Improved Growth Estimates for Lethrinus harak: Measuring Increments, Adjusting Ages, and Fitting Flexible Growth Models

 ,
,
Abstract
:1. Introduction
2. Results
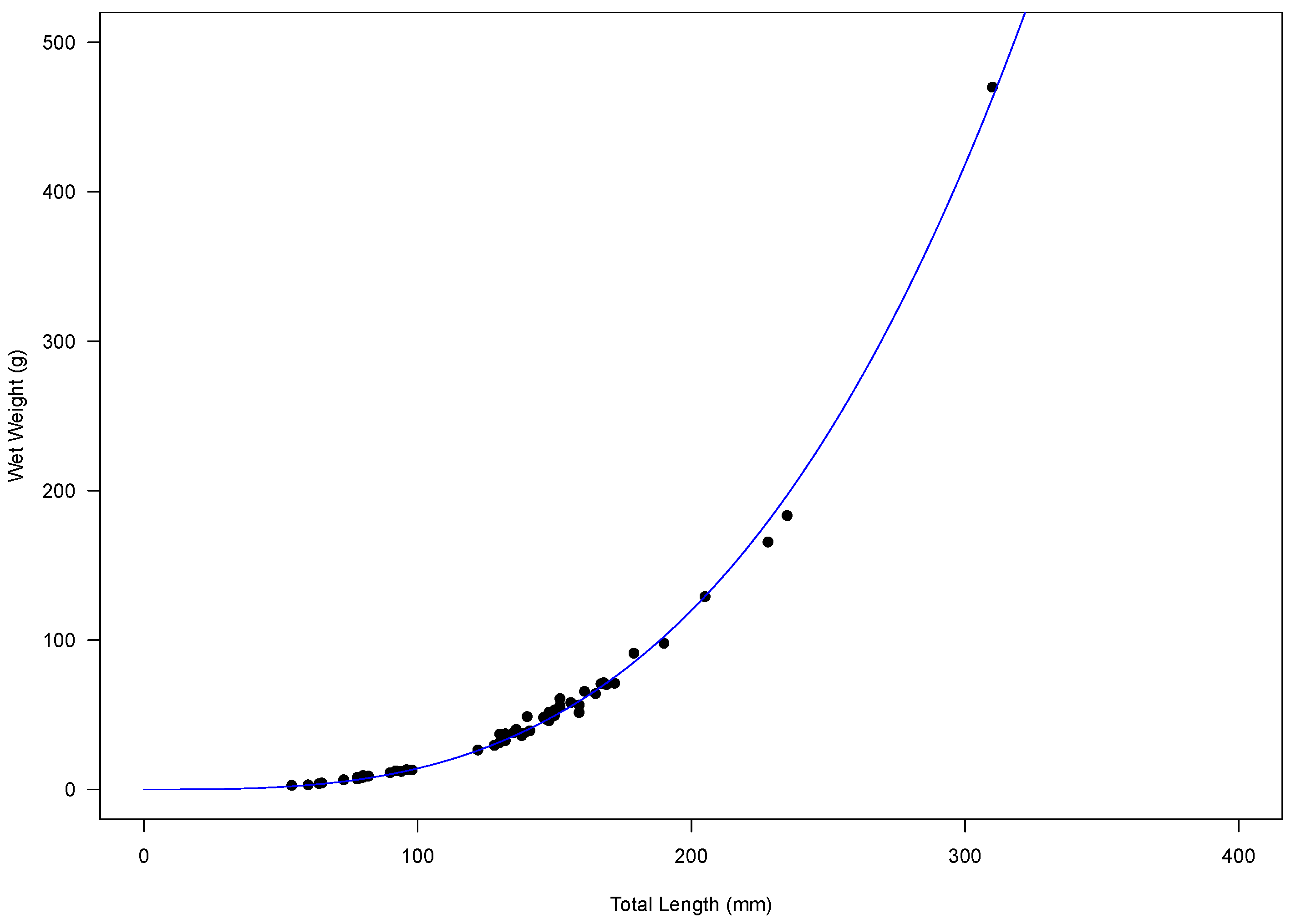
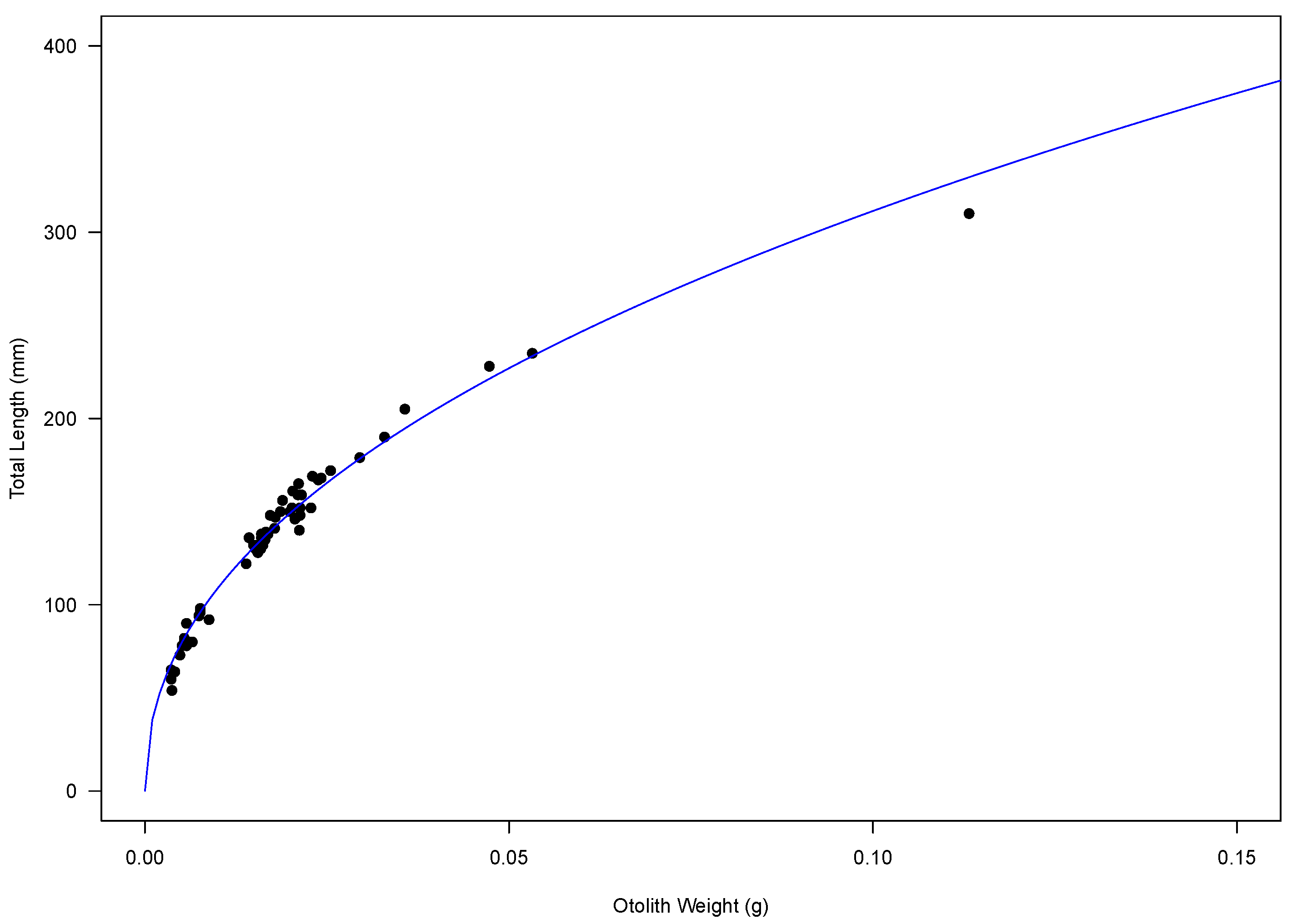
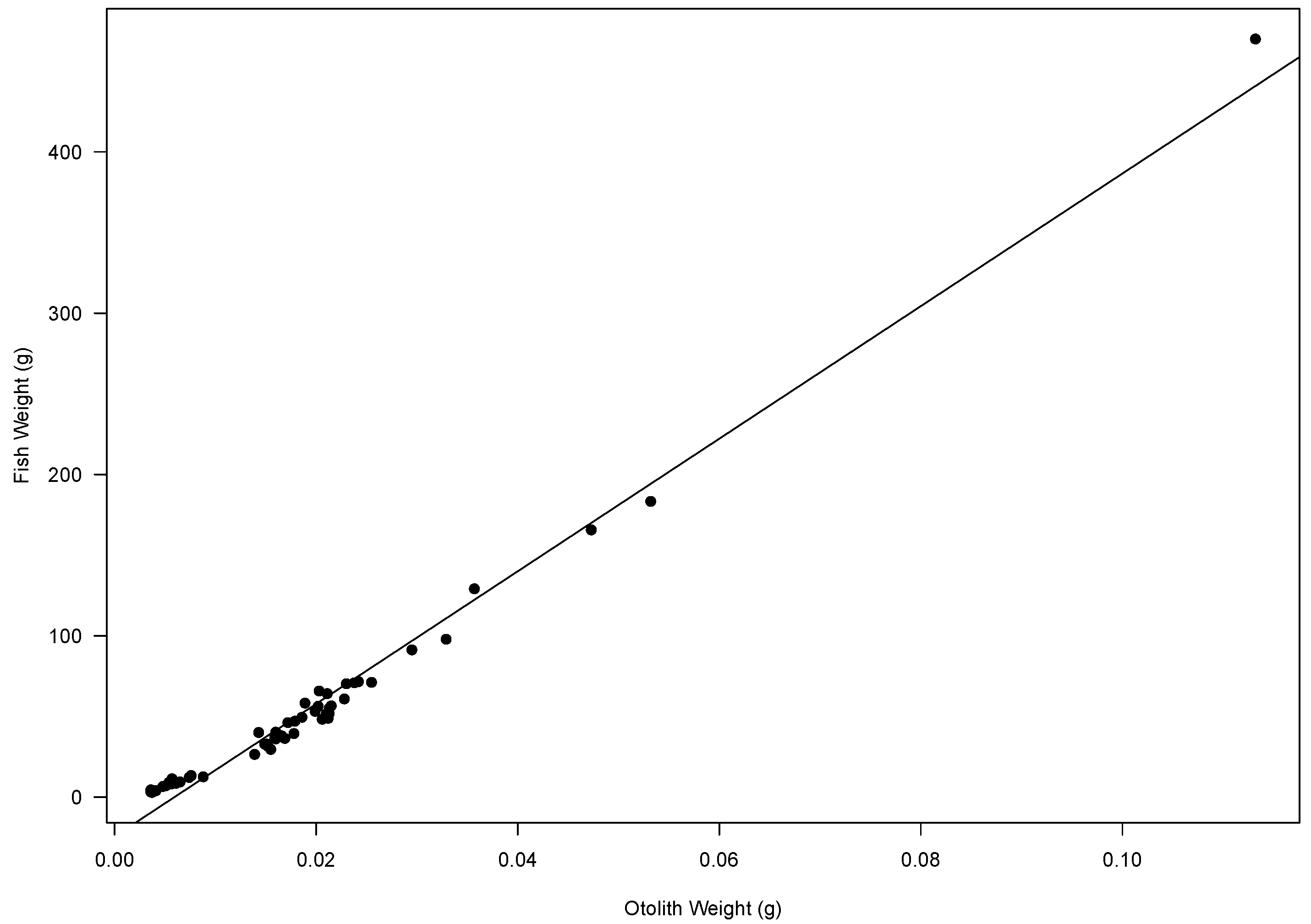
2.1. Age Estimation and Otolith Relationships
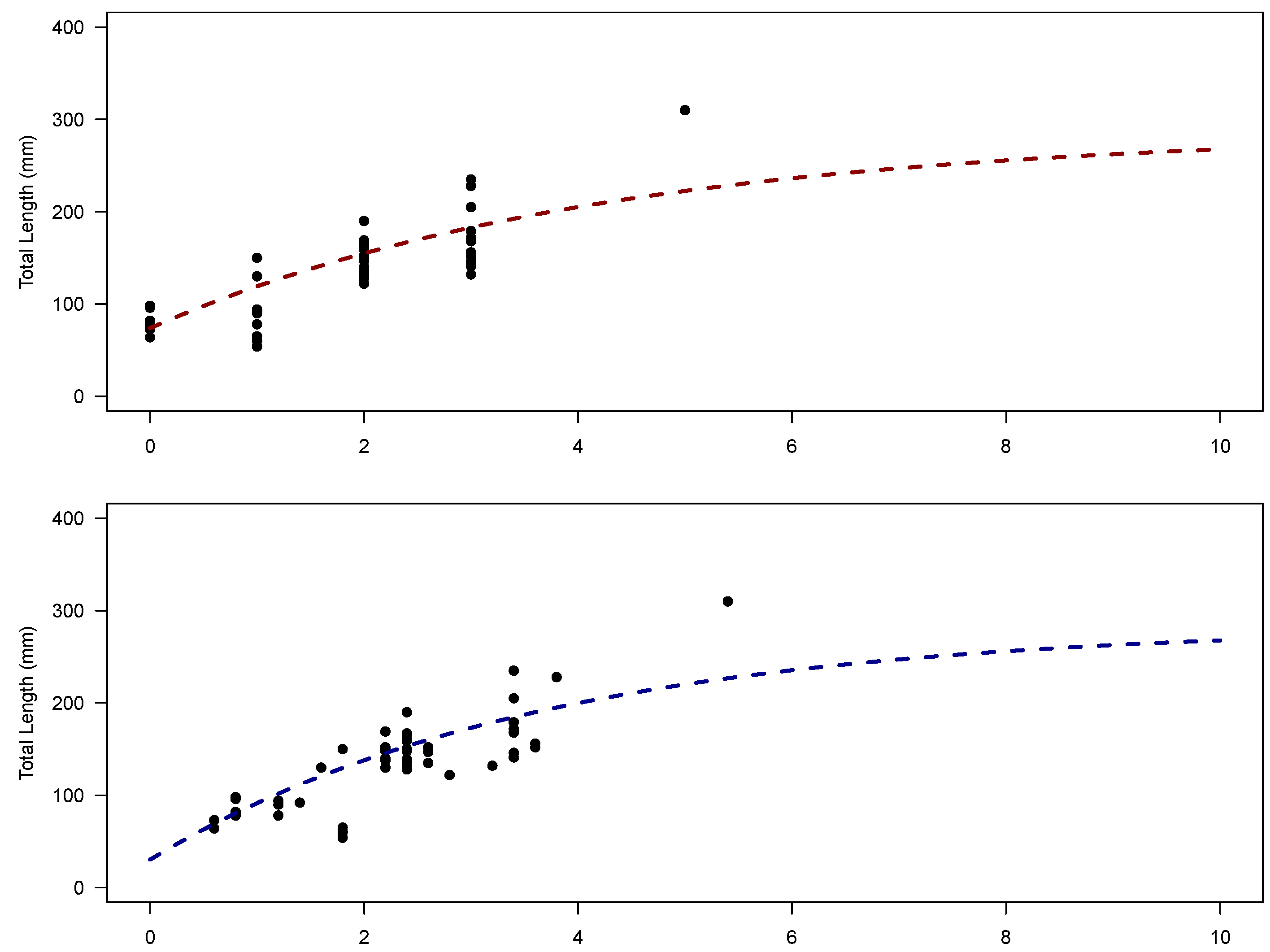
2.2. Growth
3. Discussion
Strengths and Limitations
4. Materials and Methods
4.1. Collection of Samples
4.2. Otolith Analyses
4.3. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jacquet, J.; Fox, H.; Motta, H.; Ngusaru, A.; Zeller, D. Few data but many fish: Marine small-scale fisheries catches for Mozambique and Tanzania. Afr. J. Mar. Sci. 2010, 32, 197–206. [Google Scholar] [CrossRef]
- Jiddawi, N.; Öhman, M. Marine fisheries in Tanzania. J. Hum. Environ. 2002, 31, 518–527. [Google Scholar] [CrossRef]
