1. Introduction
Invasive species are a major source of biodiversity loss impacting multiple levels of ecosystems composition and structure. These impacts include hybridizing with native species, proliferating foreign pathogens, and enacting competitive displacement of native species, with the most visible impact being, perhaps, predation [
1]. The introduction of top predators in freshwater systems has the potential to reduce or extirpate populations of native species at a local level [
2]. For Portugal, comprehensive studies focusing on the diet of invading predators are limited to two species, namely, the largemouth bass (
Micropterus salmoides, Lacépède, 1802) [
3] and the pikeperch (
Sander lucioperca, L., 1758) [
4]. The rate of fish introduction in Portugal is high, with the most recent introductions related to sport angling with two top fish predators [
5,
6,
7]. It is therefore important to evaluate the impacts of predatory fishes in order to evaluate the direct impact on aquatic organisms, especially species with conservation concerns.
The most recent top predator introduced in Portuguese watersheds,
Silurus glanis, was described as a large-sized opportunistic omnivorous predator [
8] with acute predatory senses that make it an apex predator [
9]. It feeds mostly on benthic organisms and is able to adapt its diet to the available spectrum of prey items found within a system [
10,
11]. Typically, the predominant prey type found in the
S. glanis diet is whichever prey is more abundant in the system, provided its size and habitat use are suitable for
S. glanis [
12]. Its capacity to adapt to different prey types is remarkable, such as preying on birds found in river banks in regions of France and Spain [
13].
Works concerning the European catfish diet habits were mostly done within its native range, where the fish community structure and environmental conditions are very different than those found in Portuguese rivers [
14]. For the introduced range, few works are present in the literature, which are either preliminary or based on low sample numbers (e.g., [
15,
16]); nevertheless, they provide valuable information indicating that, in the invasive context,
S. glanis seems to continue to be highly adaptable to the spectrum of prey available and incorporates native species into its diet [
17]. In Spain, a study by Carol et al. (2009) [
18] focused on reservoir populations and their diets, observing that the basis of the diet consisted of crustaceans, mostly crayfish (
Procambarus clarkii, Girard, 1852), with fish prey representing the largest portion of biomass consumed, mostly non-native species. More recently, Guillerault et al. (2017) [
16] showed that this species predates in migratory fish, so its impact could be larger than expected. This fact was previously supported by Syväranta et al. (2010) [
17], who found evidence of the European eel (
Anguilla anguilla, L., 1758) being part of this species’ diet in France.
The previously described
S. glanis habits are worrying in the Portuguese context, because this species invaded the Tagus River drainage, arriving to the lower Tagus in 2006 [
6,
19]. Some threatened native fish, such as the European eel, the long-snouted barbel (
Luciobarbus comizo, Steindachner, 1864), and the sea lamprey (
Petromyzon marinus, L., 1758), occupy benthic areas, the same spatial niche as
S. glanis, making them potential prey for this species and adding to the pressure these species’ populations are already under. In addition, Portuguese watersheds are not home to any native piscivorous fish [
20], so the introduction of predators of this kind may have its impacts enlarged by a form of fish naivety [
2]. Furthermore, the water temperature in Portugal is generally higher than in its native range, so a higher metabolism, possibly related to higher activity levels, is expected, and associated with a larger impact on aquatic communities in the rivers where it occurs [
14].
The Tagus is the longest river in the Iberian Peninsula. A river with great historical importance, it was once one of the main routes for the trade of goods between both Iberian countries. Despite retaining some of its economic importance, traversing the river is now nearly impossible due to the many dams and weirs found along its course, with the main objective of energy production [
21]. Due to its great extension, the river crosses many different ecosystems of extreme ecological importance, like the Spanish plateau, the Mediterranean forest valleys, and the marshlands close to its mouth. A place of refuge during the last ice ages, it is home to many endemic species, many of which are threatened, like the long-snouted barbel or the Lisbon arched-mouth nase (
Iberochondrostoma olisiponense) [
22]. It is also an important habitat for diadromous species like the European eel, the sea lamprey, the Allis shad (
Alosa alosa, L. 1758), and the Twait shad (
Alosa fallax, Lacépède, 1800). Sadly, in recent years, ever-increasing episodes of severe pollution, drought, and the arrival of invasive species have accentuated the loss of this rich biodiversity [
22]. The aim of this work is to describe the dietary habits of
S. glanis, an apex predator that recently arrived to this region with a unique fauna. This information is essential to demonstrate the species impact and move forward toward the management of this species.
2. Results
In total, of the 96 fishes captured, 35 were collected in lentic and the remaining 61 in lotic habitats of Tagus River. Fish where no prey was found were discarded for the remaining analysis, leaving us with a final sample size of 28 for lentic habitats and 38 for the lotic habitats (
Table 1). Overall, the mean length in each size class presented little variation between environments (
Table 1).
