
Phosphorus Absorption and Excretion in Hybrid Sturgeon (Huso dauricus♀ X Acipenser schrenckii♂) Intubated with Different Ca/P Ratios

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animal
2.2. Surgery Operation
2.3. Treatments
2.4. Sample Collection and Analysis
2.5. Statistical Analysis
3. Results
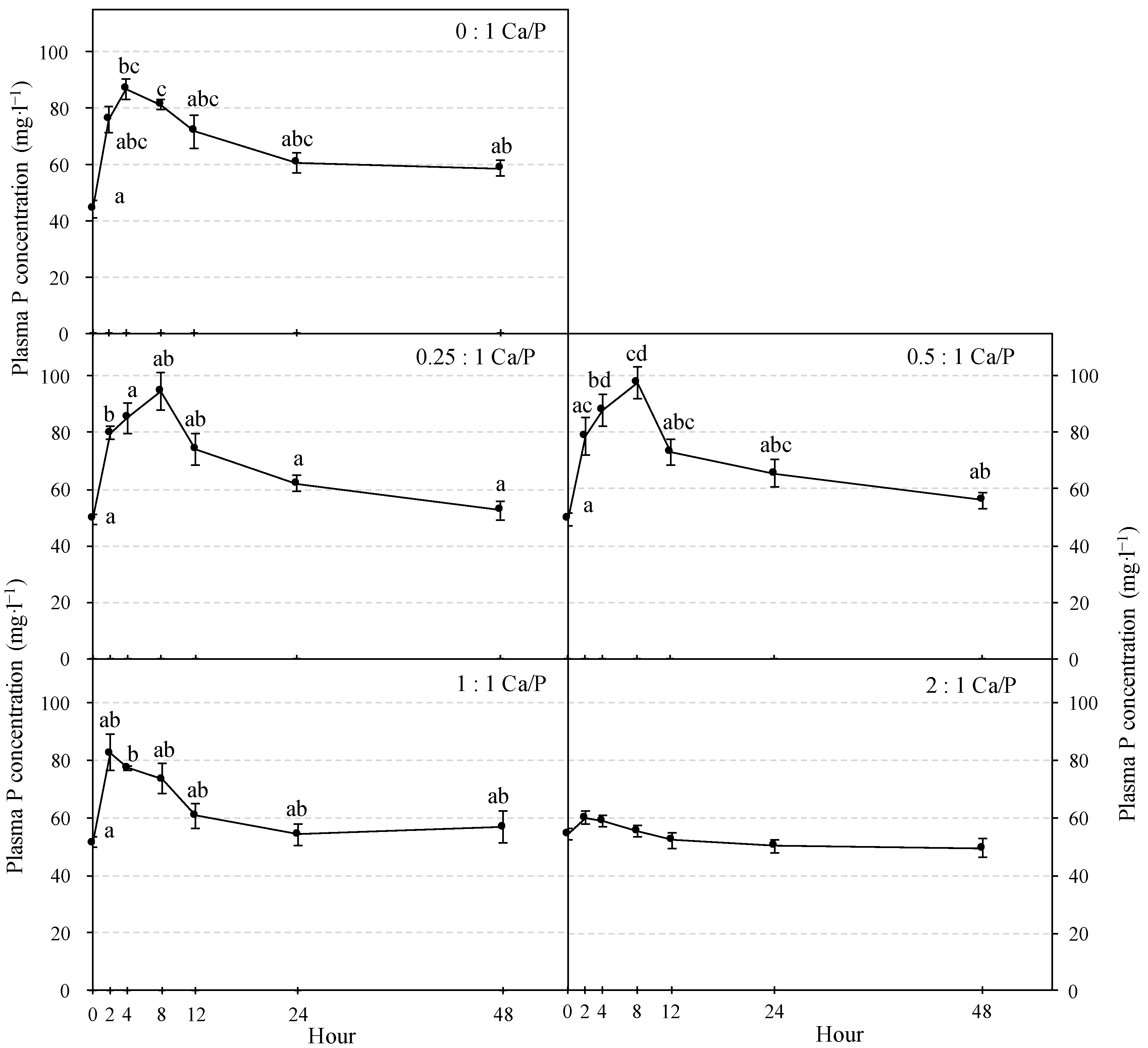
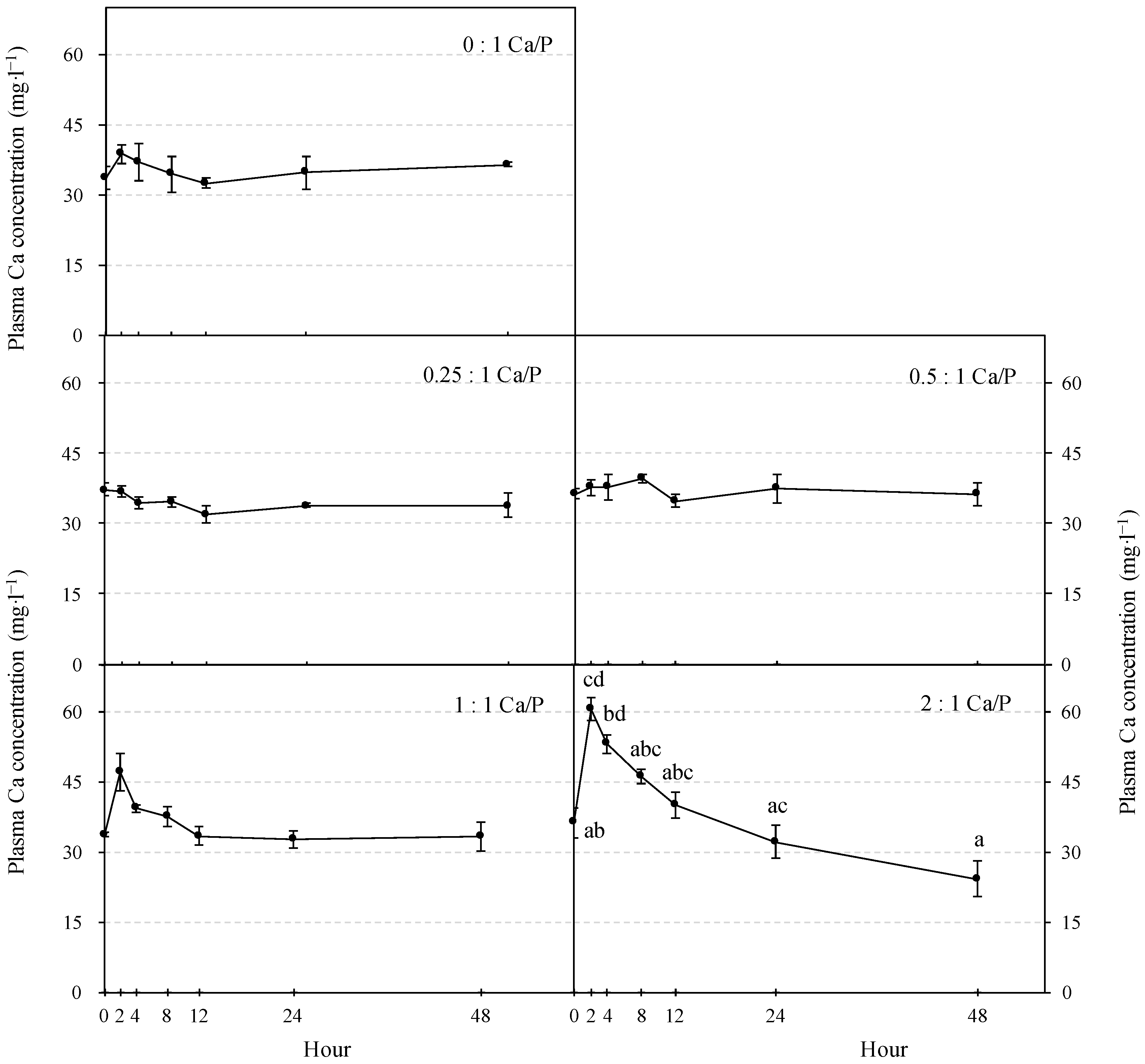
3.1. Plasma P and Ca Concentration
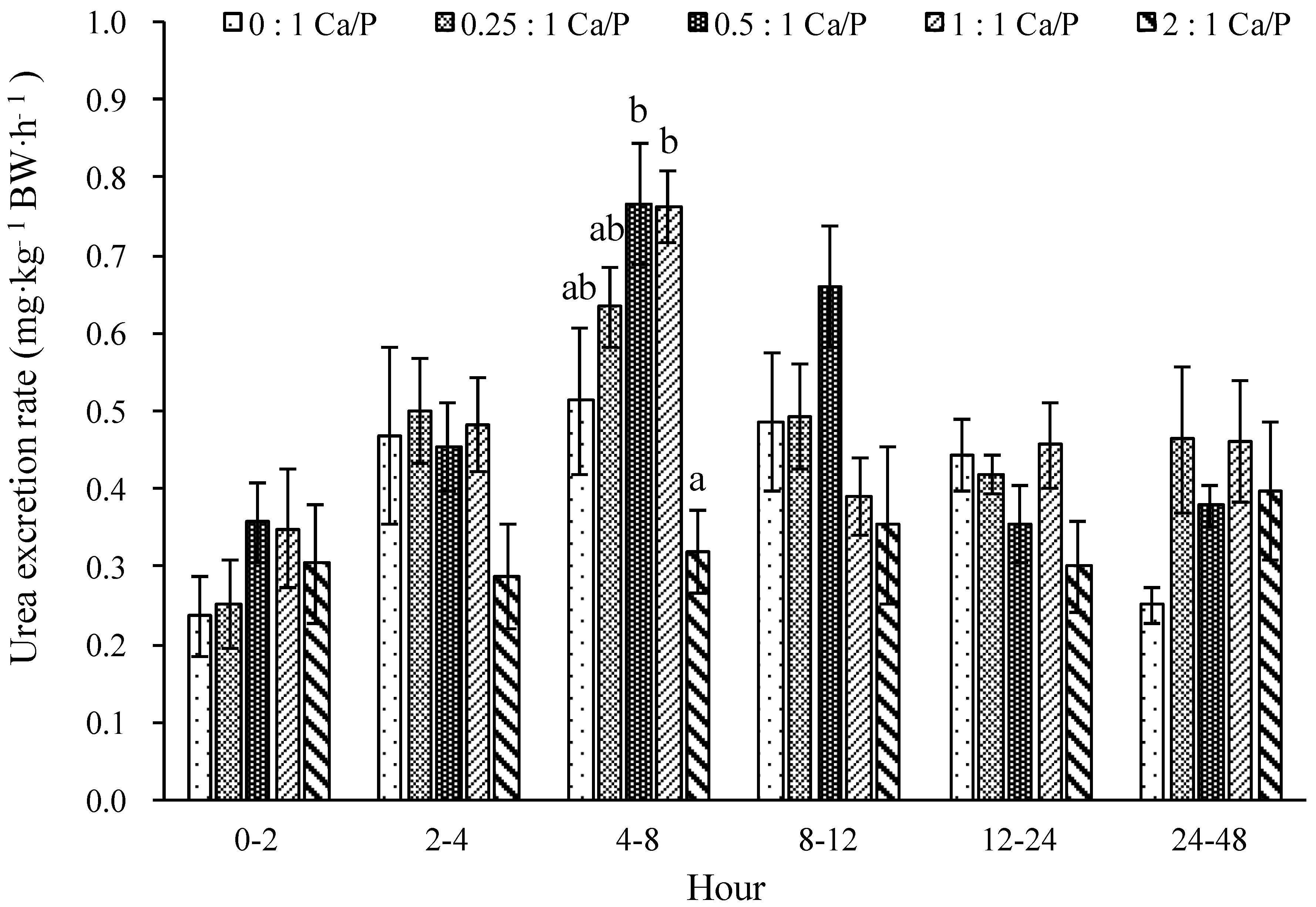
3.2. The Excretion Rate of P and Urea in Urine
3.3. Total Excretion of P, Ca, and Urea in Urine during 48 h Post Intubation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Giri, S.; Nejadhashemi, A.P.; Woznicki, S.; Zhang, Z. Analysis of best management practice effectiveness and spatiotemporal variability based on different targeting strategies. Hydrol. Process. 2014, 28, 431–445. [Google Scholar] [CrossRef]
- Liu, R.M.; Zhang, P.P.; Wang, X.J.; Chen, Y.X.; Shen, Z.Y. Assessment of effects of best management practices on agricultural non-point source pollution in Xiangxi River watershed. Agric. Water Manag. 2013, 117, 9–18. [Google Scholar] [CrossRef]
- Hashemi, F.; Olesen, J.E.; Dalgaard, T.; Børgesen, C.D. Review of scenario analyses to reduce agricultural nitrogen and phosphorus loading to the aquatic environment. Sci. Total Environ. 2016, 573, 608–626. [Google Scholar] [CrossRef]
- Sena, M.; Morris, M.R.; Seib, M.; Hicks, A. An exploration of economic valuation of phosphorus in the environment and its implications in decision making for resource recovery. Water Res. 2020, 172, 115449. [Google Scholar] [CrossRef] [PubMed]
- Prathumchai, N.; Polprasert, C.; Englande, A.J., Jr. Phosphorus leakage from fisheries sector—A case study in Thailand. Environ. Pollut. 2016, 219, 967–975. [Google Scholar] [CrossRef]
