
Experimental Study of the Environmental Effects of Summertime Cocultures of Seaweed Gracilaria lemaneiformis (Rhodophyta) and Japanese Scallop Patinopecten yessoensis in Sanggou Bay, China


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experiment Design and Management
2.2. Sample Analysis
2.3. Data Analysis
3. Results
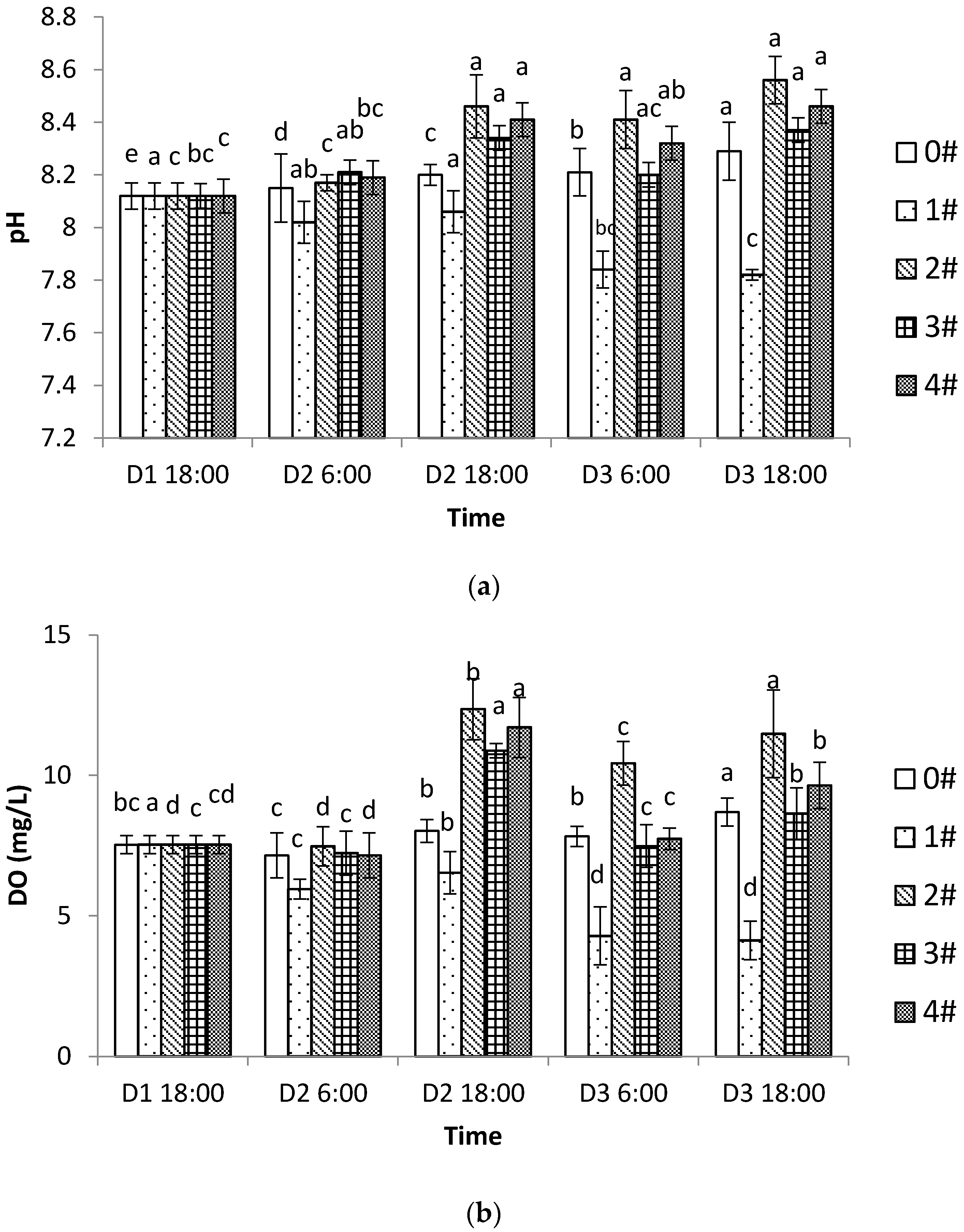
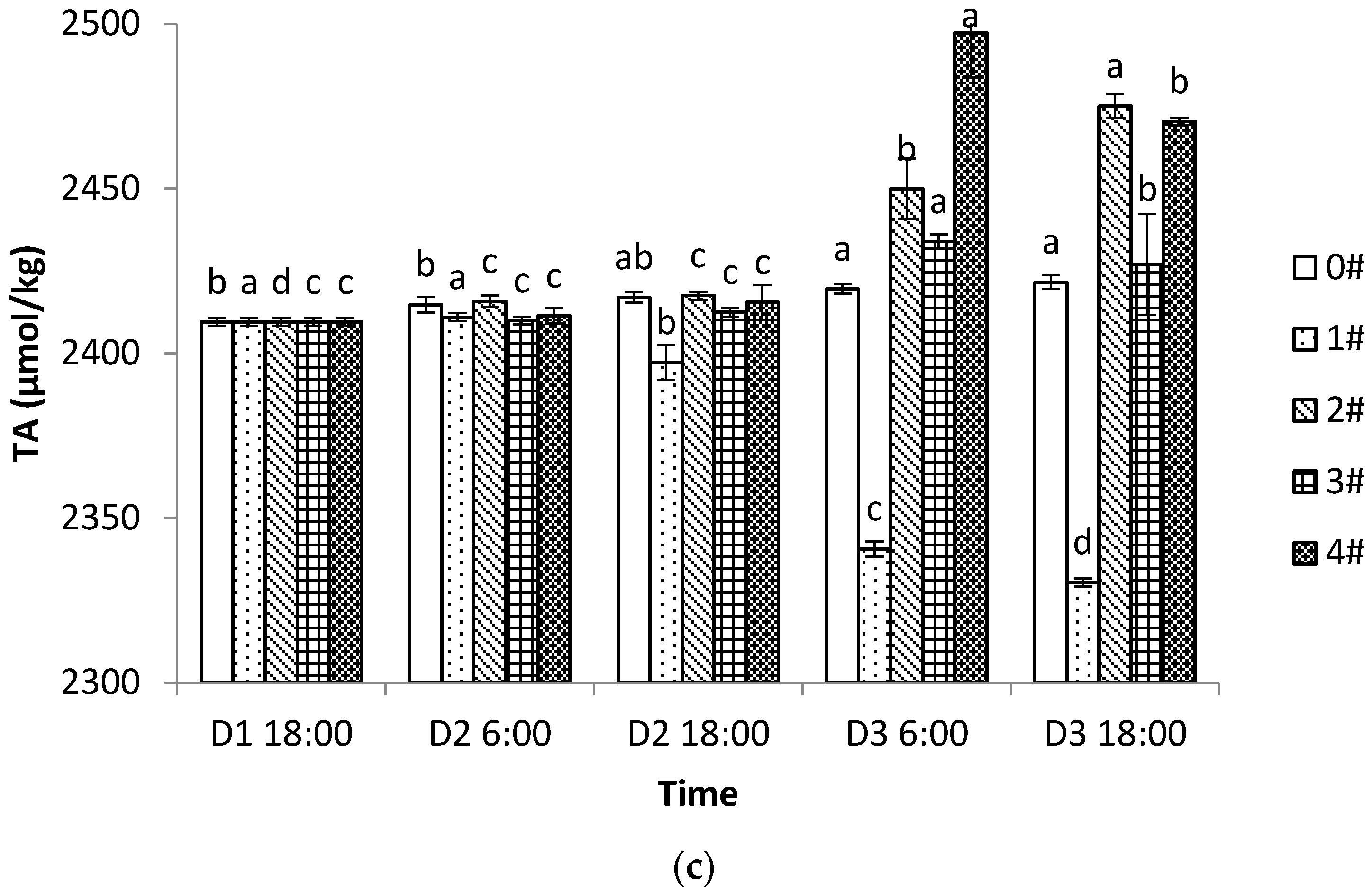
3.1. Changes of pH, DO, and TA in Different Culture Modes
3.2. Variation of Chl-a and Components in Different Culture Modes
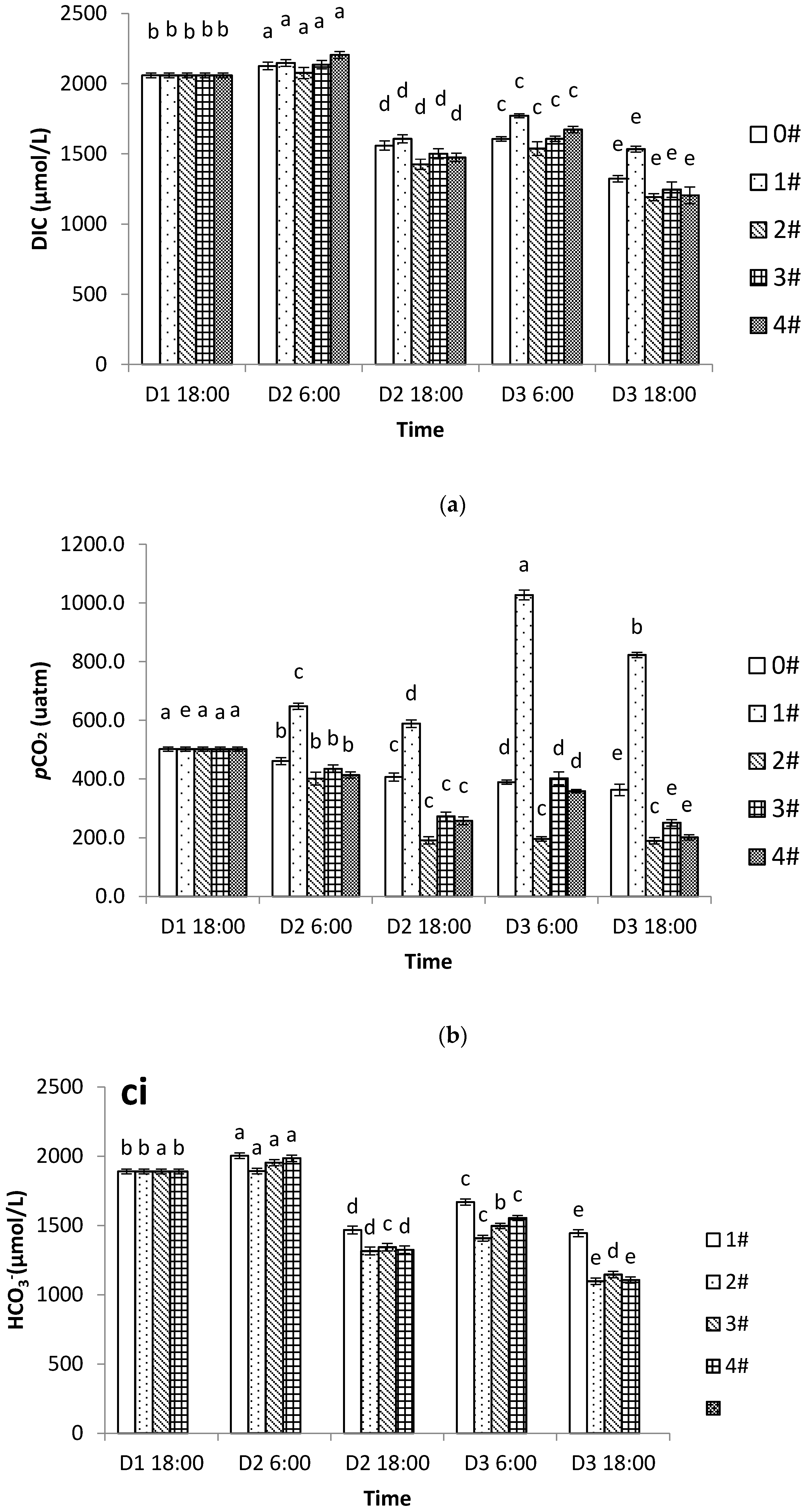
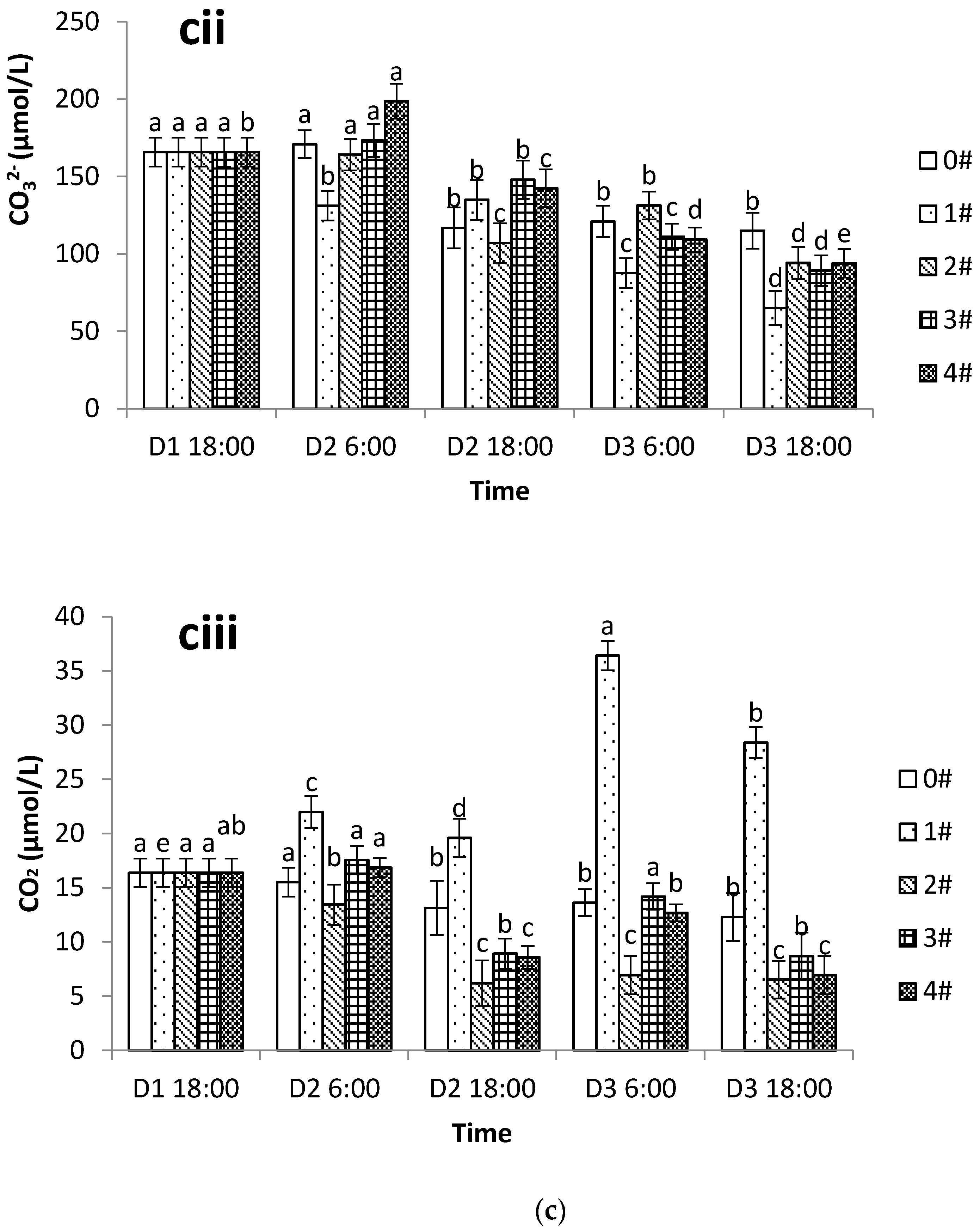
3.3. Changes in DIC, pCO2, and the Carbonate System in Different Aquaculture Modes
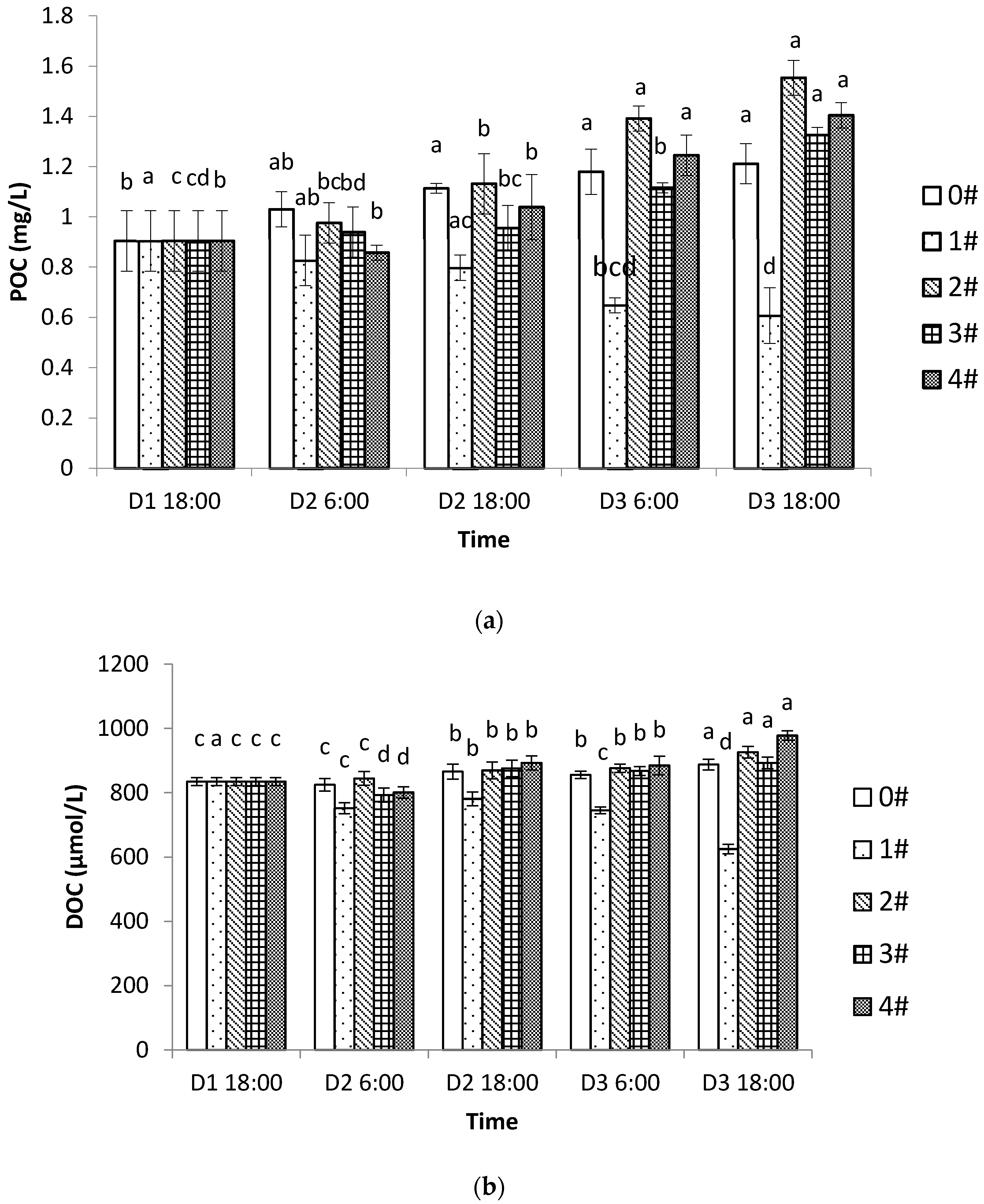
3.4. POC and DOC in Different Culture Modes
3.5. Variation of Nutrients in Different Culture Modes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, Y. Comparison of Culture Effect, Discharge of Nitrogen and Phosphorus and Environmental Influence for Three Kinds of Cages. Master’s Thesis, Huazhong Agriculture University, Wuhan, China, 2012. (In Chinese with English abstract). [Google Scholar]
- Fang, J.; Fang, J.; Chen, Q.; Mao, Y.; Jiang, Z.; Du, M.; Gao, Y.; Lin, F. Assessing the effects of oyster/kelp weight ratio on water column properties: An experimental IMTA study at Sanggou Bay, China. J. Oceanol. Limnol. 2019, 38, 3–4. [Google Scholar] [CrossRef]
- Reid, G.K.; Lefebvre, S.; Filgueira, R.; Robinson, S.M.C.; Broch, O.J.; Dumas, A.; Chopin, T.B.R. Performance measures and models for open-water integrated multi-trophic aqua-culture. Rev. Aquac. 2018, 12, 47–75. [Google Scholar] [CrossRef]
- Troell, M.; Halling, C.; Neori, A.; Chopin, T.; Buschmann, A.; Kautsky, N.; Yarish, C. Integrated mariculture: Asking the right questions. Aquaculture 2003, 226, 69–90. [Google Scholar] [CrossRef]