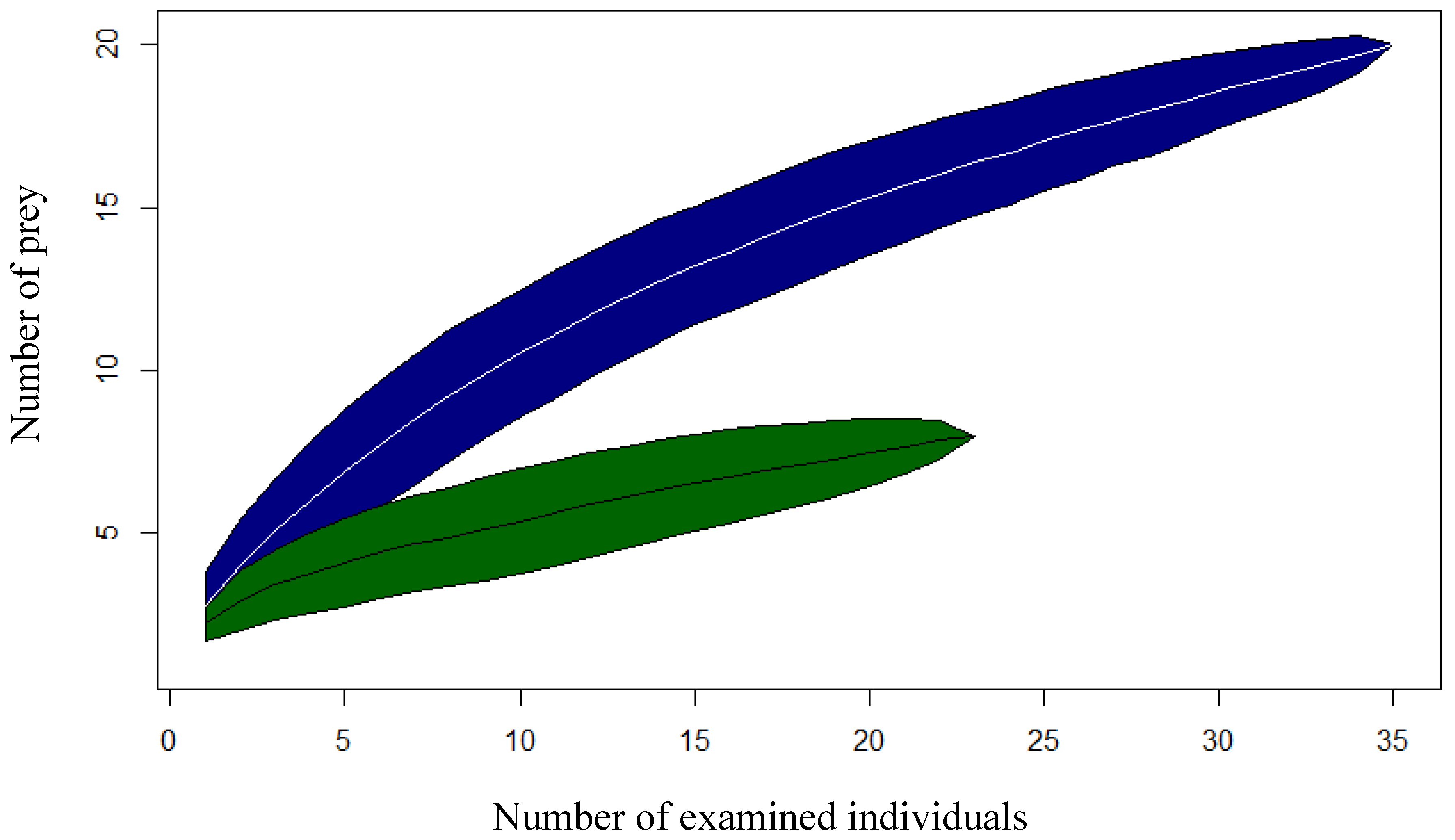
Prey cumulation curves showed that the number of prey items continued to increase as more stomachs were analyzed and did not stabilize with our studied number of individuals (
Figure 1). The more accentuated curvature was observed for the lotic system when compared to the lentic system, thereby indicating how much broader the dietary niche seems to be in this environment.
The diet of the fishes captured in the lotic section differed greatly from that of the fishes from the lentic sectors (
Table 2). Despite both relying heavily on crustaceans, the lentic fish ate almost solely based on this prey type. These fish also displayed a much larger red swamp crayfish (
Procambarus clarkii (Girard, 1852)) component when compared to the lotic system (Mann–Whitney U: W = 499; N
lotic = 38; N
lentic = 28; P < 0.001). The lotic fish composition (18.3%), consisted mostly of benthic fish (75.4% of fish prey found), but also of a portion of pelagic fishes (24.6% of fish prey found), which exceeded the number of fish prey found in the lentic system (Mann–Whitney U: W = 533; N
lotic = 38; N
lentic = 28; P < 0.01). In the lentic sample, fish prey represented less than 10% of the total number of prey found and came from only two of the individuals sampled. In the lentic environment, aside from the freshwater shrimp (
Atyaephyra desmaresti, Millet, 1831) and amphipods, the remainder of the diet was completely constituted by non-native species.