- Barcellos, D.; Queiroz, H.M.; Nóbrega, G.N.; Filho, R.L.D.O.; Santaella, S.T.; Otero, X.L.; Ferreira, T.O. Phosphorus enriched effluents increase eutrophication risks for mangrove systems in northeastern Brazil. Mar. Pollut. Bull. 2019, 142, 58–63. [Google Scholar] [CrossRef]
- Fraser, T.W.K.; Witten, P.E.; Albrektsen, S.; Breck, O.; Fontanillas, R.; Nankervis, L.; Thomsen, T.H.; Koppe, W.; Sambraus, F.; Fjelldal, P.G. Phosphorus nutrition in farmed Atlantic salmon (Salmo salar): Life stage and temperature effects on bone pathologies. Aquaculture 2019, 511, 734246. [Google Scholar] [CrossRef]
- Lall, S.P. The Minerals. In Fish Nutrition, 3rd ed.; Halver, J.E., Hardy, R.W., Eds.; Academic Press: London, UK, 2002; pp. 259–308. [Google Scholar]
- Xu, H.; Zhang, X.; Wei, Y.; Sun, B.; Jia, L.; Liang, M. Effects of dietary phosphorus level and stocking density on tiger puffer Takifugu rubripes: Growth performance, body composition, lipid metabolism, deposition of phosphorus and calcium, serum biochemical parameters, and phosphorus excretion. Aquaculture 2020, 529, 735709. [Google Scholar] [CrossRef]
- McDaniel, N.K.; Sugiura, S.H.; Kehler, T.; Fletcher, J.W.; Coloso, R.M.; Weis, P.; Ferraris, R.P. Dissolved oxygen and dietary phosphorus modulate utilization and effluent partitioning of phosphorus in rainbow trout (Oncorhynchus mykiss) aquaculture. Environ. Pollut. 2005, 138, 350–357. [Google Scholar] [CrossRef]
- Sugiura, S.H. Effects of dietary phosphorus restriction on phosphorus balance in rainbow trout Oncorhynchus mykiss. Aquacult. Sci. 2015, 63, 245–253. [Google Scholar] [CrossRef]
- Lellis, W.A.; Barrows, F.T.; Hardy, R.W. Effects of phase-feeding dietary phosphorus on survival, growth, and processing characteristics of rainbow trout Oncorhynchus mykiss. Aquaculture 2004, 242, 607–616. [Google Scholar] [CrossRef]
- Mai, K.; Zhang, C.; Ai, Q.; Duan, Q.; Xu, W.; Zhang, L.; Liufu, Z.; Tan, B. Dietary phosphorus requirement of large yellow croaker, Pseudosciaena crocea R. Aquaculture 2006, 251, 346–353. [Google Scholar] [CrossRef]
