
A Multidisciplinary Approach Evaluating Soybean Meal-Induced Enteritis in Rainbow Trout Oncorhynchus mykiss



Abstract
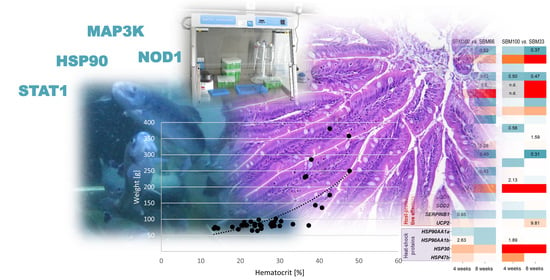
:
1. Introduction
2. Materials and Methods
2.1. SBM Feeding Trial
2.1.1. Diets
2.1.2. Fish, Husbandry Conditions and Feeding Trial
2.1.3. Sampling
2.2. Assessment of Body Condition Indices
2.3. Haematocrit Determination
2.4. Differential Blood Count
2.5. Quantification of Serum Cortisol Levels
2.6. Histopathological Evaluation of the Intestine
2.7. Real-Time Quantitative PCR (RT-qPCR)
2.8. Data Analysis and Statistics
3. Results
3.1. Overall Health, Growth and Performance
3.2. Quantitative Nutrient Compositions of the Fish Body
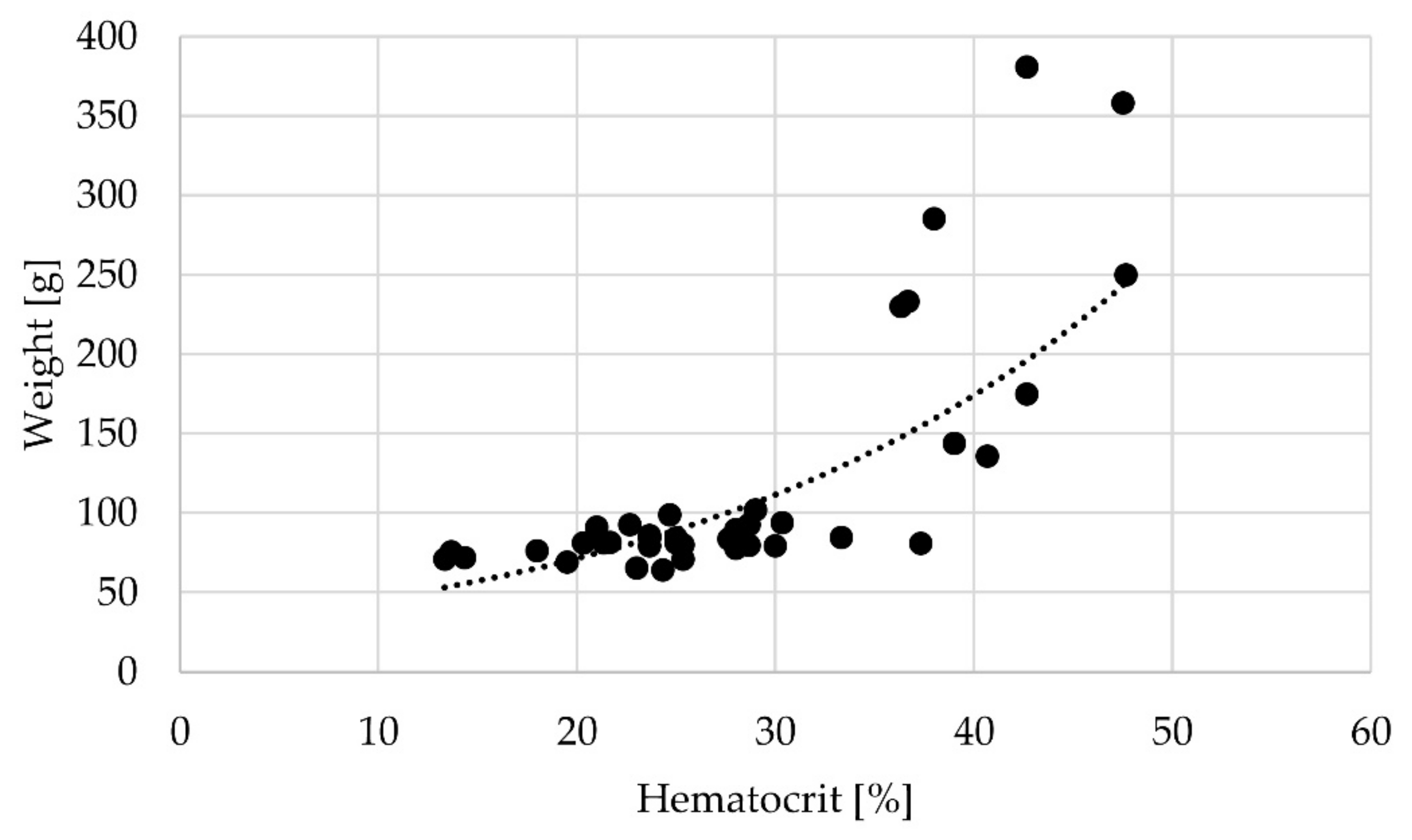
3.3. Haematocrit
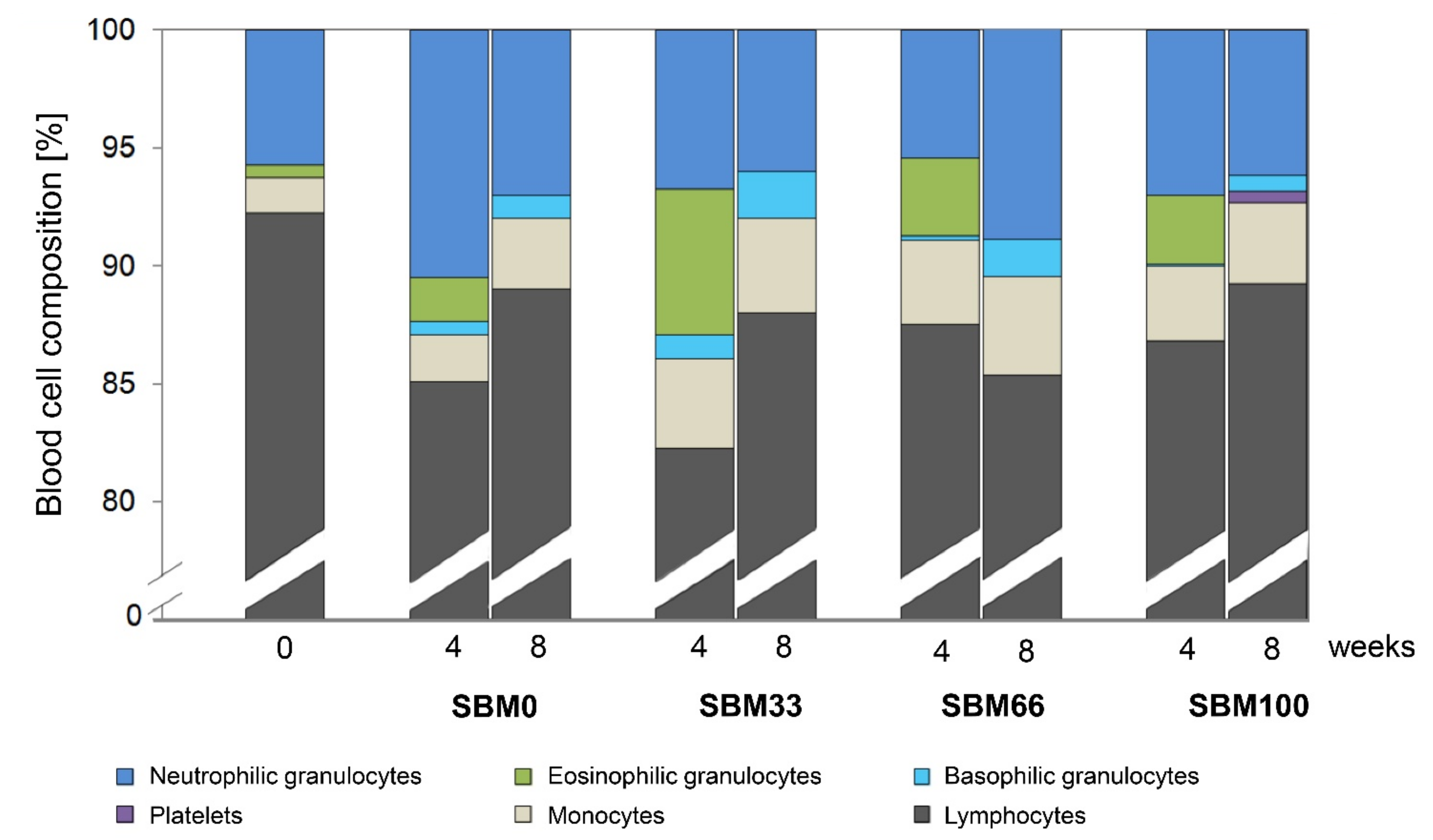
3.4. Differential Blood Count
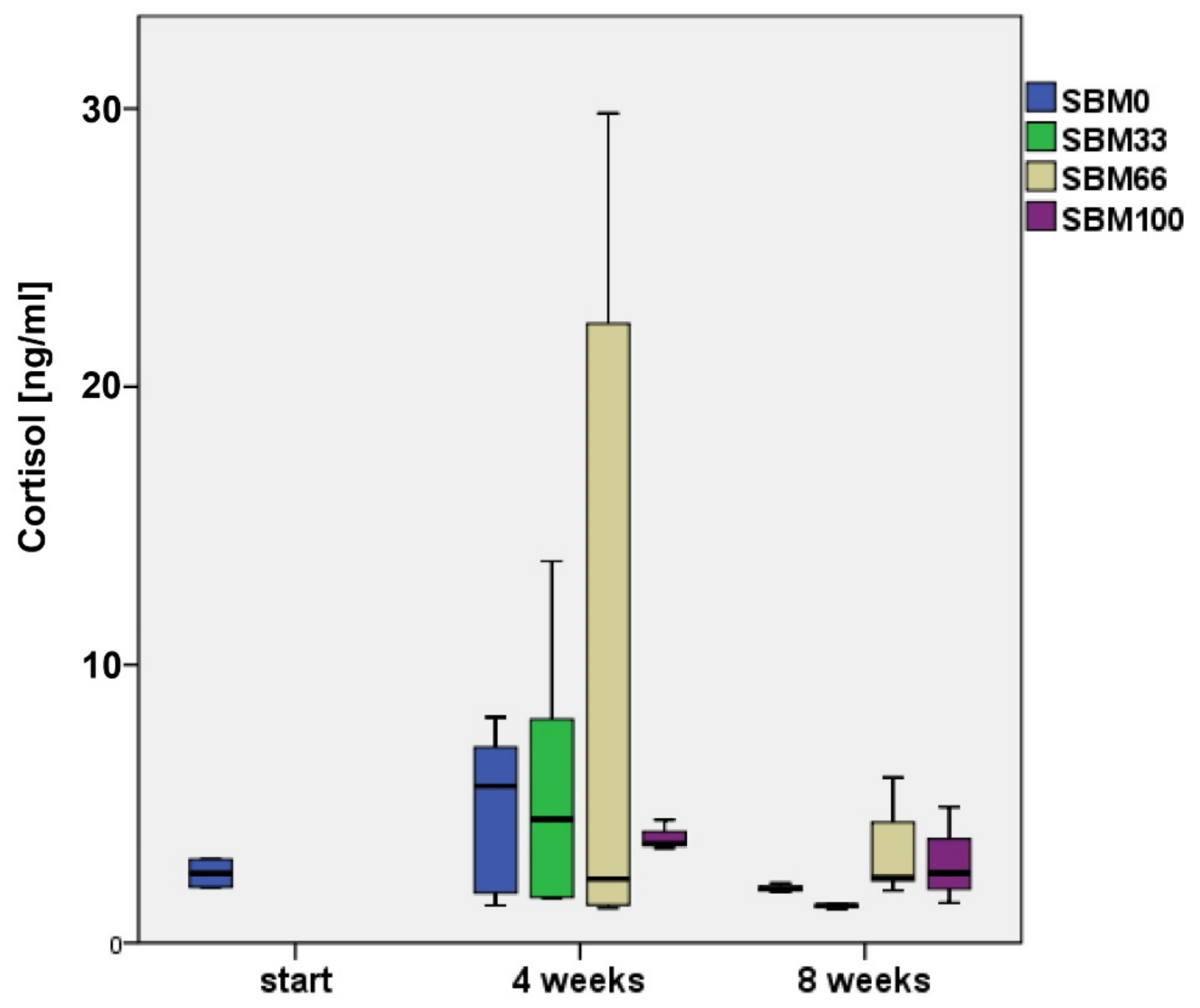
3.5. Cortisol
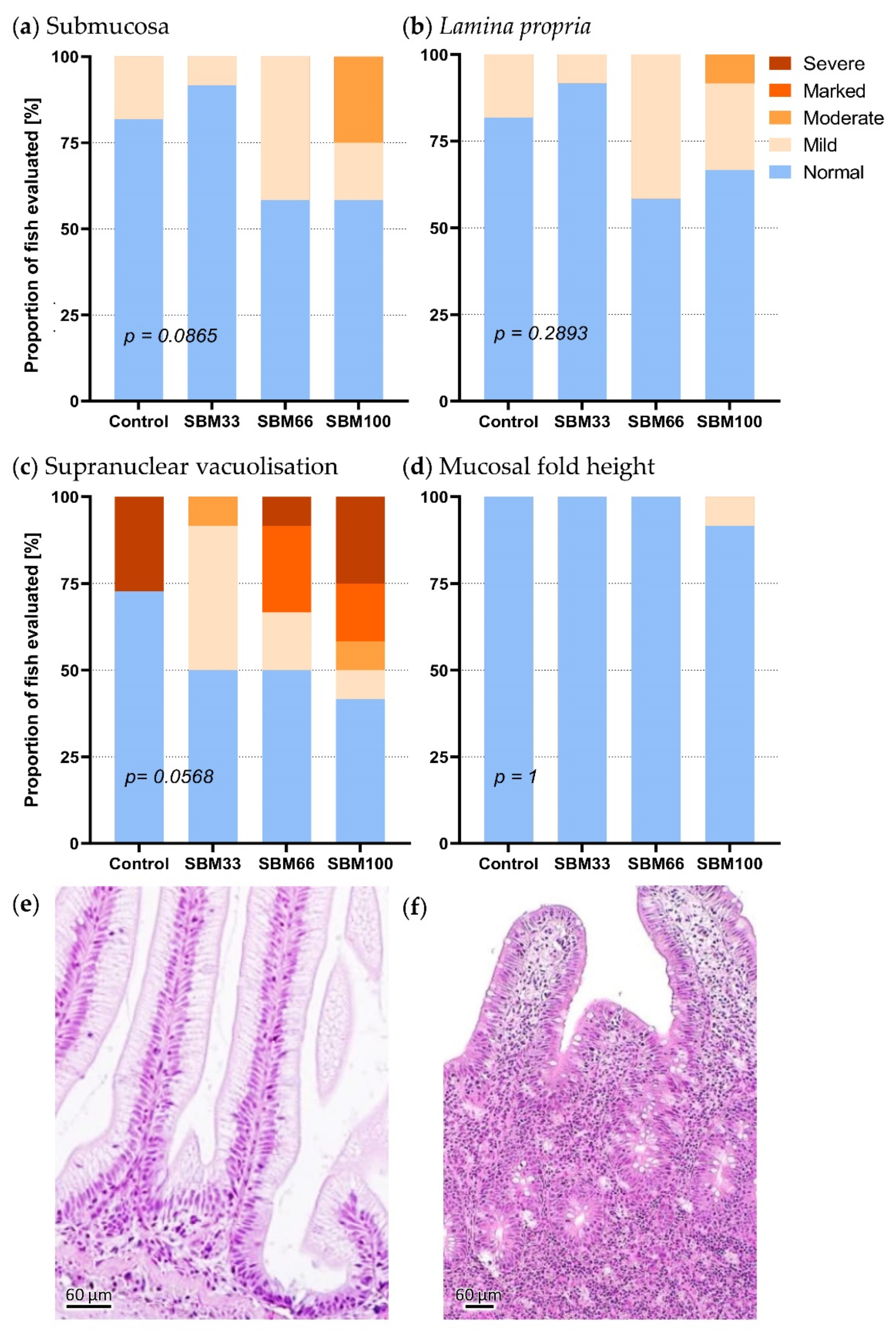
3.6. Histopathology
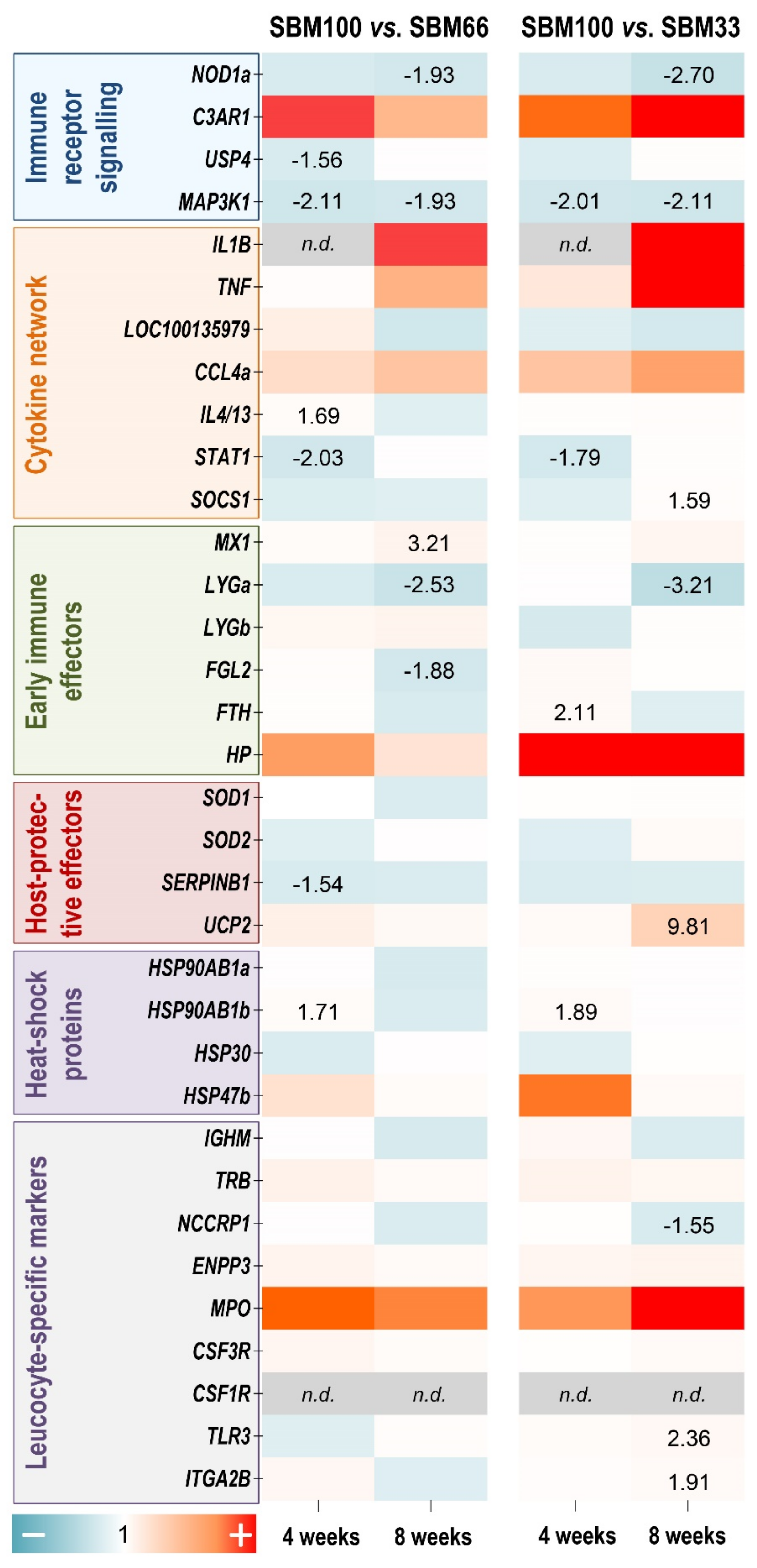
3.7. Gene Profiling
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Farstad, W. Ethics in animal breeding. Reprod. Domest. Anim. 2018, 53, 4–13. [Google Scholar] [CrossRef]
- Bergleiter, S.; Berner, N.; Censkowsky, U.; Julià-Camprodon, G. Naturland e.V. and Organic Services GmbH Organic Aquaculture 2009 Production and Markets. 2009. Available online: www.naturland.de (accessed on 4 January 2022).
- Tschirren, L.; Bachmann, D.; Güler, A.C.; Blaser, O.; Rhyner, N.; Seitz, A.; Zbinden, E.; Wahli, T.; Segner, H.; Refardt, D. MyFishCheck: A Model to Assess Fish Welfare in Aquaculture. Animals 2021, 11, 145. [Google Scholar] [CrossRef]
- Hvas, M.; Folkedal, O.; Oppedal, F. Fish welfare in offshore salmon aquaculture. Rev. Aquac. 2021, 13, 836–852. [Google Scholar] [CrossRef]
- Rabadan, C.G.; Spreadbury, C.; Consuegra, S.; de Leaniz, C.G. Development, validation and testing of an Operational Welfare Score Index for farmed lumpfish Cyclopterus lumpus L. Aquaculture 2021, 531, 735777. [Google Scholar] [CrossRef]
- Stien, L.H.; Bracke, M.B.M.; Folkedal, O.; Nilsson, J.; Oppedal, F.; Torgersen, T.; Kittilsen, S.; Midtlyng, P.J.; Vindas, M.A.; Øverli, Ø.; et al. Salmon Welfare Index Model (SWIM 1.0): A semantic model for overall welfare assessment of caged Atlantic salmon: Review of the selected welfare indicators and model presentation. Rev. Aquac. 2013, 5, 33–57. [Google Scholar] [CrossRef]
- Ings, J.S.; Oakes, K.D.; Vijayan, M.M.; Servos, M.R. Temporal changes in stress and tissue-specific metabolic responses to municipal wastewater effluent exposure in rainbow trout. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2012, 156, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Rebl, A.; Verleih, M.; Nipkow, M.; Altmann, S.; Bochert, R.; Goldammer, T. Gradual and Acute Temperature Rise Induces Crossing Endocrine, Metabolic, and Immunological Pathways in Maraena Whitefish (Coregonus maraena). Front. Genet. 2018, 9, 241. [Google Scholar] [CrossRef] [PubMed]