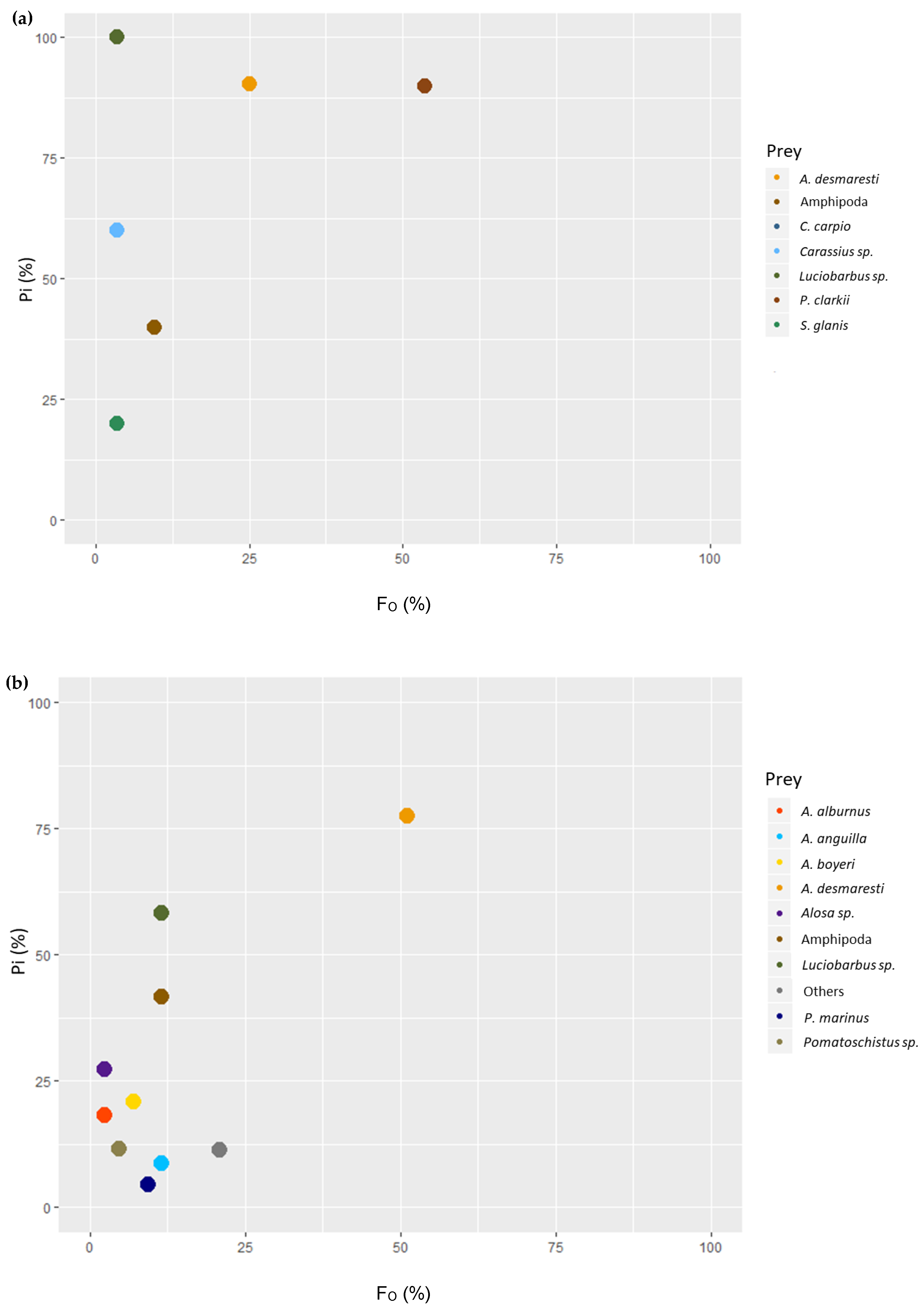
In the lentic environment, some degree of specialization toward the consumption of crustaceans was observed, while fish prey were rare (
Figure 2). As for the lotic environment, a broad dietary niche seemed to form due to the number of different fish species found without any of them becoming dominant in the diet; however, the same degree of specialization was observable toward the most abundant prey,
A. desmaresti (
Figure 2).
