
Feeding Habits and Prey Composition of Six Mesopelagic Fish Species from an Isolated Central Mediterranean Basin

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
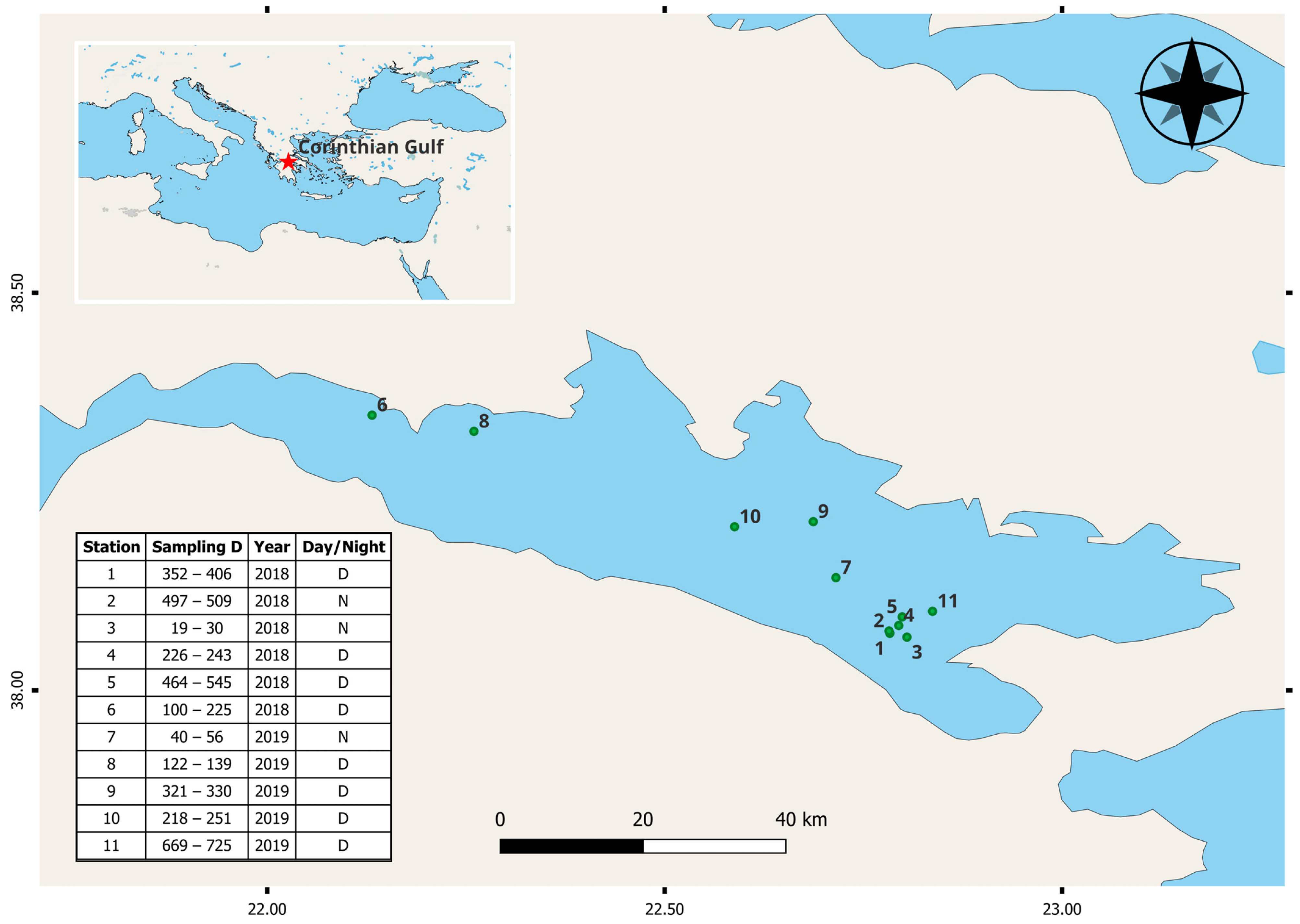
2.1. Study Area and Sample Collection
2.2. Laboratory and Data Analysis
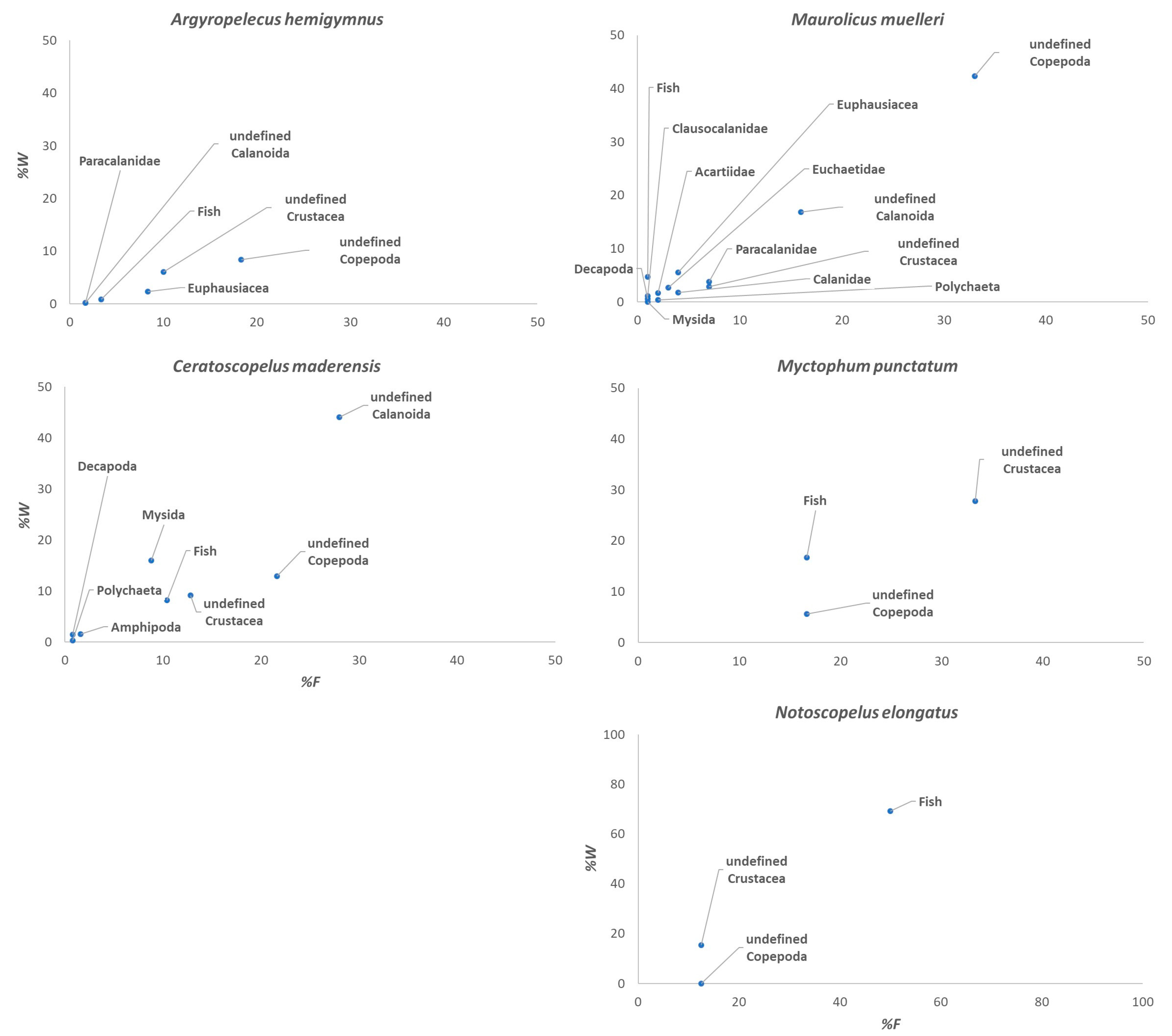
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Catul, V.; Gauns, M.; Karuppasamy, P.K. A Review on Mesopelagic Fishes Belonging to Family Myctophidae. Rev. Fish Biol. Fish. 2011, 21, 339–354. [Google Scholar] [CrossRef]
- Irigoien, X.; Klevjer, T.A.; Røstad, A.; Martinez, U.; Boyra, G.; Acuña, J.L.; Bode, A.; Echevarria, F.; Gonzalez-Gordillo, J.I.; Hernandez-Leon, S.; et al. Large Mesopelagic Fishes Biomass and Trophic Efficiency in the Open Ocean. Nat. Commun. 2014, 5, 3271. [Google Scholar] [CrossRef] [PubMed]
- Gjoesaeter, J.; Kawaguchi, K. A Review of the World Resources of Mesopelagic Fish; FAO: Rome, Italy, 1980; Volume 193. [Google Scholar]
- Bernal, A.; Olivar, M.P.; Maynou, F.; Fernández de Puelles, M.L. Diet and Feeding Strategies of Mesopelagic Fishes in the Western Mediterranean. Prog. Oceanogr. 2015, 135, 1–17. [Google Scholar] [CrossRef]
- Watanabe, H.; Kawaguchi, K.; Hayashi, A. Feeding Habits of Juvenile Surface-Migratory Myctophid Fishes (Family Myctophidae) in the Kuroshio Region of the Western North Pacific. Mar. Ecol. Prog. Ser. 2002, 236, 263–272. [Google Scholar] [CrossRef]
- Kaartvedt, S.; Langbehn, T.J.; Aksnes, D.L. Enlightening the Ocean’s Twilight Zone. ICES J. Mar. Sci. 2019, 76, 803–812. [Google Scholar] [CrossRef]
- Hulley, P.A. Myctophidae. In Check-List of the Fishes of the Eastern Tropical Atlantic; Quero, J.C., Hureau, J.C., Karrer, C., Post, A., Saldanha, L., Eds.; UNESCO: Paris, France, 1990; Volume 1, pp. 398–467. [Google Scholar]
- Valls, M.; Olivar, M.P.; Fernández de Puelles, M.L.; Molí, B.; Bernal, A.; Sweeting, C.J. Trophic Structure of Mesopelagic Fishes in the Western Mediterranean Based on Stable Isotopes of Carbon and Nitrogen. J. Mar. Syst. 2014, 138, 160–170. [Google Scholar] [CrossRef]
- Modica, L.; Cartes, J.E.; Velasco, F.; Bozzano, A. Juvenile Hake Predation on Myctophidae and Sternoptychidae: Quantifying an Energy Transfer between Mesopelagic and Neritic Communities. J. Sea Res. 2015, 95, 217–225. [Google Scholar] [CrossRef]