- Musharraf, M.; Khan, M.A. Dietary Phosphorus Requirement of Fingerling Indian Major Carp, Labeo rohita (Hamilton). J. World Aquac. Soc. 2019, 50, 469–484. [Google Scholar] [CrossRef]
- Zafar, N.; Khan, M.N. Determination of dietary phosphorus requirement of stinging catfish Heteropneustes fossilis based on feed conversion, growth, vertebrae phosphorus, whole body phosphorus, hematology and antioxidant status. Aquac. Nutr. 2018, 24, 1577–1586. [Google Scholar] [CrossRef]
- Hung, S.S.O. Recent advances in sturgeon nutrition. Anim. Nutr. 2017, 3, 191–204. [Google Scholar] [CrossRef]
- Jin, J.L.; Wang, C.F.; Tang, Q.; Xie, C.X.; Dai, Z.G. Dietary phosphorus affected growth performance, body composition, antioxidant status and total P discharge of young hybrid sturgeon (♀Huso huso × ♂Acipenser schrenckii) in winter months. J. Appl. Ichthyol. 2012, 28, 697–703. [Google Scholar] [CrossRef]
- Jin, J.; Chu, Z.; Chen, X.; Liang, X. Responses of hybrid sturgeon (Huso dauricus ♀ × Acipenser schrenckii ♂) to oral administration of phosphorus. Aquac. Res. 2020, 51, 1428–1436. [Google Scholar] [CrossRef]
- Xu, Q.Y.; Xu, H.; Wang, C.; Zheng, Q.; Sun, D. Studies on dietary phosphorus requirement of juvenile Siberian sturgeon Acipenser baerii. J. Appl. Ichthyol. 2011, 27, 709–714. [Google Scholar] [CrossRef]
- Chen, X.H.; Wu, J.P.; Tang, D.; Chu, Z.P.; Li, Y.; Liu, W.; Li, L.X. Analysis of nutrient components of sturgeon feeds in China. Freshw. Fish. 2018, 48, 67–76. [Google Scholar] [CrossRef]
- Hua, K.; Bureau, D.P. Modelling digestible phosphorus content of salmonid fish feeds. Aquaculture 2006, 254, 455–465. [Google Scholar] [CrossRef]
- Wang, C.; Tang, Q.; Luo, W.; Zhao, Y. Modeling phosphorus metabolism in fish species: An example for juvenile yellow catfish, Pelteobagrus fulvidraco. Aquac. Int. 2016, 24, 281–294. [Google Scholar] [CrossRef]
