
Feeding Selectivity of Ruditapes philippinarum on Phytoplankton

Abstract
:1. Introduction
2. Materials and Methods
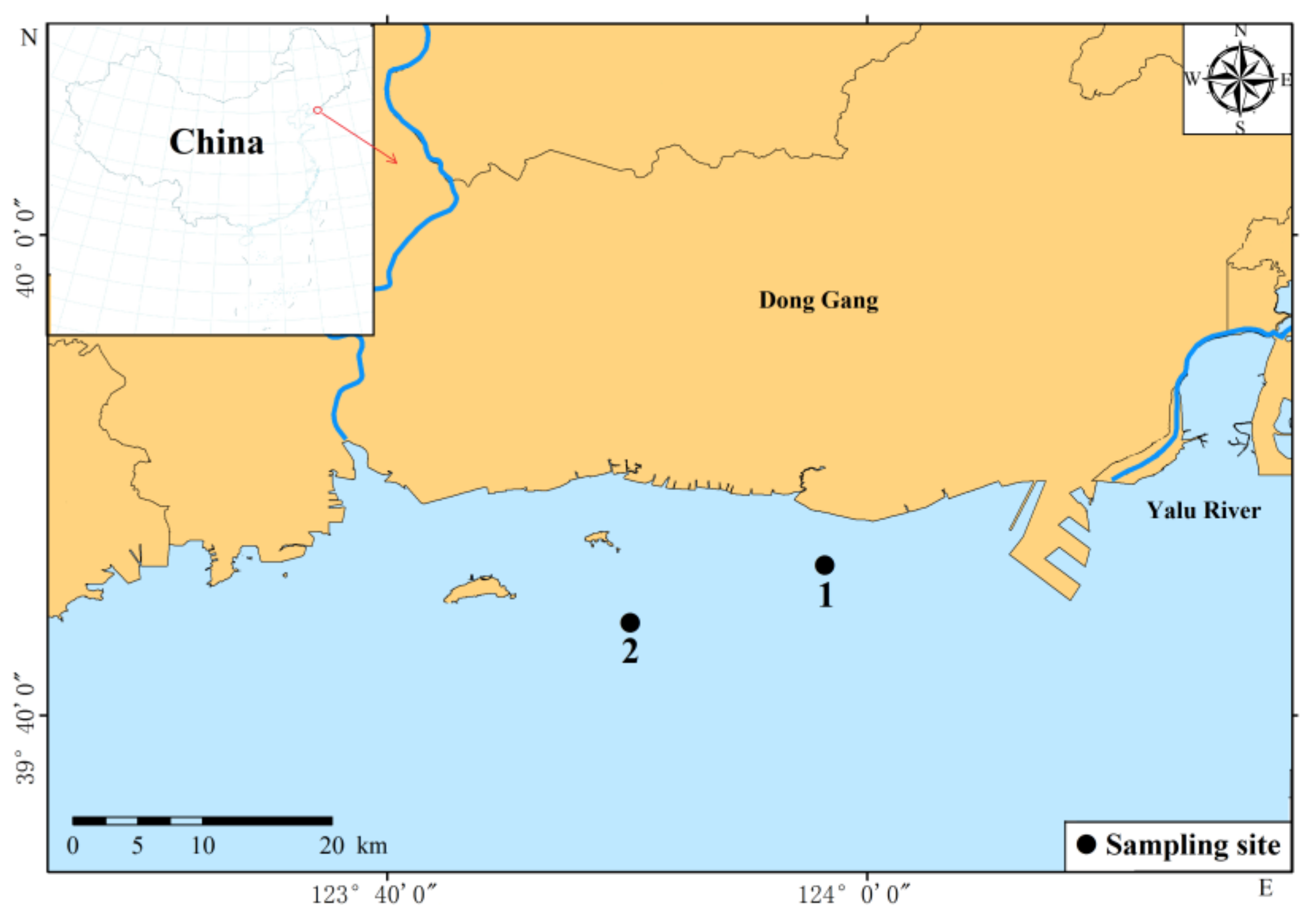
2.1. Sample Collection
2.2. Genomic DNA Extraction
2.3. PCR Amplification of the 18S rDNA V4 Variable Region
2.4. Data Analysis
2.4.1. Quality Control and Analysis of Sequencing Data
2.4.2. Dominance
