
Effect of Astragalus membranaceus on Transcriptome and Survival of Hybrid Yellow Catfish (Pseudobagrus vachellii ♂ × Tachysurus fulvidraco ♀) in Response to Aeromonas hydrophila Challenge


Abstract
:1. Introduction
2. Materials and Methods
2.1. A. membranaceus Extract and Feeds Preparation
2.2. Fish
2.3. Relative Percent Survival (RPS)
2.4. RNA Extraction and Library Preparation
2.5. RNA Sequencingand Function Annotation
2.6. DEGs Identification
2.7. RNA Sequencing and Transcriptome Assembling
3. Result and Discussion
3.1. Transcriptome and Unigene Annotation
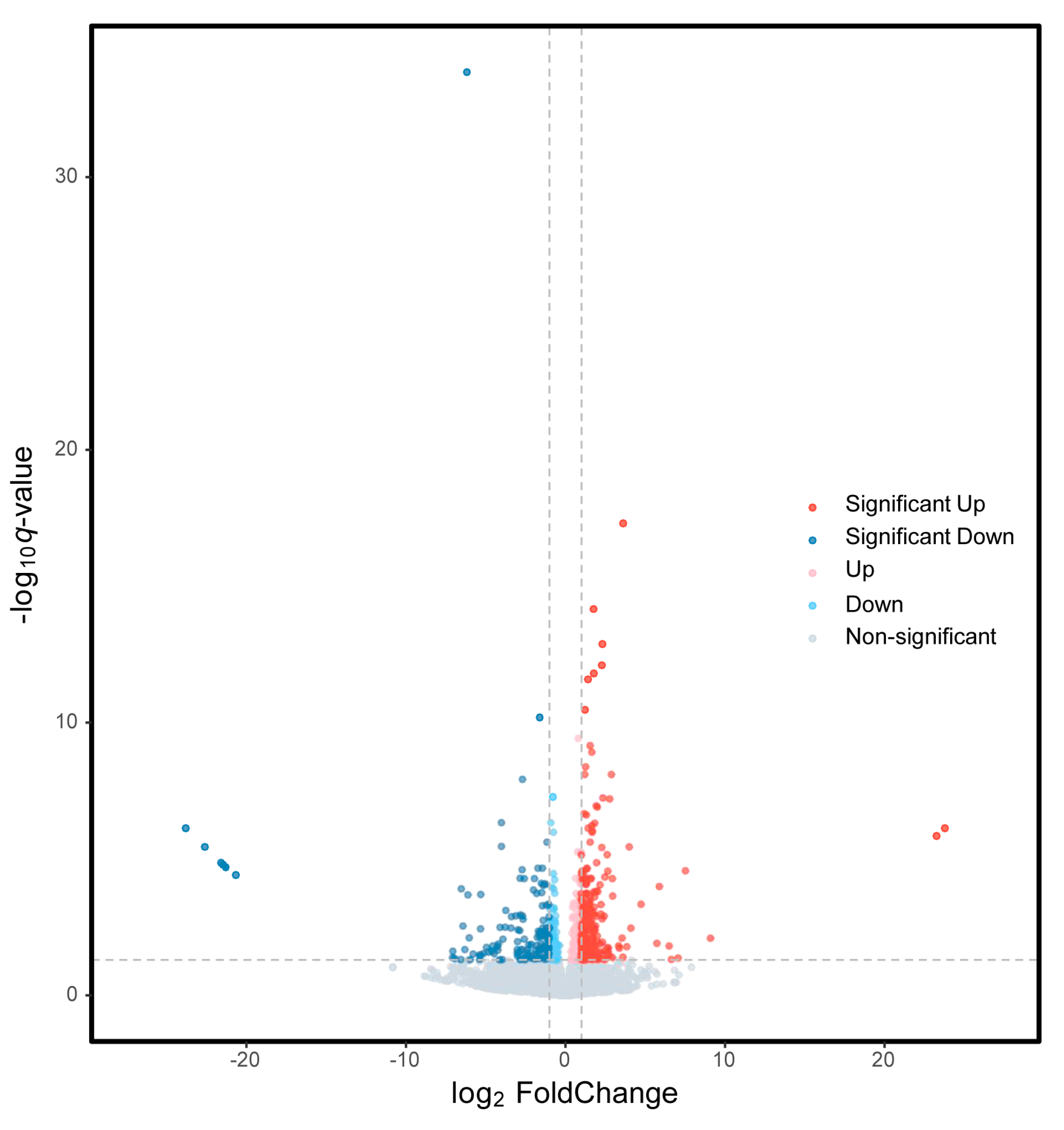
3.2. DEGs Analysis
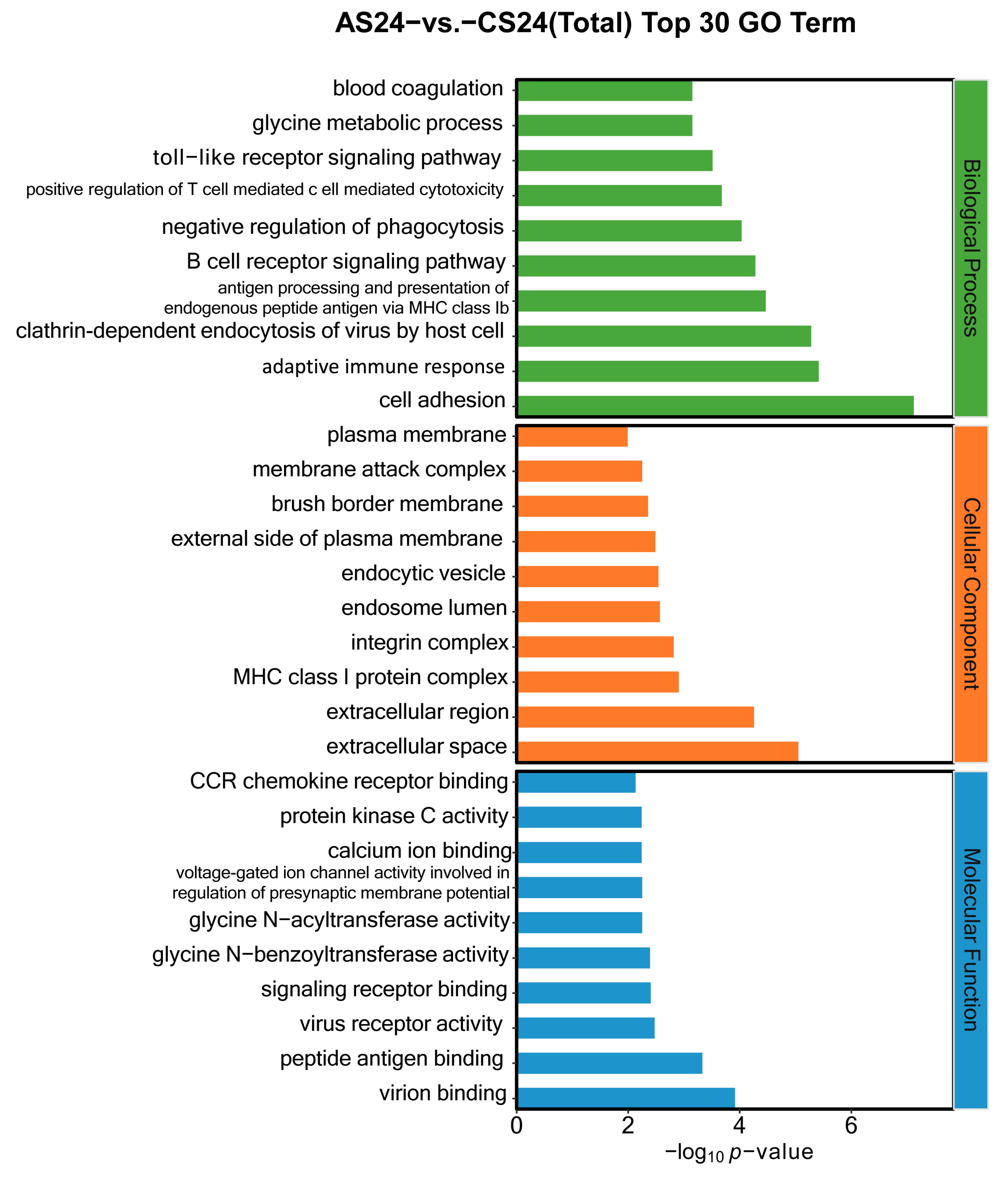
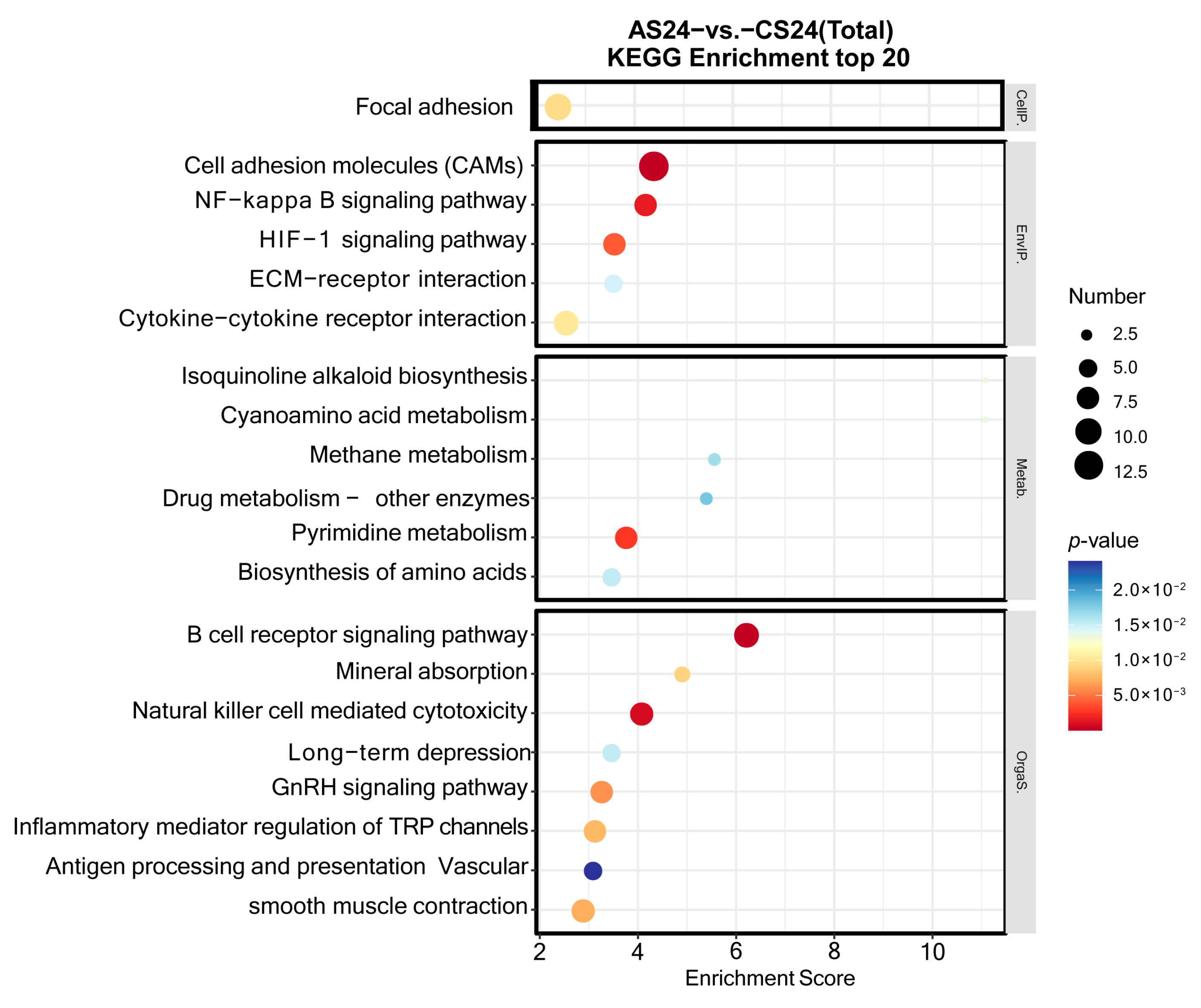
3.3. Functional Annotation
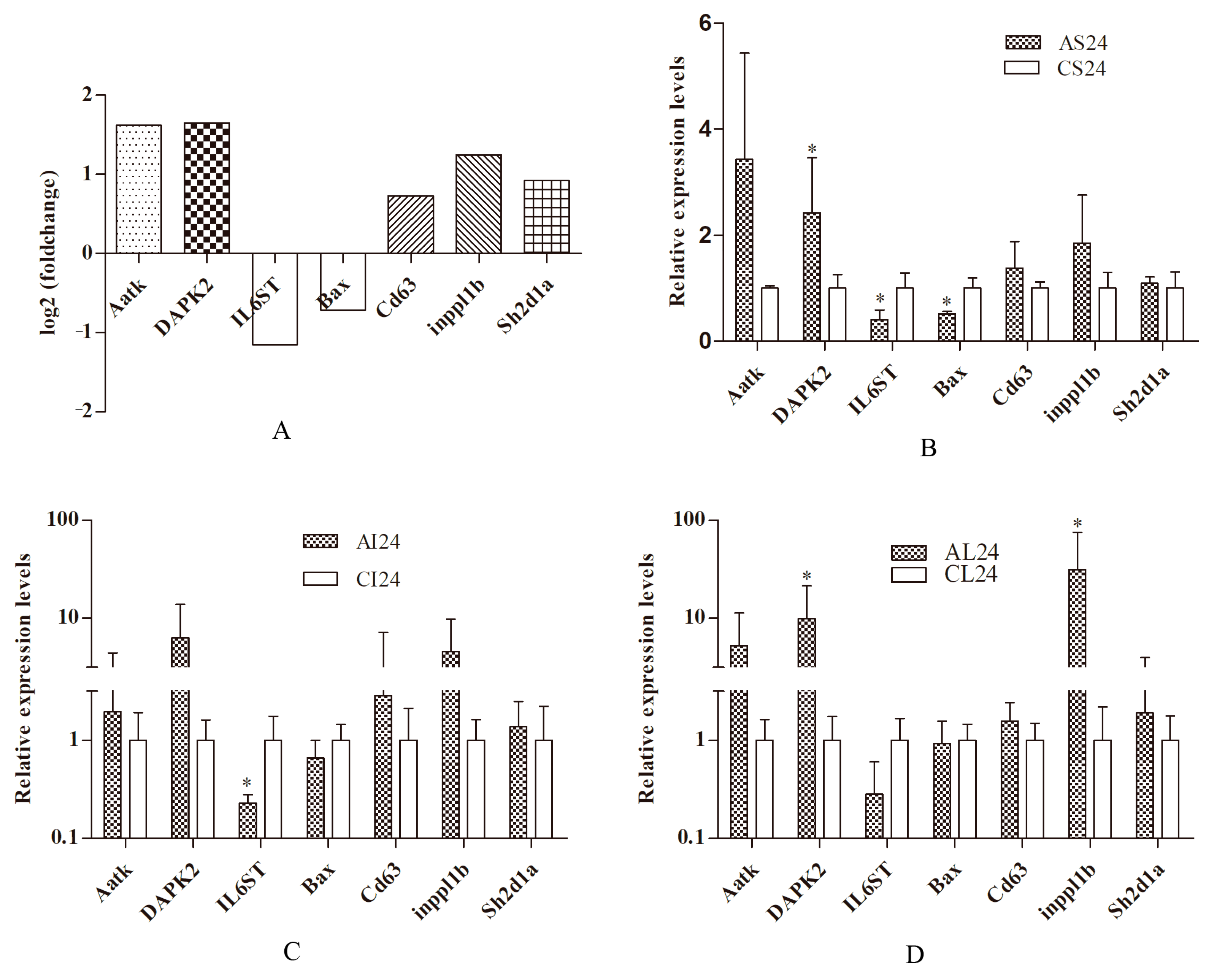
3.4. qRTPCR Analysis
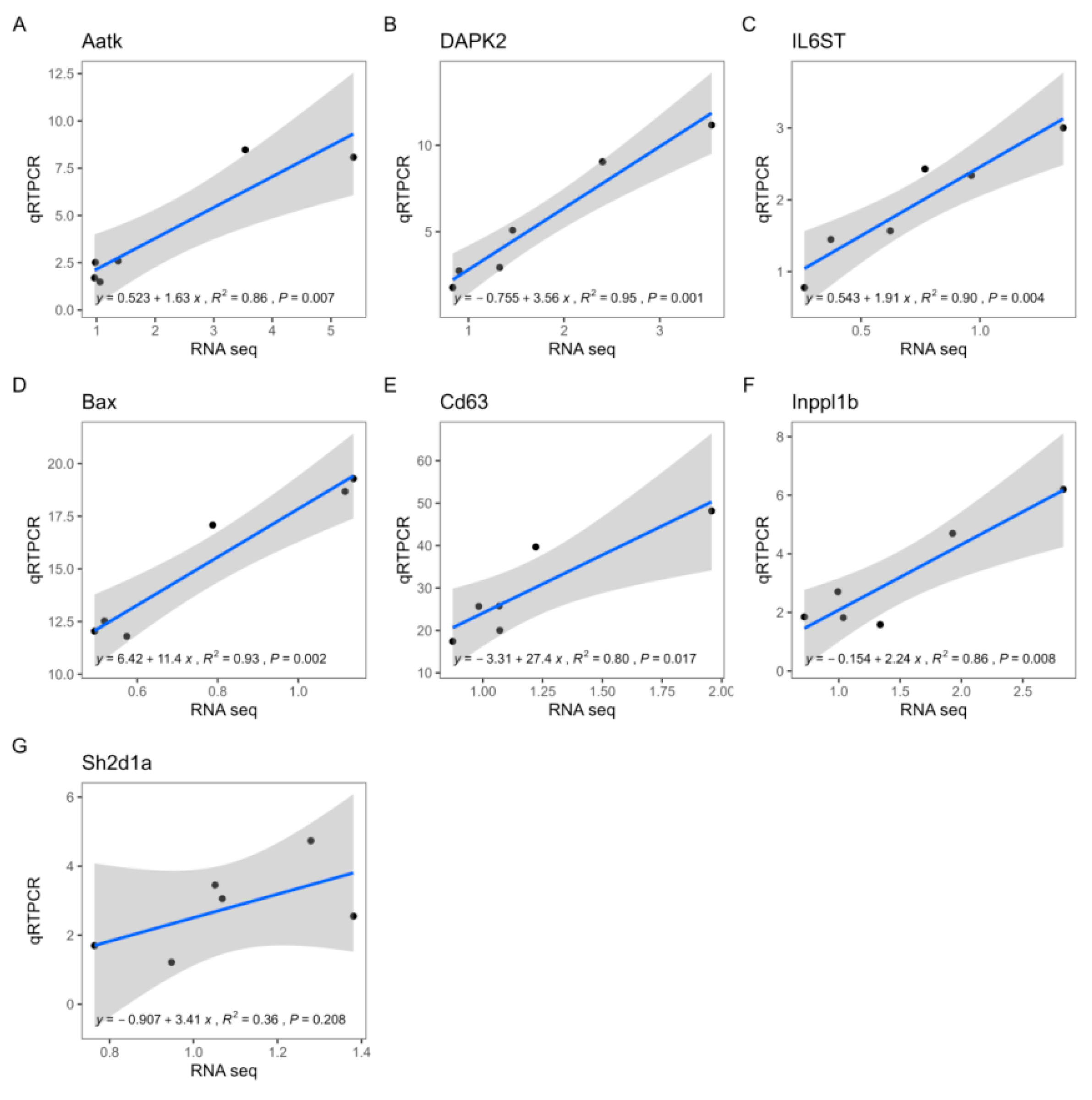
3.5. Correlation between qRTPCR and RNA-Seq
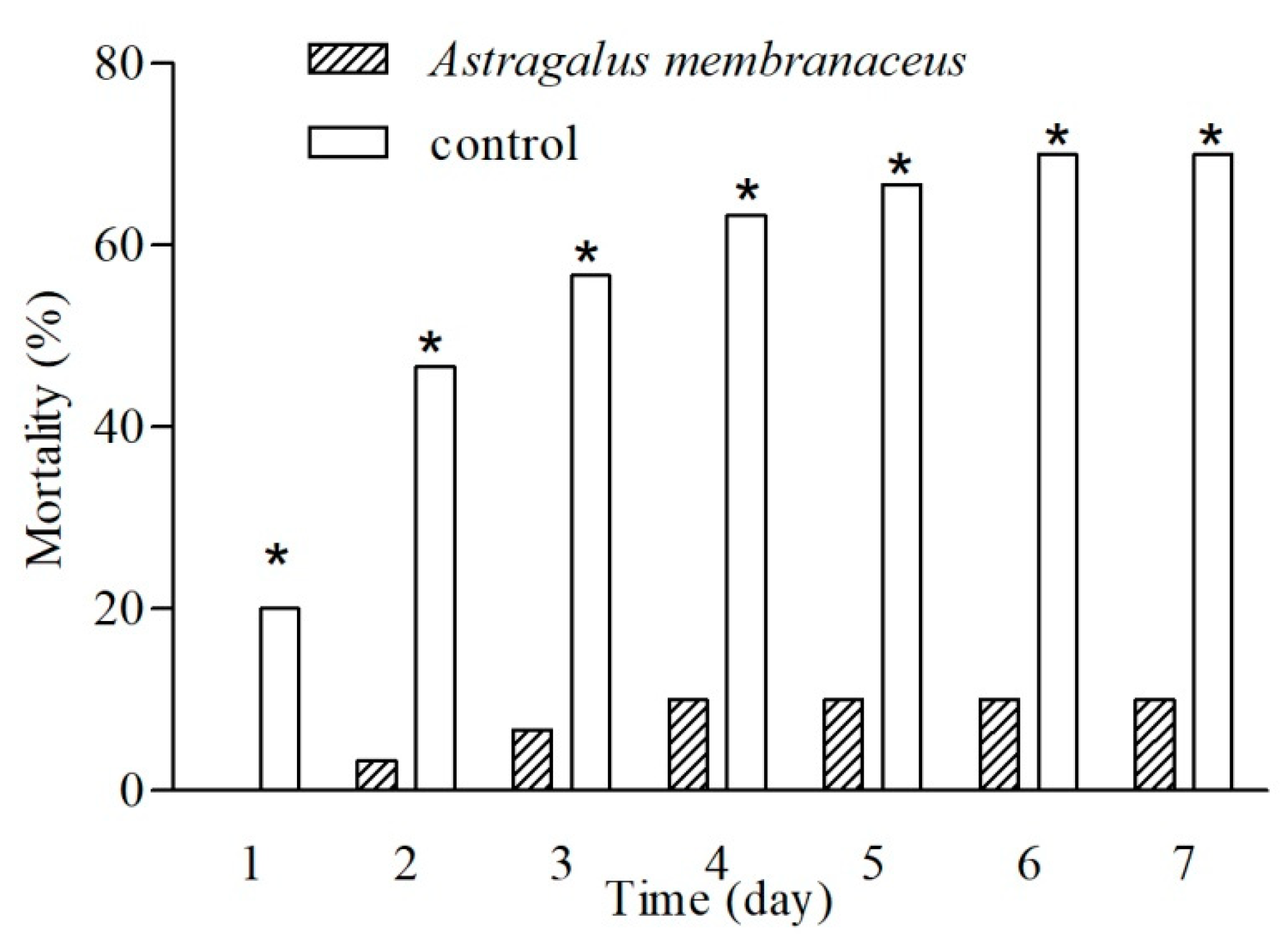
3.6. Disease Resistance
4. Conclusions and Prospects
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Shao, W.; Fan, Q.; Zhang, C.; Wang, L. Comparative analysis of nutritive composition in muscle of “huangyou no. 1”, yellow catfish and darkbarbel catfish. J. Huazhong Agric. Univ. 2018, 37, 76–82. [Google Scholar]
- Pan, T.; Yan, M. The screening of traditional Chinese herbs on nonspecific immune response and protection of Pacific white shrimp (Litopenaeus vannamei) from Vibrio harveyi infection. Aquac. Int. 2020, 28, 767–776. [Google Scholar] [CrossRef]