- D’Iglio, C.; Porcino, N.; Savoca, S.; Profeta, A.; Perdichizzi, A.; Armeli Minicante, E.; Salvati, D.; Soraci, F.; Rinelli, P.; Giordano, D. Ontogenetic Shift and Feeding Habits of the European Hake (Merluccius Merluccius L., 1758) in Central and Southern Tyrrhenian Sea (Western Mediterranean Sea): A Comparison between Past and Present Data. Ecol. Evol. 2022, 12, e8634. [Google Scholar] [CrossRef] [PubMed]
- Walkrr, M.G.; Nichols, J.H. Predation on Benthosema Glaciale (Myctouhidae) by Spawning Mackerel (Scomber Scombrus). J. Fish Biol. 1993, 42, 618–620. [Google Scholar] [CrossRef]
- Pusineri, C.; Vasseur, Y.; Hassani, S.; Meynier, L.; Spitz, J.; Ridoux, V. Food and Feeding Ecology of Juvenile Albacore, Thunnus Alalunga, off the Bay of Biscay: A Case Study. ICES J. Mar. Sci. 2005, 62, 116–122. [Google Scholar] [CrossRef]
- Polunin, A.; Morales-Nin, B.; Pawsey, W.; Cartes, J.; Pinnegar, J.; Moranta, J. Feeding Relationships in Mediterranean Bathyal Assemblages Elucidated by Carbon and Nitrogen Stable-Isotope Data. Mar. Ecol. Prog. Ser. 2001, 220, 13–23. [Google Scholar] [CrossRef]
- Papadimitraki, M.; Maar, K.; Jónasdóttir, S.H. Meso- and Bathypelagic Fish Feeding Ecology: A Meta-Analysis on Fatty Acids and Stable Isotope Trophic Studies. Deep Sea Res. Part Oceanogr. Res. Pap. 2023, 198, 104083. [Google Scholar] [CrossRef]
- Stergiou, I.K.; Karpouzi, V.S. Feeding Habits and Trophic Levels of Mediterranean Fish. Rev. Fish Biol. Fish. 2002, 11, 217–254. [Google Scholar] [CrossRef]
- Karachle, P.K.; Stergiou, K.I. An Update on the Feeding Habits of Fish in the Mediterranean Sea (2002–2015). Mediterr. Mar. Sci. 2017, 18, 43–52. [Google Scholar] [CrossRef]
- Battaglia, P.; Andaloro, F.; Esposito, V.; Granata, A.; Guglielmo, L.; Guglielmo, R.; Musolino, S.; Romeo, T.; Zagami, G. Diet and Trophic Ecology of the Lanternfish Electrona Risso (Cocco 1829) in the Strait of Messina (Central Mediterranean Sea) and Potential Resource Utilization from the Deep Scattering Layer (DSL). J. Mar. Syst. 2016, 159, 100–108. [Google Scholar] [CrossRef]
- Bernal, A.; Tuset, V.M.; Olivar, M.P. Multiple Approaches to the Trophic Role of Mesopelagic Fish around the Iberian Peninsula. Animals 2023, 13, 886. [Google Scholar] [CrossRef] [PubMed]
- Kapelonis, Z.; Siapatis, A.; Machias, A.; Somarakis, S.; Markakis, K.; Giannoulaki, M.; Badouvas, N.; Tsagarakis, K. Seasonal Patterns in the Mesopelagic Fish Community and Associated Deep Scattering Layers of an Enclosed Deep Basin. Sci. Rep. 2023, 13, 17890. [Google Scholar] [CrossRef] [PubMed]