- Tang, Q.S.; Fang, J.G.; Zhang, J.H.; Jiang, Z.J.; Liu, H.M. Impacts of multiple stressors on coastal ocean ecosystems and integrated multi-trophic aquaculture. Prog. Fish. Sci. 2013, 34, 1–11, (In Chinese with English abstract). [Google Scholar]
- Tang, Q.; Zhang, J.; Fang, J. Shellfish and seaweed mariculture increase atmospheric CO2 absorption by coastal ecosystems. Mar. Ecol. Prog. Ser. 2011, 424, 97–104. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.; Fang, J. Effects of mussel-kelp ratios in integrated mariculture on the carbon dioxide system in Sanggou Bay. J. Sea Res. 2021, 167, 101983. [Google Scholar] [CrossRef]
- Jiang, Z.; Wang, G.; Fang, J.; Mao, Y. Growth and food sources of Pacific oyster Crassostrea gigas integrated culture with Sea bass Lateolabrax japonicus in Ailian Bay, China. Aquac. Int. 2012, 21, 45–52. [Google Scholar] [CrossRef]
- Chopin, T.; Robinson, S.; Troell, M.; Neori, A.; Buschmann, A.; Fang, J. Multitrophic Integration for Sustainable Marine Aquaculture. Encycl. Ecol. 2008, 2463–2475. [Google Scholar] [CrossRef]
- Ning, Z.; Liu, S.; Zhang, G.; Ning, X.; Li, R.; Jiang, Z.; Fang, J.; Zhang, J. Impacts of an integrated multi-trophic aquaculture system on benthic nutrient fluxes: A case study in Sanggou Bay, China. Aquac. Environ. Interact. 2016, 8, 221–232. [Google Scholar] [CrossRef] [Green Version]
- Chauvaud, L.; Thompson, J.K.; Cloern, J.; Thouzeau, G. Clams as CO2 generators: The Potamocorbula amurensis example in San Francisco Bay. Limnol. Oceanogr. 2003, 48, 2086–2092. [Google Scholar] [CrossRef] [Green Version]
- Suikkanen, S.; Silvia, P.; Engström-Öst, J.; Lehtiniemi, M.; Lehtinen, S.; Brutemark, A. Climate Change and Eutrophication Induced Shifts in Northern Summer Plankton Communities. PLoS ONE 2013, 8, e66475. [Google Scholar] [CrossRef] [Green Version]
- Joseph, S.S.; Winston, R.J.; Tirpak, R.A.; Wituszynski, D.M.; Boening, K.M.; Martin, J.F. The seasonality of nutrients and sediment in residential stormwater runoff: Impli-cations for nutrient-sensitive waters. J. Environ. Manag. 2020, 276, 111248. [Google Scholar]
