
Effects of High Dietary Starch Levels on the Growth Performance, Liver Function, and Metabolome of Largemouth Bass (Micropterus salmoides)
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Diets
2.2. Fish Management
2.3. Sample Collection
2.4. Chemical Analysis
2.5. Hepatic Transcriptome Analysis
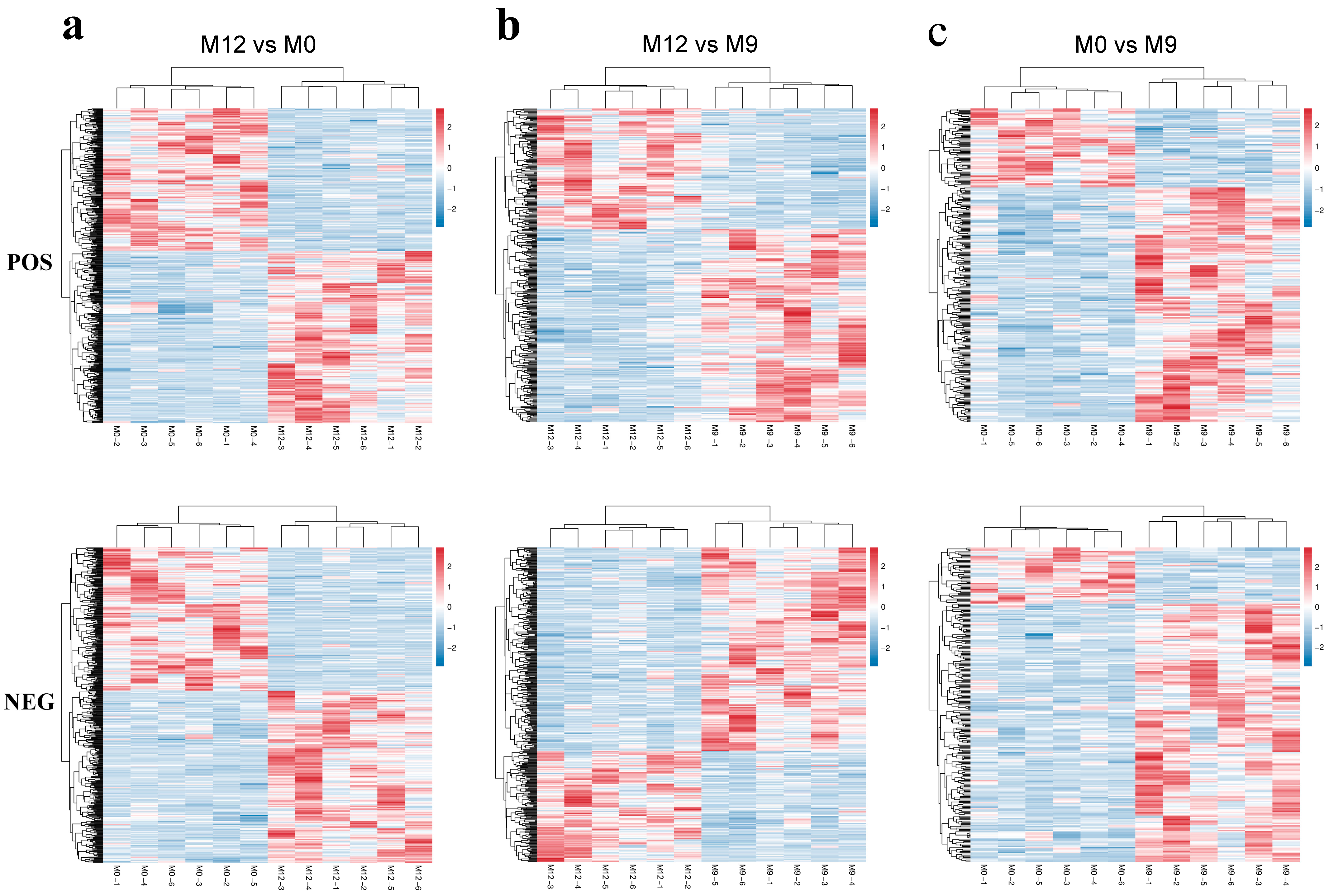
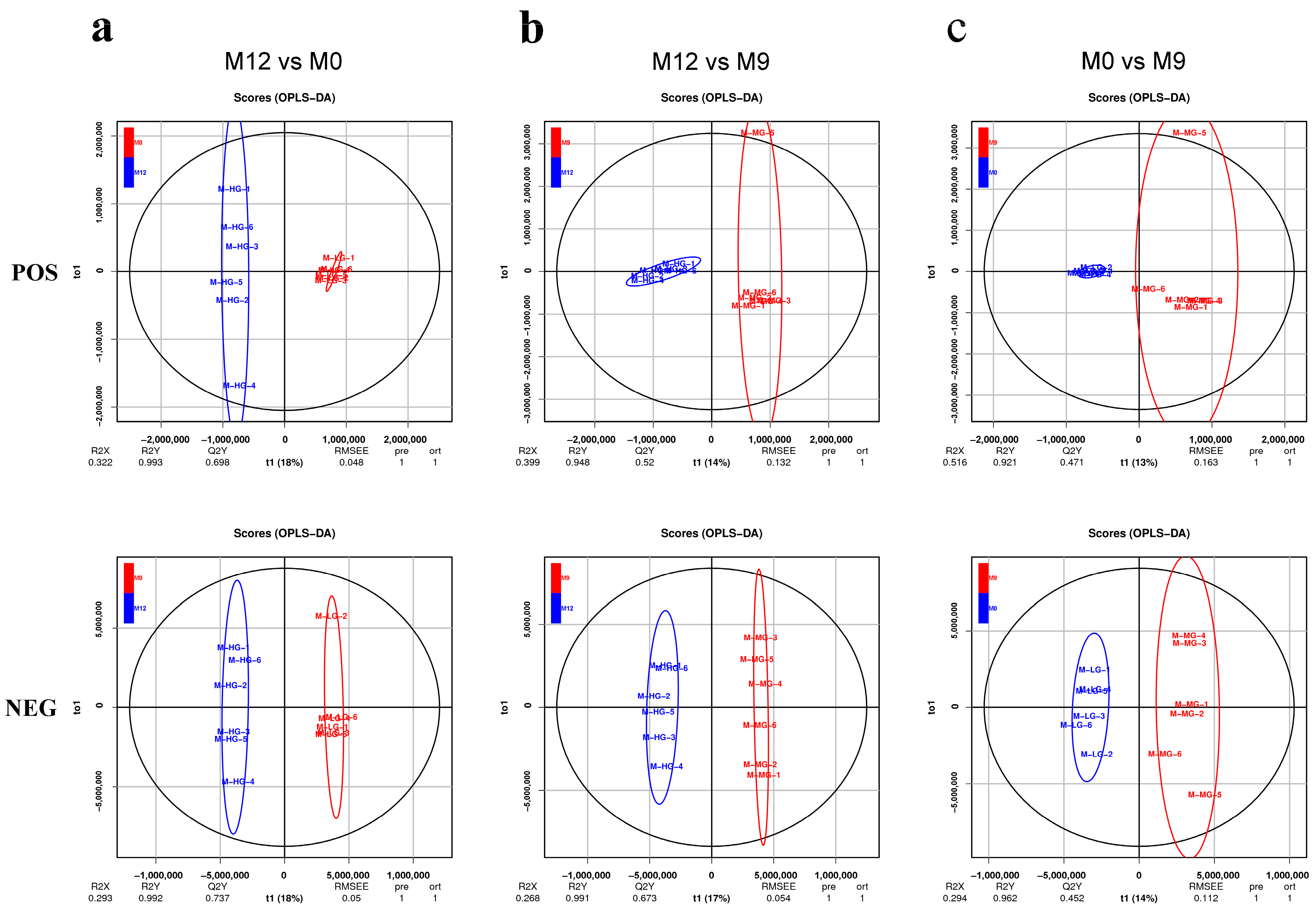
2.6. LC-MS/MS Non-Target Metabonomics
2.7. Calculations
2.8. Statistical Analysis
3. Results
3.1. Growth Performance and Feed Utilization
3.2. Morphometric Parameters and Chemical Composition
3.3. Intestinal Digestive and Absorption Enzyme Activity
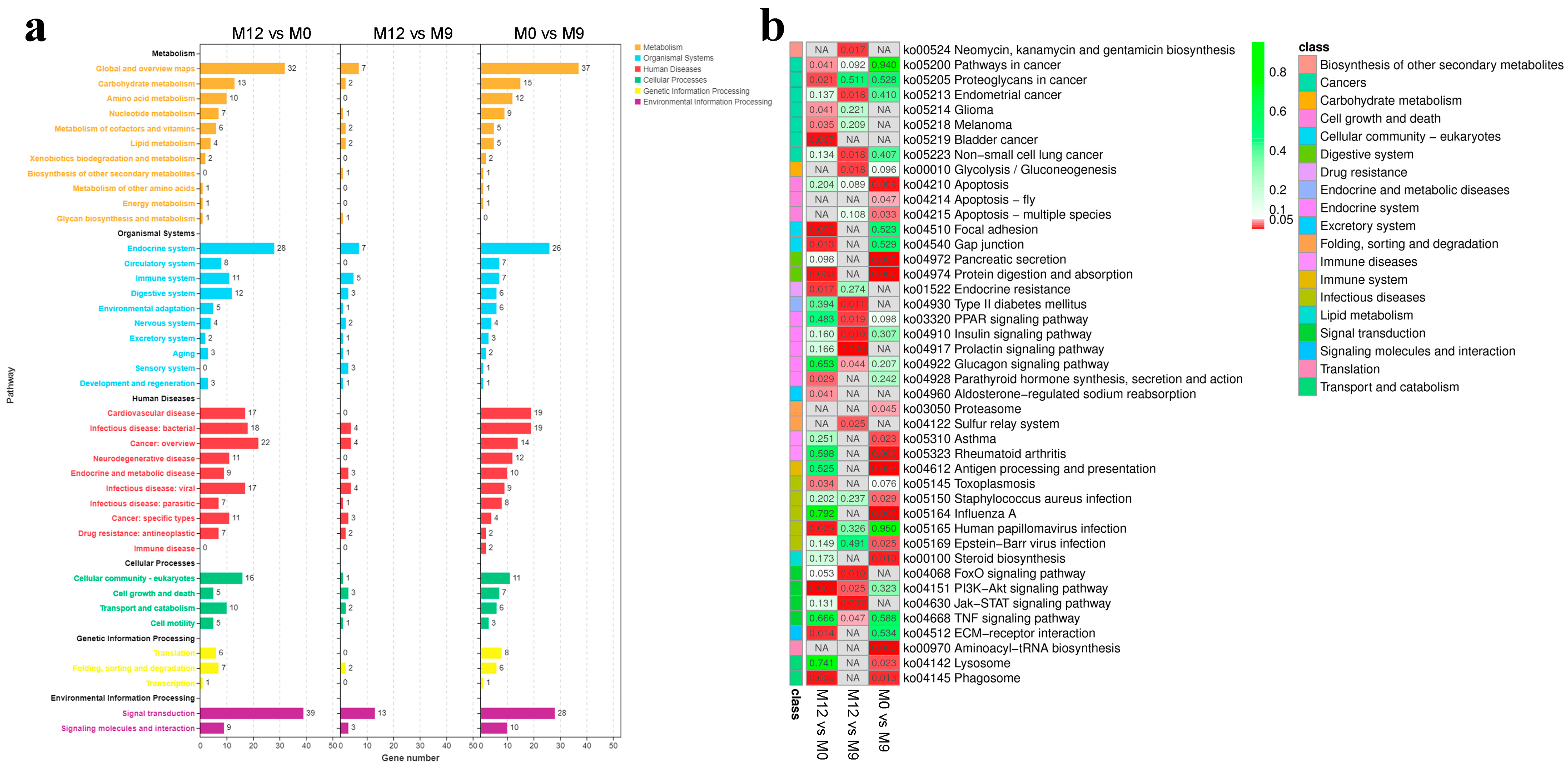
3.4. Detection and Functional Enrichment Analysis of DEGs in Liver
3.5. Liver Metabolome
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sobiecki, J.G.; Appleby, P.N.; Bradbury, K.E.; Key, T.J. High compliance with dietary recommendations in a cohort of meat eaters, fish eaters, vegetarians, and vegans: Results from the European Prospective Investigation into Cancer and Nutrition-Oxford study. Nutr. Res. 2016, 36, 464–477. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.J.; Zhou, Q.C.; Liang, H.O.; Yang, J.; Zhao, L.M. Effects of dietary carbohydrate sources on the growth performance and hepatic carbohydrate metabolic enzyme activities of juvenile cobia (Rachycentron canadum Linnaeus.). Aquac. Res. 2010, 42, 99–107. [Google Scholar] [CrossRef]
- Prakash, S.; Maas, R.M.; Horstmann, P.; Elbers, J.J.; Kokou, F.; Schrama, J.W.; Philip, A.J.P. Effect of dietary starch, amylase and ash on nutrient digestibility, faecal waste production and faecal characteristics of rainbow trout, (Oncorhynchus mykiss). Aquaculture 2024, 583, 740612. [Google Scholar] [CrossRef]
- Hoseini, S.M.; Taheri Mirghaed, A.; Ghelichpour, M.; Pagheh, E.; Iri, Y.; Kor, A. Effects of dietary tryptophan supplementation and stocking density on growth performance and stress responses in rainbow trout (Oncorhynchus mykiss). Aquaculture 2020, 519, 734908. [Google Scholar] [CrossRef]
