1. Introduction
The Alanine, Serine, Cysteine Transporter 2 (ASCT2) is a Na
+-dependent, broad scope neutral amino acid exchanger encoded by the SLC1A5 gene, which is the fifth member of the SLC1 family [
1,
2]. ASCT2 is highly expressed in many kinds of cancer cells and plays a key role in their growth [
3,
4,
5]. It catalyzes the obligatory exchange of substrate amino acids, including alanine, serine, cysteine, threonine, glutamine and so on [
6]. In addition to its role as a transporter, it also acts as a receptor for several retroviruses, such as RD-114 virus and simian retrovirus 4 [
7,
8]. Therefore, ASCT2 has increasingly gained attention over the past few years.
With the development of high-throughput sequencing technology, the sequences of SLC1A5 have been predicted in many species, which contribute to the research in molecular mechanism of ASCT2 as well as its related signaling pathways. Nevertheless, most previous studies of ASCT2 were focused on humans as well as a few mammals. In contrast to numerous research hotspots in human physiology and pathology, the research in aquaculture regarding ASCT2 molecular characterization and nutrition regulation, which can contribute to the research on the improvement of feed formula, has been sparse.
Although ASCT2 was named by the acronym of Alanine, Serine, Cysteine Transporter 2, the preferred substrate of ASCT2 is glutamine [
9,
10]. Glutamine is the major oxidative fuel of the enterocyte, an essential precursor for nucleic acid biosynthesis in all cells and a key link between carbon metabolism of carbohydrates and proteins [
11,
12,
13]. It is a vehicle for nitrogen transfer between tissues, which plays an important role in cell growth [
14]. Hu et al. reported that 7.5 g/kg glutamine and 2.5 g/kg glutamine dipeptide could remarkably increase the expression of ASCT2 in grass carp (
Ctenopharyngodon idelus) [
15]. Therefore, it is necessary to study the transport mechanism of glutamine in ASCT2. Moreover, Kanai et al. reported that glutamate was also one of the transport substrates of ASCT2 [
16]. In the intestine, glutamate is taken up from the lumen or is produced in the epithelium from glutamine through glutaminase activity [
17]. It is an important nutrient in the maintenance of intestinal mucosal activity. Thus, the regulatory effect of glutamate on ASCT2 is also worthy of our attention.
Protein is the core component in fish feed. Therefore, researchers improve the feed formula by adjusting the protein content, protein source and additives. In our previous research, we reported the effects of different dietary protein levels on the expression levels of peptide transporter 1 (PepT1), L-Type amino acid transporter 2 (LAT2), and glutamate dehydrogenase (GDH) in crucian carp [
18,
19,
20]. These proteins showed different preferences for dietary protein levels. Therefore, studying the expression pattern of ASCT2 in fish fed under the different protein levels will help to deepen the understanding of ASCT2 and improve the dietary formula.
Crucian carp is one of the most important freshwater species for Chinese aquaculture. Triploid crucian carp (3
n = 150) has the characteristics of resilience, sterile traits and fast growth rate [
21]. However, the molecular mechanism of nutrition regulation on triploid crucian carp is still spare. Hence, we cloned ASCT2 cDNA from triploid crucian carp and studied the expression pattern in different tissues and diurnal variation. Furthermore, we analyzed the regulation of ASCT2 by dietary protein levels and protein source, as well as feed additives (glutamate and glutamine) in triploid crucian carp. The results of this study will contribute to enrich the knowledge of fish ASCT2 and have a potential application in improving fish feed formulation in aquaculture.
2. Materials and methods
2.1. Fish
All the triploid crucian carp in this experiment were provided by the Hunan Institute of Aquatic Science. Before the experiment, all of the fish were kept in a 300-L circular fiberglass container in an indoor recirculating freshwater system at 25 °C for 2 weeks of adaptation.
2.2. RNA Isolation and cDNA Synthesis
Total RNAs were isolated from the specimens of fish using Trizol reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions. The integrity of RNA samples was tested by agarose gel electrophoresis, and the quality of RNA samples was determined by calculating through the A260/A280 and A260/A230 ratios spectrophotometer (BioPhotometer Eppendorf, Hamburg, Germany). To eliminate endogenous DNA contamination, 1 g RNA was digested by DNase I prior to the cDNA synthesis. RNA was reversely transcribed to cDNA by a PrimeScript RT reagent kit with gDNA eraser (TaKaRa, Dalian, China). The products were separated by a 1% agarose gel and then cloned and tested.