- Methot, R.D. Frame Trawl for Sampling Pelagic Juvenile Fish. CalCOFI Rep. 1986, 27, 267–278. [Google Scholar]
- Trégouboff, G.; Rose, M. Manuel de Planctonologie Méditerranéenne (Tome I & II); Centre National de la Recherche Scientifique: Paris, France, 1957. [Google Scholar]
- Hyslop, E.J. Stomach Contents Analysis—A Review of Methods and Their Application. J. Fish Biol. 1980, 17, 411–429. [Google Scholar] [CrossRef]
- Amundsen, P.-A.; Sánchez-Hernández, J. Feeding Studies Take Guts—Critical Review and Recommendations of Methods for Stomach Contents Analysis in Fish. J. Fish Biol. 2019, 95, 1364–1373. [Google Scholar] [CrossRef]
- Windell, J.T. Food Analysis and Rate of Digestion. In Methods for Assessment of Fish Production in Fresh Waters, 2nd ed.; Ricker, W.E., Ed.; Blackwell Scientific Publications: Oxford, UK, 1971; pp. 215–226. [Google Scholar]
- Pauly, D.; Froese, R.; Saa, P.; Palomares, M.L.; Christensen, V.; Rius, J. Trophlab Manual; ICLARM: Manila, Philippines, 2000. [Google Scholar]
- Christensen, V.; Pauly, D. ECOPATH II—A Software for Balancing Steady-State Ecosystem Models and Calculating Network Characteristics. Ecol. Model. 1992, 61, 169–185. [Google Scholar] [CrossRef]
- Shannon, C.; Weaver, W. The Mathematical Theory of Communication; University of illinois Press: Champaign, IL, USA, 1949. [Google Scholar]
- Iglesias-Rios, R.; Mazzoni, R. Measuring Diversity: Looking for Processes That Generate Diversity. Nat. Conserv. 2014, 12, 156–161. [Google Scholar] [CrossRef]
- Levins, R. Evolution in Changing Environments: Some Theoretical Explorations. (MPB-2); Princeton University Press: Princeton, NJ, USA, 1968; ISBN 978-0-691-07959-2. [Google Scholar]
- Feinsinger, P.; Spears, E.E.; Poole, R.W. A Simple Measure of Niche Breadth. Ecology 1981, 62, 27–32. [Google Scholar] [CrossRef]
- Schoener, T.W. Nonsynchronous Spatial Overlap of Lizards in Patchy Habitats. Ecology 1970, 51, 408–418. [Google Scholar] [CrossRef]
- Wallace, R.K., Jr. An Assessment of Diet-Overlap Indexes. Trans. Am. Fish. Soc. 1981, 110, 72–76. [Google Scholar] [CrossRef]
- Wallace, R.K.; Ramsey, J.S. Reliability in Measuring Diet Overlap. Can. J. Fish. Aquat. Sci. 1983, 40, 347–351. [Google Scholar] [CrossRef]
- Zaret, T.M.; Rand, A.S. Competition in Tropical Stream Fishes: Support for the Competitive Exclusion Principle. Ecology 1971, 52, 336–342. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. Primer V6: User Manual—Tutorial; PRIMER-E Limited: Auckland, New Zealand, 2006. [Google Scholar]