- Lin, S.M.; Shi, C.M.; Mu, M.M.; Chen, Y.J.; Luo, L. Effect of high dietary starch levels on growth, hepatic glucose metabolism, oxidative status and immune response of juvenile largemouth bass, Micropterus salmoides. Fish. Shellfish. Immunol. 2018, 78, 121–126. [Google Scholar] [CrossRef]
- Guo, J.L.; Kuang, W.M.; Zhong, Y.F.; Zhou, Y.L.; Chen, Y.J.; Lin, S.M. Effects of supplemental dietary bile acids on growth, liver function and immunity of juvenile largemouth bass (Micropterus salmoides) fed high-starch diet. Fish. Shellfish. Immunol. 2020, 97, 602–607. [Google Scholar] [CrossRef]
- Luchese, C.L.; Rodrigues, R.B.; Tessaro, I.C. Cassava starch-processing residue utilization for packaging development. Int. J. Biol. Macromol. 2021, 183, 2238–2247. [Google Scholar] [CrossRef]
- Fan, M.; Hu, T.; Zhao, S.; Xiong, S.; Xie, J.; Huang, Q. Gel characteristics and microstructure of fish myofibrillar protein/cassava starch composites. Food Chem. 2017, 218, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Oliveira Cavalheiro, J.M.; Oliveira de Souza, E.; Bora, P.S. Utilization of shrimp industry waste in the formulation of tilapia (Oreochromis niloticus Linnaeus) feed. Bioresour. Technol. 2007, 98, 602–606. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Huang, Q.; Hu, T.; Xiong, S.; Zhao, S. Effects and mechanism of modified starches on the gel properties of myofibrillar protein from grass carp. Int. J. Biol. Macromol. 2014, 64, 17–24. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of AOAC; AOAC International: Gaithersburg, MD, USA, 2005. [Google Scholar]
- Yuan, X.Y.; Zhang, X.T.; Xia, Y.T.; Zhang, Y.Q.; Wang, B.; Ye, W.W.; Ye, Z.F.; Qian, S.C.; Huang, M.M.; Yang, S.; et al. Transcriptome and 16S rRNA analyses revealed differences in the responses of largemouth bass (Micropterus salmoides) to early Aeromonas hydrophila infection and immunization. Aquaculture 2021, 541, 736759. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Ning, X.; Peng, Y.; Tang, P.; Zhang, Y.; Wang, L.; Zhang, W.; Zhang, K.; Ji, J.; Yin, S. Integrated analysis of transcriptome and metabolome reveals distinct responses of Pelteobagrus fulvidraco against Aeromonas veronii infection at invaded and recovering stage. Int. J. Mol. Sci. 2022, 23, 10121. [Google Scholar] [CrossRef]
- Wang, J.; Li, X.; Han, T.; Yang, Y.; Jiang, Y.; Yang, M.; Xu, Y.; Harpaz, S. Effects of different dietary carbohydrate levels on growth, feed utilization and body composition of juvenile grouper Epinephelus akaara. Aquaculture 2016, 459, 143–147. [Google Scholar] [CrossRef]
- Mahanama, D.; Radampola, K.; Heenkenda, E. Effect of cassava starch sources on growth and feed utilization of Nile Tilapia fingerlings (Oreochromis niloticus) reared under two dietary protein levels. Aquac. Stud. 2021, 21, 169–179. [Google Scholar] [CrossRef]
- Olurin, K.B. Growth of African catfish Clarias gariepinus fingerlings, fed different levels of cassava. Int. Res. J. Agric. Rural. Dev. 2013, 22, 4334–4343. [Google Scholar]
- Boonkusol, D.; Tongbai, W. Beneficial effects of soybean and cassava in local wisdom feed diets on growth performance with nutritional and economic analysis of snakehead fish (Channa striata). OnLine J. Biol. Sci. 2022, 22, 463–468. [Google Scholar] [CrossRef]
- Ding, X.; Nie, X.; Yuan, C.; Jiang, L.; Ye, W.; Qian, L. Effects of dietary multienzyme complex supplementation on growth performance, digestive capacity, histomorphology, blood metabolites and hepatic glycometabolism in snakehead (Channa argus). Animals 2022, 12, 380. [Google Scholar] [CrossRef]
- Liu, Y.; Fan, J.; Huang, H.; Zhou, H.; Cao, Y.; Zhang, Y.; Jiang, W.; Zhang, W.; Deng, J.; Tan, B. High dietary non-starch polysaccharides detrimental to nutrient digestibility, digestive enzyme activity, growth performance, and intestinal morphology in largemouth bass, Micropterus salmoides. Front. Nutr. 2022, 9, 1015371. [Google Scholar] [CrossRef]