- NRC. Nutrient Requirements of Fish and Shrimp; Natlional Academic Press: Washington, DC, USA, 2011. [CrossRef]
- Ye, C.-X.; Liu, Y.-J.; Tian, L.-X.; Mai, K.-S.; Du, Z.-Y.; Yang, H.-J.; Niu, J. Effect of dietary calcium and phosphorus on growth, feed efficiency, mineral content and body composition of juvenile grouper, Epinephelus coioides. Aquaculture 2006, 255, 264–271. [Google Scholar] [CrossRef]
- Hassaan, M.S.; Soltan, M.A.; Agouz, H.M.; Badr, A.M. Influences of calcium/phosphorus ratio on supplemental microbial phytase efficiency for Nile tilapia (Oreochromis niloticus). Egypt. J. Aquat. Res. 2013, 39, 205–213. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, C.C.; Palacios, M.C.A.; Perez, G.M.; Ross, L.C. Phosphorus and calcium requirements in the diet of the American cichlid Cichlasoma urophthalmus (Günther). Aquac. Nutr. 2000, 6, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Deng, D.F.; Refstie, S.; Hemre, G.-I.; Crocker, C.E.; Chen, H.Y.; Cech, J.J., Jr.; Hung, S.S.O. A new technique of feeding, repeated sampling of blood and continuous collection of urine in white sturgeon. Fish Physiol. Biochem. 2000, 22, 191–197. [Google Scholar] [CrossRef]
- Tashjian, D.H.; Teh, S.J.; Sogomonyan, A.; Hung, S.S.O. Bioaccumulation and chronic toxicity of dietary L-selenomethionine in juvenile white sturgeon (Acipenser transmontanus). Aquat. Toxicol. 2006, 79, 401–409. [Google Scholar] [CrossRef] [Green Version]
- Ai, F.; Wang, L.; Li, J.; Xu, Q. Effects of a-ketoglutarate (AKG) supplementation in low phosphorous diets on the growth, phosphorus metabolism and skeletal development of juvenile mirror carp (Cyprinus carpio). Aquaculture 2019, 507, 393–401. [Google Scholar] [CrossRef]