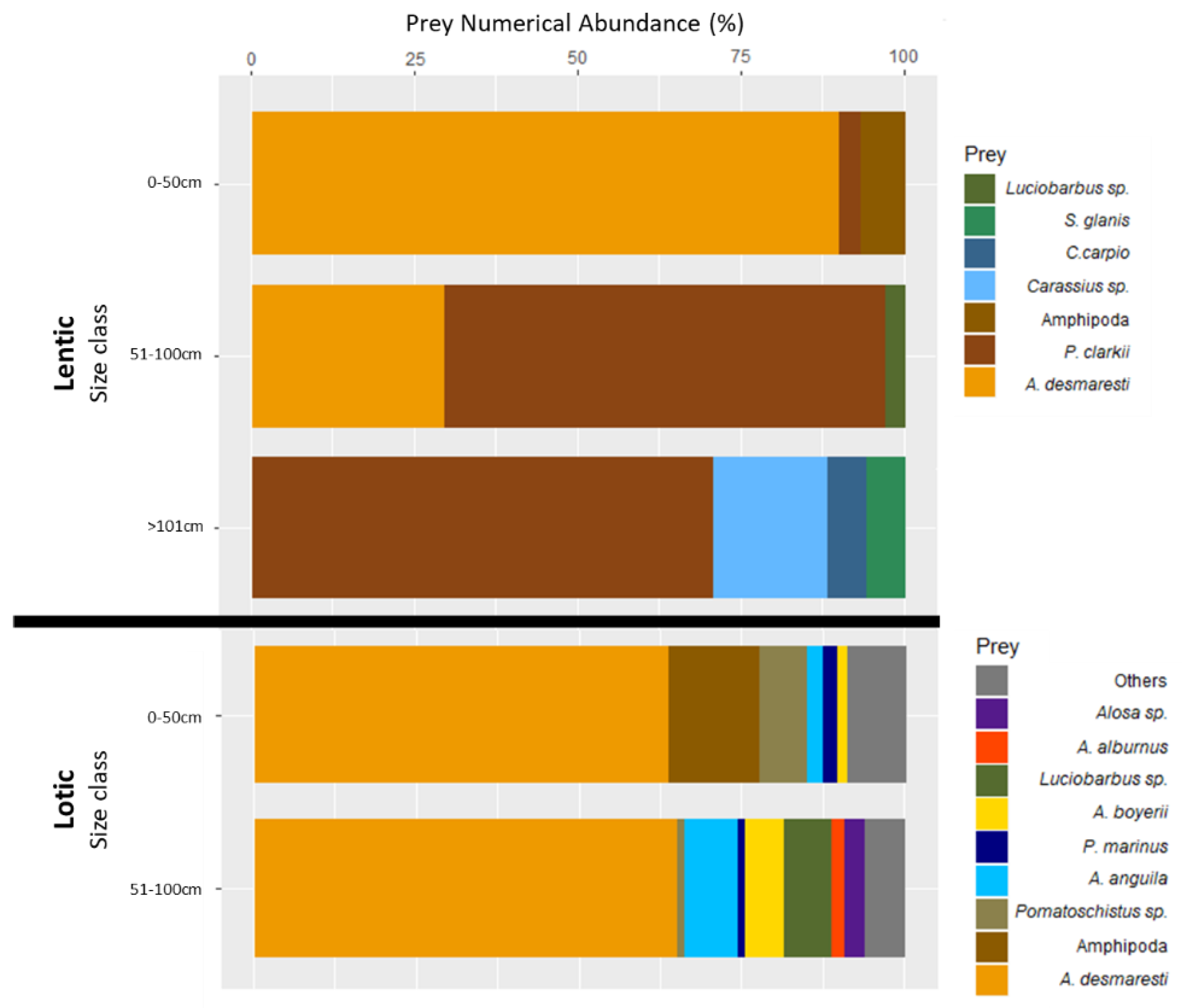
In the lentic environments, small-sized fish (Class 1) had a diet that consisted almost exclusively of crustaceans, nearly 90%, while in the lotic environment it was about 65%, also complemented with small fishes (
Figure 3). However, while in the lotic environment the main crustacean prey was
A. desmaresti, in the lentic environment it was crayfish, being dominant in Class 2 and Class 3. Furthermore, we observed remarkable prey diversity in the lotic environment, with small European catfish eating mostly benthic fish (such as eel), while medium-sized fishes had a larger proportion of pelagic fish prey (such as shad,
Alosa sp.) (
Figure 3). The Mann–Whitney U test revealed no differences between Class 1 and Class 2 for the lotic system, however, it did show significant differences between Class 1 and Class 2 in the lentic system in the amount of
A. desmaresti consumed (Mann–Whitney U: W = 52; N
lentic,C1 = 8; N
lentic,C2 = 15; P < 0.05) and of crayfish (Mann–Whitney U: W = 11; N
lentic,C1 = 8; N
lentic,C2 = 15; P < 0.05).