- Barman, D.; Nen, P.; Mandal, S.C.; Kumar, V. Immunostimulants for aquaculture health management. J. Mar. Sci. Res. Dev. 2013, 3, 1000134. [Google Scholar] [CrossRef]
- Labh, S.N.; Shakya, S.R. Application of Immunostimulants as an Alternative to Vaccines for Health Management in Aquaculture; AkiNik Publications: New Delhi, India, 2014. [Google Scholar]
- Sakai, M. Current research status of fish immunostimulants. Aquaculture 1999, 172, 63–92. [Google Scholar] [CrossRef]
- Ardó, L.; Yin, G.; Xu, P.; Váradi, L.; Szigeti, G.; Jeney, Z.; Jeney, G. Chinese herbs (Astragalus membranaceus and Lonicera japonica) and boron enhance the non-specific immune response of Nile tilapia (Oreochromis niloticus) and resistance against Aeromonas hydrophila. Aquaculture 2008, 275, 26–33. [Google Scholar] [CrossRef]
- Wang, C.; Liu, H.; Mu, G.; Lu, S.; Wang, D.; Jiang, H.; Sun, X.; Han, S.; Liu, Y. Effects of traditional Chinese medicines on immunity and culturable gut microflora to Oncorhynchus masou. Fish Shellfish Immunol. 2019, 93, 322–327. [Google Scholar] [CrossRef]
- Xia, Y.-T.; Cheng, E.H.-C.; Xia, Y.-J.; Wu, Q.-Y.; Zhang, L.H.-L.; Lin, S.-Y.; Dong, T.T.-X.; Qin, Q.-W.; Wang, W.-X.; Tsim, K.W.-K. Characterization of a macrophagic-like cell line derived from rabbit fish (Siganus fuscescens): An illustration of anti-inflammatory responses of the herbal extract of Scutellaria baicalensis. Fish Shellfish Immunol. Rep. 2021, 2, 100036. [Google Scholar] [CrossRef] [PubMed]