- Su, J.; Cai, W.-J.; Brodeur, J.; Chen, B.; Hussain, N.; Yao, Y.; Ni, C.; Testa, J.M.; Li, M.; Xie, X.; et al. Chesapeake Bay acidification buffered by spatially decoupled carbonate mineral cycling. Nat. Geosci. 2020, 13, 441–447. [Google Scholar] [CrossRef]
- Nikinmaa, M. Climate change and ocean acidification—Interactions with aquatic toxicology. Aquat. Toxicol. 2013, 126, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Kanerva, M.; Kiljunen, M.; Torniainen, J.; Nikinmaa, M.; Dutz, J.; Vuori, K.A. Environmentally driven changes in Baltic salmon oxidative status during marine migration. Sci. Total Environ. 2020, 742, 140259. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Yang, H.; Hu, H.; Liu, Y.; Mao, Y.; Zhou, H.; Xu, X.; Zhang, F. Bioremediation potential of the macroalga Gracilaria lemaneiformis (Rhodophyta) integrated into fed fish culture in coastal waters of north China. Aquaculture 2006, 252, 264–276. [Google Scholar] [CrossRef]
- Fei, X.G.; Lu, S.; Bao, Y.; Wilkes, R.; Yarish, C. Seaweed cultivation in China. World Aquac. 1998, 29, 22–24. [Google Scholar]
- Fei, X.G.; Bao, Y.; Lu, S. Seaweed cultivation—Traditional way and its reformation. Aquac. Oceanol. Limnol. Sin. 1999, 17, 193–199. [Google Scholar]
- Zhang, M.; Fang, J.; Zhang, J.; Bin, L.; Ren, S.; Mao, Y.; Gao, Y. Effect of Marine Acidification on Calcification and Respiration of Chlamys farreri. J. Shellfish Res. 2011, 30, 267–271. [Google Scholar]
- Sugden, A.M. Threats of coastal hypoxia. Science 2017, 356, 38. [Google Scholar] [CrossRef] [PubMed]
- Levin, L.; Breitburg, D.L. Linking coasts and seas to address ocean deoxygenation. Nat. Clim. Chang. 2015, 5, 401–403. [Google Scholar] [CrossRef]
- Vaquer-Sunyer, R.; Duarte, C.M. Thresholds of hypoxia for marine biodiversity. Proc. Natl. Acad. Sci. USA 2008, 105, 15452–15457. [Google Scholar] [CrossRef] [Green Version]