- Quinn, N.L.; McGowan, C.R.; Cooper, G.A.; Koop, B.F.; Davidson, W.S. Ribosomal genes and heat shock proteins as putative markers for chronic, sublethal heat stress in Arctic charr: Applications for aquaculture and wild fish. Physiol. Genom. 2011, 43, 1056–1064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernier, N.J.; Craig, P.M. CRF-related peptides contribute to stress response and regulation of appetite in hypoxic rainbow trout. Am. J. Physiol. Integr. Comp. Physiol. 2005, 289, R982–R990. [Google Scholar] [CrossRef]
- Delaney, M.A.; Klesius, P.H. Hypoxic conditions induce Hsp70 production in blood, brain and head kidney of juvenile Nile tilapia Oreochromis niloticus (L.). Aquaculture 2004, 236, 633–644. [Google Scholar] [CrossRef]
- Caipang, C.M.; Berg, I.; Brinchmann, M.F.; Kiron, V. Short-term crowding stress in Atlantic cod, Gadus morhua L. modulates the humoral immune response. Aquaculture 2009, 295, 110–115. [Google Scholar] [CrossRef]
- Yarahmadi, P.; Miandare, H.K.; Hoseinifar, S.H.; Gheysvandi, N.; Akbarzadeh, A. The effects of stocking density on hemato-immunological and serum biochemical parameters of rainbow trout (Oncorhynchus mykiss). Aquac. Int. 2014, 23, 55–63. [Google Scholar] [CrossRef]
- Korytář, T.; Nipkow, M.; Altmann, S.; Goldammer, T.; Köllner, B.; Rebl, A. Adverse Husbandry of Maraena Whitefish Directs the Immune System to Increase Mobilization of Myeloid Cells and Proinflammatory Responses. Front. Immunol. 2016, 7, 631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebrahimi, G. Effects of Rearing Tank Background Color on Growth Performance in Juvenile Common Carp, Cyprinus carpio L. Agric. J. 2011, 6, 213–217. [Google Scholar] [CrossRef] [Green Version]
- Lopez, P.A.; Díaz-Gil, C.; Saraiva, J.; Moranta, D.; Castanheira, M.; Nuñez-Velázquez, S.; Ledesma-Corvi, S.; Mora-Ruiz, M.; Grau, A. Effects of structural environmental enrichment on welfare of juvenile seabream (Sparus aurata). Aquac. Rep. 2019, 15, 100224. [Google Scholar] [CrossRef]
- Benhaïm, D.; Bégout, M.-L.; Péan, S.; Manca, M.; Prunet, P.; Chatain, B. Impact of a plant-based diet on behavioural and physiological traits in sea bass (Dicentrarchus labrax). Aquat. Living Resour. 2013, 26, 121–131. [Google Scholar] [CrossRef] [Green Version]