- Pan, T.; Yan, M.; Chen, S.; Wang, X. Effects of Ten Traditional Chinese Herbs on Immune Response and Disease Resistance of Sciaenops Ocellatus (Actinopterygii: Perciformes: Sciaenidae). Acta Ichthyol. Piscat. 2013, 43, 41–49. [Google Scholar] [CrossRef]
- Adel, M.; Yeganeh, S.; Dawood, M.A.O.; Safari, R.; Radhakrishnan, S. Effects of pediococcus pentosaceus supplementation on growth performance, intestinal microflora and disease resistance of white shrimp, litopenaeus vannamei. Aquac. Nutr. 2017, 23, 1401–1409. [Google Scholar] [CrossRef]
- Jeney, G.; Jeney, Z. Application of immunostimulants for modulation of the non-specific defense mechanisms in sturgeon hybrid: Acipenser ruthenus x a. Baerii. J. Appl. Ichthyol. 2002, 18, 416–419. [Google Scholar] [CrossRef]
- Zeng, P.; Li, J.; Chen, Y.; Zhang, L. The structures and biological functions of polysaccharides from traditional Chinese herbs. Prog. Mol. Biol. Transl. Sci. 2019, 163, 423–444. [Google Scholar] [CrossRef]
- Shan, H.; Zheng, X.; Li, M. The Effects of Astragalus membranaceus Active Extracts on Autophagy-Related Diseases. Int. J. Mol. Sci. 2019, 20, 1904. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Wang, Z.; Huang, L.; Zheng, S.; Wang, D.; Chen, S.; Zhang, H.; Yang, S. Review of the botanical characteristics, phytochemistry, and pharmacology of astragalus membranaceus (huangqi). Phytother. Res. 2014, 28, 1275–1283. [Google Scholar] [CrossRef]