- Fang, J.H.; Zhang, J.H.; Wu, W.G.; Mao, Y.Z.; Gao, Y.P.; Jiang, Z.J.; Fang, J.G. Carbon and nitrogen budget and environmental optimization in an integrated cage culture model of Japanese flounder with Perinereis aibuhitensis. J. Fish. Sci. 2014, 21, 390–397, (In Chinese with English abstract). [Google Scholar]
- Mao, Y.; Yang, H.; Zhou, Y.; Ye, N.; Fang, J. Potential of the seaweed Gracilaria lemaneiformis for integrated multi-trophic aquaculture with scallop Chlamys farreri in North China. J. Appl. Phycol. 2009, 21, 649–656. [Google Scholar] [CrossRef]
- Phillips, J.; Hurd, C. Nitrogen ecophysiology of intertidal seaweeds from New Zealand: N uptake, storage and utilisation in relation to shore position and season. Mar. Ecol. Prog. Ser. 2003, 264, 31–48. [Google Scholar] [CrossRef]
- Pedersen, A.; Kraemer, G.; Yarish, C. The effects of temperature and nutrient concentrations on nitrate and phosphate uptake in different species of Porphyra from Long Island Sound (USA). J. Exp. Mar. Biol. Ecol. 2004, 312, 235–252. [Google Scholar] [CrossRef]
- Lartigue, J.; Sherman, T.D. Response of Enteromorpha sp. (Chlorophyceae) to a nitrate pulse: Nitrate uptake, inorganic nitrogen storage and nitrate reductase activity. Mar. Ecol. Prog. Ser. 2005, 292, 147–157. [Google Scholar] [CrossRef]
- Zou, D.H.; Xia, J.R.; Yang, Y.F. Photosynthetic use of exogenous inorganic carbon in the agarophyte Gracilaria lemaneiformis (Rhodophyta). Aquaculture 2004, 237, 421–431. [Google Scholar] [CrossRef]
- Fisher, T.; Peele, E.; Ammerman, J.; Harding, L. Nutrient limitation of phytoplankton in Chesapeake Bay. Mar. Ecol. Prog. Ser. 1992, 82, 51–63. [Google Scholar] [CrossRef]
- Justić, D.; Rabalais, N.N.; Turner, R.E.; Dortch, Q. Changes in nutrient structure of river-dominated coastal waters: Stoichiometric nutrient balance and its consequences. Estuar. Coast. Shelf Sci. 1995, 40, 339–356. [Google Scholar] [CrossRef]