2.3. Cloning of cDNA of ASCT2
The primers of SLC1A5 were designed based on conserved sequences of ASCT2/SLC1A5 in other teleosts (
Table 1). Polymerase chain reactions (PCRs) were carried out using a volume of 20 μL with 1 μL of cDNA (approximately 20 ng), 10 μL of 2 × Taq PCR Master Mix (Tiangen, Beijing, China), 8 μL of ddH
2O, and 0.5 μL of each primer (100 μM). The temperature profile during amplification was as follows: initial denaturation at 94 °C for 5 min, followed by 35 cycles at 94 °C for 30 s, 55 °C for 1 min, and 72 °C for 2 min. A final extension step was performed at 72 °C for 10 min. The PCR products were separated by polyacrylamide gel electrophoresis (PAGE). The DNA fragments were purified using a gel extraction kit (UNIQ-10 Spin Column DNA Gel Extraction Kit for PAGE, Sangon, Shanghai, China) and ligated into the pMD19-T vector. The plasmids were transformed into
E.coli DH5α and purified. The inserted DNA fragments in the pMD19-T vector were sequenced using an automated DNA sequencer (ABI PRISM 3730, Applied Biosystems, Carlsbad, CA, USA). All the PCRs in this study were repeated, and the results were consistent.
2.4. Phylogenetic Relationships
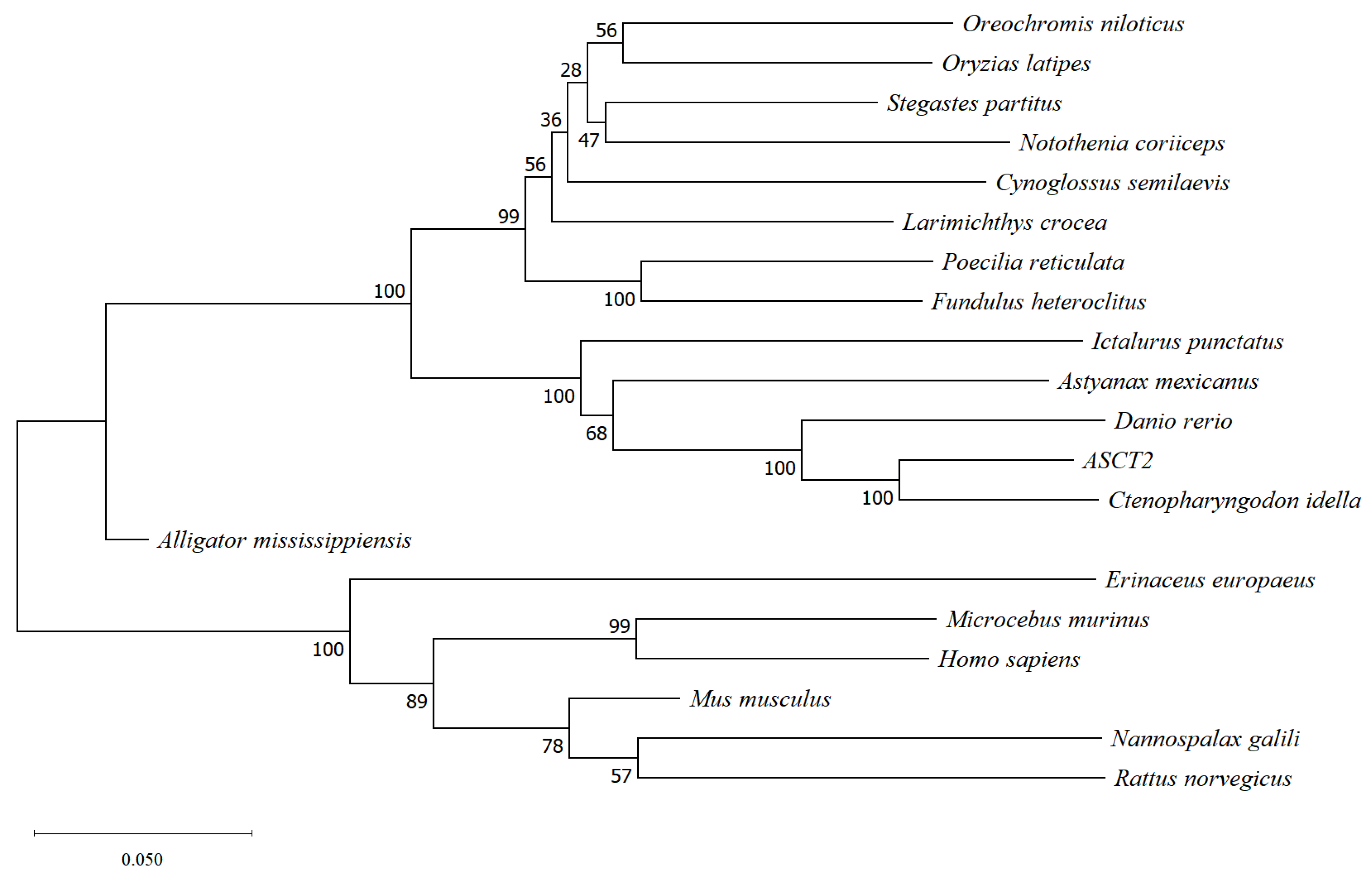
Phylogenetic analysis was conducted using the sequences of SLC1A5 homologs from triploid crucian carp and other vertebrates according to the methods of Mao et al. [
