The algae used in this experiment were discovered and identified in a fish pond in Enugu, Nigeria (see introduction section). The algae were grown in open ponds and were harvested with a scoop net and transferred into a white base plastic bath of 20 L capacity. The algae water complex was allowed to settle for three hours. After 3 h the water was decanted and the resultant algae were dried at regular temperature of 40 °C using an electric oven. The dried meal was ground to dust using an electric grinder and stored at −20 °C till used. Microalga C. vulgaris grows very fast because it has mixotrophic growth, the capability of combining autotrophic and heterotrophic nutrition techniques by performing photosynthesis, as well as ingesting organic matter, even in a dark environment [55,56].
4.2. Feed Formulation
Four types of feed (Feed 1, F1 to feed 4 F4, were designed to vary in algal meal inclusion as 25%, 15%, 5% and 0%. The feed varied in composition of corn meal as follows F1, 40%, F2, 43% and F3, 53% and F4, 43. Millet meal, (purchased from Scionure feed ingredients market Enugu Nigeria) was also added in F1 as 23%, F2, 30%, F3, 30% and F4 30%. The proximate composition of chlorella meal and millet are tabulated in Table 3. There was no fortification of the diets with amino acids and algal meal was the main source of proteins in the feed. The rest of the basal ingredients were same for all diets. The ingredients were measured and mixed with an electric mixer and 150 mL of water was added to the feed. The dough was preconditioned at 120 °C and then pelleted. The pellets were dried at 40 °C to constant water activity of 8%. The feeds were stored at −20 °C until used.
4.3. Experimental Set Up
The research was set up at the fish research wet lab of Godfrey Okoye University Emene Enugu, Enugu state, Nigeria. Three plastic aquariums per treatment were used for this experiment. The aquariums had a dimensioned width of 50 cm, a length of 80 cm and a height 30 cm. A total of 12 aquariums were used for the experiment. The tanks were randomly distributed to ensure uniformity and subjected to 12:12 day: night light regime. The wet lab is roofed and offers shade, although the four sides are all exposed to daylight. The light intensity was 30 lux measured with a lux meter. The experiment was carried out in stagnant water, but this was completely changed daily. Water change was with rubber siphon, which gently removes the waste and water without disturbing the fish. Water change was done by 8–9 a.m. daily. Before actual feeding experiment started the fish were acclimated for 9 d, during which period they were fed with 35% protein commercial diet.
4.4. Feeding of Fish and Hygiene
The fish were fed to satiation three times daily. Feeding was done by hand and care was taken to avoid overfeeding of the fish. Feeding was done in the morning hours between 8–9 a.m. and in the afternoon between 1–2 p.m. and in the evening 6–7 p.m. The aquariums were cleaned every morning and fecal matter was removed from the tanks with any uneaten food. The water in the aquariums was changed daily to maintain good culture condition. The water used in the experiments was from university water reservoir but was filtered before use. Physico-chemical parameters of the water culture system were measured every 5 d throughout the period of the experiment. Water was aerated during the experiment and the average temperature (mean + SD, n = 22) 30.0 ± 1.5 °C. The water total gas pressure was 101 ± 2.07 (P4 Tracker Total Gas Pressure saturometer; Point Four Systems Inc., Richmond, VA, USA). Dissolved oxygen content was 4.05 ± 1.09 mg L−1 (YSI oxygen meter, Yellow Springs, OH, USA) and pH was 7.05 ± 0.77. Ammonia concentration was 0.6 ± 0.6 mg/L (Tetra ammonia kit, Malvern, PA, USA).
4.4.1. Weight Measurement of Fish
Fish were weighed per replicates of the treatment diets. The fish weighed together and the average of the treatment feed was used. Weighing was done with electronic balance sensitive to 0.0001 g. The catfish were not fed for about 12 h before weighing.
4.4.2. Growth and Nutritional Parameters Analyzed
At the end of the experiment five fish were taken from each aquarium for analyses of proximate composition, hepatosomatic index (HSI) and peritoneal fat somatic index (FSI). The fish were dissected and liver and visceral fat of the fish were removed and weighed (to 0.001 g). These five fish per aquarium were then pooled, ground together and freeze dried for calculation of water content, and stored at −80 °C to be used in analyzing proximate composition. Moisture content of the feeds was determined by oven drying feed samples at 70 °C for two days while freeze dried feed samples were used for lipid and protein analyses.