- Redfield, A.C.; Ketchum, B.H.; Richard, F.A. The influence of organism on the composition of seawater. Sea 1963, 2, 26–77. [Google Scholar]
- Prins, T.C.; Escaravage, V.; Smaal, A.C.; Peeters, J.C.H. Nutrient cycling and phytoplankton dynamics in relation to mussel grazing in a mesocosm experiment. Ophelia 1995, 41, 289–315. [Google Scholar] [CrossRef]
- Souchu, P.; Vaquer, A.; Collos, Y.; Landrein, S.; Deslous-Paoli, J.; Bibent, B. Influence of shellfish farming activities on the biogeochemical composition of the water column in Thau lagoon. Mar. Ecol. Prog. Ser. 2001, 218, 141–152. [Google Scholar] [CrossRef] [Green Version]
- Dupuy, C.; Vaquer, A.; Lam-Höai, T.; Rougier, C.; Mazouni, N.; Lautier, J.; Collos, Y.; Le Gall, S. Feeding rate of the oyster Crassostrea gigas in a natural planktonic community of the Mediterranean Thau Lagoon. Mar. Ecol. Prog. Ser. 2000, 205, 171–184. [Google Scholar] [CrossRef]
- Lu, J.C.; Huang, L.F.; Xiao, T.; Jiang, Z.; Zhang, W. The effects of Zhikong scallop (Chlamys farreri) on the microbial food web in a phospho-rus-deficient mariculture system in Sanggou Bay, China. Aquaculture 2015, 448, 341–349. [Google Scholar] [CrossRef]
- Han, T.; Jiang, Z.; Fang, J.; Zhang, J.; Mao, Y.; Zou, J.; Huang, Y.; Wang, D. Carbon dioxide fixation by the seaweed Gracilaria lemaneiformis in integrated multi-trophic aquaculture with the scallop Chlamys farreri in Sanggou Bay, China. Aquac. Int. 2013, 21, 1035–1043. [Google Scholar] [CrossRef]
- Schlitzer, R. Export and sequestration of particulate organic carbon in the north pacific from inverse modeling. In Proceedings of the Workshop of the JGOFS North Pacific Process Study Synthesis Group, Sapporo, Japan, 2 October 2002. [Google Scholar]
- Han, T.; Shi, R.; Qi, Z.; Huang, H.; Wu, F.; Gong, X. Biogenic acidification of Portuguese oyster Magallana angulata mariculture can be mediated through introducing brown seaweed Sargassum hemiphyllum. Aquaculture 2020, 520, 734972. [Google Scholar] [CrossRef]