- Morales, G.A.; Azcuy, R.L.; Casaretto, M.E.; Márquez, L.; Hernández, A.J.; Gómez, F.; Koppe, W.; Mereu, A. Effect of different inorganic phosphorus sources on growth performance, digestibility, retention efficiency and discharge of nutrients in rainbow trout (Oncorhynchus mykiss). Aquaculture 2018, 495, 568–574. [Google Scholar] [CrossRef]
- Andrews, J.W.; Murai, T.; Campbell, C. Effects of dietary calcium and phosphorous on growth, food conversion, bone ash and hematocrit levels of catfish. J. Nutr. 1973, 103, 766–771. [Google Scholar] [CrossRef] [Green Version]
- Cowey, C.B.; Sargent, J.R. Nutrition. In Fish Physiology; Hoar, W.S., Randall, D.J., Brett, J.R., Eds.; Academic Press: New York, NY, USA, 1979; Volume VIII, pp. 1–69. [Google Scholar]
- Hossain, M.A.; Yoshimatsu, T. Dietary calcium requirement in fishes. Aquac. Nutr. 2014, 20, 1–11. [Google Scholar] [CrossRef]
- Zafar, N.; Khan, M.A. Quantification of dietary calcium requirement of fingerling Heteropneustes fossilis based on growth, feed conversion efficiency, mineralization and serum alkaline phosphatase activity. J. Anim. Physiol. Anim. Nutr. 2019, 103, 1959–1968. [Google Scholar] [CrossRef] [PubMed]
- Cheng, K.-M.; Hu, C.-Q.; Liu, Y.-N.; Zheng, S.-X.; Qi, X.-J. Effects of dietary calcium, phosphorus and calcium/phosphorus ratio on the growth and tissue mineralization of Litopenaeus vannamei reared in low-salinity water. Aquaculture 2006, 251, 472–483. [Google Scholar] [CrossRef]
- Nwanna, L.C.; Oni, O.V. Determination of Optimum Calcium and Phosphorous Ratio for the Production of African Catfish Clarias gariepinus (Burchell, 1822). J. Appl. Sci. Environ. Manag. 2018, 22, 689. [Google Scholar] [CrossRef] [Green Version]
- Jahan, P.; Watanabe, T.; Satoh, S.; Kiron, V. A laboratory-based assessment of phosphorus and nitrogen loading from currently available commercial carp feeds. Fish. Sci. 2002, 68, 579–586. [Google Scholar] [CrossRef]