- Krogdahl, Å.; Hemre, G.I.; Mommsen, T.P. Carbohydrates in fish nutrition: Digestion and absorption in postlarval stages. Aquac. Nutr. 2005, 11, 103–122. [Google Scholar] [CrossRef]
- Gaylord, T.G.; BARROWS, F.T.; RAWLES, S.D.; LIU, K.; BREGITZER, P.; HANG, A.; OBERT, D.E.; MORRIS, C. Apparent digestibility of nutrients and energy in extruded diets from cultivars of barley and wheat selected for nutritional quality in rainbow trout Oncorhynchus mykiss. Aquac. Nutr. 2009, 15, 306–312. [Google Scholar] [CrossRef]
- Hu, G.; Gu, W.; Sun, P.; Bai, Q.; Wang, B. Transcriptome analyses reveal lipid metabolic process in liver related to the difference of carcass fat content in rainbow trout (Oncorhynchus mykiss). Int. J. Genom. 2016, 2016, 7281585. [Google Scholar] [CrossRef]
- Jin, G.; Zhang, L.; Mai, K.; Chen, X.; Xu, S.; Ai, Q. Effects of different dietary lipid sources on growth performance, hepatic lipid deposition and transcriptome response in spotted sea bass (Lateolabrax maculatus). Aquaculture 2023, 566, 739143. [Google Scholar] [CrossRef]
- Ma, D.; Fan, J.; Zhu, H.; Su, H.; Jiang, P.; Yu, L.; Liao, G.; Bai, J. Histologic examination and transcriptome analysis uncovered liver damage in largemouth bass from formulated diets. Aquaculture 2020, 526, 735329. [Google Scholar] [CrossRef]
- Caseras, A.; Metón, I.; Fernández, F.; Baanante, I.V. Glucokinase gene expression is nutritionally regulated in liver of gilthead sea bream (Sparus aurata). Biochim. Et Biophys. Acta (BBA) Gene Struct. Expr. 2000, 1493, 135–141. [Google Scholar] [CrossRef]
- Belghit, I.; Philip, A.J.P.; Maas, R.M.; Lock, E.-J.; Eding, E.H.; Espe, M.; Schrama, J.W. Impact of dietary glutamate and glycine on growth and nutrient utilization in rainbow trout (Oncorhynchus mykiss). Aquaculture 2023, 568, 739311. [Google Scholar] [CrossRef]
- Omura, Y.; Inagaki, M. Immunocytochemical localization of taurine in the fish retina under light and dark adaptations. Amino Acids 2000, 19, 593–604. [Google Scholar] [CrossRef]