- Costello, M.J. Predator Feeding Strategy and Prey Importance: A New Graphical Analysis. J. Fish Biol. 1990, 36, 261–263. [Google Scholar] [CrossRef]
- Badouvas, N.; Somarakis, S.; Tsagarakis, K. Length–Weight Relations of 16 Mesopelagic Fishes (Actinopterygii: Myctophiformes and Stomiiformes) from the Eastern Mediterranean Sea. Acta Ichthyol. Piscat. 2022, 52, 279–283. [Google Scholar] [CrossRef]
- Watanabe, H.; Moku, M.; Kawaguchi, K.; Ishimaru, K.; Ohno, A. Diel Vertical Migration of Myctophid Fishes (Family Myctophidae) in the Transitional Waters of the Western North Pacific. Fish. Oceanogr. 1999, 8, 115–127. [Google Scholar] [CrossRef]
- Aksnes, D.L.; Løtvedt, A.S.; Lindemann, C.; Calleja, M.L.; Morán, X.A.G.; Kaarvedt, S.; Thingstad, T.F. Effects of Migrating Mesopelagic Fishes on the Biological Carbon Pump. Mar. Ecol. Prog. Ser. 2023, 717, 107–126. [Google Scholar] [CrossRef]
- Olivar, M.P.; Castellón, A.; Sabatés, A.; Sarmiento-Lezcano, A.; Emelianov, M.; Bernal, A.; Yang, Y.; Proud, R.; Brierley, A.S. Variation in Mesopelagic Fish Community Composition and Structure between Mediterranean and Atlantic Waters around the Iberian Peninsula. Front. Mar. Sci. 2022, 9, 1028717. [Google Scholar]
- Hopkins, T.L.; Baird, R.C. Feeding Ecology of Four Hatchetfishes (Sternoptychidae) in the Eastern Gulf of Mexico. Bull. Mar. Sci. 1985, 36, 260–277. [Google Scholar]
- Merrett, N.R.; Roe, H.S.J. Patterns and Selectivity in the Feeding of Certain Mesopelagic Fishes. Mar. Biol. 1974, 28, 115–126. [Google Scholar] [CrossRef]
- Davison, P.; Asch, R. Plastic Ingestion by Mesopelagic Fishes in the North Pacific Subtropical Gyre. Mar. Ecol. Prog. Ser. 2011, 432, 173–180. [Google Scholar] [CrossRef]
- Rochman, C.M.; Lewison, R.L.; Eriksen, M.; Allen, H.; Cook, A.-M.; Teh, S.J. Polybrominated Diphenyl Ethers (PBDEs) in Fish Tissue May Be an Indicator of Plastic Contamination in Marine Habitats. Sci. Total Environ. 2014, 476–477, 622–633. [Google Scholar] [CrossRef]
- Wieczorek, A.M.; Morrison, L.; Croot, P.L.; Allcock, A.L.; MacLoughlin, E.; Savard, O.; Brownlow, H.; Doyle, T.K. Frequency of Microplastics in Mesopelagic Fishes from the Northwest Atlantic. Front. Mar. Sci. 2018, 5, 339138. [Google Scholar]
- Torre, M.; Digka, N.; Anastasopoulou, A.; Tsangaris, C.; Mytilineou, C. Anthropogenic Microfibres Pollution in Marine Biota. A New and Simple Methodology to Minimize Airborne Contamination. Mar. Pollut. Bull. 2016, 113, 55–61. [Google Scholar] [CrossRef]