- Mullally, M.; Mimeault, C.; Rojas, M.O.; Vindas, P.S.; Garcia, M.; Alvarez, L.P.; Moon, T.W.; Gilmour, K.M.; Trudeau, V.L.; Arnason, J.T. A botanical extract of Souroubea sympetala and its active principle, betulinic acid, attenuate the cortisol response to a stressor in rainbow trout, Oncorhynchus mykiss. Aquaculture 2017, 468, 26–31. [Google Scholar] [CrossRef]
- Oxley, A.; Jolly, C.; Eide, T.; Jordal, A.-E.O.; Svardal, A.; Olsen, R.-E. The combined impact of plant-derived dietary ingredients and acute stress on the intestinal arachidonic acid cascade in Atlantic salmon (Salmo salar). Br. J. Nutr. 2010, 103, 851–861. [Google Scholar] [CrossRef] [Green Version]
- Olsen, R.L.; Hasan, M.R. A limited supply of fishmeal: Impact on future increases in global aquaculture production. Trends Food Sci. Technol. 2012, 27, 120–128. [Google Scholar] [CrossRef]
- Barnes, M.E.; Brown, M.L.; Bruce, T.; Sindelar, S.; Neiger, R. Rainbow Trout Rearing Performance, Intestinal Morphology, and Immune Response after Long-term Feeding of High Levels of Fermented Soybean Meal. N. Am. J. Aquac. 2014, 76, 333–345. [Google Scholar] [CrossRef]
- Oliva-Teles, A.; Gouveia, A.; Gomes, E.; Rema, P. The effect of different processing treatments on soybean meal utilization by rainbow trout, Oncorhynchus mykiss. Aquaculture 1994, 124, 343–349. [Google Scholar] [CrossRef]
- Voorhees, J.M.; Barnes, M.E.; Chipps, S.R.; Brown, M.L. Effects of Exercise and Bioprocessed Soybean Meal Diets during Rainbow Trout Rearing. Open Biol. J. 2019, 7, 1–13. [Google Scholar] [CrossRef]
- Bruni, L.; Secci, G.; Husein, Y.; Faccenda, F.; de Medeiros, A.C.L.; Parisi, G. Is it possible to cut down fishmeal and soybean meal use in aquafeed limiting the negative effects on rainbow trout (Oncorhynchus mykiss) fillet quality and consumer acceptance? Aquaculture 2021, 543, 736996. [Google Scholar] [CrossRef]
- Rumsey, G.L.; Siwicki, A.K.; Anderson, D.P.; Bowser, P.R. Effect of soybean protein on serological response, non-specific defense mechanisms, growth, and protein utilization in rainbow trout. Vet.-Immunol. Immunopathol. 1994, 41, 323–339. [Google Scholar] [CrossRef]
- Van den Ingh, T.S.G.A.M.; Krogdahl, Å.; Olli, J.J.; Hendriks, H.G.C.J.M.; Koninkx, J.G.J.F. Effects of soybean-containing diets on the proximal and distal intestine in Atlantic salmon (Salmo salar): A morphological study. Aquaculture 1991, 94, 297–305. [Google Scholar] [CrossRef]
- Francis, G.; Makkar, H.P.S.; Becker, K. Antinutritional factors present in plant-derived alternate fish feed ingredients and their effects in fish. Aquaculture 2001, 199, 197–227. [Google Scholar] [CrossRef]
- Krogdahl, A.; Lea, T.B.; Olli, J.J. Soybean proteinase inhibitors affect intestinal trypsin activities and amino acid digestibilities in rainbow trout (Oncorhynchus mykiss). Comp. Biochem. Physiol. Part A Physiol. 1994, 107, 215–219. [Google Scholar] [CrossRef]