22]. The nucleotide sequences of these species were derived from the National Center for Biotechnology Information database. Evolutionary analyses were conducted in MEGA-X [
23]. Evolutionary history was inferred using the Neighbor-Joining method [
24]. The tree was drawn to scale, with the same units for branch lengths and the evolutionary distances used to infer the phylogenetic tree. The evolutionary distances were computed using the maximum likelihood method and measured as the number of base substitutions per site. All positions containing gaps and missing data were eliminated from analysis.
2.5. Quantitative Real-Time PCR (qRT-PCR)
The ASCT2 expression levels were determined by qRT-PCR in a Prism 7500 Sequence Detection System (Applied Biosystems, Foster City, CA, USA). The primers were designed using Primer Premier 5.0 software and the single band of amplification is determined by agarose gel electrophoresis (
Table 1). Beta-actin was used as an endogenous control. Five microliters of first-strand cDNAs (in a dilution of 1:20) as templates was added to 20 μL PCR solution containing 20 nmol/L primers and 10 μL SYBR Green PCR Master Mix (Applied Biosystems). For each sample, three repeats were performed with the following condition: 50 °C for 5 min and 95 ℃ for 10 min followed by 40 cycles at 95 °C for 15 s and 60 °C for 45 s. The efficiency of the assay was determined by the amplification of a dilution series of cDNA, and the analysis of relative mRNA expression was performed using the 2
−ΔΔCt method [
25].
2.6. ASCT2 Expression in Various Tissues and Diurnal Variation
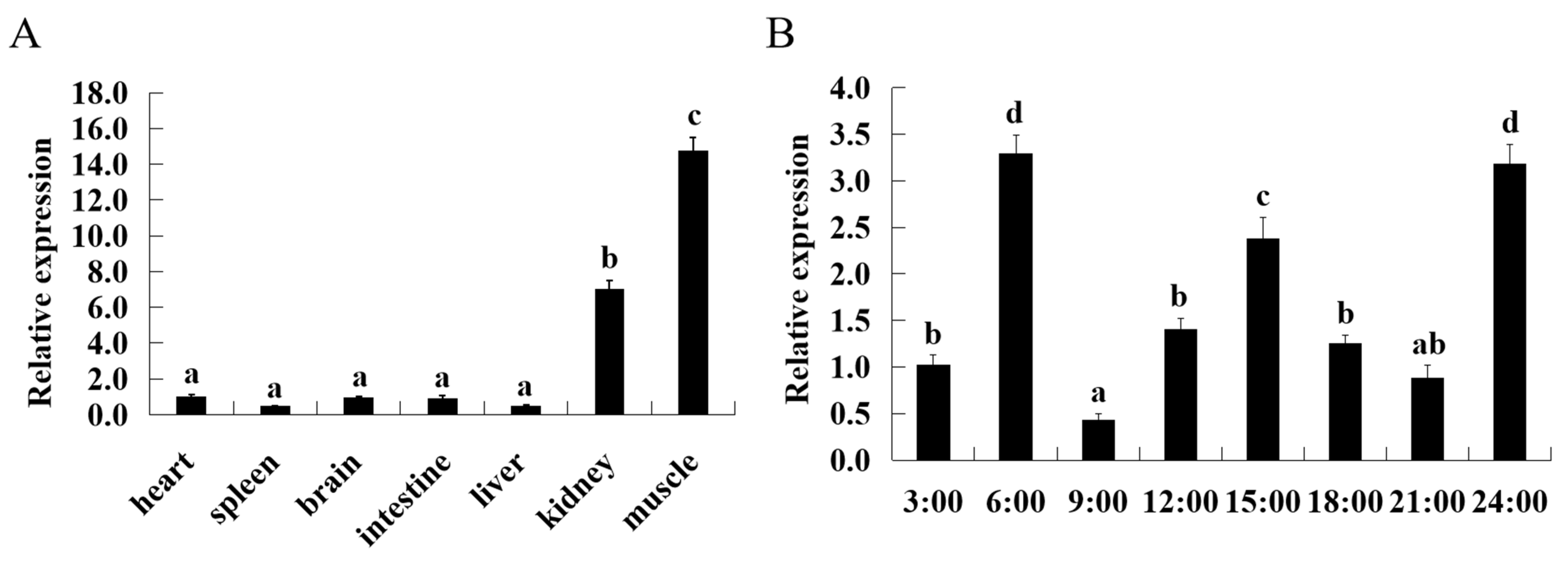
Before the experience, the fish (body weight 11.72 ± 0.11 g) were acclimatized for 2 weeks to an indoor recirculating aquaculture system comprising 4 glass fiber tanks (1500 L). They were fed twice daily at 8:30 a.m. and 2:30 p.m., respectively. In order to explore the expression differences of ASCT2 among different tissues, different tissues were collected from triploid crucian carp, including heart, spleen, brain, intestine, liver, kidney, and muscle. Later, the samples of muscle in triploid crucian carp were collected at eight different times (00:00, 03:00, 06:00, 09:00, 12:00, 15:00, 18:00, 21:00, and 24:00). Three replicates for each experimental group were designed. Total RNAs were isolated from these samples; after synthesis of cDNAs, ASCT2 expressions were assayed by qRT-PCR.
2.7. The Effect of Glutamate and Glutamine on ASCT2
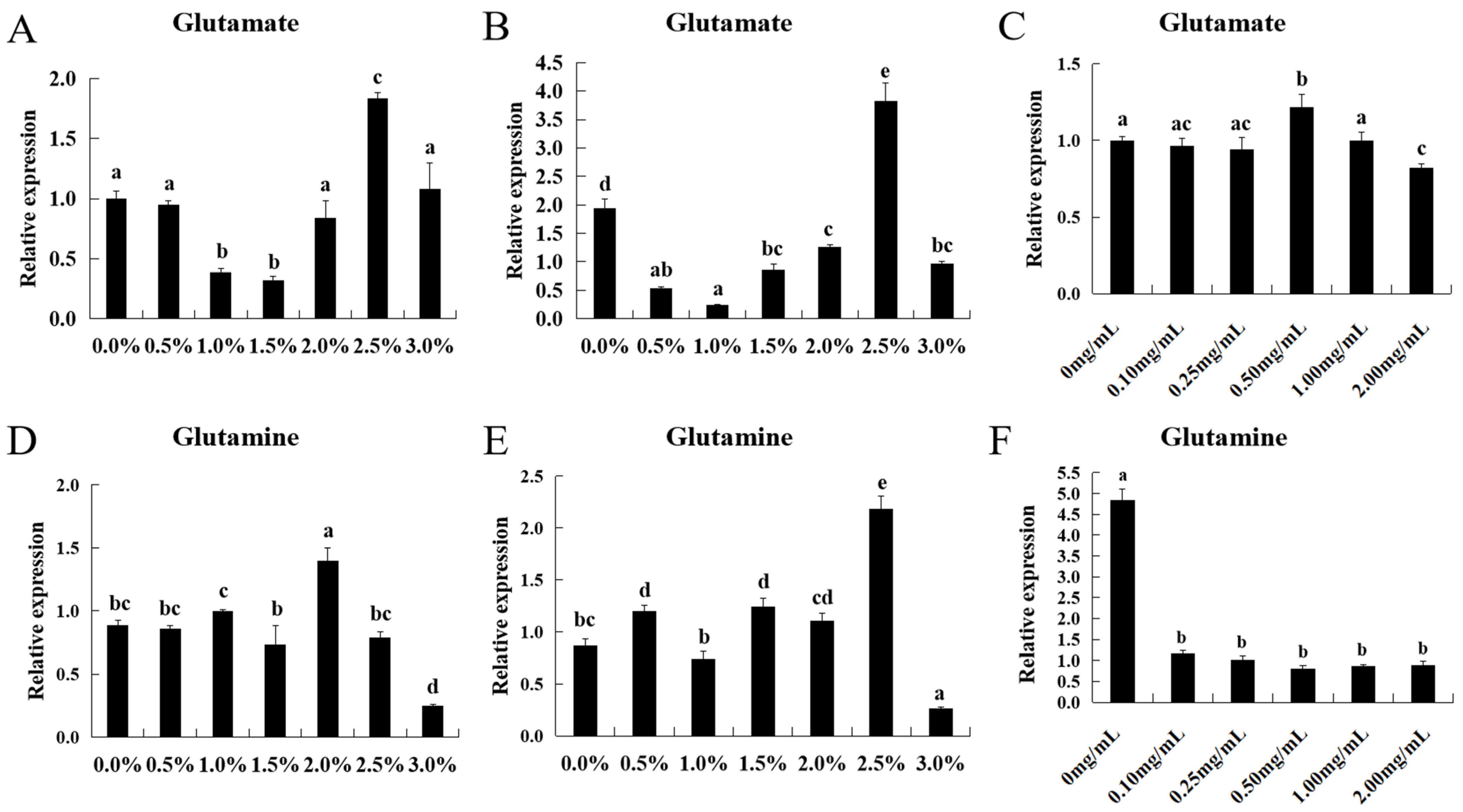