- Bush, E.R.; Short, R.E.; Milner-Gulland, E.J.; Lennox, K.; Samoilys, M.; Hill, N. Mosquito net use in an artisanal East African fishery. Conserv. Lett. 2016, 10, 451–459. [Google Scholar] [CrossRef]
- King, J.; McFarlane, G. Marine fish life history strategies: Applications to fishery management. Fish. Manag. Ecol. 2003, 10, 249–264. [Google Scholar] [CrossRef]
- Carpenter, K. Lethrinidae: Emperor (emperor snappers). In The Living Marine Resources of the Western Central Pacific; Carpenter, K., Niems, V., Eds.; Bony fishes, part 3 (Menidae to Pomacentridae); FAO: Rome, Italy, 2001; Volume 5, pp. 3004–3050. [Google Scholar]
- Sobo, F. Community participation in fisheries management in Tanzania. In Proceedings of the IIFET 2012 Tanzania Proceedings, Dar es Salaam, Tanzania, 16–20 July 2012. [Google Scholar]
- Lasi, F. Age, Growth, Mortality Rates and Reproductive Biology of Three Fijian Emperor Fishes (Pisces: Lethrinidae) Lethrinus harak. Master’s Thesis, University of the South Pacific, Suva, Fiji, 2003. [Google Scholar]