- Gonzalez, J.; Bahmad, H.F.; Ocejo, S.; Abreu, A.; Popp, M.; Gogola, S.; Fernandez, V.; Recine, M.; Poppiti, R. The usefulness of elastin staining to detect vascular invasion in cancer. Int. J. Mol. Sci. 2023, 24, 15264. [Google Scholar] [CrossRef]
- Li, P.; Mai, K.; Trushenski, J.; Wu, G. New developments in fish amino acid nutrition: Towards functional and environmentally oriented aquafeeds. Amino Acids 2009, 37, 43–53. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, J.; Wang, Q.; Han, F.; Shi, L.; Han, C.; Huang, Z.; Xu, L. Effects of insufficient serine on health and selenoprotein expression in rats and their offspring. Front. Nutr. 2022, 9, 1012362. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.-Z.; Yang, S.; Wu, G. Free radicals, antioxidants, and nutrition. Nutrition 2002, 18, 872–879. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | Percentage (%) | ||||
|---|---|---|---|---|---|
| M12 | M9 | M6 | M3 | M0 | |
| Fish meal a | 40 | ||||
| Chicken meal | 6 | ||||
| Yeast extract | 6 | ||||
| Enzymolysis intestinal membrane protein powder | 5 | ||||
| Spray-dried animal blood cells | 2.5 | ||||
| Corn protein powder | 4 | ||||
| Shrimp meal | 4 | ||||
| Cottonseed meal | 6 | ||||
| Soya bean meal | 3.5 | ||||
| Fermented soybean meal | 5 | ||||
| Wheat gluten | 0 | 3 | 6 | 9 | 12 |
| Cassava starch | 12 | 9 | 6 | 3 | 0 |
| Ca(H2PO4) | 2 | ||||
| Soybean oil | 4 | ||||
| Premix b | 3 | ||||
| Total | 100 | ||||
| Nutrient levels | |||||
| Moisture | 6.08 | 5.41 | 5.21 | 5.89 | 5.08 |
| Crude protein | 49.90 | 50.53 | 51.05 | 51.59 | 52.08 |
| Crude lipid | 13.76 | 12.89 | 13.74 | 13.65 | 13.03 |
| Crude ash | 11.81 | 12.47 | 12.46 | 12.24 | 12.49 |
| Starch | 12.71 | 11.66 | 10.70 | 9.83 | 8.91 |
| Final BW (g/Fish) | Weight Gain Rate (WGR, %) | Specific Growth Rate (SGR, %/d) | Feed Conversion Rate (FCR, %) | Protein Efficiency Ratio (PER) | Protein Retention Efficiency (PRE, %) | |