- Mo, W.Y.; Lun, C.H.I.; Choi, W.M.; Man, Y.B.; Wong, M.H. Enhancing growth and non-specific immunity of grass carp and Nile tilapia by incorporating Chinese herbs (Astragalus membranaceus and Lycium barbarum) into food waste based pellets. Environ. Pollut. 2016, 219, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Elabd, H.; Wang, H.-P.; Shaheen, A.; Matter, A. Astragalus membranaceus nanoparticles markedly improve immune and anti-oxidative responses; and protection against Aeromonas veronii in Nile tilapia Oreochromis niloticus. Fish Shellfish Immunol. 2019, 97, 248–256. [Google Scholar] [CrossRef]
- Wang, X.; Ding, L.; Yan, M.; Chai, X.; Lu, R.; Wang, Q.; Li, F. Polysaccharides, Saponins, and Water Decoction of Astragalus membranaceus Significantly Enhance the Non-Specific Immune Response of Spotted Maigre (Nibea albiflora). Isr. J. Aquac. Bamidgeh 2012, 64, 20621. [Google Scholar] [CrossRef]
- Abdel-Latif, H.M.; Ahmed, H.A.; Shukry, M.; Chaklader, M.R.; Saleh, R.M.; Khallaf, M.A. Astragalus membranaceus extract (ame) enhances growth, digestive enzymes, antioxidant capacity, and immunity of pangasianodon hypophthalmus juveniles. Fishes 2022, 7, 319. [Google Scholar] [CrossRef]
- Shi, F.; Lu, Z.; Yang, M.; Li, F.; Zhan, F.; Zhao, L.; Li, Y.; Li, Q.; Li, J.; Li, J. Astragalus polysaccharides mediate the immune response and intestinal microbiota in grass carp (ctenopharyngodon idellus)—Sciencedirect. Aquaculture 2020, 534, 736205. [Google Scholar] [CrossRef]
- Marioni, J.C.; Mason, C.E.; Mane, S.M.; Stephens, M.; Gilad, Y. RNA-seq: An assessment of technical reproducibility and comparison with gene expression arrays. Genome Res. 2008, 18, 1509–1517. [Google Scholar] [CrossRef]