In order to analyze the effects of different proportions of dietary glutamate and glutamine additives on ASCT2, the fish were randomly divided into 13 groups (
n = 15 for each group, body weight of fish was 300.35 ± 0.04 g). The formula for the control group contained 32% CP, while the formula for the glutamate/glutamine groups was supplemented with 0.5, 1.0, 1.5, 2.0, 2.5, 3.0 g/kg glutamate or glutamine (
Table 2 and
Table 3). The fish were acclimatized for 2 weeks to an outdoor cage (1.2 m × 1.2 m × 2.0 m, water depth: 1.8 m). They were fed on the control diet twice daily at 8:30 a.m. and 2:30 p.m., respectively. There were 585 acclimatized fish of the same age and size (each weighing 300.00 ± 10.15 g) batch-weighed and randomly distributed into 39 cages. For each of the 39 diet treatments, triplicate cages were set up with each cage having 15 fish. The fish were fed manually to apparent satiation twice daily (at 8:30 a.m. and 2:30 p.m.). Feeding was done for 60 days with each of the 13 diets to the allocated cages. After 60 d, the fish were fasted for 24 h and then sacrificed with 50 mg L
−1 MS-222 (tricaine methane sulfonate, Sigma-Aldrich, St. Louis, MO, USA) before sampling and the ASCT2 expression levels of intestine and muscle in triploid crucian carp (
n = 3 for each group) were assayed by qRT-PCR.