- Andersen, V.; Gubanova, A.; Nival, P.; Ruellet, T. Zooplankton Community During the Transition from Spring Bloom to Oligotrophy in the Open NW Mediterranean and Effects of Wind Events. 2. Vertical Distributions and Migrations. J. Plankton Res. 2001, 23, 243–261. [Google Scholar] [CrossRef]
- Scotto di Carlo, B.; Ianora, A.; Fresi, E.; Hure, J. Vertical Zonation Patterns for Mediterranean Copepods from the Surface to 3000 m at a Fixed Station in the Tyrrhenian Sea. J. Plankton Res. 1984, 6, 1031–1056. [Google Scholar] [CrossRef]
- Brugnano, C.; Bergamasco, A.; Granata, A.; Guglielmo, L.; Zagami, G. Spatial Distribution and Community Structure of Copepods in a Central Mediterranean Key Region (Egadi Islands—Sicily Channel). J. Mar. Syst. 2010, 81, 312–322. [Google Scholar] [CrossRef]
- Staby, A.; Aksnes, D.L. Follow the Light—Diurnal and Seasonal Variations in Vertical Distribution of the Mesopelagic Fish Maurolicus Muelleri. Mar. Ecol. Prog. Ser. 2011, 422, 265–273. [Google Scholar] [CrossRef]
- Christiansen, S.; Titelman, J.; Kaartvedt, S. Nighttime Swimming Behavior of a Mesopelagic Fish. Front. Mar. Sci. 2019, 6, 787. [Google Scholar] [CrossRef]
- Kristoffersen, J.B.; Salvanes, A.G.V. Life History of Maurolicus Muelleri in Fjordic and Oceanic Environments. J. Fish Biol. 1998, 53, 1324–1341. [Google Scholar] [CrossRef]
- Kaartvedt, S.; Christiansen, S.; Røstad, A.; Aksnes, D. Mesopelagic Fishes in a Hurry at Low Latitudes. Mar. Ecol. Prog. Ser. 2022, 694, 149–156. [Google Scholar] [CrossRef]
- Staby, A.; Srisomwong, J.; Rosland, R. Variation in DVM Behaviour of Juvenile and Adult Pearlside (Maurolicus Muelleri) Linked to Feeding Strategies and Related Predation Risk. Fish. Oceanogr. 2013, 22, 90–101. [Google Scholar] [CrossRef]
- Prihartato, P.K.; Aksnes, D.L.; Kaartvedt, S. Seasonal Patterns in the Nocturnal Distribution and Behavior of the Mesopelagic Fish Maurolicus Muelleri at High Latitudes. Mar. Ecol. Prog. Ser. 2015, 521, 189–200. [Google Scholar] [CrossRef]
- Hulley, P.A. Myctophidae. In Fishes of the North-Eastern Atlantic and the Mediterranean; Whitehead, P.J.P., Bauchot, M.L., Hureau, J.C., Nielsen, J., Tortonese, E., Eds.; UNESCO: Paris, France, 1984; Volume 1. [Google Scholar]
- Froese, R.J.; Pauly, D. Fishbase. World Wide Web Electronic Publication. Available online: http://www.fishbase.org (accessed on 1 February 2024).
