

Anti-Aging Potential of Bioactive Phytoconstituents Found in Edible Medicinal Plants: A Review

 , , and
, , and
Abstract
1. Introduction
2. Materials Used for the Study
3. Proposed Mechanism of Action of the Bioactive Compounds
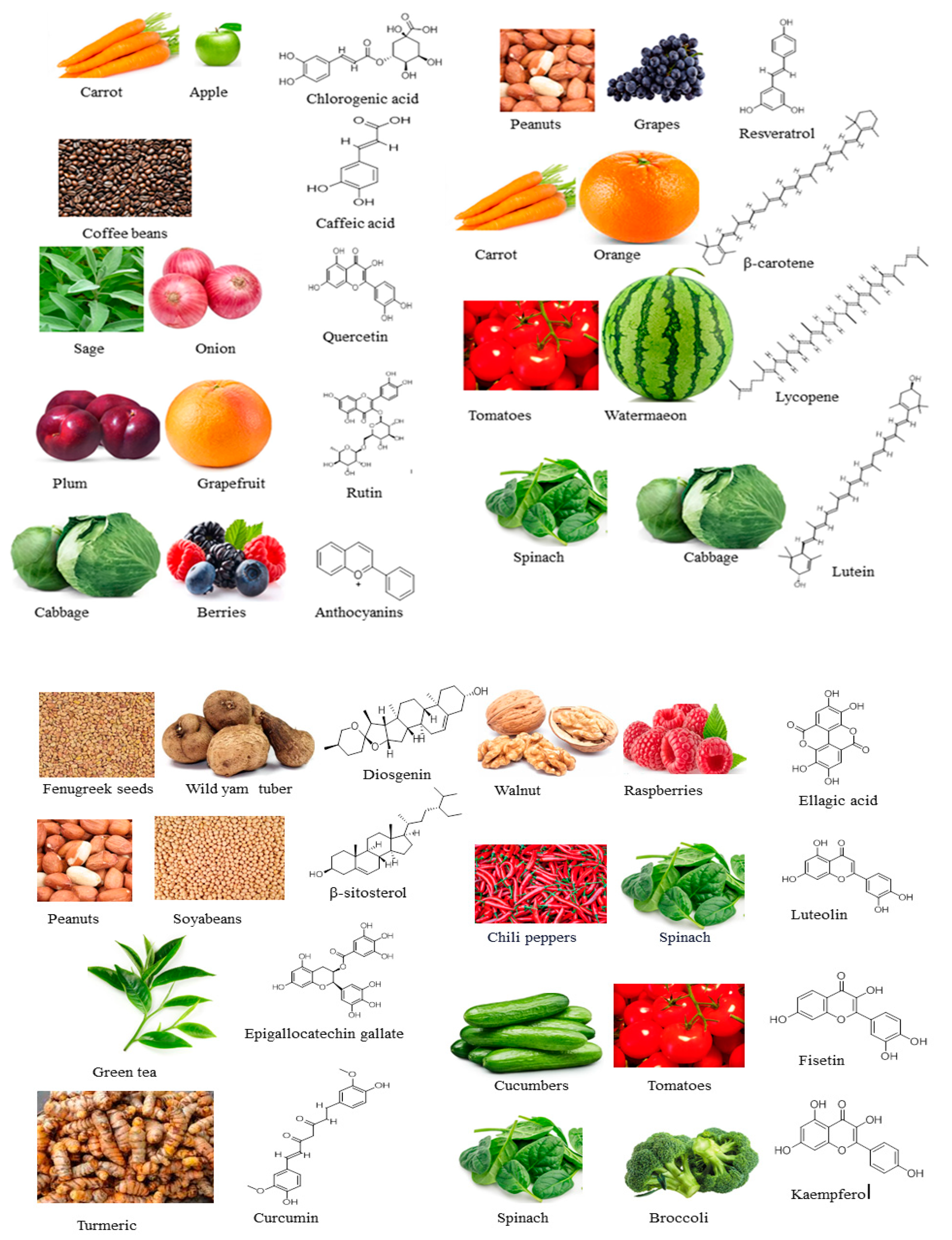
4. Bioactive Compounds
4.1. Chlorogenic Acid
4.2. Quercetin
4.3. Rutin
4.4. Anthocyanins
4.5. Resveratrol
4.6. Epigallocatechin Gallate
4.7. Caffeic Acid/Dihydrocaffeic Acid
4.8. Lycopene
4.9. Lutein
4.10. β-Sitosterol
4.11. Curcumin
4.12. Luteolin
4.13. Fisetin
4.14. Kaempferol
4.15. β-Carotene
4.16. Diosgenin
4.17. Ellagic Acid