In order to deepen explore the effects of additive of glutamate and glutamine on ASCT2, in vitro, muscle cell of triploid crucian carp was used to carry out the experiment. During the experiment, the muscle was rapidly separated from triploid crucian carp (25 g) after being anesthetized with 2-phenoxyethanol. The muscle was cut into pieces by using scissors. After 3 washes with PBS containing 500 U/mL penicillin (Gibco, Waltham, MA, USA) and 500 U/mL streptomycin (Gibco, Waltham, MA, USA), minced muscle tissues were seeded in dishes with DMEM/F12 containing 20% FBS (Gibco, Waltham, MA, USA), 500 U/mL penicillin, 500 U/mL streptomycin and 15 ng/mL fibroblast growth factor (FGF, PEPROTECH, East Windsor, NJ, USA) in 28 °C. Afterward, culture medium was replaced with fresh medium by half every 3 days. Subculture was carried out at a split ratio of 1:2 when primary cell cultures grew to above 90% of confluence. After subculture of 3 passages, the cells were inoculated into 6-well plates and randomly divided into 6 groups of 4 wells. After passaging, the cells had adhered well and were normally extended (3rd day). The original culture solution was aspirated using a sterile pipette and gently washed with 2 mL PBS. Complete cell medium containing different concentrations of glutamate/glutamine (0, 0.10, 0.25, 0.50, 1.00 and 2.00 mg/mL) was added to each group of cells. Because HEPES was added to DMEM/F12 medium, the pH of the medium remained stable (pH = 7.2). After 8 h of culture, we removed the cell culture medium and collected the cells. The relative mRNA expression levels of ASCT2 were detected by qRT-PCR.
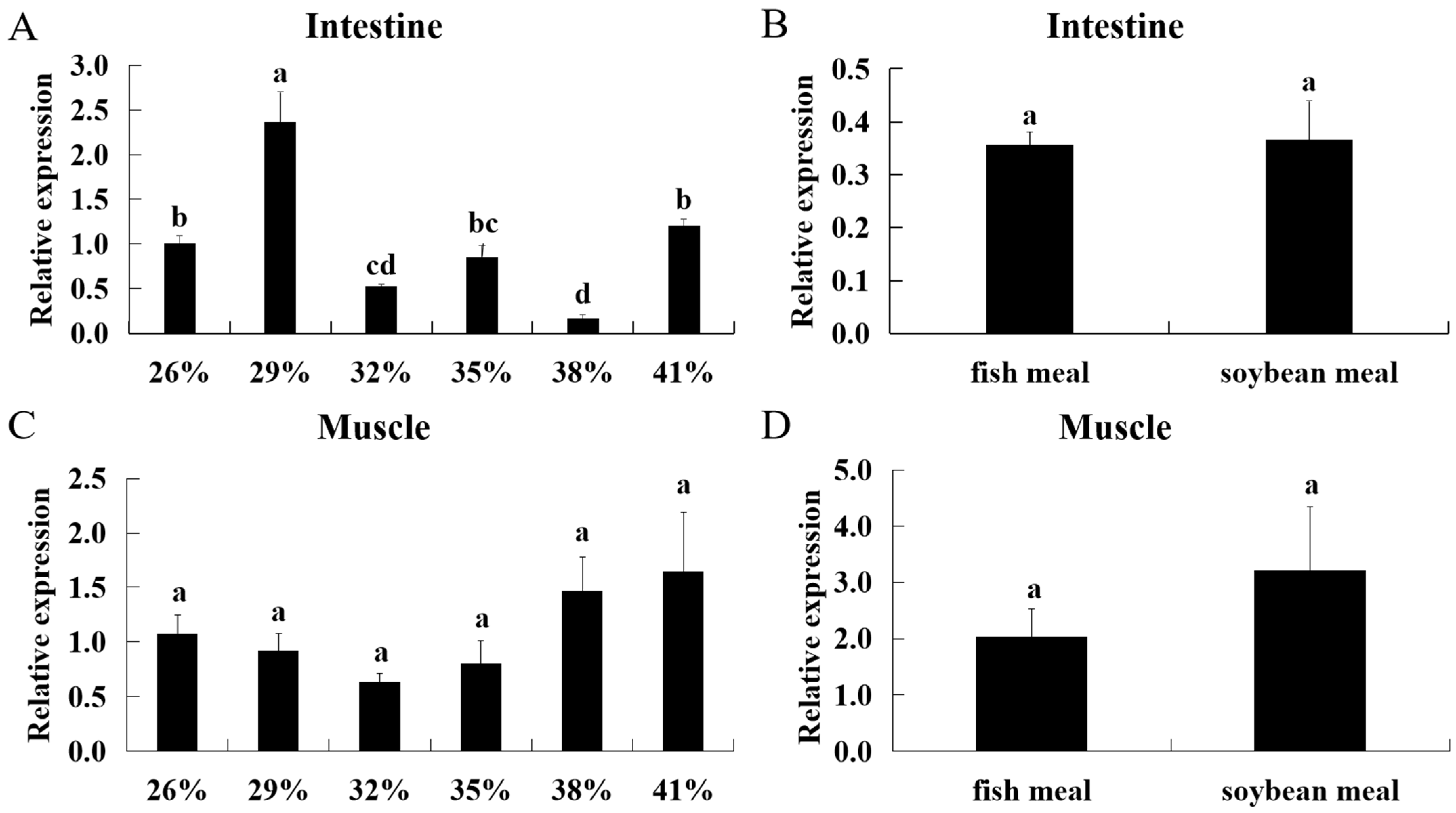
2.8. Regulation of Dietary Protein Levels and Protein Sources on ASCT2
To reveal the effects of dietary protein levels on triploid crucian carp intestine ASCT2 expression, 6 isocaloric diets with 5 different dietary protein levels at 26, 29, 32, 35, 38 and 41%, respectively, were formulated (
Table 4). To reveal the effects of different protein sources on triploid crucian carp intestine ASCT2 expression, two protein diets of fish meal and soybean meal were formulated (