- Ren, L.H.; Zhang, J.H.; Fang, J.G.; Tang, Q.H.; Liu, Y.; Du, M.R. The diurnal rhythm of respiration, excretion and calcification in oyster Crassostre gigas. Prog. Fish. Sci. 2013, 1, 75–81, (In Chinese with English abstract). [Google Scholar]
- Zhang, J.H.; Fang, J.G.; Tang, Q.S.; Ren, L.H. Carbon sequestration rate of the scallop Chlamys farreri cultivated in different areas of Sanggou Bay. Prog. Fish. Sci. 2013, 34, 12–16, (In Chinese with English abstract). [Google Scholar]
- Zhang, Y.; Zhang, J.; Liang, Y.; Li, H.; Li, G.; Chen, X.; Zhao, P.; Jiang, Z.; Zou, D.; Liu, X.; et al. Carbon sequestration processes and mechanisms in coastal mariculture environments in China. Sci. China Earth Sci. 2017, 60, 2097–2107. [Google Scholar] [CrossRef]
- Li, H.; Zhang, Y.; Liang, Y.; Chen, J.; Zhu, Y.; Zhao, Y.; Jiao, N. Impacts of maricultural activities on characteristics of dissolved organic carbon and nutrients in a typical raft-culture area of the Yellow Sea, North China. Mar. Pollut. Bull. 2018, 137, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Jiao, N.; Cai, R.; Zheng, Q.; Tang, K.; Liu, J.; Jiao, F.; Wallace, D.; Chen, F.; Li, C.; Amann, R.; et al. Unveiling the enigma of refractory carbon in the ocean. Natl. Sci. Rev. 2018, 5, 459–463. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mode | Japanese Scallop (g) | Seaweed (g) | Ratios |
|---|---|---|---|
| Control (0#) | 0 | 0 | / |
| Group 1 (1#) | 400.3 ± 2.4 | 0 | / |
| Group 2 (2#) | 0 | 399.7 ± 3.22 | / |
| Group 3 (3#) | 401.3 ± 2.59 | 400.4 ± 7.52 | 1:1 |
| Group 4 (4#) | 398.87 ± 8.98 | 639.08 ± 6.15 | 1:1.6 |
| Source | Sum of Squares | df | Mean Squares | F | p | |
|---|---|---|---|---|---|---|
| DO | Mode | 239.505 | 4 | 59.876 | 1373.056 | 0.000 |
| Time | 120.056 | 4 | 30.014 | 688.265 | 0.000 | |
| Mode × Time | 228.045 | 16 | 14.253 | 326.839 | 0.000 | |
| pH | Mode | 1.266 | 4 | 0.316 | 222.854 | 0.000 |