2.5. Statistical Analysis
3. Results
3.1. Phytoplankton Community Structure in the Stomachs of Shellfish and Seawater
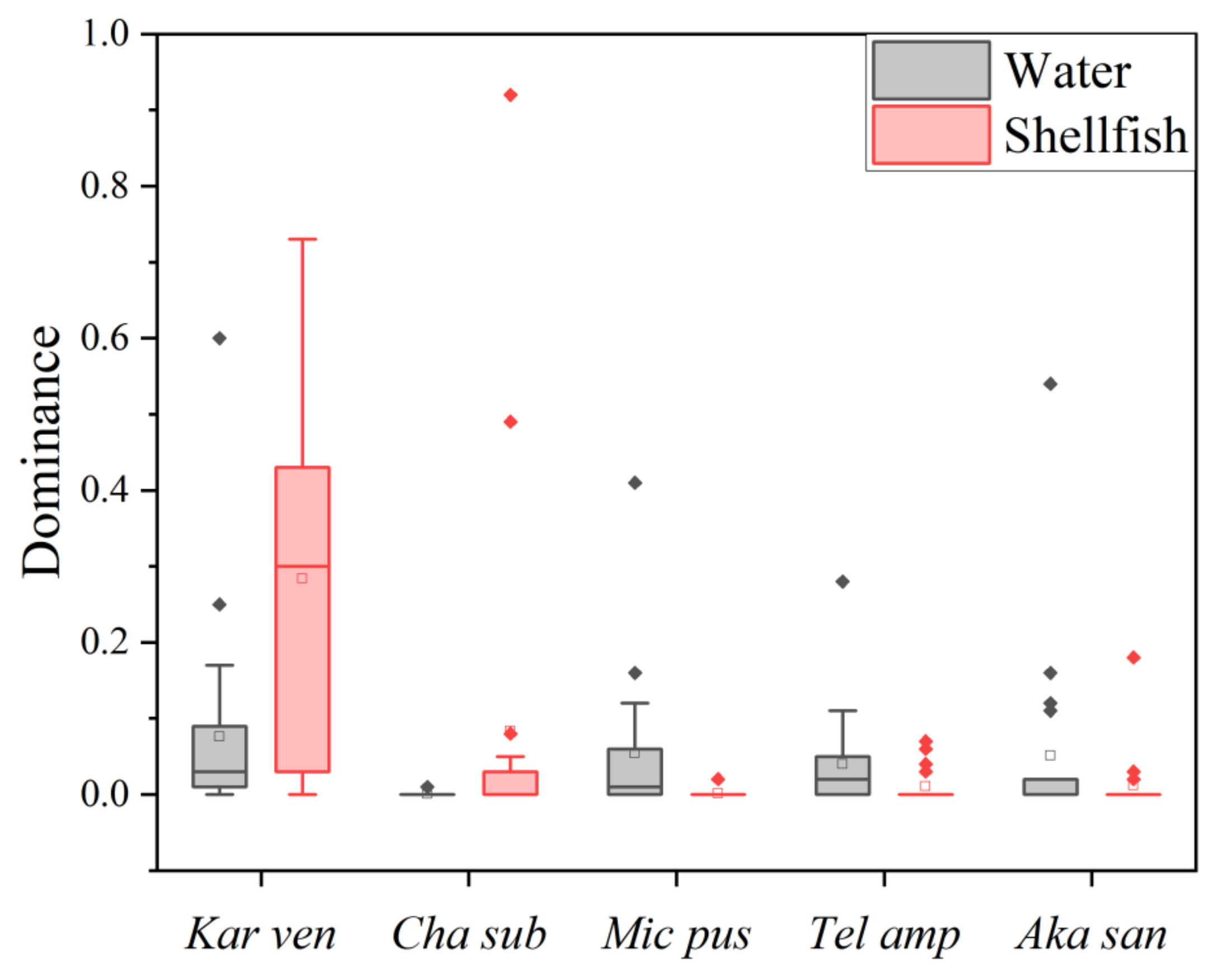
3.1.1. Dominant Species
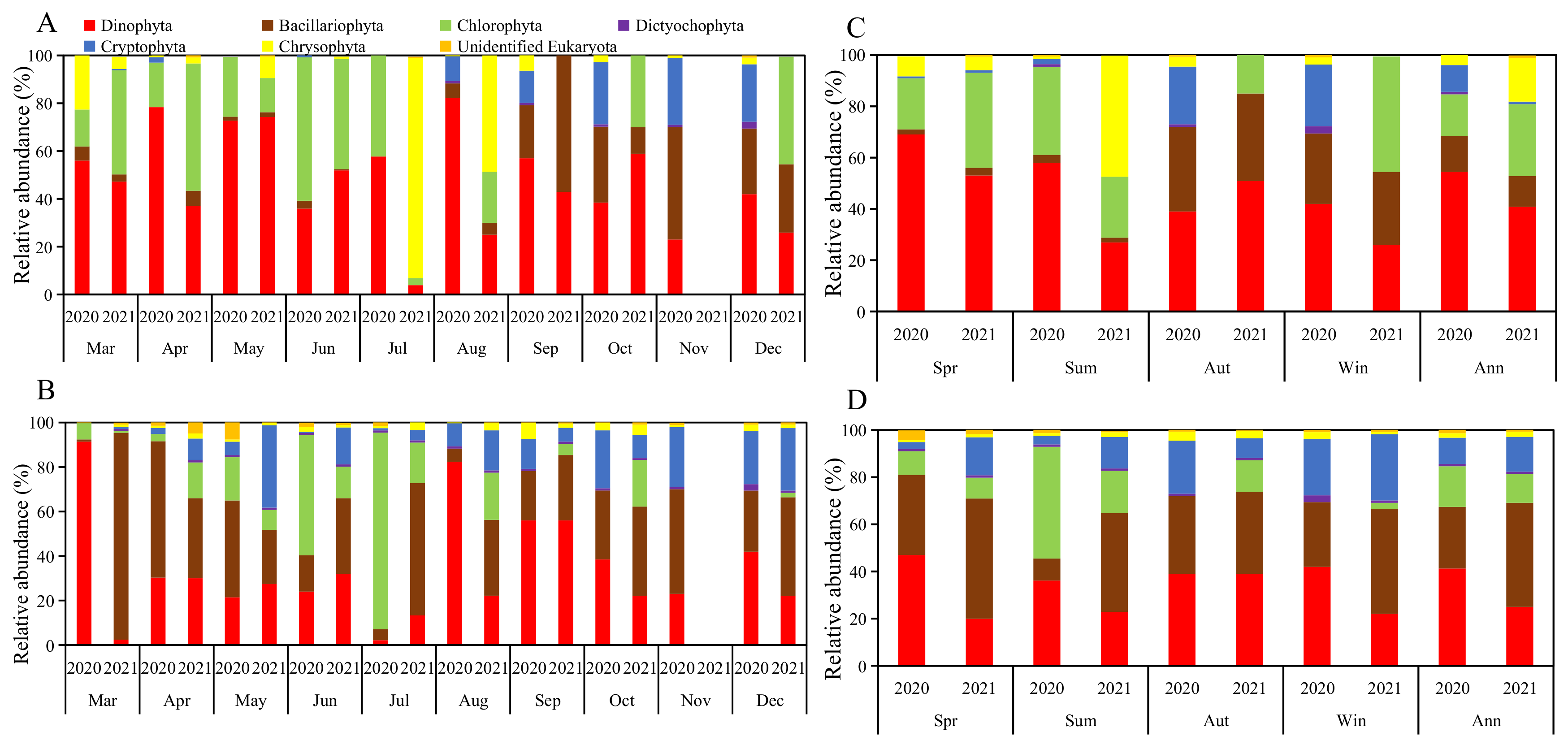
3.1.2. Phytoplankton Groups
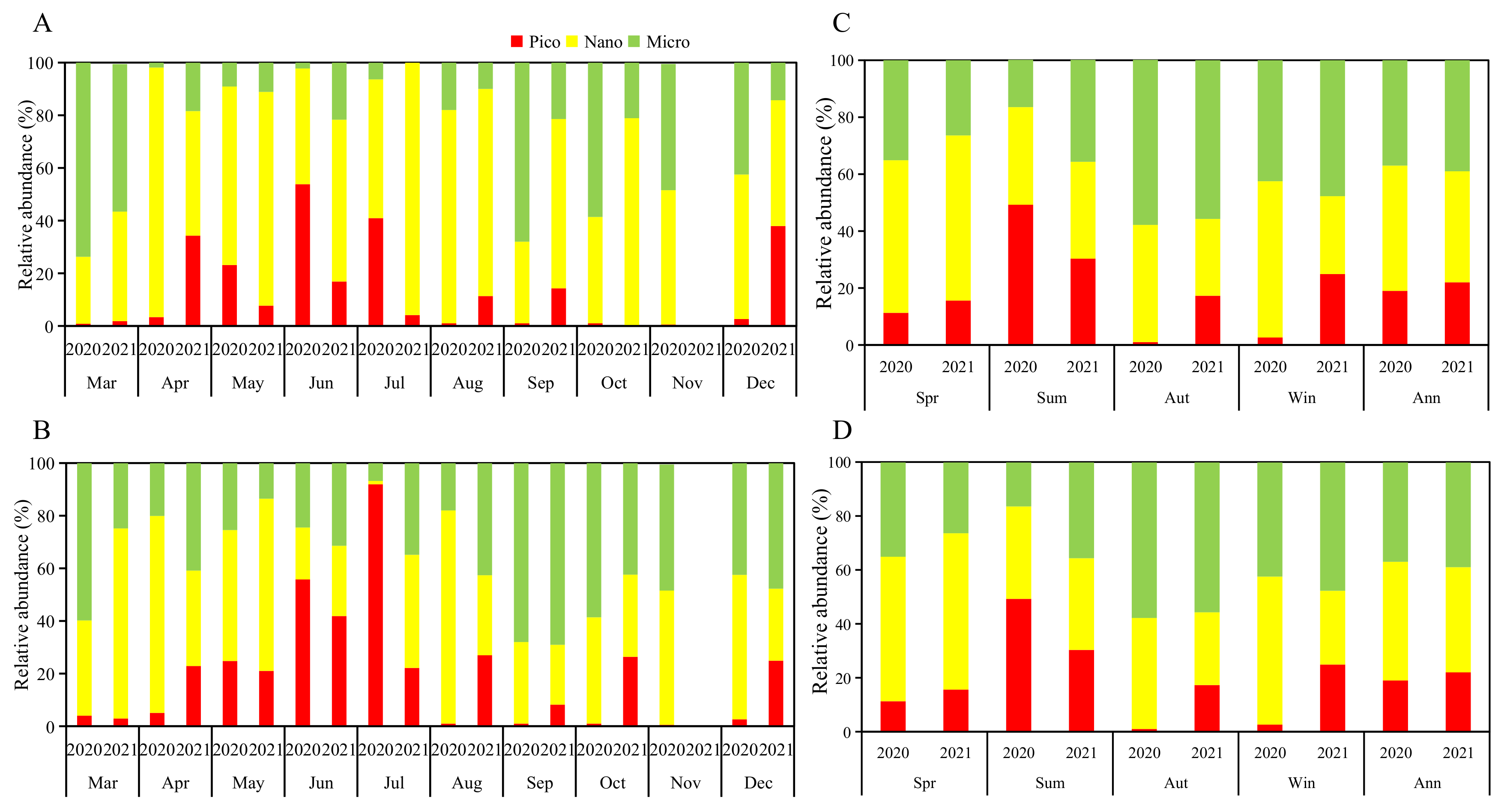
3.1.3. Phytoplankton Sizes
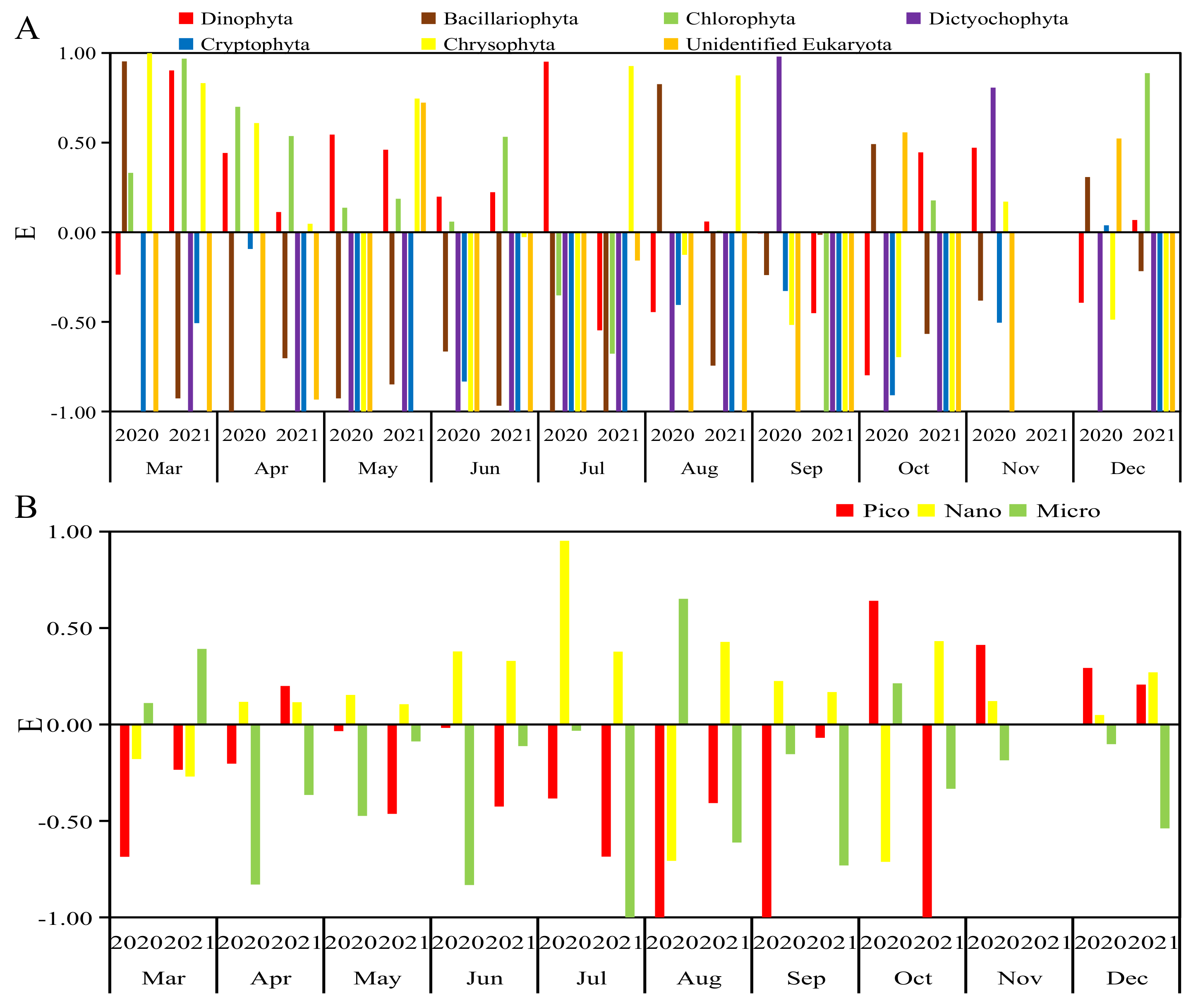