- Kinzer, J. Observation on Feeding Habit of Mesopelagic Fish Benthosema Glaciale (Myctophidae) off NW Africa. In Oceanic Sound Scattering Prediction; Zahuranec, B.J., Andersen, N.R., Eds.; Plenum Press: New York, NY, USA, 1977; Volume 5, pp. 381–392. [Google Scholar]
- Sameoto, D.D. Feeding of Lantern Fish Benthosema Glaciale off the Nova Scotia Shelf. Mar. Ecol. Prog. Ser. 1988, 44, 113–129. [Google Scholar] [CrossRef]
- Contreras, T.; Olivar, M.P.; Bernal, A.; Sabatés, A. Comparative Feeding Patterns of Early Stages of Mesopelagic Fishes with Vertical Habitat Partitioning. Mar. Biol. 2015, 162, 2265–2277. [Google Scholar] [CrossRef]
- Voss, R.; Dickmann, M.; Schmidt, J.O. Feeding Ecology of Sprat (Sprattus sprattus L.) and Sardine (Sardina pilchardus W.) Larvae in the German Bight, North Sea. Oceanologia 2009, 51, 117–138. [Google Scholar] [CrossRef]
- Eduardo, L.N.; Lucena-Frédou, F.; Mincarone, M.M.; Soares, A.; Le Loc’h, F.; Frédou, T.; Ménard, F.; Bertrand, A. Trophic Ecology, Habitat, and Migratory Behaviour of the Viperfish Chauliodus Sloani Reveal a Key Mesopelagic Player. Sci. Rep. 2020, 10, 20996. [Google Scholar] [CrossRef] [PubMed]
- Gartner, J.V.; Crabtree, R.E.; Sulak, K.J. Feeding At Depth. In Fish Physiology; Randall, D.J., Farrell, A.P., Eds.; Deep-Sea Fishes; Academic Press: Cambridge, MA, USA, 1997; Volume 16, pp. 115–193. [Google Scholar]
- Battaglia, P.; Andaloro, F.; Consoli, P.; Esposito, V.; Malara, D.; Musolino, S.; Pedà, C.; Romeo, T. Feeding Habits of the Atlantic Bluefin Tuna, Thunnus thynnus (L. 1758), in the Central Mediterranean Sea (Strait of Messina). Helgol. Mar. Res. 2013, 67, 97–107. [Google Scholar] [CrossRef]
- Giménez, J.; Marçalo, A.; García-Polo, M.; García-Barón, I.; Castillo, J.J.; Fernández-Maldonado, C.; Saavedra, C.; Santos, M.B.; de Stephanis, R. Feeding Ecology of Mediterranean Common Dolphins: The Importance of Mesopelagic Fish in the Diet of an Endangered Subpopulation. Mar. Mammal Sci. 2018, 34, 136–154. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Argyropelecus hemigymnus | Benthosema glaciale | Ceratoscopelus maderensis | Maurolicus muelleri | Myctophum punctatum | Notoscopelus elongatus |
|---|---|---|---|---|---|---|
| N | 79 | 16 | 210 | 92 | 14 | 26 |
| TL range | 22–44 | 30–65 | 32–79 | 24–52 | 56–74 | 68–122 |
| Mean TL (±SD) | 32.0 ± 4.6 | 44.6 ± 9.8 | 57.0 ± 7.5 | 36.6 ± 8.6 | 62.1 ± 6.4 | 80.6 ± 13.8 |
| %VI | 41.77% | 56.25% | 55.71% | 17.39% | 66.27% | 73.08% |
| Prey item N | 88 | 15 | 1591 | 661 | 14 | 13 |
| TROPH ± SE | 2.6 ± 0.46 | 2.73 ± 0.44 | 3.14 ± 0.43 | 3.01 ± 0.29 | 3.03 ± 0.51 | 3.98 ± 0.7 |
| OI | 0.212 | 0.194 | 0.185 | 0.084 | 0.260 | 0.490 |
| H’ | 1.304 | 0.683 | 1.830 | 1.572 | 1.33 | 1.213 |
| Bn | 0.282 | 0.196 | 0.545 | 0.386 | 0.36 | 0.291 |