Ash content was determined by incineration samples in a muffle furnace at 550 °C for 24 h. The ash percentage was calculated as weight of ash/weight of sample ×100. The dry matter ratio (DMR) measures the quantity of dry matter in feed needed to produce 1 kg of dry matter of fish.
Total lipids were analyzed using a modified chloroform methanol method. Lipids in the samples were extracted twice with 2:1 chloroform:methanol mixture. Lipid extraction was after modified methods of Parrish [57], and Kainz et al. [58]. Whole body protein was analyzed by Kjeldahl method, using Tecator Kjeltech model 1002 (Tecator, Kjeltect, Höganäs, Sweden). Protein percentage was expressed as %N × 6.25.
4.4.3. Calculations and Statistical Analyses
Specific growth rate (SGR, % day−1) was determined as:
SGR = 100 × (Ln final weight (g) − Ln initial weight (g)) (period in days)−1, where Ln is the natural logarithm of the final and initial weights.
Food conversion ratio (FCR) = feed fed (g) × weight gain−1 (g)
Protein efficiency ratio (PER) = weight gain (g) × protein intake−1 (g)
Hepatosomatic index (HSI) = 100 × liver weight (g)/fish weight (g)
Daily feed intake (daily satiation feeding) = Weight of feed given in (g) − uneaten feed (rejects)
DMR = FCR × (%DM in feed) × (%DM in culture species)−1
(Dry matter ratio = FCR (% dry matter in feed × (% dry matter in culture species)−1
Waster production ratio (WPR) = (DMR − 1) × (%DM in culture species) × 100−1
Protein conversion ratio (PCR) = FCR × (crude protein of feed/100)
The data were analyzed using analysis of variance and Fisher’s least significant difference (FLSD0.05) was used to separate treatment means. PASW version 18 statistical software SPSS (Hong Kong, HK) was used in the data analysis.
5. Conclusions
Algal meal of C. vulgaris was well accepted by African catfish as a fish feed ingredient. Inclusion should be done with care to ensure that fish can utilize the diet. However, our results show that C. vulgaris can substitute fishmeal in C. gariepinus diets. The acceptance of the diet and the high SGR suggests it can be a cheap way of making feed since it is inexpensive to grow the algae and it can be cost saving compared to fishmeal. In contrast to fish meal, algae feeds could be a sustainable alternative source for proteins for a number of fish species. The use of the algae could lead to early harvesting, which is cost saving and beneficial to the farmer.
Conflicts of Interest
The author declares no conflict of interest.
References
- Francis, G.; Makkar, H.P.S.; Becker, K. Anti-nutritional factors present in plant-derived alternate fish feed ingredients and their effects in fish. Aquaculture 2001, 199, 197–227. [Google Scholar] [CrossRef]
- Gatlin, D.M., III; Barrows, F.T.; Brown, P.; Dabrowski, K.; Gaylord, G.T.; Hardy, R.W.; Herman, E.; Hu, G.; Krogdahl, Å.; Nelson, R.; et al. Expanding the utilization of sustainable plant products in aquafeeds: A review. Aquac. Res. 2007, 38, 551–579. [Google Scholar] [CrossRef]
- Vielma, J.; Ruohonen, K.; Gabaudan, J.; Vogel, K. Top-spraying soybean meal-based diets with phytase improves protein and mineral digestibilities but not lysine utilization in rainbow trout Oncorhynchus mykiss (Walbum). Aquac. Res. 2004, 31, 955–964. [Google Scholar] [CrossRef]
- Li, P.; Mai, K.; Trushenski, J.; Wu, G. New developments in fish amino acid nutrition: Towards functional and environmentally oriented aquafeeds. Amino Acids 2009, 37, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Uchechukwu, E.; Pirhonen, J.; Vielma, J. Effects of sesame seed meal and bambaranut meal on growth and feed utilization and body composition of Juvenile African catfish Clarias gariepinus. Iran. J. Fish. Sci. 2014, 13, 998–1013. [Google Scholar]