- Ebisawa, A.; Ozawa, T. Life-history traits of eight Lethrinus species from two local populations in waters off the Ryukyu Islands. Fish. Sci. 2009, 75, 553–566. [Google Scholar] [CrossRef]
- Nanami, A.; Yamada, H. Site fidelity, size, and spatial arrangement of daytime home range of thumbprint emperor Lethrinus harak (Lethrinidae). Fish. Sci. 2009, 75, 1109–1116. [Google Scholar] [CrossRef]
- Trianni, M.S. Life history characteristics and stock status of the thumbprint emperor (Lethrinus harak) in Saipan Lagoon. Fish. Bull. 2016, 114, 409–425. [Google Scholar] [CrossRef]
- Kulmiye, A.; Ntiba, M.; Kisia, S. Some aspects of the reproductive biology of the thumbprint emperor, Lethrinus harak (Forsskål, 1775), in Kenyan coastal waters. West. Indian Ocean J. Mar. Sci. 2002, 1, 135–144. [Google Scholar]
- Hilomen, V.V. Inter- and Intra-habitat Movement Patterns and Population Dynamics of Small Reef Fishes of Commercial and Recreational Significance. Ph.D. Dissertation, James Cook University, Townsville, Australia, 1997. [Google Scholar]
- Ebisawa, A. Reproductive and sexual characteristics in five Lethrinus species in waters off the Ryukyu Islands. Ichthyol. Res. 2006, 53, 269–280. [Google Scholar] [CrossRef]
- Ricker, W.E. Computation and interpretation of biological statistics of fish populations. Bull. Fish. Res. Board Can. 1975, 191, 382. [Google Scholar]
- Kimirei, I.; Nagelkerken, I.; Slooter, N.; Gonzalez, E.; Huijbers, C.; Mgaya, Y.; Rypel, A. Demography of fish populations reveals new challenges in appraising juvenile habitat values. Mar. Ecol. Prog. Ser. 2015, 518, 225–237. [Google Scholar] [CrossRef] [Green Version]
- Pilling, G.M.; Millner, R.S.; Easey, M.W.; Mees, C.C.; Rathacharen, S.; Azemia, R. Validation of annual growth increments in the otoliths of the lethrinid Lethrinus mahsena and the lutjanid Aprion virescens from sites in the tropical Indian Ocean, with notes on the nature of growth increments in Pristipomoides filamentosus. Fish. Bull. 2000, 98, 600. [Google Scholar]
- Fowler, A.J. Validation of annual growth increments in the otoliths of a small, tropical coral reef fish. Mar. Ecol. Prog. Ser. 1990, 64, 25–38. [Google Scholar] [CrossRef]
- Pilling, G.; Grandcourt, E.; Kirkwood, G. The utility of otolith weight as a predictor of age in the emperor Lethrinus mahsena and other tropical fish species. Fish. Res. 2003, 60, 493–506. [Google Scholar] [CrossRef]
- Francis, R.C.; Campana, S.E. Inferring age from otolith measurements: A review and a new approach. Can. J. Fish. Aquat. Sci. 2004, 61, 1269–1284. [Google Scholar] [CrossRef]
- Pentilla, J. Age Determination Methods for Northwest Atlantic Species: Technical Report 72. Atlantic Cod, Gadus morhua; Pentilla, J., Dery, L., Eds.; National Oceanic and Atmospheric Administration: Silver Spring, MA, USA, 1988; pp. 31–36.