- Musharraf, M.; Khan, M.A. Dietary calcium requirement of fingerling Indian major carp, Labeo rohita (Hamilton) based on growth performance, tissue mineralization, whole body, and serum biochemical composition. Aquac. Int. 2020, 28, 1125–1139. [Google Scholar] [CrossRef]
- Sugiura, S.H.; Hardy, R.W.; Roberts, R.J. The pathology of phosphorus deficiency in fish—A review. J. Fish Dis. 2004, 27, 255–265. [Google Scholar] [CrossRef]
- Dalsgaard, J.; Ekmann, K.S.; Pedersen, P.B.; Verlhac, V. Effect of supplemented fungal phytase on performance and phosphorus availability by phosphorus-depleted juvenile rainbow trout (Oncorhynchus mykiss), and on the magnitude and composition of phosphorus waste output. Aquaculture 2009, 286, 105–112. [Google Scholar] [CrossRef]
- Dalsgaard, J.; Pedersen, P.B. Solid and suspended/dissolved waste (N, P, O) from rainbow trout (Oncorhynchus mykiss). Aquaculture 2011, 313, 92–99. [Google Scholar] [CrossRef]
- Sugiura, S.H.; Dong, F.M.; Hardy, R.W. Primary responses of rainbow trout to dietary phosphorus concentrations. Aquac. Nutr. 2000, 6, 235–245. [Google Scholar] [CrossRef]
- Bureau, D.P.; Cho, C.Y. Phosphorus utilization by rainbow trout (Oncorhynchus mykiss): Estimation of dissolved phosphorus waste output. Aquaculture 1999, 179, 127–140. [Google Scholar] [CrossRef]
- Adedokun, S.A.; Adeola, O. Calcium and phosphorus digestibility: Metabolic limits. J. Appl. Poult. Res. 2013, 22, 600–608. [Google Scholar] [CrossRef]
- Sabbagh, Y.; Giral, H.; Caldas, Y.; Levi, M.; Schiavi, S.C. Intestinal Phosphate Transport. Adv. Chronic Kidney Dis. 2011, 18, 85–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werner, A.; Kinne, R.K. Evolution of the Na-Pi cotransport systems. Am. J. Physiol. 2001, 280, 301–312. [Google Scholar] [CrossRef]
- Marks, J.; Srai, S.K.; Biber, J.; Murer, H.; Unwin, R.J.; Debnam, E.S. Intestinal phosphate absorption and the effect of vitamin D: A comparison of rats with mice. Exp. Physiol. 2006, 91, 531–537. [Google Scholar] [CrossRef] [PubMed]