- Hardy, R.W. Utilization of plant proteins in fish diets: Effects of global demand and supplies of fishmeal. Aquac. Res. 2010, 41, 770–776. [Google Scholar] [CrossRef]
- Mustafa, M.G.; Nakagawa, H. A review: Dietary benefits of algae as an additive in fish feed. Isr. J. Aquac. Bamid. 1995, 47, 155–162. [Google Scholar]
- Fleurence, J. Seaweed proteins: Biochemical, nutritional aspects and potential uses. Trends Food Sci. Technol. 1999, 10, 25–28. [Google Scholar] [CrossRef]
- Guccione, A.; Biondi, N.; Sampietro, G.; Rodolfi, L.; Bassi, N.; Tredici, M.R. Chlorella for protein and biofuels: from strain selection to outdoor cultivation in a Green Wall Panel photobioreactor. Biotechnol. Biofuels 2014, 7, 84. [Google Scholar] [CrossRef] [PubMed]
- Patil, V.; Kallqvist, T.; Olsen, E.; Vogt, G.; Gislerod, H.R. Fatty acid composition of 12 microalgae for possible use in aquaculture feed. Aquac. Int. 2007, 15, 1–9. [Google Scholar] [CrossRef]
- Bai, S.C.; Koo, J.-W.; Kim, K.W.; Kim, S.K. Effects of Chlorella powder as a feed additive on growth performance in juvenile Korean rockfish, Sebastes schlegeli (Hilgendorf). Aquac. Res. 2001, 32 (Suppl. S1), 92–98. [Google Scholar] [CrossRef]
- Nakagawa, H. Effect of dietary algae on improvement of lipid metabolism in fish. Biomed. Pharmacother. 1997, 51, 345–348. [Google Scholar] [CrossRef]
- Valente, L.M.P.; Gouveia, A.; Rema, P.; Matos, J.; Gomes, E.F.; Pinto, I.S. Evaluation of three seaweeds Graci-laria bursa-pastoris, Ulva rigida and Gracilaria cornea as dietary ingredients in European sea bass (Dicentrarchus labrax) juveniles. Aquaculture 2006, 252, 85–91. [Google Scholar] [CrossRef]
- Görs, M.; Rhena Schumann, R.; Hepperle, D.; Karsten, U. Quality analysis of commercial Chlorella products used as dietary supplement in human nutrition. J. Appl. Phycol. 2010, 22, 265. [Google Scholar] [CrossRef]
- Ortiz, J.; Romero, N.; Robert, P.; Araya, J.; Lopez-Hernández, J.; Bozzo, C.; Navarretea, E.; Osorioa, A.; Riosa, A. Dietary fiber, amino acid, fatty acid and tocopherol contents of the edible seaweeds Ulva lactuca and Durvillaea antarctica. Food Chem. 2006, 99, 98–104. [Google Scholar] [CrossRef]
- Dawczynski, C.; Schubert, R.; Jahreis, G. Amino acids, fatty acids, and dietary fibre in edible seaweed products. Food Chem. 2007, 103, 891–899. [Google Scholar] [CrossRef]
- Sardesa, V.M. Nutritional role of polyunsaturated fatty acid. J. Nutr. Biochem. 1992, 3, 154–166. [Google Scholar] [CrossRef]
- Catarina, G.A.; Meireles, L.A.; Amaro, H.M.; Xavier, M.F. Changes in lipid class and fatty acid composition of cultures of Pavlova lutheri, in response to light intensity. J. Am. Oil Chem. Soc. 2010, 87, 791–801. [Google Scholar]
- Amaya, E.A.; Davis, D.A.; Rouse, D.B. Replacement of fishmeal in practical diets for the Pacific white shrimp (Litopenaeus vannamei) reared under pond conditions. Aquaculture 2007, 262, 393–401. [Google Scholar] [CrossRef]
- Azaza, M.S.; Mensi, F.; Ksouri, J.; Dhraief, M.N.; Brini, B.; Abdelmouleh, A.; Kraiem, M.M. Growth of Nile tilapia (Oreochromis niloticus L.) fed with diets containing graded levels of green algae ulva meal (Ulvarigida) reared in geothermal waters of southern Tunisia. J. Appl. Ichthyol. 2008, 24, 202–207. [Google Scholar] [CrossRef]
- Norambuena, F.; Hermon, K.; Skrzypczyk, V.; Emery, J.A.; Sharon, Y.; Beard, A.; Turchini, G.M. algae in fish feed: Performances and fatty acid metabolism in juvenile atlantic salmon. PLoS ONE 2015, 10, e0124042. [Google Scholar] [CrossRef] [PubMed]