3.2. Selectivity Index (E) Value
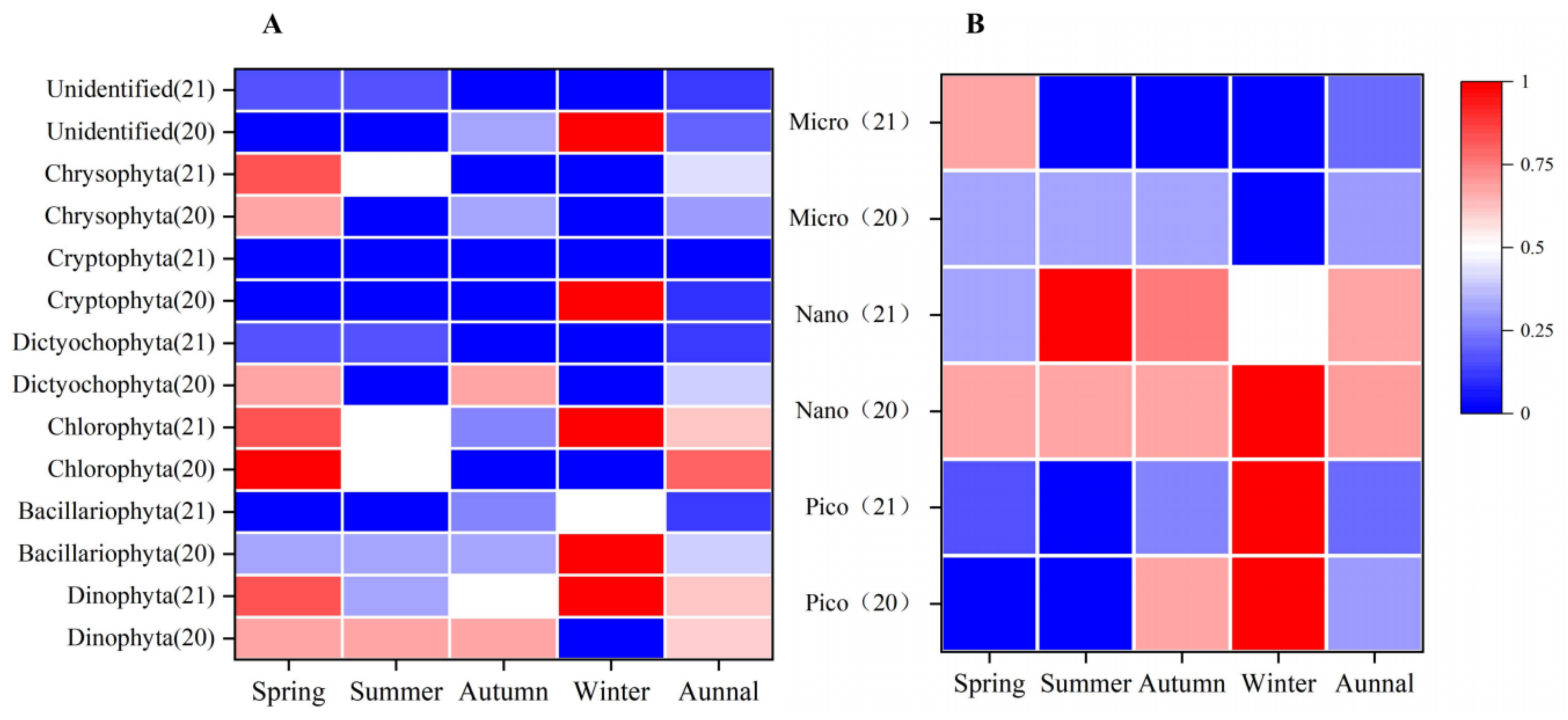
3.2.1. Selectivity for Phytoplankton Groups
3.2.2. Selectivity for Phytoplankton of Different Sizes
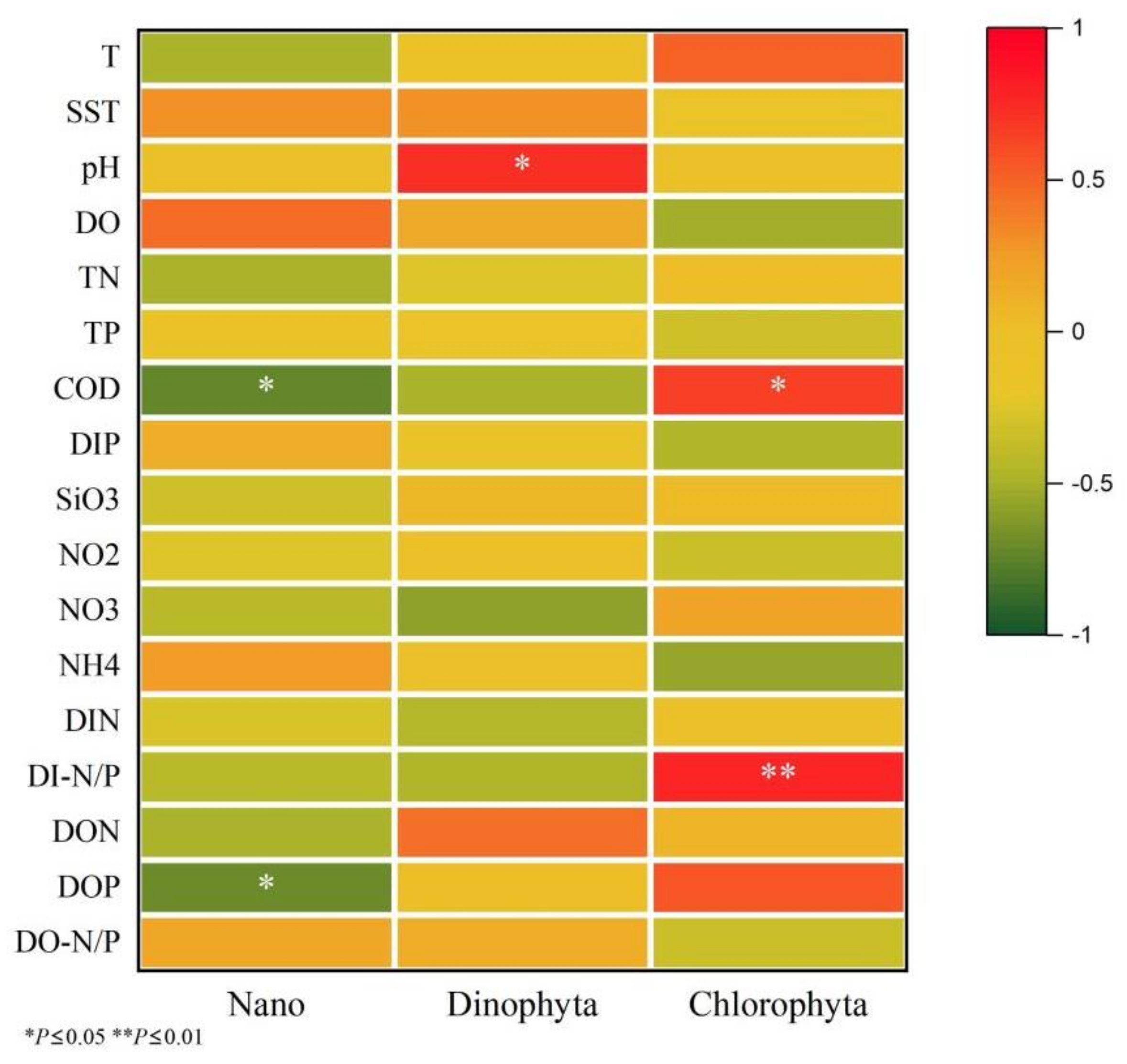
3.3. Relationship between the Environmental Factors and Feeding Selectivity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cranford, P.J.; Li, W.; Strand, Ø.; Strohmeier, T. Phytoplankton Depletion by Mussel Aquaculture: High Resolution Mapping, Ecosystem Modeling and Potential Indicators of Ecological Carrying Capacity. ICES CM Doc 2008/ H: 12. International Council for the Exploration of the Sea, Copenhagen. 2008. Available online: http://ices.dk/sites/pub/CM%20Doccuments/CM-2008/H/H1208.pdf (accessed on 26 May 2022).