- Proszkowiec-Weglarz, M.; Angel, R. Calcium and phosphorus metabolism in broilers: Effect of homeostatic mechanism on calcium and phosphorus digestibility. J. Appl. Poult. Res. 2013, 22, 609–627. [Google Scholar] [CrossRef]
- Amirkolaie, A.K. Reduction in the environmental impact of waste discharged by fish farms through feed and feeding. Rev. Aquac. 2011, 3, 19–26. [Google Scholar] [CrossRef]
- Damasceno, F.M. Modelling the Dynamics of Phosphorus Digestion in Rainbow Trout. Ph.D. Dissertation, The University of Guelph, Guelph, ON, Canada, 2019. [Google Scholar]
- Huber, K.; Hempel, R.; Rodehutscord, M. Adaptation of Epithelial Sodium-Dependent Phosphate Transport in Jejunum and Kidney of Hens to Variations in Dietary Phosphorus Intake. Poult. Sci. 2006, 85, 1980–1986. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, J.; Chu, Z.; Ruan, R.; Liu, W.; Chen, X.; Li, C. Phosphorus Absorption and Excretion in Hybrid Sturgeon (Huso dauricus♀ X Acipenser schrenckii♂) Intubated with Different Ca/P Ratios. Fishes 2022, 7, 138. https://doi.org/10.3390/fishes7030138
Jin J, Chu Z, Ruan R, Liu W, Chen X, Li C. Phosphorus Absorption and Excretion in Hybrid Sturgeon (Huso dauricus♀ X Acipenser schrenckii♂) Intubated with Different Ca/P Ratios. Fishes. 2022; 7(3):138. https://doi.org/10.3390/fishes7030138
Chicago/Turabian StyleJin, Jiali, Zhipeng Chu, Rui Ruan, Wei Liu, Xihua Chen, and Chuangju Li. 2022. "Phosphorus Absorption and Excretion in Hybrid Sturgeon (Huso dauricus♀ X Acipenser schrenckii♂) Intubated with Different Ca/P Ratios" Fishes 7, no. 3: 138. https://doi.org/10.3390/fishes7030138
APA StyleJin, J., Chu, Z., Ruan, R., Liu, W., Chen, X., & Li, C. (2022). Phosphorus Absorption and Excretion in Hybrid Sturgeon (Huso dauricus♀ X Acipenser schrenckii♂) Intubated with Different Ca/P Ratios. Fishes, 7(3), 138. https://doi.org/10.3390/fishes7030138