- Gallego, I.; Jesús Casas, J.; Fuentes-Rodríguez, F.; Juan, M.; Sánchez-Castillo, P.; Pérez-Martínez, C. Culture of Spirogyra africana from farm ponds for long-term experiments and stock maintenance. Biotechnol. Agron. Soc. Environ. 2013, 17, 423–430. [Google Scholar]
- Kim, J.-H.; Kim, Y.H.; Lee, I.K. Morphotaxonomy of the genus Spirogyra (Zygnemataceae, Chlorophyta) in Korea. Algae 2004, 19, 91–105. [Google Scholar] [CrossRef]
- Ghazala, B.; Hena, L.; Zarina, A.; Shameel, M. Taxonomic survey of fresh water algae at the campus of BZ University of Multan, Pakistan. Int. J. Phycol. Phycochem. 2009, 5, 77–99. [Google Scholar]
- Masud-ul-Hasan, A.Z.; Shameel, M. Microtaxonomical studies on Chlorophycota and Vaucherophycota from Jauharabad District, Pakistan. Int. J. Phycol. Phycochem. 2010, 6, 141–154. [Google Scholar]
- Trifa, F.K.; Othman, F.A.; Omer, A.T. Oil and fatty acid composition of spirogyra and chara species from Bestan SWR spring water in Sulaimani-Kurdistan Region of Iraq. Egypt. J. Exp. Biol. 2013, 9, 159–162. [Google Scholar]
- Kay, R.A. Microalgae as food and supplement. Critical reviews. Food Sci. Nutr. 1991, 30, 555–573. [Google Scholar]
- Safi, C.; Charton, M.; Pignolet, O.; Silvestre, F.; Vaca-Garcia, C.; Pontalier, P.-Y. Influence of microalgae cell wall characteristics on protein extractability and determination of nitrogen-to-protein conversion factors. J. Appl. Phycol. 2013, 25, 523–529. [Google Scholar] [CrossRef]
- Daniel, N.; Sivaramakrishnan, T.; Saravanan, K.; Shalini, B.; Arunjyoti, B.; Sankar, R.; Dann Roy, S. A review on microalgae as potential fish feed ingredient. J. Andeman Sci. Assoc. 2016, 1, 140–144. [Google Scholar]
- Bakhtiyar, Y.; Langer, S.; Karlopia, S.K.; Ahmed, I. Growth, survival and proximate body composition of Labeo rohita larvae fed artificial food and natural food organisms under laboratory condition. Int. J. Fish. Aquac. 2011, 3, 114–117. [Google Scholar]
- Radhakrishnan, S.; Saravana Bhavan, P.; Seenivasan, C.; Muralisankar, T. Effect of dietary replacement of fishmeal with Chlorella vulgaris on growth performance, energy utilization and digestive enzymes in Macrobrachium rosenbegii postlarvae. Int. J. Fish. Aquac. 2015, 7, 62–70. [Google Scholar]
- Berliner, M.D. Proteins in Chlorella vulgaris. Microbios 1986, 46, 199–203. [Google Scholar]
- Seyfabadi, J.; Ramezanpour, Z.; Amini Khoeyi, Z. Protein, fatty acid, and pigment content of Chlorella vulgaris under different light regimes. J. Appl. Phycol. 2011, 23, 721–726. [Google Scholar] [CrossRef]
- Lubitz, J.A. The protein quality, digestibility, and composition of micro-algae, Chlorella 71105. J. Food Sci. 1963, 28, 229–232. [Google Scholar] [CrossRef]
- Zeinhom, M.M. Nutritional and Physiological Studies on Fish. Ph.D. Thesis, Faculty of Agriculture, Zagazig University, Zagazig, Egypt, 2004. [Google Scholar]
- Shaaban, M. Green microalgae water extracts as foliar feeding to wheat plants. Pak. J. Biol. Sci. 2001, 4, 628–632. [Google Scholar]
- Shi, X.; Chen, F.; Chen, G.-H.; Pan, Y.-X.; Zhu, X.-M.; Liu, X.; Luo, Z. Fishmeal can be totally replaced by a mixture of rapeseed meal and chlorella meal in diets for crucian carp (Carassius auratus gibelio). Aquac. Res. 2017, 1–9. [Google Scholar] [CrossRef]
- Pakravan, S.; Akbarzadeh, A.; Sajjadi, M.M.; Hajimoracloo, A.; Noori, F. Chlorella vulgaris meal improved growth performance, digestive enzyme activities, fatty acid composition and tolerance of hypoxia and ammonia stress in juvenile Pacific white shrimp Litopenaeus vannamei. Aquac. Nutr. 2017. [Google Scholar] [CrossRef]