- Liu, Y.; Xin, Z.-Z.; Zhang, D.-Z.; Wang, Z.-F.; Zhu, X.-Y.; Tang, B.-P.; Jiang, S.-H.; Zhang, H.-B.; Zhou, C.-L.; Chai, X.-Y.; et al. Transcriptome analysis of yellow catfish (Pelteobagrus fulvidraco) liver challenged with polyriboinosinic polyribocytidylic acid (poly I:C). Fish Shellfish Immunol. 2017, 68, 395–403. [Google Scholar] [CrossRef]
- Tao, Y.-F.; Qiang, J.; Dagoudo, M.; Zhu, H.-J.; Bao, J.-W.; Ma, J.-L.; Li, M.-X.; Xu, P. Transcriptome profiling reveals differential expression of immune-related genes in gills of hybrid yellow catfish (Tachysurus fulvidraco ♀ × Pseudobagrus vachellii ♂) under hypoxic stress: Potential NLR-mediated immune response. Fish Shellfish Immunol. 2021, 119, 409–419. [Google Scholar] [CrossRef]
- Zhang, J.; Ma, W.; Song, X.; Lin, Q.; Gui, J.-F.; Mei, J. Characterization and Development of EST-SSR Markers Derived from Transcriptome of Yellow Catfish. Molecules 2014, 19, 16402–16415. [Google Scholar] [CrossRef]
- Chen, X.; Mei, J.; Wu, J.; Jing, J.; Ma, W.; Zhang, J.; Dan, C.; Wang, W.; Gui, J.-F. A Comprehensive Transcriptome Provides Candidate Genes for Sex Determination/Differentiation and SSR/SNP Markers in Yellow Catfish. Mar. Biotechnol. 2015, 17, 190–198. [Google Scholar] [CrossRef]
- Bian, L.; Liu, C.; Chen, S.; Zhao, F.; Ge, J.; Tan, J. Transcriptome analysis of gene expression patterns during embryonic development in golden cuttlefish (sepia esculenta). Genes Genom. 2018, 40, 253–263. [Google Scholar] [CrossRef]