- Harris, P.J.; Wyanski, D.M.; White, D.B.; Mikell, P.P.; Eyo, P.B. Age, growth, and reproduction of greater amberjack off the southeastern US Atlantic coast. Trans. Am. Fish. Soc. 2007, 136, 1534–1545. [Google Scholar] [CrossRef]
- Midway, S.R.; Wagner, T.; Arnott, S.A.; Biondo, P.; Martinez-Andrade, F.; Wadsworth, T.F. Spatial and temporal variability in growth of southern Flounder (Paralichthys lethostigma). Fish. Res. 2015, 167, 323–332. [Google Scholar] [CrossRef]
- Plummer, M. rjags: Bayesian Graphical Models using MCMC; CRAN: Vienna, Austria, 2016. [Google Scholar]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Charnov, E.L. Fish growth: Bertalanffy k is proportional to reproductive effort. Environ. Biol. Fishes 2008, 83, 185–187. [Google Scholar] [CrossRef]
- Prince, J.; Hordyk, A.; Valencia, S.R.; Loneragan, N.; Sainsbury, K. Revisiting the concept of Beverton–Holt life-history invariants with the aim of informing data-poor fisheries assessment. ICES J. Mar. Sci. 2014, 72, 194–203. [Google Scholar] [CrossRef]
- Pauly, D. On the interrelationships between natural mortality, growth parameters, and mean environmental temperature in 175 fish stocks. ICES J. Mar. Sci. 1980, 39, 175–192. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Measure | 1st Annulus Length | 2nd Annulus Length | 3rd Annulus Length | 4th Annulus Length | 5th Annulus Length |
|---|---|---|---|---|---|
| Mean | 0.97 | 1.40 | 1.57 | 2.12 | 2.38 |
| Standard deviation | 0.15 | 0.12 | 0.13 | – | – |
| Minimum | 0.63 | 1.16 | 1.47 | 2.12 | 2.38 |
| Maximum | 1.28 | 1.65 | 1.90 | 2.12 | 2.38 |
| n | 46 | 37 | 12 | 1 | 1 |
| Study | (mm) | K | Location | |
|---|---|---|---|---|
| Unadjusted ages (this study) | 291 (210, 395) | 0.23 (0.13, 0.41) | (,) | Tanzania |
| Adjusted ages (this study) | 278 (200, 383) | 0.29 (0.15, 0.53) | (, 0.1) | Tanzania |
| Ebisawa and Ozawa (2009) | 284 | 0.509 | Japan | |
| Lasi (2003), constrained | 285 | 0.90 | Fiji | |
| Lasi (2003), unconstrained | 325 | 0.27 | Fiji | |
| Trianni (2016), combined sexes | 301 | 0.259 | Saipan |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Midway, S.R.; Ostrowski, A.; West, L.; Hernandez, M.; Robertson, M.D. Improved Growth Estimates for Lethrinus harak: Measuring Increments, Adjusting Ages, and Fitting Flexible Growth Models. Fishes 2018, 3, 31. https://doi.org/10.3390/fishes3030031
Midway SR, Ostrowski A, West L, Hernandez M, Robertson MD. Improved Growth Estimates for Lethrinus harak: Measuring Increments, Adjusting Ages, and Fitting Flexible Growth Models. Fishes. 2018; 3(3):31. https://doi.org/10.3390/fishes3030031
Chicago/Turabian StyleMidway, Stephen R., Andrew Ostrowski, Lindsey West, Mario Hernandez, and Matthew D. Robertson. 2018. "Improved Growth Estimates for Lethrinus harak: Measuring Increments, Adjusting Ages, and Fitting Flexible Growth Models" Fishes 3, no. 3: 31. https://doi.org/10.3390/fishes3030031
APA StyleMidway, S. R., Ostrowski, A., West, L., Hernandez, M., & Robertson, M. D. (2018). Improved Growth Estimates for Lethrinus harak: Measuring Increments, Adjusting Ages, and Fitting Flexible Growth Models. Fishes, 3(3), 31. https://doi.org/10.3390/fishes3030031