- Dawah, M.A.; Ibrahim, A.N.; Eladel, H.M. Amino acids content of Chlorella vulgaris and Scenedesmus bijuga raised on domestic sewage and agricultural drainage water. In Proceedings of the 1st Scientific Conference of the Egyptian Aquaculture Society, El-Arish, Egypt, 13–15 December 2002. [Google Scholar]
- Dawah, M.A.; Khater, A.M.; Shaker, I.M.A.; Ibrahim, N.A. Production of Scenedesmus bijuga (Chlorophyceae) in large scale in outdoor tanks and its use in feeding monosex Nile tilapia (Oreochromis niloticus) fry. J. Egypt. Acad. Soc. Environ. Dev. 2002, 2, 113–125. [Google Scholar]
- Van Der Meeren, T.; Mangor-Jensen, A.; Pickova, J. The effect of green water and light intensity on survival, growth and lipid composition in Atlantic cod (Gadus morhua) during intensive larval rearing. Aquaculture 2007, 265, 206–2173. [Google Scholar] [CrossRef]
- Wang, Y.; Hu, M.; Cao, L.; Yang, Y.; Wang, W. Effects of daphnia (Moina micrura) plus chlorella (Chlorella pyrenoidosa) or microparticle diets on growth and survival of larval loach (Misgurnus anguillicaudatus). Aquac. Int. 2008, 16, 361–368. [Google Scholar] [CrossRef]
- Cahu, C.L.; Zambonino Infante, J.L.; Péres, A.; Quazuguel, P.; Le Gall, M.M. Algal addition in sea bass (Dicentrarchus labrax) larvae rearing: Effect on digestive enzymes. Aquaculture 1998, 161, 479–489. [Google Scholar] [CrossRef]
- Güroy, D.; Güroy, B.; Merrifield, D.L.; Ergün, S.; Tekinay, A.A.; Yiğit, M. Effect of dietary Ulva and Spirulina on weight loss and body composition of rainbow trout, Oncorhynchus mykiss(Walbaum), during a starvation period. J. Anim. Physiol. Anim. Nutr. 2011, 95, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Nandeesha, M.C.; Gangadhar, B.; Varghese, T.J.; Keshavanath, P. Effect of feeding Spirulina platensis on the growth, proximate composition and organoleptic quality of common carp, Cyprinus carpio L. Aquac. Res. 1998, 29, 305–312. [Google Scholar] [CrossRef]
- Badwy, T.M.; Ibrahim, E.M.; Zeinhom, M.M. Partial replacement of fishmeal with dried microalga (Chlorella spp. and Scenedesmus spp.) in Nile tilapia (Oreochromis niloticus) diets. In Proceedings of the 8th International Symposium on Tilapia in Aquaculture, Cairo, Egypt, 12–14 October 2008; pp. 801–811. [Google Scholar]
- Miller, M.R.; Nichols, P.D.; Carter, C.G. n-3 Oil sources for use in aquaculture—Alternatives to the unsustainable harvest of wild fish. Nutr. Res. Rev. 2008, 21, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.Z.; Jauncey, K. Optimal dietary carbohydrate to lipid ratio in African catfish Clarias gariepinus (Burchell 1822). Aquac. Int. 2004, 12, 169–180. [Google Scholar] [CrossRef]
- Ematipour, G.R.; Brown, M.L.; Gatlin, D.M., III. Effects of dietary carbohydrate: Lipid ratio on growth and body composition of hybrid striped bass. J. World Aquac. Soc. 1992, 23, 128–132. [Google Scholar] [CrossRef]
- Enyidi, U.; Pirhonen, J.; Kettunen, J.; Vielma, J. Effect of Feed Protein:Lipid Ratio on Growth Parameters of African Catfish Clarias gariepinus after Fish Meal Substitution in the Diet with Bambaranut (Voandzeia subterranea) Meal and Soybean (Glycine max) Meal. Fishes 2017, 2, 1. [Google Scholar] [CrossRef]
- Tan, Q.; Xie, S.; Zhu, X.; Lei, W.; Yang, Y. Effect of dietary carbohydrate- to-lipid ratios on the growth and feed utilization in Chinese longsnout catfish (Leiocassis longirostris Gunther). J. Appl. Ichthyol. 2007, 23, 605–610. [Google Scholar] [CrossRef]
- Luo, Y.; Xie, X. Effects of high carbohydrate and high lipid diets on growth, body composition and glucose metabolism in southern catfish at two temperatures. Aquac. Res. 2010, 41, e431–e437. [Google Scholar] [CrossRef]