|---|---|---|---|---|---|---|
| M12 | 234.773 ± 4.664 a | 181.7 ± 5.5 a | 0.930 ± 0.020 a | 0.837 ± 0.050 | 1.670 ± 0.095 | 1.820 ± 0.060 a |
| M9 | 261.990 ± 7.029 b | 214.7 ± 8.5 b | 1.030 ± 0.020 b | 0.893 ± 0.021 | 1.770 ± 0.040 | 2.200 ± 0.085 b |
| M6 | 256.243 ± 3.925 b | 207.7 ± 5.0 b | 1.013 ± 0.015 b | 0.857 ± 0.057 | 1.677 ± 0.110 | 1.933 ± 0.045 a |
| M3 | 251.163 ± 5.329 ab | 204.7 ± 11.6 b | 0.993 ± 0.021 b | 0.870 ± 0.056 | 1.687 ± 0.106 | 1.920 ± 0.062 a |
| M0 | 251.733 ± 18.367 ab | 202.0 ± 21.9 ab | 0.993 ± 0.064 b | 0.933 ± 0.074 | 1.783 ± 0.144 | 1.980 ± 0.219 a |
| Condition Factor (CF, g/cm3) | Viscerosomatic Index (VSI, %) | Hepatosomatic Index (HSI, %) | Enterosomatic Index (ESI, %) | Gonadosomatic Index (GSI, %) | ||
|---|---|---|---|---|---|---|
| Male | M12 | 2.290 ± 0.159 | 7.213 ± 0.214 ab | 2.437 ± 0.175 a | 0.580 ± 0.017 | 0.647 ± 0.058 |
| M9 | 2.367 ± 0.075 | 6.713 ± 0.120 b | 1.970 ± 0.036 b | 0.550 ± 0.010 | 0.737 ± 0.021 | |
| M6 | 2.337 ± 0.029 | 7.290 ± 0.598 a | 2.407 ± 0.212 a | 0.550 ± 0.040 | 0.727 ± 0.137 | |
| M3 | 2.260 ± 0.108 | 6.657 ± 0.042 b | 2.187 ± 0.057 ab | 0.577 ± 0.038 | 0.690 ± 0.075 | |
| M0 | 2.317 ± 0.083 | 6.663 ± 0.264 b | 2.123 ± 0.172 b | 0.533 ± 0.015 | 0.697 ± 0.049 | |
| Female | M12 | 2.380 ± 0.108 a | 7.140 ± 0.220 a | 2.520 ± 0.203 a | 0.517 ± 0.006 | 1.607 ± 0.148 |
| M9 | 2.433 ± 0.101 a | 6.803 ± 0.211 ab | 2.283 ± 0.215 ab | 0.567 ± 0.047 | 1.947 ± 0.516 | |
| M6 | 2.290 ± 0.053 b | 7.140 ± 0.195 a | 2.423 ± 0.195 a | 0.573 ± 0.038 | 1.757 ± 0.234 | |
| M3 | 2.163 ± 0.021 b | 6.680 ± 0.378 bc | 2.350 ± 0.344 ab | 0.560 ± 0.036 | 1.857 ± 0.172 | |
| M0 | 2.323 ± 0.04 a | 6.353 ± 0.040 c | 1.980 ± 0.171 b | 0.543 ± 0.031 | 1.810 ± 0.377 |
| Moisture | Crude Protein | Crude Lipid | Crude Ash | |||
|---|---|---|---|---|---|---|
| Whole fish | Male | M12 | 68.740 ± 1.498 | 17.740 ± 0.750 | 8.743 ± 0.486 ab | 4.317 ± 0.188 a |