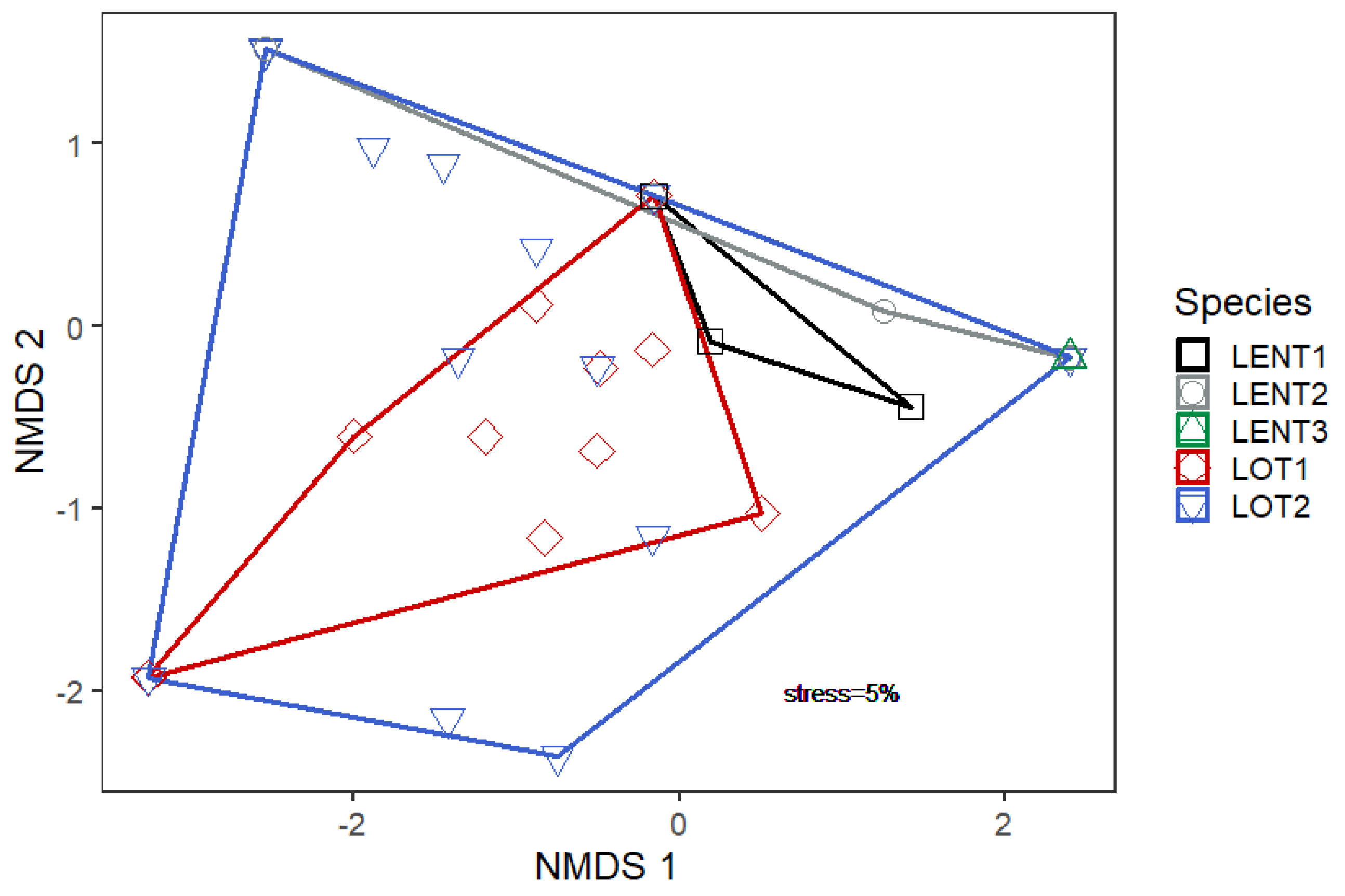
The NMDS (
Figure 4) showed that fishes in the lotic system exhibited greater prey diversity than those in the lentic system. As size increased in the lotic system, there was an expansion in prey diversity, with medium-sized individuals’ (LOT2) prey spectrum overlapping that of small-sized individuals (LOT1). For the lentic system, the different size classes exhibited diverging predatory niches.
3. Discussion
This work constitutes the first dietary study of European catfish in Portuguese freshwaters, assessing feeding behavior and potential impact in two different habitats in the Lower Tagus region. This species appropriates the most common food resource available in each habitat. In the lentic environment, non-native fish eat predominantly crustaceans, such as the freshwater shrimp and red swamp crayfish, and in lotic systems the European catfish ate mostly freshwater shrimp, closely followed by fish. Piscivory tended to increase with individual size in both studied habitats, and its feeding behavior was considerably broader in the lotic environment. This first set of data showed that non-native predators presented different impacts depending on their environment; in this case, the European catfish could have an important impact on the fish communities in the lotic habitats of the Lower Tagus River.
This study presents preliminary data regarding species impact in the Lower Tagus River. Firstly, this study relied on fishermen’s fishing efforts (predominately in fishing season, i.e., February and July) and, consequently, the number of fish captured varied along the year, with the bulk of the data coinciding with spring and early summer. However, the fish during the winter presented low activity [
23] and, most likely, any collected European catfish would present low number of prey items and prey diversity. Additionally, the size of the collected fish was highly dependent on the fishing net mesh size. Nevertheless, the observed tendency for piscivory seemed to be consistent in both studied habitats and comparable with other studies (e.g., [
17,
18]).