- Satoh, K.-I.; Nakagawa, H.; Kasahara, S. Effect of ulva meal supplementation on disease resistance of red sea bream. Nippon Suisan Gakkaishi 1987, 53, 1115–1120. [Google Scholar] [CrossRef]
- Rocha, R.J.; Ribeiro, L.; Costa, R.; Dinis, M.T. Does the presence of microalgae influence fish larvae prey capture? Aquac. Res. 2008, 39, 362–369. [Google Scholar] [CrossRef]
- Ogawa, T.; Aiba, S. Bioenergetic analysis of mixotrophic growth in Chlorella Vulgaris and Scenedesmus acutus. Biotechnol. Bioeng. 1981, 23, 1121–1132. [Google Scholar] [CrossRef]
- Liang, Y.; Sarkany, N.; Cui, Y. Biomass and lipid productivities of Chlorella vulgaris under autotrophic, heterotrophic and mixotrophic growth conditions. Biotechnol. Lett. 2009, 31, 1043–1049. [Google Scholar] [CrossRef] [PubMed]
- Parrish, C.C. Determination of total lipid classes and fatty acids in aquatic samples. In Lipids in Freshwater Ecosystems; Wetzel, R.G., Art, M.T., Wainmann, B.C., Eds.; Springer: New York, NY, USA, 1999; pp. 4–20. [Google Scholar]
- Kainz, M.; Arts, M.; Mazumder, A. Essential fatty acids in the planktonic food web and their ecological role for higher trophic level. Limnol. Oceanogr. 2004, 49, 1784–1793. [Google Scholar] [CrossRef]
Figure 1.
Relationship between the specific growth rate (SGR)of African catfish C. gariepinus fed with algae C. vulgaris diets as substitute of fishmeal for 60 d.
Figure 1.
Relationship between the specific growth rate (SGR)of African catfish C. gariepinus fed with algae C. vulgaris diets as substitute of fishmeal for 60 d.

Figure 2.
Relationship of protein conversion ratio and food conversion ratio (FCR) of African catfish C. gariepinus fed diets varying Chlorella vulgaris meal and fishmeal for 60 d.
Figure 2.
Relationship of protein conversion ratio and food conversion ratio (FCR) of African catfish C. gariepinus fed diets varying Chlorella vulgaris meal and fishmeal for 60 d.


{kind=link}
{kind=link}
{kind=link}
Table 1.
The growth and nutrient utilization of African catfish Clarias gariepinus fed diets varying in composition of freshwater algae as follows: F1, 25%; F2, 15%; F3, 5%, and; F4 0% for 60 days.
Table 1.
The growth and nutrient utilization of African catfish Clarias gariepinus fed diets varying in composition of freshwater algae as follows: F1, 25%; F2, 15%; F3, 5%, and; F4 0% for 60 days.
| Feed | F1 | F2 | F3 | F4 |
|---|---|---|---|---|
| Ini. no | 30 | 30 | 30 | 30 |
| Final no | 30 | 30 | 30 | 30 |
| Ini. Av. Wt | 1.09 ± 0.05 | 1.09 ± 0.05 | 1.09 ± 0.05 | 1.09 ± 0.05 |
| Fin. Av. Wt | 122.11 ± 2.51 a | 109.32 ± 1.09 b | 90.34 ± 0.55 c | 63.59 ± 1.30 d |
| FCR | 1.92 ± 0.05 a | 1.88±0.02 a | 2.22 ± 0.11 b | 2.98 ± 0.01 c |
| AWG | 121.02 ± 0.04 a | 108.23 ± 0.06 b | 89.25 ± 0.02 c | 62.50 ± 0.01 d |
| SGR | 7.86 ± 0.45 a | 7.68 ± 0.21 a | 7.36 ± 0.44 a | 6.77 ± 0.07 b |
| HSI | 1.48 ± 0.01 a | 1.53 ± 0.1 a | 1.87 ± 0.08 b | 2.50 ± 0.59 c |
| PCR | 0.61 ± 0.02 a | 0.60 ± 0.03 a | 0.69 ± 0.11 b | 0.95 ± 0.23 c |
| PER | 2.46 ± 0.22 b | 2.30 ± 0.13 b | 2.07 ± 0.03 c | 2.02 ± 0.09 a |
| DFI | 3.88 ± 0.15 a | 3.39 ± 0.04 b | 3.30 ± 0.24 b | 3.11 ± 0.08 b |
| DMR | 7.95 ± 0.12 a | 8.11 ± 0.43 a | 8.84 ± 0.37 b | 12.34 ± 0.22 c |
| WPR | 1.54 ± 0.10 a | 1.5 ± 0.22 a | 1.81 ± 0.32 b | 2.50 ± 0.21 c |
Ini. no = initial number of fish, Final no = final number of fish, Ini. Av. Wt = initial average weight, Fin. Av. Wt = final average weight, FCR = food conversion ratio, AWG = average weight gain, SGR = specific growth rate, HSI = hepatosomatic index, PCR = protein conversion ratio, PER = protein efficiency ratio, WPR = waste production ratio, DMR = dry matter ratio and DFI = daily feed intake. Means not followed by same superscripts are significantly different p < 0.05.