| Time | 0.409 | 4 | 0.102 | 72.051 | 0.000 | |
| Mode × Time | 0.672 | 16 | 0.042 | 29.575 | 0.000 | |
| TA | Mode | 36,383.561 | 4 | 9095.890 | 7517.265 | 0.000 |
| Time | 4551.065 | 4 | 1137.766 | 940.303 | 0.000 | |
| Mode × Time | 45,434.731 | 16 | 2839.671 | 2346.835 | 0.000 | |
| Source | Sum of Squares | df | Mean Squares | F | p | |
|---|---|---|---|---|---|---|
| Mode | 21.224 | 4 | 5.306 | 124.961 | 0.000 | |
| Chl-a | Time | 249.662 | 4 | 62.415 | 1469.937 | 0.000 |
| Mode × Time | 30.099 | 16 | 1.881 | 44.303 | 0.000 | |
| Source | Sum of Squares | df | Mean Squares | F | p | |
|---|---|---|---|---|---|---|
| DIC | Mode | 215,895.095 | 4 | 53,973.774 | 10,709.048 | 0.000 |
| Time | 7,725,702.666 | 4 | 1,931,425.667 | 383,218.162 | 0.000 | |
| Mode × Time | 198,111.298 | 16 | 12,381.956 | 2456.730 | 0.000 | |
| pCO2 | Mode | 1,664,176.577 | 4 | 416,044.144 | 80,420.904 | 0.000 |
| Time | 307,442.537 | 4 | 76,860.634 | 14,857.081 | 0.000 | |
| Mode × Time | 823,088.365 | 16 | 51,443.023 | 9943.883 | 0.000 | |
| HCO3− | Mode | 250,760.705 | 4 | 62,690.176 | 2072.403 | 0.000 |
| Time | 6,416,415.953 | 4 | 1,604,103.988 | 53,028.231 | 0.000 | |
| Mode × Time | 190,286.215 | 16 | 11,892.888 | 393.153 | 0.000 | |
| CO32− | Mode | 5749.88 | 4 | 1437.475 | 47.520 | 0.000 |
| Time | 66,728.177 | 4 | 16,682.044 | 551.473 | 0.000 | |
| Mode × Time | 11,856.723 | 16 | 741.045 | 24.497 | 0.000 | |
| CO2 | Mode | 1929.804 | 4 | 482.451 | 15.949 | 0.000 |
| Time | 433.633 | 4 | 108.408 | 3.584 | 0.012 | |
| Mode × Time | 1073.704 | 16 | 67.107 | 2.218 | 0.016 | |
| Source | Sum of Squares | df | Mean Squares | F | p | |
|---|---|---|---|---|---|---|
| DOC | Mode | 48,740.061 | 4 | 12,185.015 | 50.286 | 0.000 |
| Time | 125,626.612 | 4 | 31,406.653 | 129.611 | 0.000 | |
| Mode × Time | 76,374.501 | 16 | 4773.406 | 19.699 | 0.000 | |
| POC | Mode | 1.599 | 4 | 0.400 | 53.790 | 0.000 |
| Time | 1.057 | 4 | 0.264 | 35.548 | 0.000 | |