- Zhang, J.H. Effect of Filter Feeding Shellfish Mariculture on the Ecosystem and the Evaluation of Ecology Carrying Capacity (Graduate School of Chinese Academy of Sciences (Institute of Oceanography), Qing Dao, Shan Dong, China). Personal Communication. 2008. Available online: https://kns.cnki.net/kcms/detail/detail.aspx?FileName=2008117213.nh&DbName=CDFD2008 (accessed on 10 June 2022).
- Shumway, S.E.; Cucci, T.L.; Newell, R.C.; Yentsch, C.M. Particle selection, ingestion, and absorption in fifilterfeeding bivalves. J. Exp. Mar. Biol. Ecol. 1985, 91, 77–92. [Google Scholar] [CrossRef]
- Ward, J.E.; Shumway, S.E. Separating the grain from the chaffff: Particle selection in suspension- and depositfeeding bivalves. J. Exp. Mar. Biol. Ecol. 2004, 300, 83–130. [Google Scholar] [CrossRef]
- Ten Winkel, E.H.; Davids, C. Food selection by Dreissena polymorpha Pallas (Mollusca: Bivalvia). Freshw. Biol. 1982, 12, 553–558. [Google Scholar] [CrossRef]
- Baker, S.M.; Levinton, J.S.; Ward, J.E. Particle transport in the zebra mussel, Dreissena polymorpha (Pallas). Biol. Bull. 2000, 199, 116–125. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.S.; Ross, A.H.; Hayden, B.J. Comparison of assimilation efficiency on diets of nine phytoplankton species of the greenshell mussel Perna canaliculus. J. Shellfish. Res. 2006, 25, 887–892. [Google Scholar]
- Safi, K.A.; Hewitt, J.E.; Talman, S.G. The effect of high inorganic seston loads on prey selection by the suspension feeding bivalve, Atrina zelandica. J. Exp. Mar. Biol. Ecol. 2007, 344, 136–148. [Google Scholar] [CrossRef]
- Safi, K.A.; Hayden, B. Differential grazing on natural planktonic populations by the mussel Perna canaliculus. Aquat. Biol. 2010, 11, 113–125. [Google Scholar] [CrossRef]
- Espinosa, E.P.; Cerrato, R.M.; Wikfors, G.H.; Allam, B. Modeling food choice in the two suspension-feeding bivalves, Crassostrea virginica and Mytilus edulis. Mar. Biol. 2016, 163, 40. [Google Scholar] [CrossRef]
- Rosa, M.; Ward, J.E.; Holohan, B.A.; Shumway, S.E.; Wikfors, G.H. Physicochemical surface properties of microalgae and their combined effects on particle selection by suspension-feeding bivalve molluscs. J. Exp. Mar. Biol. Ecol. 2017, 486, 59–68. [Google Scholar] [CrossRef]
- Song, L.; Wu, J.; Liu, W.D.; Song, Y.; Wang, N. Diversity of marine nanophytoplankton and picophytoplankton in Changxing Island offshore waters of Bohai Sea. Res. Environ. Sci. 2016, 29, 1635–1642. [Google Scholar] [CrossRef]
- Song, L.; Wu, J.; Du, J.; Li, N.; Song, G.J.; Wang, K.; Sun, M.; Wang, P. The characteristics and distribution of eukaryotic phytoplankton community in Liaodong Bay, China. Ocean. Sci. J. 2019, 54, 183–203. [Google Scholar] [CrossRef]
- Prokopowich, C.D.; Gregory, T.R.; Crease, T.J. The correlation between rDNA copy number and genome size in eukaryotes. Genome 2003, 46, 48–50. [Google Scholar] [CrossRef]
- Zhu, F.; Massana, R.; Nota, F.; Marie, D.; Vaulot, D. Mapping of picoeucaryotes in marine ecosystems with quantitative PCR of the 18S rRNA gene. FEMS Microbiol. Ecol. 2005, 52, 79–92. [Google Scholar] [CrossRef]
- Godhe, A.; Asplund, M.E.; Harnstrom, K.; Saravanan, V.; Tyagi, A.; Karunasagar, I. Quantification of diatom and dinoflagellate biomasses in coastal marine seawater samples by Real-Time PCR. Appl. Env. Microb. 2008, 74, 7174–7182. [Google Scholar] [CrossRef]
- Song, L.; Bi, X.D.; Song, G.J.; Du, J.; Wu, J.H.; Wang, Z.S.; Hu, C.K. Size-fractionated eukaryotic microalgae and its influencing factors Dachangshan Island and its adjacent waters. China Environ. Sci. 2020, 40, 2627–2634. [Google Scholar] [CrossRef]
- Liu, R.Y. Checklist of Marine Biota of China Seas, 1st ed.; Science Press: Beijing, China, 2008; pp. 301–870. [Google Scholar]
- Xu, X.; Yu, Z.M.; Cheng, F.J.; He, L.Y.; Cao, X.H.; Song, X.X. Molecular diversity and ecological characteristics of the eukaryotic phytoplankton community in the coastal waters of the Bohai sea, China. Harmful Algae 2017, 61, 13–22. [Google Scholar] [CrossRef]
- Yu, J. Establishment of Efficient Detection Technology for Plankton Diversity and Its Application in Brown Tide Research in Bohai Sea. (Ocean University of China, Qing Dao, Shan Dong, China). Personal Communication. 2014. Available online: https://kns.cnki.net/kcms/detail/detail.aspx?FileName=1014328607.nh&DbName=CMFD2015 (accessed on 20 April 2022).