| Argyropelecus hemigymnus | Benthosema glaciale | Ceratoscopelus maderensis | Maurolicus muelleri | Myctophum punctatum | Notoscopelus elongatus | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Prey Items | %N | %F | %W | %IRI | %N | %F | %W | %IRI | %N | %F | %W | %IRI | %N | %F | %W | %IRI | %N | %F | %W | %IRI | %N | %F | %W | %IRI |
| Crustacea (Total) | 90.91 | 40.00 | 17.12 | 53.33 | 57.14 | 37.50 | 99.56 | 73.60 | 85.10 | 99.09 | 79.00 | 78.88 | 50.00 | 50.00 | 33.33 | 23.08 | 25.00 | 15.38 | ||||||
| Unidentified Crustacea | 10.23 | 10.00 | 6.05 | 9.31 | 53.33 | 57.14 | 37.50 | 100.00 | 0.88 | 12.80 | 9.13 | 2.86 | 1.66 | 7.00 | 2.81 | 0.77 | 28.57 | 33.33 | 27.78 | 66.05 | 7.69 | 12.50 | 15.38 | 4.11 |
| Copepoda (Total) | 75.00 | 21.67 | 8.77 | 96.98 | 49.60 | 56.97 | 96.52 | 66.00 | 70.05 | 21.43 | 16.67 | 5.56 | 15.38 | 12.50 | ||||||||||
| Unidentified Copepoda | 73.86 | 18.33 | 8.35 | 86.15 | 19.74 | 21.60 | 12.86 | 15.73 | 53.71 | 33.00 | 42.38 | 78.50 | 21.43 | 16.67 | 5.56 | 15.81 | 15.38 | 12.50 | 2.74 | |||||
| Calanoida (Total) | 1.14 | 3.33 | 0.42 | 77.25 | 28.00 | 44.11 | 42.81 | 33.00 | 27.67 | |||||||||||||||
| Unidentified Calanoida | 1.67 | 0.21 | 0.02 | 77.25 | 28.00 | 44.11 | 75.92 | 22.39 | 16.00 | 16.84 | 15.54 | |||||||||||||
| Acartiidae | 0.76 | 2.00 | 1.60 | 0.12 | ||||||||||||||||||||
| Calanidae (Total) | 8.47 | 4.00 | 1.74 | |||||||||||||||||||||
| Unidentified Calanidae | 3.48 | 1.00 | 1.07 | 0.11 | ||||||||||||||||||||
| Calanus sp. (Total) | 4.99 | 3.00 | 66.84 | |||||||||||||||||||||
| Unidentified Calanus sp. | 0.91 | 1.00 | 0.02 | |||||||||||||||||||||
| Calanus helgolandicus | 4.08 | 2.00 | 0.67 | 0.24 | ||||||||||||||||||||
| Euchaetidae (Total) | 0.61 | 3.00 | 2.67 | |||||||||||||||||||||
| Unidentified Euchaetidae | 0.45 | 2.00 | 2.14 | 0.13 | ||||||||||||||||||||
| Euchaeta marina | 0.15 | 1.00 | 53.48 | 1.33 | ||||||||||||||||||||
| Clausocalanidae | 0.30 | 1.00 | 1.07 | |||||||||||||||||||||
| Ctenocalanus vanus | 0.30 | 1.00 | 1.07 | 0.03 | ||||||||||||||||||||
| Paracalanidae | 1.14 | 1.67 | 0.21 | 10.29 | 7.00 | 3.74 | ||||||||||||||||||
| Paracalanus parvus | 1.14 | 1.67 | 0.21 | 0.13 | 10.29 | 7.00 | 3.74 | 2.43 | ||||||||||||||||
| Malacostraca | 5.68 | 8.33 | 2.30 | 1.70 | 11.20 | 18.99 | 0.91 | 6.00 | 6.02 | |||||||||||||||
| Amphipoda | 0.19 | 1.60 | 1.56 | 0.06 | ||||||||||||||||||||
| Decapoda (Total) | 0.06 | 0.80 | 1.44 | 0.15 | 1.00 | 0.53 | ||||||||||||||||||
| Decapoda (larva) | 0.06 | 0.80 | 1.44 | 0.03 | ||||||||||||||||||||
| Lucifer typus | 0.15 | 1.00 | 0.53 | 0.02 | ||||||||||||||||||||
| Euphausiacea | 5.68 | 8.33 | 2.30 | 3.80 | 0.30 | 4.00 | 5.48 | 0.57 | ||||||||||||||||
| Mysida | 1.45 | 8.80 | 15.99 | 3.43 | 0.45 | 1.00 | 0.01 | |||||||||||||||||
| Fish | 2.27 | 3.33 | 0.84 | 0.59 | 0.25 | 10.40 | 8.17 | 1.96 | 0.15 | 1.00 | 4.68 | 0.12 | 14.29 | 16.67 | 16.67 | 18.14 | 61.54 | 50.00 | 69.23 | 93.15 | ||||
| Polychaeta | 0.00 | 0.80 | 0.24 | 0.00 | 0.76 | 2.00 | 0.40 | 0.06 | ||||||||||||||||