- Kim, W.-J.; Lee, K.; Lee, D.; Kim, H.-C.; Nam, B.-H.; Jung, H.; Yi, S.-J.; Kim, K. Transcriptome profiling of olive flounder responses under acute and chronic heat stress. Genes Genom. 2021, 43, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Wang, Y.; Zhang, Z.; He, S. Comprehensive transcriptome analysis reveals accelerated genic evolution in a tibet fish, gymnodiptychus pachycheilus. Genome Biol. Evol. 2015, 7, 251–261. [Google Scholar] [CrossRef] [PubMed]
- Carruthers, M.; Yurchenko, A.A.; Augley, J.J.; Adams, C.E.; Herzyk, P.; Elmer, K.R. De novo transcriptome assembly, annotation and comparison of four ecological and evolutionary model salmonid fish species. BMC Genom. 2018, 19, 4379. [Google Scholar]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Tatusov, R.L.; Galperin, M.Y.; Natale, D.A.; Koonin, E.V. The COG database: A tool for genome-scale analysis of protein functions and evolution. Nucleic Acids Res. 2000, 28, 33–36. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D. Gene ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef]
- Minoru, K.; Susumu, G.; Shuichi, K.; Yasushi, O.; Masahiro, H. The kegg resource for deciphering the genome. Nucleic Acids Res. 2004, 32, D277. [Google Scholar]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Huber, W. Differential Expression of rna-seq Data at the Gene Level–The Deseq Package. Heidelberg, Germany: European Molecular Biology Laboratory (EMBL) 10 (2012): F1000research. Available online: https://www.genomatix.de/online_help/help_regionminer/DESeq_1.10.1.pdf (accessed on 8 December 2021).