Table 5). The diets of different protein levels and sources were prepared by thoroughly mixing dry ingredients with oil and then water until a stiff dough was obtained. The dough was then passed through a meat mincer equipped with a 2 mm diameter, and the products were dried using an electrical fan at 28 °C. After drying, the material was broken up into regular pieces sieved to a convenient pellet size and stored at 20 °C. Juvenile triploids (body weight 11.72 ± 0.11 g) were cultured in 8 fiberglass tanks (1.2 m height 0.8 m depth;
n = 25 per tank). Before the experiment, the fish were fasted for 24 h and then fed with different diets for 60 d. The fish were fed manually to apparent satiation twice daily, at 8:30 a.m. and 2:30 p.m. After 60 d, the fish were fasted for 24 h and then sacrificed with 50 mg L
−1 MS-222 (tricaine methane sulfonate, Sigma-Aldrich, St. Louis, MO, USA) before sampling. Finally, the intestine of triploid crucian carp (
n = 3 for each group) was collected for gene expression analysis by qRT-PCR.
2.9. Statistical Analysis
The data analysis was measured by using the SPSS 18 analysis program and represented as means ± standard error (SE) [
26]. The significant difference between groups was confirmed using one-way ANOVA analysis. Differences between mean values of individual treatments were separated using the Least Significance Difference (LSD) test at
p < 0.05.
4. Discussion
For aquaculture, dietary protein plays a crucial role in growth and development. ASCT2, which is a transporter of glutamine, plays an important role in dietary protein absorption. In this study, the sequence and expression characterization of ASCT2 in triploid crucian carp were analyzed. The sequence of ASCT2 in triploid crucian carp was cloned, and its ORF region shared the highest sequence homology with that in Cyprinus carpio, followed by Sinocyclocheilus anshuiensis. The phylogenetic tree showed that the ASCT2 of triploid crucian carp shared high sequence homology in cyprinid fish. Moreover, compared with amphibians and mammals, ASCT2 showed the higher homology among fish analyzed in this study. Taken together, it indicates that the evolution of ASCT2 is in accordance with classical taxonomy.