| Mode × Time | 1.219 | 16 | 0.076 | 10.248 | 0.000 | |
| Mode | Time | SiO4 | DIN | PO4 | Si:P | N:P | Si:N |
|---|---|---|---|---|---|---|---|
| All gourps | Initial | 14.88 ± 2.35 | 16.04 ± 1.32 | 1.57 ± 0.27 | 9.48 | 10.22 | 0.93 |
| 0# | Final | 14.32 ± 1.22 | 3.85 ± 0.87 | 0.48 ± 0.12 | 29.83 | 8.02 | 3.72 |
| 1# | Final | 6.56 ± 1.01 | 18.86 ± 2.21 | 1.32 ± 0.24 | 4.97 | 14.29 | 0.35 |
| 2# | Final | 14.77 ± 1.25 | 2.37 ± 1.06 | 0.30 ± 0.11 | 36.93 | 7.9 | 6.23 |
| 3# | Final | 8.21 ± 0.97 | 9.64 ± 1.13 | 0.49 ± 0.25 | 16.76 | 19.67 | 0.60 |
| 4# | Final | 8.94 ± 1.33 | 8.9 ± 1.21 | 0.34 ± 0.10 | 26.29 | 26.17 | 0.69 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Wang, X.; Wu, W.; Yang, J.; Wu, N.; Zhang, J. Experimental Study of the Environmental Effects of Summertime Cocultures of Seaweed Gracilaria lemaneiformis (Rhodophyta) and Japanese Scallop Patinopecten yessoensis in Sanggou Bay, China. Fishes 2021, 6, 53. https://doi.org/10.3390/fishes6040053
Liu Y, Wang X, Wu W, Yang J, Wu N, Zhang J. Experimental Study of the Environmental Effects of Summertime Cocultures of Seaweed Gracilaria lemaneiformis (Rhodophyta) and Japanese Scallop Patinopecten yessoensis in Sanggou Bay, China. Fishes. 2021; 6(4):53. https://doi.org/10.3390/fishes6040053
Chicago/Turabian StyleLiu, Yi, Xinmeng Wang, Wenguang Wu, Jun Yang, Ningning Wu, and Jihong Zhang. 2021. "Experimental Study of the Environmental Effects of Summertime Cocultures of Seaweed Gracilaria lemaneiformis (Rhodophyta) and Japanese Scallop Patinopecten yessoensis in Sanggou Bay, China" Fishes 6, no. 4: 53. https://doi.org/10.3390/fishes6040053
APA StyleLiu, Y., Wang, X., Wu, W., Yang, J., Wu, N., & Zhang, J. (2021). Experimental Study of the Environmental Effects of Summertime Cocultures of Seaweed Gracilaria lemaneiformis (Rhodophyta) and Japanese Scallop Patinopecten yessoensis in Sanggou Bay, China. Fishes, 6(4), 53. https://doi.org/10.3390/fishes6040053