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T.; et al. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2007, 36, D480–D484. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.-N.; Xin, Z.-Z.; Chai, X.-Y.; Jiang, S.-H.; Li, C.-F.; Zhang, H.-B.; Ge, B.-M.; Zhang, D.-Z.; Zhou, C.-L.; Tang, B.-P. Characterization of immune-related genes in the yellow catfish Pelteobagrus fulvidraco in response to LPS challenge. Fish Shellfish Immunol. 2016, 56, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Zhu, R.; Liu, X.-X.; Lv, X.; Li, S.-Y.; Li, Y.-D.; Yu, X.-J.; Wang, X.-G. Deciphering transcriptome profile of the yellow catfish (Pelteobagrus fulvidraco) in response to Edwardsiella ictaluri. Fish Shellfish Immunol. 2017, 70, 593–608. [Google Scholar] [CrossRef]
- Li, Z.; Liu, X.; Cheng, J.; He, Y.; Wang, X.; Wang, Z.; Qi, J.; Yu, H.; Zhang, Q. Transcriptome profiling provides gene resources for understanding gill immune responses in Japanese flounder (Paralichthys olivaceus) challenged with Edwardsiella tarda. Fish Shellfish Immunol. 2018, 72, 593–603. [Google Scholar] [CrossRef]
- Taylor, G.A. IRG proteins: Key mediators of interferon-regulated host resistance to intracellular pathogens. Cell. Microbiol. 2007, 9, 1099–1107. [Google Scholar] [CrossRef]
- Hunn, J.P.; Feng, C.G.; Sher, A.; Howard, J.C. The immunity-related GTPases in mammals: A fast-evolving cell-autonomous resistance system against intracellular pathogens. Mamm. Genome 2010, 22, 43–54. [Google Scholar] [CrossRef] [PubMed]
- MacMicking, J.D. Immune control of tuberculosis by ifn-γ-inducible lrg-47. Science 2003, 302, 654–659. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Shi, Y.; Ding, T.; Duan, W.; Qiu, Z.; Zhao, Z. Functional characterization of an immunity-related GTPase gene in immune defense from obscure puffer (Takifugu obscurus). Fish Shellfish Immunol. 2020, 103, 248–255. [Google Scholar] [CrossRef]
- Boles, K.S.; Mathew, P.A. Molecular cloning of CS1, a novel human natural killer cell receptor belonging to the CD2 subset of the immunoglobulin superfamily. Immunogenetics 2001, 52, 302–307. [Google Scholar] [CrossRef] [PubMed]
- Simmons, D.P.; Nguyen, H.N.; Gomez-Rivas, E.; Jeong, Y.; Brenner, M.B. Slamf7 Engagement Super-Activates Macrophages in Acute and Chronic Inflammation; Cold Spring Harbor Laboratory: New York, NY, USA, 2020. [Google Scholar]
- Choe, U.; Pham, Q.; Kim, Y.S.; Yu, L.; Wang, T.T.Y. Identification and elucidation of cross talk between SLAM Family Member 7 (SLAMF7) and Toll-like receptor (TLR) pathways in monocytes and macrophages. Sci. Rep. 2023, 13, 1–13. [Google Scholar] [CrossRef]
- Li, K.; Underhill, D. C-type lectin receptors in phagocytosis. Curr. Top. Microbiol. Immunol. Undefined 2020, in press. [Google Scholar]
- Yu, M.; Chang, S.; Xu, J.; Zhang, H.; Jiang, Y. Genome-wide identification of endosialin family of C-type lectins in common carp (Cyprinus carpio) and their response following Aeromonas hydrophila infection. Dev. Comp. Immunol. 2022, 129, 104338. [Google Scholar] [CrossRef] [PubMed]
- Mu, L.; Yin, X.; Liu, J.; Wu, L.; Bian, X.; Wang, Y.; Ye, J. Identification and characterization of a mannose-binding lectin from Nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2017, 67, 244–253. [Google Scholar] [CrossRef]
- Liu, F.; Li, J.; Fu, J.; Shen, Y.; Xu, X. Two novel homologs of simple C-type lectin in grass carp (Ctenopharyngodon idellus): Potential role in immune response to bacteria. Fish Shellfish Immunol. 2011, 31, 765–773. [Google Scholar] [CrossRef]
- Gaozza, E.; Baker, S.J.; Vora, R.K.; Reddy, E.P. AATYK: A novel tyrosine kinase induced during growth arrest and apoptosis of myeloid cells. Oncogene 1997, 15, 3127–3135. [Google Scholar] [CrossRef]
- Haag, T.; Herkt, C.E.; Walesch, S.K.; Richter, A.M.; Dammann, R.H. The apoptosis associated tyrosine kinase gene is frequently hypermethylated in human cancer and is regulated by epigenetic mechanisms. Genes Cancer 2014, 5, 365–374. [Google Scholar] [CrossRef]
- Steinmann, S.; Scheibe, K.; Erlenbach-Wuensch, K.; Neufert, C.; Schneider-Stock, R. Death-associated protein kinase: A molecule with functional antagonistic duality and a potential role in inflammatory bowel disease (Review). Int. J. Oncol. 2015, 47, 5–15. [Google Scholar] [CrossRef]
- Xu, Y.; Feng, Y.; Li, S.; Sun, J. Identification and characterization of apoptosis-related gene serine/threonine kinase 17A (STK17A) from Japanese flounder Paralichthys olivaceus. Fish Shellfish Immunol. 2020, 98, 1008–1016. [Google Scholar] [CrossRef]