Gene is the internal factor that determines life and health, mRNA abundance in different tissues can reflect its primary function. In mammals, ASCT2 is widely expressed in many tissues, such as lung, skeletal muscle, kidney, testis, intestine, brain, placenta, and pancreas [
27,
28,
29]. Moreover, the expression levels of ASCT2 in various tissues are different [
30,
31]. In our previous study of grass carp, ASCT2 was widely expressed in intestine, liver, brain, heart, kidney, gill, muscle, hypophysis, spleen and gonad [
15]. In this study, the ASCT2 expression pattern in triploid crucian carp was analyzed. The results showed that ASCT2 is widely distributed in all the tissues examined in this study, including heart, spleen, brain, intestine, liver, kidney and muscle. It indicates that ASCT2 is widely distributed in various tissues. Nevertheless, there were significant differences in the mRNA abundance of ASCT2 in different tissues. Among them, the ASCT2 was highly expressed in muscle. This result was consistent with our previous study in grass carp, which indicated that ASCT2 played an important role in muscle. The circadian clock is a conserved pathway among all light-exposed organisms that enables them to maintain an endogenous rhythm of one day [
32]. Individual tissues show different response to circadian oscillators [
33]. Thus, we further explored the diurnal variation of ASCT2 in muscle. Within a day of 24 h, muscle appeared with high expression levels at 6:00, 15:00 and 24:00. It showed a cyclical change of 9 h during the day, which had reference value for the improvement of feeding strategy. In zebra fish, it is demonstrated that light can dramatically increase the expression of the clock genes period2 and cryptochrome 1a in vivo and in vitro [
34,
35]. In addition, the oxygen levels can also affect the circadian clock [
36]. Thus, we supposed that the variation of diurnal variation in triploid crucian carp might be related to light and oxygen levels, which needs to be further explored.
Glutamine is the main respiratory substrate of intestinal cells, which can significantly improve the morphology and function of the intestine [
11]. In this study, we found that the additions of glutamate and glutamine in daily feed increased the FCR and FR in triploid crucian carp. These results were consistent with the research in Atlantic salmon (
Salmo salar L.) and grass carp [
37,
38]. By comparing the effects of glutamate additive on expression levels of ASCT2 between intestine and muscle, we found they had a similar trend. When the addition of glutamate in the feed was low, the expression levels of ASCT2 decreased with the increase of glutamate additives. We speculate it may be related to the change of proton concentration, as ASCT2 is a Na
+-dependent neutral amino acid transporter. Then, the expression levels of ASCT2 increased with the increase of glutamate additives, which indicated that glutamate could increase the expression level of ASCT2. Later, the expression levels of ASCT2 decreased significantly with the increase of glutamate additives (
p < 0.05). We speculated that it was a self-protection mechanism of ASCT2. By comparing the effects of glutamate on the expression levels in muscle between in vivo and in vitro, we found that they shared the similar trend. It is worth noting that ASCT2 is less sensitive to glutamate in vitro compared to in vivo, which may be due to the stable proton concentration in the medium. Compared to the glutamate group, the expression pattern of ASCT2 in the glutamine group showed the more complex trend. It indicates that ASCT2 is more sensitive to glutamine compared to glutamate. When the additive of glutamine in the feed was low, the expression levels of ASCT2 in the intestines and muscles showed oscillating changes, although the changes were not obvious. However, the expression levels of ASCT2 in the intestine and muscles showed significant increase when the glutamine additive reached 2.0 and 2.5%, respectively (
p < 0.05). These results indicate that glutamine can increase the expression of ASCT2. After that, the expression levels of ASCT2 showed significant decrease (
p < 0.05), which indicated that extra addition of glutamine could not further increase the expression level of ASCT2. In vitro, the expression levels of ASCT2 showed a sharp decline in the glutamine additive groups, which might be related to a certain concentration of glutamine in medium. It also indicates that ASCT2 is more sensitive to glutamine compared to glutamate.
Compared with plant-source protein, animal-source protein contains antioxidants such as taurine, creatine and phosphocreatine, and plays an important role in the energy metabolism of tissues [
39]. In the feedstuff of fish, fish meal is the main source of protein. Given the global context of rapid development of aquaculture, the feedstuff costs are increasing and the protein-rich feedstuffs are lacking [
40,
41]. There has been an impending necessity to optimize dietary supply in tune with the different physiological needs of organisms [
41]. The previous study of triploid crucian carp demonstrated that the different protein levels can affect the transcription levels of some transfer proteins, which play the important roles in nutrition regulation, such as DGH, PepT1 and LAT2 [
18,
19,
20]. In the present study, we found fish fed a low-protein (29%) diet had the highest expression level of intestinal ASCT2 (
p < 0.05). On the contrary, the lowest expression level occurred when fish were fed the diet containing 38% protein (
p < 0.05). These results indicate that the lowprotein diet can promote the absorption of glutamine. Although the anti-nutritional factors in plant protein could affect nutrient absorption, the results in the present study showed that there was no significant effect on the expression levels of ASCT2 between fish meal and soybean meal.


 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}