The lotic habitats generally exhibit communities that are more diverse, which increases the available prey diversity for
S. glanis and allows for a wider dietary niche. Verissimo et al. (2018) [
22] documented a total of 14 fish species in the Lower Tagus. The European catfish in the Tarn river, a tributary of the Garrone in France, seems to also feed on most of the available species found there [
17]. Most of the diet in the lotic habitat consisted of native species, with
A. desmarestii, a small freshwater shrimp, being the most abundant prey (over 60%). The presence of prey like
A. anguilla,
Alosa sp., and
P. marinus is worrying for two main reasons. Firstly, the populations of these three species have seen a sharp decline over the past few decades; therefore, added pressure on these populations may further aggravate this trend [
24,
25]. In addition, these species are central to the economy of anglers and fisheries, as well as the restaurants that depend on them across the Tagus River, and their decline will most definitely affect the future prospects of these commercial activities. In fact, Guillerault et al. (2017) [
16] mentioned that the European catfish could represent an important predatory pressure on migratory fish, namely shad and lamprey.
In lentic habitats, the diet consisted mostly of invasive species, with the staple prey being
P. clarkii. This observation was consistent with that found in some Spanish reservoirs [
18], however, other reservoir populations exhibited fairly different niches. The prey diversity found was lesser than that of lotic sites, but this was to be expected, given that reservoirs are areas of high stress and disturbance mostly populated by non-native species.
Due to abundance being calculated based on prey number rather than volume, it is important to note that this alternative approach may change how these data are viewed, as fishes, who generally have a larger biomass than the freshwater crustaceans analyzed here, may become the most predated group and therefore be even more impacted than what was observed in this study.
In general, the typology of prey found within S. glanis stomachs seemed to be directly correlated with the size of the mouth, given that larger prey were found in larger individuals. In lotic habitats, individuals of Class 1 were already preying on other, smaller fish species like goby (Pomatoschistus sp.) and juveniles of both eel and lamprey. Class 2 individuals fed on benthic fishes, with 10% of the diet surprisingly consisting of pelagic species, such as A. boyerii and shad (Alosa sp.). It is also worth noting that, in these habitats, almost all (81.1%) of the diet comprised native species, and of these, roughly 20% were native fish species.
As for the lentic habitats, the mouth size/prey size relation was easier to observe with Class 1 individuals feeding strictly on crustaceans with a vast majority of freshwater shrimp, Class 2 fishes replacing the shrimp with crayfish and, finally, Class 3 individuals including large fish such as carp in their diet. The predation on pelagic prey demonstrated a plastic response of this species predatory behavior, enabling it to obtain new food resources [
13]. In fact, the different dietary habits observed in both environments attested to this broad adaptive capacity. Indeed, Copp et al. (2009) [
14] observed that the European catfish presented an enormous variation across the species range.
The prey cumulation curves did not stabilize, indicating that a larger sampling effort is needed to evaluate the full predatory niche this species is using in its current distribution. In fact, there is a new prey item registered for every two individuals in the lotic system, and one new prey item registered for every three individuals in the lentic system. However, when compared to the work of Guillerault et al. (2017) [
16], we saw that for the same number of individuals sampled (N = 16), they found six different prey species, while in the Tagus, 14 species were identified. The same work also described DNA metabarcoding as a more efficient method of diet composition analysis; perhaps, if the same had been applied in this case, the number of identified prey would be higher. In sum, the number of species affected by
S. glanis predation in the Portuguese Tagus River was larger than that found for France.
The larger prey diversity observed in the present study could be a result of native fish prey naivety, considering that this non-native fish was only established about a decade ago. As stated previously, in the Portuguese freshwater courses, piscivorous fishes were originally absent and, as such, the native fish species have not developed strategies to evade fish predation. This “inadaptation” toward this threat may explain why smaller S. glanis are able to prey on fish in the lotic system. Most non-native fishes found in reservoirs share the same native distribution with S. glanis, therefore, fishes like carp and goldfish may have some innate mechanism of defense that has not yet been silenced. Studying the diet of this recent apex predator constitutes an unique opportunity to demonstrate the early and considerable impact of predation on a unique fish community of the Lower Tagus River that could encompass more than 20 native fish, some endemic to this region.