- Jiang, X.J. Analysis of Picoplankton Abundance and Study of Genetic Diversity of Microeukaryotes in the in the North Yellow Sea. (Ocean University of China, Qing Dao, Shan Dong, China). Personal Communication. 2009. Available online: https://kns.cnki.net/kcms/detail/detail.aspx?FileName=2009171363.nh&DbName=CMFD2009 (accessed on 6 May 2022).
- Pan, H.Z. Study on Feeding Selectivity of Different Phytoplankton Assemblages by Several Typical Bivalve Species in the Yellow Sea and Bohai Sea. (Jinan University, Guang Zhou, Guang Dong, China). Personal Communication. 2020. Available online: https://kns.cnki.net/kcms/detail/detail.aspx?FileName=1021514063.nh&DbName=CMFD2021 (accessed on 20 April 2022).
- Lavaud, R.; Artigaud, S.; Grand, F.L.; Donval, A.; Soudant, P.; Flye-Sainte-Marie, J.; Strohmeier, T.; Strand, O.; Leynaert, A.; Beker, B.; et al. New insights into the seasonal feeding ecology of Pecten maximus using pigments, fatty acids and sterols analyses. Mare Ecol. Prog. Ser. 2018, 590, 109–129. [Google Scholar] [CrossRef]
- Bougrier, S.; Hawkins, A.J.S.; Héral, M. Preingestive selection of different microalgal mixtures in Crassostrea gigas and Mytilus edulis, analysed by flow cytometry. Aquaculture 1997, 150, 123–134. [Google Scholar] [CrossRef]
- Langdon, C.J.; Waldock, M.J. The effect of algal and artificial diets on the growth and fatty acid composition of Crassostrea gigas Spat. J. Mar. Biol. Assoc. UK 1981, 61, 431–448. [Google Scholar] [CrossRef]
- Jiang, T.; Liu, L.; Li, Y.; Zhang, J.; Tan, Z.; Wu, H.; Jiang, T.; Lu, S. Occurrence of marine algal toxins in oyster and phytoplankton samples in Daya Bay, South China Sea. Chemosphere 2017, 183, 80–88. [Google Scholar] [CrossRef]
- Bricelj, V.M.; Lee, J.H.; Cembella, A.D. Influence of dinoflagellate cell toxicity on uptake and loss of paralytic shellfish toxins in the northern quahog Mercenaria mercenaria. Mar. Ecol. Prog. 1991, 74, 33–46. [Google Scholar] [CrossRef]
- Navarro, J.M.; Labraña, W.; Chaparro, O.R.; Cisternas, B.; Ortíz, A. Physiological Constraints in Juvenile Ostrea chilensis Fed the Toxic Dinoflagellate Alexandrium catenella. Estuaries Coasts 2016, 39, 1133–1141. [Google Scholar] [CrossRef]
- Villanueva, P.A.; Navarro, J.M. Pre-ingestive selection efficiency in two populations of the razor clam Tagelus dombeii with different histories of exposure to paralytic shellfish poisoning (PSP). Mar. Freshw. Behav. Phy. 2016, 49, 291–300. [Google Scholar] [CrossRef]
- Navarro, J.M.; Leiva, G.E.; Martinez, G.; Aguilera, C. Interactive effects of diet and temperature on the scope for growth of the scallop Argopecten purpuratus during reproductive conditioning. J. Exp. Mar. Biol. Ecol. 2000, 247, 67–83. [Google Scholar] [CrossRef]
- Ding, R.Y.; Cha, D.J.; Xie, Y.X.; Xin, Q.D.; Li, X.Y.; Li, Y.; Cui, H.W.; Pan, H.Z.; Tan, Z.J.; Jiang, T. Feeding Selectivity of Phytoplankton by Four Bivalve Species in Aquaculture Area of Qinhuangdao [J/OL]. J. Yantai Univ. (Nat. Sci. Eng. Ed.) 2022, 1–11. [Google Scholar] [CrossRef]
- Sun, X.M.; Yang, F.F.; Wu, J.Y.; Li, T.; Xiang, W.Z. Evaluation of the oil-producing performance of oleaginous Picochlorum species. Fish. Mod. 2014, 41, 43–48. [Google Scholar] [CrossRef]
- Molina, F.R.; Paggi, J.C.; Devercelli, M. Zooplanktophagy in the natural diet and selectivity of the invasive mollusk Limnoperna fortunei. Biol. Invasions 2010, 12, 1647–1659. [Google Scholar] [CrossRef]
- Loret, P.; Pastoureaud, A.; Cedric, B.; Delesalle, B. Phytoplankton composition and selective feeding of the pearl oyster Pinctada margaritifera in the Takapoto lagoon (Tuamotu Archipelago, French Polynesia): In situ study using optical microscopy and HPLC pigment analysis. Mar. Ecol. Prog. Ser. 2000, 199, 55–67. [Google Scholar] [CrossRef]
- Cucci, T.L.; Shumway, S.E.; Newell, R.C.; Selvin, R.; Yentsch, C.M. Flow cytometry: A new method for characterization of differential ingestion, digestion and egestion by suspension feeders. Mar. Ecol. Prog. Ser. 1985, 24, 201–204. [Google Scholar] [CrossRef]
- Jiang, T.; Wang, L.; Zhang, F.; Fang, X.; Lu, L.; Zhang, J.; Wang, W.; Qu, K.; Chai, C. Selective feeding of bay scallop Argopecten irradians on phytoplankton community revealed by HPLC analysis of phytopigments in Bohai Sea, China. J. Oceanol. Limnol. 2019, 37, 1747–1755. [Google Scholar] [CrossRef]
- Strohmeier, T.; Strand, Ø.; Alunno-Bruscia, M.; Duinker, A.; Cranford, P.J. Variability in particle retention efficiency by the mussel Mytilus edulis. J. Exp. Mar. Biol. Ecol. 2012, 412, 96–102. [Google Scholar] [CrossRef] [Green Version]
- Rosa, M.; Ward, J.E.; Ouvrard, M.; Holohan, B.A.; Espinosa, E.P.; Shumway, S.E.; Allam, B. Examining the physiological plasticity of particle capture by the blue mussel, Mytilus edulis (L.): Confounding factors and potential artifacts with studies utilizing natural seston. J. Exp. Mar. Biol. Ecol. 2015, 473, 207–217. [Google Scholar] [CrossRef]