| M9 | 67.117 ± 1.805 | 17.917 ± 0.677 | 9.390 ± 0.631 a | 3.780 ± 0.035 b | ||
| M6 | 67.500 ± 0.637 | 17.173 ± 1.210 | 8.873 ± 0.605 ab | 3.840 ± 0.156 b | ||
| M3 | 66.987 ± 0.982 | 17.073 ± 1.159 | 9.390 ± 0.498 a | 4.000 ± 0.241 ab | ||
| M0 | 67.783 ± 1.134 | 17.387 ± 0.110 | 8.223 ± 0.649 b | 4.183 ± 0.228 a | ||
| Female | M12 | 68.627 ± 0.442 a | 17.067 ± 0.511 | 8.997 ± 0.055 | 4.033 ± 0.298 ab | |
| M9 | 67.980 ± 0.592 a | 17.480 ± 0.339 | 9.217 ± 0.532 | 3.837 ± 0.158 ab | ||
| M6 | 66.443 ± 0.798 b | 17.343 ± 0.910 | 8.513 ± 1.028 | 4.047 ± 0.193 ab | ||
| M3 | 67.433 ± 0.307 ab | 16.627 ± 0.100 | 8.723 ± 0.879 | 4.227 ± 0.163 a | ||
| M0 | 68.767 ± 1.474 a | 16.913 ± 0.395 | 8.737 ± 0.756 | 3.710 ± 0.329 b | ||
| Muscle | Male | M12 | 77.830 ± 0.469 a | 19.180 ± 0.301 b | 1.327 ± 0.135 a | 1.163 ± 0.040 |
| M9 | 77.390 ± 0.442 ab | 19.177 ± 0.095 b | 1.603 ± 0.083 ab | 1.187 ± 0.035 | ||
| M6 | 77.020 ± 0.337 ab | 19.360 ± 0.340 ab | 1.660 ± 0.400 ab | 1.207 ± 0.031 | ||
| M3 | 76.880 ± 0.347 a | 19.497 ± 0.352 ab | 1.913 ± 0.225 b | 1.170 ± 0.061 | ||
| M0 | 77.023 ± 0.737 ab | 19.763 ± 0.413 a | 1.657 ± 0.217 ab | 1.173 ± 0.049 | ||
| Female | M12 | 76.817 ± 1.453 | 19.903 ± 1.209 | 1.590 ± 0.235 | 1.28 ± 0.026 a | |
| M9 | 77.617 ± 0.629 | 19.317 ± 0.520 | 1.350 ± 0.263 | 1.200 ± 0.036 b | ||
| M6 | 76.427 ± 0.344 | 19.730 ± 0.414 | 1.803 ± 0.497 | 1.170 ± 0.010 b | ||
| M3 | 76.733 ± 0.674 | 20.020 ± 0.291 | 1.677 ± 0.613 | 1.200 ± 0.046 b | ||
| M0 | 77.133 ± 0.733 | 19.783 ± 0.257 | 1.467 ± 0.156 | 1.193 ± 0.029 b | ||
| Liver | Male | M12 | 71.007 ± 1.402 | 9.090 ± 0.423 b | 1.043 ± 0.294 b | 0.897 ± 0.057 b |
| M9 | 72.150 ± 1.459 | 10.227 ± 0.388 a | 1.207 ± 0.468 a | 0.973 ± 0.081 bc | ||
| M6 | 70.903 ± 0.976 | 9.753 ± 0.559 ab | 1.833 ± 0.195 ab | 1.093 ± 0.012 a | ||
| M3 | 70.913 ± 1.877 | 9.750 ± 0.509 ab | 1.637 ± 0.205 ab | 1.120 ± 0.046 a | ||
| M0 | 72.027 ± 1.868 | 9.800 ± 0.770 ab | 1.273 ± 0.482 ab | 1.080 ± 0.095 ac | ||