4. Materials and Methods
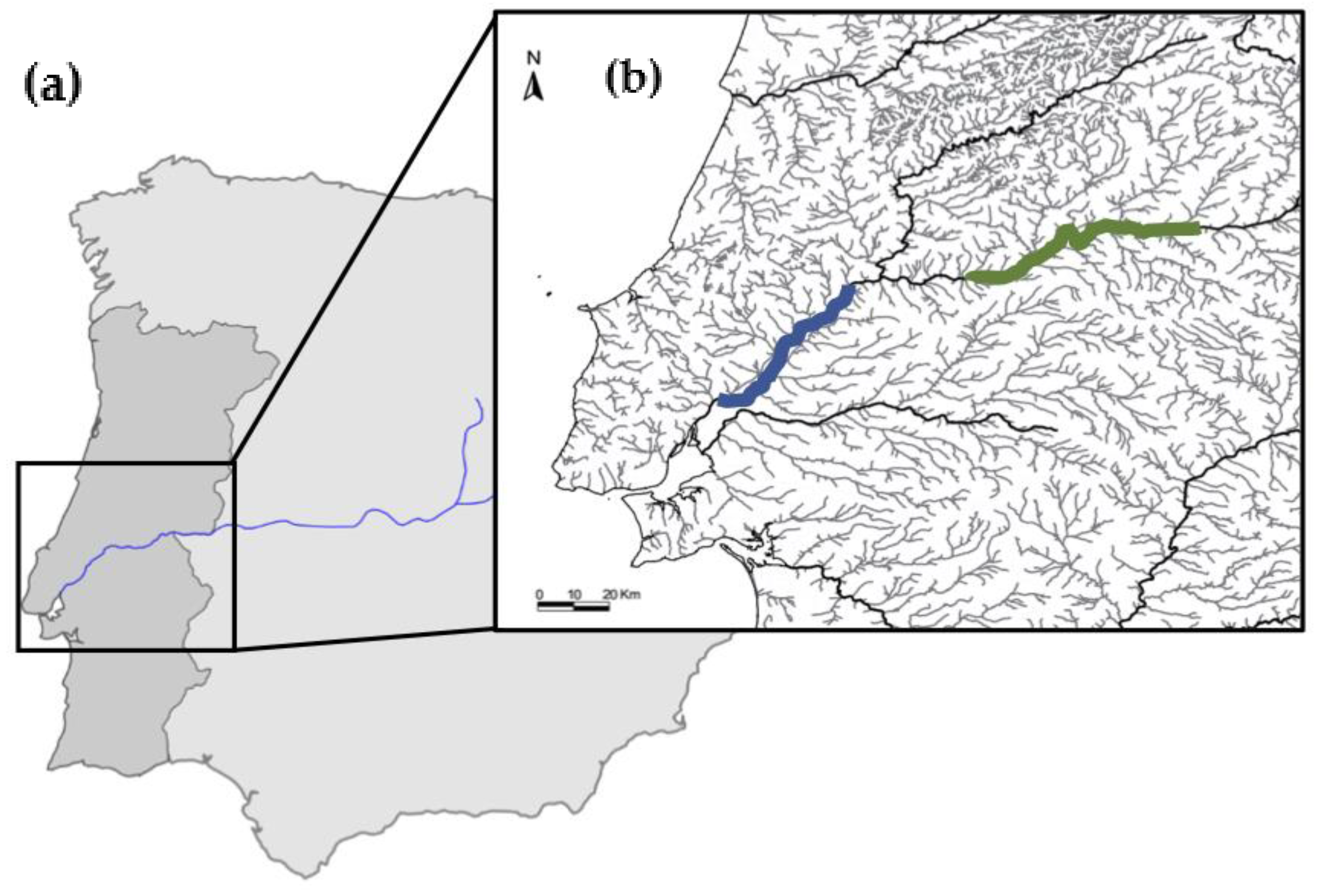
The fish used in this study were obtained opportunistically between April 2017 and November 2018 by fishermen located in two distinct areas, namely, the flowing section of the Tagus River (Santarém–Azambuja), referred to as lotic environment, and the lentic environment (Belver, Fratel, and Cedilho reservoirs) (
Figure 5). Fish were generally captured with stationary nets of various sizes and lengths. The Tagus is a river system with many obstacles through its course, like weirs and dams. Belver, Fratel, and Cedilho are hydroelectric dams with relatively small reservoirs that allow for the constant flow of water, which is maximized during periods of electrical production.
In the laboratory, fish were measured for total length (TL) to the nearest millimeter, when possible, and standard length (SL) to the nearest millimeter. They were weighed (total weight (TW) to the nearest gram) and a gut incision was made to extract the digestive tract, liver and gonads. The fishes were then weighed for eviscerated weight (EW) to the nearest gram. Stomach content analysis followed Knoppel (1970) [
26]. Contents were placed in a Petri dish, observed under a stereomicroscope (Leica MZ 125), and were posteriorly identified to the lowest possible taxa using identification keys [
27] as well as expert opinions.
A prey accumulation curve, which measured the number of different prey items as a function of sample effort, was obtained for each environment (lotic and lentic). This was done using the Vegan R package, via a randomizing methodology with 1000 permutations, adapted from [
28].
Fishes were separated into two groups according to place of capture, lentic environment and lotic environment, and then sub-divided into three size classes for each group using TL measurements, i.e., Class 1: 0–50 cm, Class 2: 50–100 cm, and Class 3: >100 cm. The relative numerical prey abundance (A
N, %) was calculated as described by Hyslop (1980) [
29]. The frequency of occurrence (F
O, %) was, given the total number of stomachs containing prey, the percentage of each of the prey items listed. Variation between systems and size classes in numeric prey abundance was evaluated using a Mann–Whitney U test, specifically pertaining to the four most preyed-upon items.