- Dunphy, B.J.; Hall, J.A.; Jeffs, A.G.; Wells, R.M.G. Selective particle feeding by the Chilean oyster, Ostrea chilensis; implications for nursery culture and broodstock conditioning. Aquaculture 2006, 261, 594–602. [Google Scholar] [CrossRef]
- Møhlenberg, F.; Riisgård, H.U. Efficiency of particle retention in 13 species of suspension feeding bivalves. Ophelia 1978, 17, 239–246. [Google Scholar] [CrossRef]
- Tammes, P.M.L.; Dral, A.D.G. Observations on the straining of suspensions by mussels. Arch. Meerl. Zool 1956, 11, 87–112. [Google Scholar] [CrossRef]
- Wang, F.; Dong, S.L.; Fan, R.Q.; Gao, L. The Observation of the Gills of Four Species Filtering-Feeding Bivalve Using Scanning Electron Microscope. J. Ocean. Univ. Qing Dao 1998, 2, 73–74; 76–77. [Google Scholar] [CrossRef]
- Riisgård, H.U. Efficiency of particle retention and filtration rate in 6 species of Northeast American bivalves. Mar. Ecol. Prog. Ser. 1988, 45, 217–223. [Google Scholar] [CrossRef]
- Jørgensen, C.B. Comparative studies on the function of gills in suspension feeding bivalves, with special reference to effects of serotonin. Biol. Bull. 1976, 151, 331–343. [Google Scholar] [CrossRef]
- Ward, J.E.; Sanford, L.P.; Newell, R.I.E.; MacDonald, B.A. A new explanation of particle capture in suspension-feeding bivalve molluscs. Limnol. Oceanogr. 1998, 43, 741–752. [Google Scholar] [CrossRef]
- Naddafi, R.; Pettersson, K..; Eklöv, P. The effe.ect of seasonal variation in selective feeding by zebra mussels (Dreissena polymorpha) on phytoplankton community composition. Freshw. Biol. 2007, 52, 823–842. [Google Scholar] [CrossRef]
- Dong, B.; Xue, Q.Z.; Li, J. The effect of temperature on the filtration rate, clearance rate and absorption efficiency of manila clam. Ruditapes Philippinarum. Mar. Fish. Res. 2000, 1, 37–42. Available online: https://doi.org/10.3969/j.issn.1000-7075.2000.01.007 (accessed on 3 May 2022).
- Wei, X.M.; Xu, Z.C.; He, J.J. Effects of temperature and specific gravity of seawater on the multiplication of calcareous chaetoceros. TaiWan Strait 1986, 1, 97–100. Available online: https://doi.org/CNKI:SUN:TWHX.0.1986-01-015 (accessed on 21 March 2022).
- Lora-Vilchis, M.C.; Cruz, R.V.; Reynoso-Granados, T.; Voltolina, D. Evaluation of five microalgae diets for juvenile pen shells Atrina maura. J. World Aquacult Soc. 2004, 35, 232–236. [Google Scholar] [CrossRef]
- Li, F.X. Size Fraction of Phytoplankton and Ecological Contribution of Microbial Loop in Larre-Scale Bivalve Mariculture Area. (Shanghai Ocean University, Shang Hai, China). Personal Communication. 2019. Available online: https://kns.cnki.net/kcms/detail/detail.aspx?FileName=1019667251.nh&DbName=CMFD2020 (accessed on 20 May 2022).
- Bayne, B.L.; Widdows, J.; Newell, R.I.E. Physiological measurements on estuarine bivalve molluscs in the field. Biol. Benthic Org. 1977, 57–68. [Google Scholar] [CrossRef]
- Rubenstein, D.E.; Koehl, M.A.R. The mechanisms of filter feeding: Some theoretical considerations. Am. Nat. 1977, 111, 981–994. [Google Scholar] [CrossRef]
- Labarbera, M. Particle capture by a Pacific brittle star: Experimental test of the aerosol suspension feeding model. Science 1978, 201, 1147–1149. [Google Scholar] [CrossRef]
- Jørgensen, C.B. Fluid mechanical aspects of suspension feeding. Mar. Ecol. Prog. Ser. 1983, 11, 89–103. [Google Scholar] [CrossRef]
- Silvester, N.R.; Sleigh, M.A. Hydrodynamic aspects of particle capture by Mytilus. J. Mar. Biol. Assoc. UK 1984, 64, 859–879. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test items | Detection Method | According to Standards |
|---|---|---|
| Silicate (SiO3) | Silicon content-Molybdenum blue spectrophotometric method | GB17378.4-2007/17.2 |
| Water temperature (T) | Surface water thermometer | GB17378.4-2007/25.1 |
| pH | pH meter method | GB17378.4-2007/26 |
| Salinity (SST) | CTD method | GB17378.4-2007/29.2 |
| Dissolved oxygen (DO) | Iodometry | GB17378.4-2007/31 |
| Ammonia nitrogen (NH4) | Hypobromite oxidation method | GB17378.4-2007/36.2 |
| Nitrite (NO2) | Naphthalene ethylenediamine spectrophotometry | GB17378.4-2007/37 |
| Nitrate (NO3) | Zn-Cd reduction method | GB17378.4-2007/38.2 |
| Inorganic phosphorus (DIP) | Phosphorus molybdenum blue spectrophotometry | GB17378.4-2007/39.1 |
| Total phosphorus (TP) | Potassium persulfate oxidation method | GB17378.4-2007/40 |
| Total nitrogen (TN) | Potassium persulfate oxidation method | GB17378.4-2007/41 |
| Chemical oxygen demand (COD) | Alkaline potassium permanganate method | GB17378.4-2007/32 |
| Group | Species Name (Stomach Contents) | Y | Species Name (in Seawater) | Y |
|---|---|---|---|---|
| Bacillariophyta | Guinardia striata * | 0.11 | Chaetoceros sp. NIES-3971 ** | 0.08 |
| Rhizosolenia setigera * | 0.05 | Thalassiosira nordenskioeldii * | 0.05 | |
| Bacillariophyta sp. GSL077 ** | 0.04 | Rhizosolenia setigera ** | 0.04 | |
| Skeletonema menzellii ** | 0.04 | |||
| Brockmanniella brockmannii * | 0.03 | |||
| Thalassiosira curviseriata * | 0.03 | |||
| Chaetoceros calcitrans ** | 0.03 | |||