- Krogdahl, A.; Penn, M.; Thorsen, J.; Refstie, S.; Bakke, A.M. Important antinutrients in plant feedstuffs for aquaculture: An update on recent findings regarding responses in salmonids. Aquac. Res. 2010, 41, 333–344. [Google Scholar] [CrossRef]
- Liener, I.E. Implications of antinutritional components in soybean foods. Crit. Rev. Food Sci. Nutr. 1994, 34, 31–67. [Google Scholar] [CrossRef]
- Baeverfjord, G.; Krogdahl, A. Development and regression of soybean meal induced enteritis in Atlantic salmon, Salmo salar L., distal intestine: A comparison with the intestines of fasted fish. J. Fish Dis. 1996, 19, 375–387. [Google Scholar] [CrossRef]
- Chikwati, E.M.; Venold, F.F.; Penn, M.H.; Rohloff, J.; Refstie, S.; Guttvik, A.; Hillestad, M.; Krogdahl, A. Interaction of soyasaponins with plant ingredients in diets for Atlantic salmon, Salmo salar L. Br. J. Nutr. 2012, 107, 1570–1590. [Google Scholar] [CrossRef] [Green Version]
- Knudsen, D.; Jutfelt, F.; Sundh, H.; Sundell, K.; Koppe, W.; Frøkiær, H. Dietary soya saponins increase gut permeability and play a key role in the onset of soyabean-induced enteritis in Atlantic salmon (Salmo salar L.). Br. J. Nutr. 2008, 100, 120–129. [Google Scholar] [CrossRef] [Green Version]
- Merrifield, D.L.; Olsen, R.E.; Myklebust, R.; Ringø, E.; El-Shemy, H. Dietary Effect of Soybean (Glycine max) Products on Gut Histology and Microbiota of Fish. Soybean Nutr. 2011, 231–250. [Google Scholar] [CrossRef] [Green Version]
- Booman, M.; Forster, I.; Vederas, J.C.; Groman, D.B.; Jones, S.R. Soybean meal-induced enteritis in Atlantic salmon (Salmo salar) and Chinook salmon (Oncorhynchus tshawytscha) but not in pink salmon (O. gorbuscha). Aquaculture 2018, 483, 238–243. [Google Scholar] [CrossRef]
- Buttle, L.; Burrells, A.; Good, J.; Williams, P.; Southgate, P. The binding of soybean agglutinin (SBA) to the intestinal epithelium of Atlantic salmon, Salmo salar and Rainbow trout, Oncorhynchus mykiss, fed high levels of soybean meal. Vet.-Immunol. Immunopathol. 2001, 80, 237–244. [Google Scholar] [CrossRef]
- Zhang, J.-X.; Guo, L.-Y.; Feng, L.; Jiang, W.-D.; Kuang, S.-Y.; Liu, Y.; Hu, K.; Jiang, J.; Li, S.-H.; Tang, L.; et al. Soybean β-Conglycinin Induces Inflammation and Oxidation and Causes Dysfunction of Intestinal Digestion and Absorption in Fish. PLoS ONE 2013, 8, e58115. [Google Scholar] [CrossRef] [Green Version]