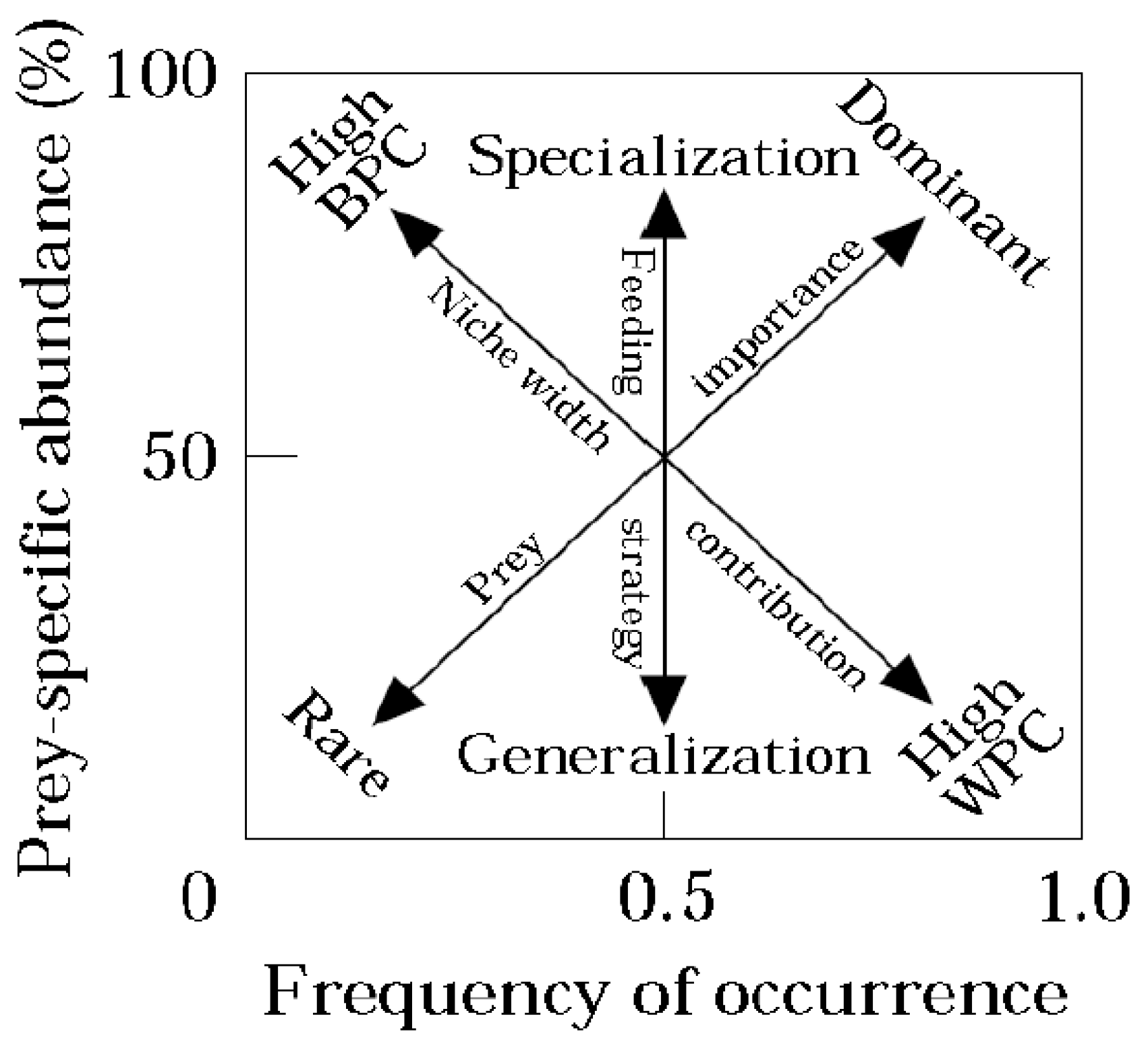
A modified Costello [
30] graphical method was used to illustrate the feeding behavior for each river system. This method plotted F
O (%) and prey specific abundance (P
i, %) to classify a species’ prey-selection behavior (1).
Pi (%) was calculated as a function of
Psi, the amount of prey items for a prey type (
i) found in the stomachs, and
Pst, the total amount of prey items of each type of prey found in each stomach that contained prey
i. The graphical representation allowed for the characterization of feeding behavior by analyzing the distribution of points along the diagonals and axes of the diagram (
Figure 6).
Feeding strategy was characterized by the vertical axis, with the bottom representing generalization and top representing specialization. The importance of each prey was characterized diagonally, from rare prey in the lower left and dominant prey in the upper right. Individuals with a narrow predatory niche were associated with prey items found in the upper left region and individuals with a broad niche were associated with prey items in the lower right region (
Figure 6) [
30]. This analysis was produced using Excel 2013.
A non-metric multidimensional scaling analysis (NMDS) was performed using the R package Vegan: Community Ecology Package [
31] to visualize the prey spectrum variation between and within each group of fish (lentic and lotic and size class within each environment). This approach was based in the Bray–Curtis similarity index [
32]. The prime components were understood as tendencies for prey niche dimensions. A larger number of iterations was used in order to reduce stress values to <0.2, as the standard approach resulted in lack of component convergence; this was possibly due to low sample size. A follow up analysis of similarity (ANOSIM) was attempted, but trust in the results was not satisfactory and therefore was not included, following Gkenas et al. (2019) [
33].






 ) and lentic section (
) and lentic section (  ).
).



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}