| Female | M12 | 71.290 ± 1.365 | 9.417 ± 0.545 b | 1.387 ± 0.369 | 0.993 ± 0.110 | |
| M9 | 71.820 ± 2.036 | 10.417 ± 0.829 ab | 1.603 ± 0.366 | 1.080 ± 0.113 | ||
| M6 | 70.353 ± 0.946 | 9.473 ± 0.386 ab | 1.817 ± 0.042 | 1.163 ± 0.104 | ||
| M3 | 70.267 ± 0.325 | 10.160 ± 0.618 ab | 1.817 ± 0.320 | 1.220 ± 0.227 | ||
| M0 | 71.463 ± 1.629 | 10.577 ± 0.300 a | 1.520 ± 0.195 | 1.227 ± 0.032 |
| Trypsin (U/mgprot) | Lipase (U/gprot) | Amylase (U/mgprot) | |
|---|---|---|---|
| M12 | 68.523 ± 4.412 | 87.950 ± 6.642 | 0.300 ± 0.060 a |
| M9 | 62.203 ± 6.487 | 80.803 ± 3.413 | 0.240 ± 0.050 ab |
| M6 | 62.223 ± 2.480 | 78.920 ± 17.403 | 0.203 ± 0.015 b |
| M3 | 69.497 ± 7.191 | 99.270 ± 29.941 | 0.183 ± 0.067 b |
| M0 | 70.010 ± 6.900 | 91.013 ± 18.132 | 0.223 ± 0.042 ab |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, L.; Guo, J.; Li, Q.; Jiang, J.; Chen, J.; Gao, L.; Yang, B.; Peng, J. Effects of High Dietary Starch Levels on the Growth Performance, Liver Function, and Metabolome of Largemouth Bass (Micropterus salmoides). Fishes 2024, 9, 256. https://doi.org/10.3390/fishes9070256
Sun L, Guo J, Li Q, Jiang J, Chen J, Gao L, Yang B, Peng J. Effects of High Dietary Starch Levels on the Growth Performance, Liver Function, and Metabolome of Largemouth Bass (Micropterus salmoides). Fishes. 2024; 9(7):256. https://doi.org/10.3390/fishes9070256
Chicago/Turabian StyleSun, Lihui, Jianlin Guo, Qian Li, Jianhu Jiang, Jianming Chen, Lingmei Gao, Bicheng Yang, and Jun Peng. 2024. "Effects of High Dietary Starch Levels on the Growth Performance, Liver Function, and Metabolome of Largemouth Bass (Micropterus salmoides)" Fishes 9, no. 7: 256. https://doi.org/10.3390/fishes9070256
APA StyleSun, L., Guo, J., Li, Q., Jiang, J., Chen, J., Gao, L., Yang, B., & Peng, J. (2024). Effects of High Dietary Starch Levels on the Growth Performance, Liver Function, and Metabolome of Largemouth Bass (Micropterus salmoides). Fishes, 9(7), 256. https://doi.org/10.3390/fishes9070256