- Miao, S.; Zhao, C.; Zhu, J.; Hu, J.; Dong, X.; Sun, L. Dietary soybean meal affects intestinal homoeostasis by altering the microbiota, morphology and inflammatory cytokine gene expression in northern snakehead. Sci. Rep. 2018, 8, 113. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.-R.; Wang, L.; Zhang, C.-X.; Song, K. Effects of substituting fishmeal with soybean meal on growth performance and intestinal morphology in orange-spotted grouper (Epinephelus coioides). Aquac. Rep. 2017, 5, 52–57. [Google Scholar] [CrossRef]
- Blaufuss, P.C.; Gaylord, G.; Sealey, W.M.; Powell, M.S. Effects of high-soy diet on S100 gene expression in liver and intestine of rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immunol. 2019, 86, 764–771. [Google Scholar] [CrossRef]
- Holen, E.; Winterthun, S.; Du, Z.-Y.; Krøvel, A. Inhibition of p38 MAPK during cellular activation modulate gene expression of head kidney leukocytes isolated from Atlantic salmon (Salmo salar) fed soy bean oil or fish oil based diets. Fish Shellfish Immunol. 2011, 30, 397–405. [Google Scholar] [CrossRef]
- NRC (National Research Council). Nutrient Requirements of Fish and Shrimp; Natlional Academic Press: Washington, DC, USA, 2011. [CrossRef]
- Rebl, A.; Anders, E.; Wimmers, K.; Goldammer, T. Characterization of Dehydrodolichyl diphosphate synthase gene in rainbow trout (Oncorhynchus mykiss). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2009, 152, 260–265. [Google Scholar] [CrossRef]
- Bowers, R.M.; Lapatra, S.E.; Dhar, A.K. Detection and quantitation of infectious pancreatic necrosis virus by real-time reverse transcriptase-polymerase chain reaction using lethal and non-lethal tissue sampling. J. Virol. Methods 2008, 147, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Rebl, A.; Rebl, H.; Korytář, T.; Goldammer, T.; Seyfert, H.-M. The proximal promoter of a novel interleukin-8-encoding gene in rainbow trout (Oncorhynchus mykiss) is strongly induced by CEBPA, but not NF-κB p65. Dev. Comp. Immunol. 2014, 46, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Kortner, T.M.; Valen, E.C.; Kortner, H.; Marjara, I.S.; Krogdahl, A.; Bakke, A.M. Candidate reference genes for quantitative real-time PCR (qPCR) assays during development of a diet-related enteropathy in Atlantic salmon (Salmo salar L.) and the potential pitfalls of uncritical use of normalization software tools. Aquaculture 2011, 318, 355–363. [Google Scholar] [CrossRef]
- Andersen, C.L.; Jensen, J.L.; Ørntoft, T.F. Normalization of Real-Time Quantitative Reverse Transcription-PCR Data: A Model-Based Variance Estimation Approach to Identify Genes Suited for Normalization, Applied to Bladder and Colon Cancer Data Sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [Green Version]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034.1. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Green, T.; Smullen, R.; Barnes, A. Dietary soybean protein concentrate-induced intestinal disorder in marine farmed Atlantic salmon, Salmo salar is associated with alterations in gut microbiota. Vet. Microbiol. 2013, 166, 286–292. [Google Scholar] [CrossRef]