- Nagy, N.; Matskova, L.; Kis, L.L.; Hellman, U.; Klein, G.; Klein, E. The proapoptotic function of SAP provides a clue to the clinical picture of X-linked lymphoproliferative disease. Proc. Natl. Acad. Sci. USA 2009, 106, 11966–11971. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zhao, J.; Li, Y.; Zou, Y.; Lu, B.; Chen, Y.; Ma, Y.; Xu, H. Evaluation of differentially expressed immune-related genes in intestine of Pelodiscus sinensis after intragastric challenge with lipopolysaccharide based on transcriptome analysis. Fish Shellfish Immunol. 2016, 56, 417–426. [Google Scholar] [CrossRef]
- Brady, H.J.; Gil-Gómez, G. Molecules in focus bax. The pro-apoptotic bcl-2 family member, bax. Int. J. Biochem. Cell Biol. 1998, 30, 647–650. [Google Scholar] [CrossRef] [PubMed]
- Schröder, J.; Lüllmann-Rauch, R.; Himmerkus, N.; Pleines, I.; Nieswandt, B.; Orinska, Z.; Koch-Nolte, F.; Schröder, B.; Bleich, M.; Saftig, P. Deficiency of the tetraspanin cd63 associated with kidney pathology but normal lysosomal function. Mol. Cell. Biol. 2009, 29, 1083–1094. [Google Scholar] [CrossRef]
- Priyathilaka, T.T.; Bathige, S.; Herath, H.; Lee, S.; Lee, J. Molecular identification of disk abalone (Haliotis discus discus) tetraspanin 33 and CD63: Insights into potent players in the disk abalone host defense system. Fish Shellfish Immunol. 2017, 69, 173–184. [Google Scholar] [CrossRef]
- Ooms, L.M.; Horan, K.A.; Rahman, P.; Seaton, G.; Gurung, R.; Kethesparan, D.S.; Mitchell, C.A. The role of the inositol polyphosphate 5-phosphatases in cellular function and human disease. Biochem. J. 2009, 419, 29–49. [Google Scholar] [CrossRef] [PubMed]
- Dimitriadis, E.; White, C.; Jones, R.; Salamonsen, L. Cytokines, chemokines and growth factors in endometrium related to implantation. Hum. Reprod. Updat. 2005, 11, 613–630. [Google Scholar] [CrossRef]
- Bai, D.; Wu, X.; Zhu, G.; Guo, Y.; Yang, G.; Ning, B.; Xing, K. Astragalus Polysaccharides Enhance Cellular Immune Response and Disease Resistance in Yellow Catfish. Isr. J. Aquac. 2012, 64. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Primers Sequences |
|---|---|
| aatk | F: GGTGATGGCTACCAATCAGA |
| R: TGCTCCCATTTGTACTCAAAG | |
| DAPK2 | F: AGGTGGTCCTCATCGTAG |
| R: TTCAGGAACTTGATCGCGT | |
| IL6ST | F: CTCAGAAGCCTACATGGTATC |
| R: TTAGGTTTAGCGTTGTTGCG | |
| Bax | F: GACCAGATCGATGGTTTAGCA |
| R: TGTCCATCATCGAAGACGC | |
| Cd63 | F: TCTTGGCCCTGATAATCGTAG |
| R: CGATGGAAGACTTCATCCG | |
| inppl1b | F: CCTGGTAATCCCTTTGCTTTAT |
| R: TGGCTTCTCATCATCTCCG | |
| Sh2d1a | F: AAAGCATTCCTTCGCAGAT |
| R: AAGTGATGTTCAGAGCCAG | |
| β-Actin | F: GCACAGTAAAGGCGTTGTGA |
| R: ACATCTGCTGGAAGGTGGAC |
| RIN Value | Raw Reads (M) | Clean Reads (M) | Clean Bases (G) | GC Percentage (%) | Q30 (%) | |
|---|---|---|---|---|---|---|
| CS241 | 9.5 | 46.25 | 44.93 | 6.54 | 46.43 | 92.01 |
| CS242 | 9.2 | 48.70 | 47.43 | 6.91 | 46.58 | 92.54 |
| CS243 | 9.7 | 50.49 | 49.17 | 7.17 | 46.31 | 92.42 |
| AS241 | 9.9 | 47.41 | 46.09 | 6.71 | 46.17 | 92.00 |
| AS242 | 9.4 | 46.40 | 45.08 | 6.56 | 46.19 | 91.97 |
| AS243 | 9.5 | 49.61 | 48.29 | 7.04 | 46.41 | 92.16 |
| Sample | Total Reads | Map Reads | Map Ratio |
|---|---|---|---|
| CS241 | 44,929,764 | 34,416,346 | 76.60% |
| CS242 | 47,428,338 | 36,342,453 | 76.63% |
| CS243 | 49,172,944 | 37,735,697 | 76.74% |
| AS241 | 46,093,010 | 34,991,235 | 75.91% |
| AS242 | 45,079,552 | 34,534,151 | 76.61% |
| AS243 | 48,294,024 | 36,864,149 | 76.33% |
| Group | Challenged Fish Number | Dead Fish Number | Mortality (%) | Relative Percent Survival (%) |
|---|---|---|---|---|
| Astragalus membranaceus | 3 × 10 | 3 | 10.00 | 85.71 |
| control | 3 × 10 | 21 | 70.00 | / |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pan, T.; Yang, M.; Jiang, H.; Li, T.; Duan, G.; Ling, J.; Gao, Q. Effect of Astragalus membranaceus on Transcriptome and Survival of Hybrid Yellow Catfish (Pseudobagrus vachellii ♂ × Tachysurus fulvidraco ♀) in Response to Aeromonas hydrophila Challenge. Fishes 2023, 8, 454. https://doi.org/10.3390/fishes8090454
Pan T, Yang M, Jiang H, Li T, Duan G, Ling J, Gao Q. Effect of Astragalus membranaceus on Transcriptome and Survival of Hybrid Yellow Catfish (Pseudobagrus vachellii ♂ × Tachysurus fulvidraco ♀) in Response to Aeromonas hydrophila Challenge. Fishes. 2023; 8(9):454. https://doi.org/10.3390/fishes8090454
Chicago/Turabian StylePan, Tingshuang, Min Yang, He Jiang, Tong Li, Guoqing Duan, Jun Ling, and Qian Gao. 2023. "Effect of Astragalus membranaceus on Transcriptome and Survival of Hybrid Yellow Catfish (Pseudobagrus vachellii ♂ × Tachysurus fulvidraco ♀) in Response to Aeromonas hydrophila Challenge" Fishes 8, no. 9: 454. https://doi.org/10.3390/fishes8090454
APA StylePan, T., Yang, M., Jiang, H., Li, T., Duan, G., Ling, J., & Gao, Q. (2023). Effect of Astragalus membranaceus on Transcriptome and Survival of Hybrid Yellow Catfish (Pseudobagrus vachellii ♂ × Tachysurus fulvidraco ♀) in Response to Aeromonas hydrophila Challenge. Fishes, 8(9), 454. https://doi.org/10.3390/fishes8090454