Table 2.
Composition of experimental diets varying in the composition of freshwater algae Chlorella vulgaris used in feeding African catfish C. gariepinus for 60 days.
Table 2.
Composition of experimental diets varying in the composition of freshwater algae Chlorella vulgaris used in feeding African catfish C. gariepinus for 60 days.
| Items | Feeds | |||
|---|---|---|---|---|
| F1 | F2 | F3 | F4 | |
| Fishmeal | 0 | 0 | 0 | 15 |
| Algae | 25 | 15 | 5 | 0 |
| Corn meal | 40 | 43 | 53 | 43 |
| Millet meal | 23 | 30 | 30 | 30 |
| Bone meal | 5 | 5 | 5 | 5 |
| Palm oil | 5 | 5 | 5 | 5 |
| Vitamin premix | 2 | 2 | 2 | 2 |
| Total | 100 | 100 | 100 | 100 |
| Proximate composition of diets | ||||
| Crude Protein % | 32.0 | 31.8 | 31.2 | 31.9 |
| Starch g | 200 | 198 | 204 | 204 |
| Crude lipids (%) | 11.9 | 11.8 | 11.6 | 11.2 |
| Ash g | 89 | 92 | 89 | 84 |
| Moisture% | 8.7 | 8.5 | 8.0 | 8.7 |
Vitamin Premix. The following vitamins were added to supply the following kg−1 diet: cholecalciferol, 1300 IU; all-race-α-tocopheryl acetate, 140 IU; menadione sodium bisulfite, 12 mg; thiamin HCl, 8 mg; riboflavin, 16 mg; calcium d-pantothenate, 17 mg; biotin, 0.2 mg; folic acid, 5 mg; vitamin B12, 0.02, niacin, 40 mg; pyridoxine HCl, 16 mg; ascorbic acid (Stay C), 80 mg, magnesium phosphate, 5000 mg, potassium carbonate, 400 mg, manganous sulfate, 10; ferrous sulfate, 5 mg; zinc sulfate, 80 mg.
Table 3.
Proximate composition of C. vulgaris and millet used in formulating diet for African catfish C. gariepinus.
Table 3.
Proximate composition of C. vulgaris and millet used in formulating diet for African catfish C. gariepinus.
| Protein | Carbohydrate | Lipids | |
|---|---|---|---|
| Millet | 20.68 ± 1.55 | 49.50 ± 3.17 | 3.64 ± 0.06 |
| C. vulgaris | 54.65 ± 0.07 | 12.09 ± 3.17 | 24.40 ± 0.09 |
© 2017 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Enyidi, U.D. Chlorella vulgaris as Protein Source in the Diets of African Catfish Clarias gariepinus. Fishes 2017, 2, 17. https://doi.org/10.3390/fishes2040017
AMA Style
Enyidi UD. Chlorella vulgaris as Protein Source in the Diets of African Catfish Clarias gariepinus. Fishes. 2017; 2(4):17. https://doi.org/10.3390/fishes2040017
Chicago/Turabian StyleEnyidi, Uchechukwu D. 2017. "Chlorella vulgaris as Protein Source in the Diets of African Catfish Clarias gariepinus" Fishes 2, no. 4: 17. https://doi.org/10.3390/fishes2040017