| Particulate organic matter | 5.68 | 55.00 | 79.54 | 46.67 | 42.86 | 62.50 | 0.19 | 15.20 | 6.49 | 18.00 | 16.04 | 35.71 | 33.33 | 50.00 | 15.38 | 25.00 | 15.38 | |||||||
| VI% | D | N |
|---|---|---|
| Argyropelecus hemigymnus | 41.03% (78) | 100% (1) |
| Benthosema glaciale | 100% (4) | 41.67% (12) |
| Ceratoscopelus maderensis | 76.83% (82) | 42.19% (128) |
| Maurolicus muelleri | 17.39% (92) | - (0) |
| Myctophum punctatum | 75.00% (12) | 0.00% (2) |
| Notoscopelus elongatus | 76.00% (25) | 0.00% (1) |
| Species | Av. Similarity (%) |
|---|---|
| Argyropelecus hemigymnus | 28.30 |
| Benthosema glaciale | 100 |
| Ceratoscopelus maderensis | 33.75 |
| Maurolicus muelleri | 35.33 |
| Myctophum punctatum | 33.33 |
| Notoscopelus elongatus | 33.55 |
| Species Groups | Key Prey Taxon Contribution (%) |
|---|---|
| A. hemigymnus—B. glaciale | Unidentified Crustacea: 41.04 |
| A. hemigymnus—C. maderensis | Calanoida: 35.35 |
| A. hemigymnus—M. muelleri | Unidentified Copepoda: 34.76 |
| A. hemigymnus—M. punctatum | Unidentified Copepoda: 34.76 |
| A. hemigymnus—N. elongatus | Fish: 36.78 |
| B. glaciale—C. maderensis | Calanoida: 35.12 |
| B. glaciale—M. muelleri | Unidentified Crustacea: 38.73 |
| B. glaciale—M. punctatum | Unidentified Copepoda: 35.62 |
| B. glaciale—N. elongatus | Fish: 45.29 |
| C. maderensis—M. muelleri | Calanoida: 41.93 |
| C. maderensis—M. punctatum | Calanoida: 34.00 |
| C. maderensis—N. elongatus | Calanoida: 32.76 |
| M. muelleri—M. punctatum | Unidentified Copepoda: 29.96 |
| M. muelleri—N. elongatus | Fish: 30.61 |
| M. punctatum—N. elongatus | Fish: 41.12 |
| Shoener’s S Index | Benthosema glaciale | Ceratoscopelus maderensis | Maurolicus muelleri | Myctophum punctatum | Notoscopelus elongatus |
|---|---|---|---|---|---|
| Argyropelecus hemigymnus | 0.783 | 0.599 | 0.638 | 0.785 | 0.556 |
| Benthosema glaciale | 0.436 | 0.421 | 0.840 | 0.543 | |
| Ceratoscopelus maderensis | 0.601 | 0.511 | 0.283 | ||
| Maurolicus muelleri | 0.461 | 0.232 | |||
| Myctophum punctatum | 0.647 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Badouvas, N.; Tsagarakis, K.; Somarakis, S.; Karachle, P.K. Feeding Habits and Prey Composition of Six Mesopelagic Fish Species from an Isolated Central Mediterranean Basin. Fishes 2024, 9, 277. https://doi.org/10.3390/fishes9070277
Badouvas N, Tsagarakis K, Somarakis S, Karachle PK. Feeding Habits and Prey Composition of Six Mesopelagic Fish Species from an Isolated Central Mediterranean Basin. Fishes. 2024; 9(7):277. https://doi.org/10.3390/fishes9070277
Chicago/Turabian StyleBadouvas, Nicholas, Konstantinos Tsagarakis, Stylianos Somarakis, and Paraskevi K. Karachle. 2024. "Feeding Habits and Prey Composition of Six Mesopelagic Fish Species from an Isolated Central Mediterranean Basin" Fishes 9, no. 7: 277. https://doi.org/10.3390/fishes9070277