- Krogdahl, A.; Bakke-McKellep, A.M.; Roed, K.H.; Baeverfjord, G. Feeding Atlantic salmon Salmo salar L. soybean products: Effects on disease resistance (furunculosis), and lysozyme and IgM levels in the intestinal mucosa. Aquac. Nutr. 2000, 6, 77–84. [Google Scholar] [CrossRef] [Green Version]
- Heikkinen, J.; Vielma, J.; Kemiläinen, O.; Tiirola, M.; Eskelinen, P.; Kiuru, T.; Navia-Paldanius, D.; von Wright, A. Effects of soybean meal based diet on growth performance, gut histopathology and intestinal microbiota of juvenile rainbow trout (Oncorhynchus mykiss). Aquaculture 2006, 261, 259–268. [Google Scholar] [CrossRef]
- Kemski, M.M.; Rappleye, C.A.; Dabrowski, K.; Bruno, R.S.; Wick, M. Transcriptomic response to soybean meal-based diets as the first formulated feed in juvenile yellow perch (Perca flavescens). Sci. Rep. 2020, 10, 3998. [Google Scholar] [CrossRef] [Green Version]
- Valenzuela, L.; Pacheco, S.; Rincón, G.; Pavez, L.; Lam, N.; Hernández, A.; Dantagnan, P.; González, F.; Jilberto, F.; Ravanal, M.; et al. Intestinal Transcriptome Analysis Reveals Enrichment of Genes Associated with Immune and Lipid Mechanisms, Favoring Soybean Meal Tolerance in High-Growth Zebrafish (Danio Rerio). Genes 2021, 12, 700. [Google Scholar] [CrossRef] [PubMed]
- Seibel, H.; Baßmann, B.; Rebl, A. Blood Will Tell: What Hematological Analyses Can Reveal About Fish Welfare. Front. Veter. Sci. 2021, 8, 194. [Google Scholar] [CrossRef]
- Hirano, M.; Osada, S.-I.; Aoki, T.; Hirai, S.-I.; Hosaka, M.; Inoue, J.-I.; Ohno, S. MEK Kinase Is Involved in Tumor Necrosis Factor α-Induced NF-κB Activation and Degradation of IκB-α. J. Biol. Chem. 1996, 271, 13234–13238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, M.; Dai, T.; Deak, J.C.; Kyriakis, J.M.; Zon, L.I.; Woodgett, J.; Templeton, D.J. Activation of stress-activated protein kinase by MEKK1 phosphorylation of its activator SEK1. Nature 1994, 372, 798–800. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Cui, K.; Fang, W.; Chen, Q.; Xu, D.; Mai, K.; Zhang, Y.; Ai, Q. High level of dietary olive oil decreased growth, increased liver lipid deposition and induced inflammation by activating the p38 MAPK and JNK pathways in large yellow croaker (Larimichthys crocea). Fish Shellfish Immunol. 2019, 94, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Jiang, W.-D.; Jiang, J.; Zhao, J.; Liu, Y.; Zhang, Y.-A.; Zhou, X.