| Thalassiosira pseudonana ** | 0.03 | |||
| Cyclotella choctawhatcheeana ** | 0.03 | |||
| Thalassiosira concaviuscula * | 0.02 | |||
| Arcocellulus cornucervis ** | 0.02 | |||
| Dinophyta | Karlodinium veneficum | 0.21 */0.34 ** | Karlodinium veneficum | 0.12 */0.03 ** |
| Polykrikos kofoidii * | 0.07 | Akashiwo sanguinea ** | 0.09 | |
| Ansanella granifera * | 0.05 | Dissodinium pseudolunula * | 0.06 | |
| Islandinium minutum ** | 0.03 | Spiniferites belerius * | 0.05 | |
| Akashiwo sanguinea * | 0.02 | Ansanella granifera * | 0.04 | |
| Alexandrium affine * | 0.02 | Alexandrium hiranoi ** | 0.03 | |
| Proterythropsis sp. * | 0.02 | Gyrodinium dominans * | 0.02 | |
| Alexandrium leei * | 0.02 | Alexandrium affine * | 0.02 | |
| Chlorophyta | Picochlorum sp. | 0.09 */0.04 ** | Ostreococcus tauri * | 0.1 |
| Pterosperma cristatum ** | 0.08 | Micromonas pusilla | 0.05 */0.06 ** | |
| Tetradesmus obliquus ** | 0.03 | Mamiella gilva ** | 0.05 | |
| Chlamydomonas raudensis ** | 0.03 | |||
| Cryptophyta | Cryptomonadales sp.* | 0.02 | Cryptomonadales sp.* | 0.07 */0.03 ** |
| Teleaulax amphioxeia* | 0.02 | Teleaulax amphioxeia ** | 0.06 | |
| Cryptomonadales environmental sample ** | 0.03 | |||
| Chrysophyta | Chattonella subsalsa** | 0.18 | Fibrocapsa japonica * | 0.01# |
| Chrysosphaerella sp.* | 0.02 | |||
| Dictyochophyta | Phaeocystis globosa* | 0.02 | Phaeocystis globose * | 0.003 # |
| Size | Species Name (Stomach Contents) | Y | Species Name (in Seawater) | Y |
|---|---|---|---|---|
| picophytoplankton | Picochlorum sp. | 0.09 */0.04 ** | Ostreococcus tauri * | 0.1 |
| Bacillariophyta sp. GSL077 ** | 0.05 | Micromonas pusilla | 0.05 */0.06 ** | |
| Mamiella gilva ** | 0.05 | |||
| Chaetoceros calcitrans ** | 0.03 | |||
| Cryptomonadales environmental sample** | 0.03 | |||
| Cryptomonadales sp. ** | 0.03 | |||
| nanophytoplankton | Karlodinium veneficum | 0.21 */0.36 ** | Karlodinium veneficum | 0.12 */0.03 ** |
| Chattonella subsalsa ** | 0.18 | Chaetoceros sp. NIES-3971 ** | 0.08 | |
| Ansanella granifera * | 0.05 | Cryptomonadales sp. * | 0.07 | |
| Chlamydomonas raudensis ** | 0.03 | Teleaulax amphioxeia | 0.02 */0.06 ** | |
| Proterythropsis sp. * | 0.02 | Thalassiosira nordenskioeldii * | 0.05 | |
| Teleaulax amphioxeia* | 0.02 | Ansanella granifera * | 0.04 | |
| Phaeocystis globose * | 0.02 | Skeletonema menzellii ** | 0.04 | |
| Cryptomonadales sp. * | 0.02 | Brockmanniella brockmannii * | 0.03 | |
| Chrysosphaerella sp. * | 0.02 | Cyclotella choctawhatcheeana ** | 0.03 | |
| Telonema subtile * | 0.02 | |||
| Arcocellulus cornucervis ** | 0.02 | |||
| microphytoplankton | Guinardia striata * | 0.11 | Akashiwo sanguinea | 0.02 */0.09 ** |
| Pterosperma cristatum ** | 0.08 | Dissodinium pseudolunula * | 0.06 | |
| Polykrikos kofoidii * | 0.07 | Spiniferites belerius * | 0.05 | |
| Rhizosolenia setigera * | 0.05 | Rhizosolenia setigera | 0.04 | |
| Tetradesmus obliquus ** | 0.02 */0.03 ** | Thalassiosira curviseriata * | 0.03 | |
| Islandinium minutum ** | 0.03 | Thalassiosira pseudonana ** | 0.03 | |
| Akashiwo sanguinea * | 0.02 | Alexandrium hiranoi ** | 0.03 | |
| Alexandrium affine * | 0.02 | Gyrodinium dominans * | 0.02 | |
| Alexandrium leei * | 0.02 | Thalassiosira concaviuscula * | 0.02 | |
| Navicula sp. 39 ** | 0.02 | Thalassiosira nodulolineata * | 0.02 | |
| Alexandrium affine * | 0.02 |
| Environmental Factors | Spring | Summer | Autumn | Winter |
|---|---|---|---|---|
| T | 8.63 | 24.27 | 19.96 | 4.57 |
| SST | 27.56 | 26.21 | 24.84 | 26.56 |
| pH | 7.99 | 8.01 | 7.93 | 7.86 |
| DO | 10.94 | 8.01 | 8.90 | 10.85 |
| TN | 0.300 | 0.440 | 0.573 | 0.570 |
| TP | 0.009 | 0.013 | 0.027 | 0.025 |
| COD | 1.20 | 1.70 | 1.63 | 1.15 |
| DIP | 0.003 | 0.005 | 0.020 | 0.019 |
| SiO3 | 0.178 | 0.930 | 1.089 | 0.868 |
| NO2 | 0.007 | 0.009 | 0.037 | 0.019 |
| NO3 | 0.152 | 0.240 | 0.304 | 0.320 |
| NH4 | 0.039 | 0.036 | 0.082 | 0.108 |
| DIN | 0.198 | 0.286 | 0.422 | 0.446 |
| DI-N/P | 245.49 | 351.74 | 76.31 | 51.08 |
| DON | 0.101 | 0.154 | 0.151 | 0.124 |
| DOP | 0.006 | 0.009 | 0.006 | 0.006 |
| DO-N/P | 38.56 | 51.78 | 64.72 | 48.32 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Song, L.; Song, G.; Wu, J.; Wang, K.; Wang, Z.; Liu, S. Feeding Selectivity of Ruditapes philippinarum on Phytoplankton. Fishes 2022, 7, 222. https://doi.org/10.3390/fishes7050222
Liu Y, Song L, Song G, Wu J, Wang K, Wang Z, Liu S. Feeding Selectivity of Ruditapes philippinarum on Phytoplankton. Fishes. 2022; 7(5):222. https://doi.org/10.3390/fishes7050222
Chicago/Turabian StyleLiu, Yin, Lun Song, Guangjun Song, Jinhao Wu, Kun Wang, Zhaohui Wang, and Suxuan Liu. 2022. "Feeding Selectivity of Ruditapes philippinarum on Phytoplankton" Fishes 7, no. 5: 222. https://doi.org/10.3390/fishes7050222
APA StyleLiu, Y., Song, L., Song, G., Wu, J., Wang, K., Wang, Z., & Liu, S. (2022). Feeding Selectivity of Ruditapes philippinarum on Phytoplankton. Fishes, 7(5), 222. https://doi.org/10.3390/fishes7050222