-Q.; Feng, L. Dietary choline deficiency and excess induced intestinal inflammation and alteration of intestinal tight junction protein transcription potentially by modulating NF-κB, STAT and p38 MAPK signaling molecules in juvenile Jian carp. Fish Shellfish Immunol. 2016, 58, 462–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Feng, L.; Jiang, W.-D.; Liu, Y.; Wu, P.; Kuang, S.-Y.; Tang, L.; Tang, W.-N.; Zhang, Y.-A.; Zhou, X.-Q. Vitamin A deficiency suppresses fish immune function with differences in different intestinal segments: The role of transcriptional factor NF-κB and p38 mitogen-activated protein kinase signalling pathways. Br. J. Nutr. 2017, 117, 67–82. [Google Scholar] [CrossRef] [Green Version]
- Booman, M.; Xu, Q.; Rise, M.L. Evaluation of the impact of camelina oil-containing diets on the expression of genes involved in the innate anti-viral immune response in Atlantic cod (Gadus morhua). Fish Shellfish Immunol. 2014, 41, 52–63. [Google Scholar] [CrossRef]
- Zheng, D.; Kille, P.; Feeney, G.P.; Cunningham, P.; Handy, R.D.; Hogstrand, C. Dynamic transcriptomic profiles of zebrafish gills in response to zinc supplementation. BMC Genom. 2010, 11, 553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craig, E.A.; Gambill, B.D.; Nelson, R.J. Heat shock proteins: Molecular chaperones of protein biogenesis. Microbiol. Rev. 1993, 57, 402–414. [Google Scholar] [CrossRef]
- Dash, S.; Das, S.; Samal, J.; Thatoi, H.N. Epidermal mucus, a major determinant in fish health: A review. Iran. J. Vet. Res. 2018, 19, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Keestra-Gounder, A.M.; Tsolis, R.M. NOD1 and NOD2: Beyond Peptidoglycan Sensing. Trends Immunol. 2017, 38, 758–767. [Google Scholar] [CrossRef] [PubMed]
- Dupont, C.; Cousin, P.; Dupont, S. IoT for Aquaculture 4.0 Smart and easy-to-deploy real-time water monitoring with IoT. In Proceedings of the 2018 Global Internet of Things Summit, Bilbao, Spain, 4–7 June 2018; pp. 1–5. [Google Scholar] [CrossRef]
- Antonucci, F.; Costa, C. Precision aquaculture: A short review on engineering innovations. Aquac. Int. 2020, 28, 41–57. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diet Composition | SBM0 | SBM33 | SBM66 | SBM100 |
|---|---|---|---|---|
| Fish meal 1 | 50.00 | 33.50 | 17.00 | 0.00 |
| Blood meal 3 | 5.00 | 5.00 | 5.00 | 5.00 |
| Gelatine 4 | 3.00 | 3.00 | 3.00 | 3.00 |
| Soybean meal 2 | 0.00 | 16.40 | 33.00 | 50.00 |
| Pea protein isolate 5 | 6.50 | 6.50 | 6.50 | 6.50 |
| Wheat gluten 6 | 7.00 | 10.59 | 14.05 | 17.25 |
| Wheat starch 7 | 19.50 | 15.00 | 10.55 | 5.50 |
| Fish oil 8 | 8.00 | 8.55 | 9.05 | 10.03 |
| Vitamins/Minerals 9 | 0.50 | 0.50 | 0.50 | 0.50 |
| Kaolin 10 | 0.00 | 0.47 | 0.85 | 1.22 |
| Calcium hydrogen phosphate 11 | 0.5 | 0.5 | 0.5 | 0.5 |
| Biolys 12 | 0.00 | 0.00 | 0.00 | 0.50 |
| SBM0 | SBM33 | SBM66 | SBM100 | |
|---|---|---|---|---|
| Dry matter (%) | 89.83 ± 0.02 | 88.35 ± 0.04 | 90.00 ± 0.02 | 90.00 ± 0.02 |
| Ash (% DM) | 10.48 ± 0.05 | 09.16 ± 0.05 | 07.86 ± 0.03 | 06.38 ± 0.00 |
| Crude protein (% DM) | 54.63 ± 0.09 | 55.40 ± 0.23 | 54.44 ± 0.13 | 54.31 ± 0.18 |
| Crude lipid (% DM) | 15.72 ± 0.04 | 14.77 ± 0.05 | 13.94 ± 0.05 | 13.61 ± 0.05 |
| Carbohydrates (% DM) 1 | 19.17 | 20.67 | 23.76 | 25.70 |
| Energy (MJkg−1) | 22.10 ± 0.01 | 22.19 ± 0.03 | 22.19 ± 0.00 | 22.44 ± 0.02 |
| SBM0 | SBM33 | SBM66 | SBM100 | |
|---|---|---|---|---|
| Initial weight [g] | 81.75 ± 0.16 | 82.09 ± 0.90 | 81.44 ± 0.85 | 82.15 ± 0.40 |
| Final weight [g] | 146.10 ± 10.49 | 138.26 ± 29.64 | 134.52 ± 8.66 | 106.74 ± 13.19 |
| K | 0.89 ± 0.24 | 0.95 ± 0.29 | 0.94 ± 0.22 | 0.88 ± 0.20 |
| DFI [%·d−1] | 2.43 ± 0.13 a,b | 2.70 ± 0.13 a | 2.69 ± 0.07 a | 2.32 ± 0.21 b |
| Start | SBM0 | SBM33 | SBM66 | SBM100 | |
|---|---|---|---|---|---|
| Moisture content [%] | 71.75 | 70.14 ± 1.540 | 74.27 ± 5.120 | 78.15 ±1.47 | 71.75 ± 1.34 |
| Ash content [%] | 02.79 | 02.79 ± 0.07 | 03.02 ± 0.330 | 03.23 ± 0.11 | 02.77 ± 0.12 |
| Protein content [%] | 16.26 | 17.01 ± 0.62 a | 16.16 ± 0.65 a,b | 15.68 ± 0.15 b | 16.34 ± 0.21 a,b |
| Fat content [%] | 09.20 | 10.05 ± 1.170 | 06.55 ± 4.800 | 06.55 ± 4.80 | 09.82 ± 1.04 |
| Energy content [MJ/kg] | 26.05 | 25.58 ± 0.440 | 23.58 ± 2.900 | 21.33 ± 1.19 | 25.73 ± 0.16 |
| SBM0 | SBM33 | SBM66 | SBM100 | |
|---|---|---|---|---|
| 4 weeks | 30.33 ± 6.17 | 29.50 ± 1.53 | 33.22 ± 7.99 | 22.67 ± 9.50 |
| 8 weeks | 23.39 ± 4.50 | 27.13 ± 11.40 | 30.40 ± 6.07 | 32.27 ± 11.25 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seibel, H.; Chikwati, E.; Schulz, C.; Rebl, A. A Multidisciplinary Approach Evaluating Soybean Meal-Induced Enteritis in Rainbow Trout Oncorhynchus mykiss. Fishes 2022, 7, 22. https://doi.org/10.3390/fishes7010022
Seibel H, Chikwati E, Schulz C, Rebl A. A Multidisciplinary Approach Evaluating Soybean Meal-Induced Enteritis in Rainbow Trout Oncorhynchus mykiss. Fishes. 2022; 7(1):22. https://doi.org/10.3390/fishes7010022
Chicago/Turabian StyleSeibel, Henrike, Elvis Chikwati, Carsten Schulz, and Alexander Rebl. 2022. "A Multidisciplinary Approach Evaluating Soybean Meal-Induced Enteritis in Rainbow Trout Oncorhynchus mykiss" Fishes 7, no. 1: 22. https://doi.org/10.3390/fishes7010022
APA StyleSeibel, H., Chikwati, E., Schulz, C., & Rebl, A. (2022). A Multidisciplinary Approach Evaluating Soybean Meal-Induced Enteritis in Rainbow Trout Oncorhynchus mykiss. Fishes, 7(1), 22. https://doi.org/10.3390/fishes7010022