{kind=link}
{kind=link}
| Bioactive Compound | Experimental Models | Condition | Effective Dose | Mechanism | References |
|---|---|---|---|---|---|
| Chlorogenic acid | Caenorhabditis elegans | Aging | 50 µM | Extension of adult mean lifespan by 20.1% through modulation of the insulin/insulin-like signaling pathway through the DAF-16 transcription factor. | [52] |
| Chlorogenic acid | Hep3B and HeLa cell lines | Aging | 20–40 µM | Increase in lifespan through the induction of SKN-1. | [54] |
| Chlorogenic acid | Yeast | Aging | 25 µM | Increase in chronological lifespan through the activation of antioxidative stress response, activation of SOD2 and SIR2 levels. | [56] |
| Chlorogenic acid | Mice, HUVECs | Vascular damage | 40 mg/kg | Suppression of angiotensin- and H2O2-induced senescence in a dose-dependent manner through increased expression of Sirt1 and eNOS. | [57] |
| Chlorogenic acid | 28 elderly individuals | Dementia | 330 mg | Significant improvement in verbal memory, attention, cognitive flexibility, motor speed and executive function. | [58] |
| Chlorogenic acid | 26 participants between 50–65 years | Dementia | 270 mg | Improvement of CNS vital signs such as psychomotor speed and motor speed. | [59] |
| Chlorogenic acid | 38 healthy participants | Subjective memory loss | 300 mg | Delayed cognitive decline and improved cognitive function. | [60] |
| Chlorogenic acid | UV-induced fibroblast cells | Increase in ROS levels due to UV exposure | 25 µg/mL | Increase in COL-3 gene expression. | [61] |
| Chlorogenic acid | Human fibroblast and keratinocytes | Exposure to ultraviolet rays | 20 µg/mL | Increase in Hyaluronic acid and collagen content and a decrease in MMP-1. | [64] |
| Chlorogenic acid | Mouse | Retinal pathogenic neovascularisation | 25–50 mg/kg/day | Significant reduction in retina vascular changes. | [66] |
| Quercetin | Simocephalus vetulus | Short lifespan | 1 mg/L | Increase in lifespan of 22%. | [73] |
| Quercetin | Human erythrocyte | Aging | 10 µM | Protection of membrane lipids from oxidative damage. | [74] |
| Quercetin | Caenorhabditis elegans | Aging | 0.15 mg/mL | Extension18.53% in lifespan by the regulation of the SIRT1 signaling pathway. | [76] |
| Quercetin | Pre-adipocytes and adipocyte cells | Aging | 20 µM | Reduction in beta galactosidase activity through the downregulation of miR-155-5p expression and upregulation of the SIRT-1 pathway. | [81] |
| Quercetin | Human skin tissues | UV-mediated aging | 20 µM | Suppressed expression of MMP-1 and the attenuation of inflammatory responses in the skin tissues due to a reduction in COX-2 expression | [82] |
| Rutin | Rats | Aging | 2% | Significant inhibition of increases in fasting blood glucose, insulin levels, and blood pressure. | [84] |
| Rutin | Human dermal fibroblast | Skin aging | 50 µM | Increase in dermal density and elasticity. | [88] |
| Rutin | Human prostrate stromal cell line | Senescence | 100 µM | Reduce the expression of senescence-associated secretory phenotype. | [89] |
| Rutin | Drosophila melanogaster | Aging | 100 µM | Extension of lifespan via mediating hormesis. | [90] |
| Rutin | Aged mice | Aging | 0.2 mg/mL | Lifespan extension of 10%. | [91] |
| Anthocyanins | Female Drosophila melanogaster | Aging | 10 mg/mL | Prolonged lifespan through the regulation of the autophagy pathway. | [96] |
| Anthocyanins | Male Drosophila melanogaster | Aging | 2.0 mg/mL | Prolonged lifespan through the regulation of the autophagy pathway. | [98] |
| Anthocyanins | Fruitflies | Aging | 20 mg/mL | Increase in lifespan of 10% through the upregulation of SOD gene and downregulation of the MTH, InR, TOR, Hep and PEPCK genes. | [99] |
| Anthocyanins | Caenorhabditis elegans | Aging | 100 µg/mL | Prevent oxidative damage through the activation of Nrf2 signaling pathway. | [103] |
| Anthocyanins | Caenorhabditis elegans | Aging | 150 µL | Upregulation of the SOD gene which resulted in a 56.25% increase in lifespan of the organism and was also associated with other factors, such as improved resistance to stress. | [104] |
| Anthocyanins | Retinal pigment epithelial cells | Aging- and light-induced damage | 10 µg/mL | Decrease in the percentage of senescent cells in the retinal pigment and also reduced intracellular ROS levels. | [105] |
| Resveratrol | SAMP8 mice model | Aging | 1 g/kg | Increased life expectancy through an increase in SIRT1 and AMPK levels. | [107] |
| Resveratrol | Bombyx mori | Aging | 500 µM | Lifespan extension through the activation of the SIRT7 pathway. | [111] |
| Resveratrol | Drosophila melanogaster | Aging | 60 mg/kg | Increase in lifespan and improvement of behavioral deficit due to oxidative damage | [112] |
| Resveratrol | Zebra fish retina | Oculopathy | 20 mg/mL | Restoration of mitochondrial integrity. | [115] |
| Resveratrol Rice | Drosophila melanogaster | Aging | 31.54 µg/L | Significant extension median lifespan. | [117] |
| Epigallocatechin gallate | Human serum albumin | Aging | 6 mg/kg | Neuroprotective potential by preventing the deposition of lipofuscin. | [121] |
| Epigallocatechin gallate | 3T3-L1 Preadipocyte cells | Senescence | 100 µM | Induce senescence through modulation of the pro- and anti-apoptotic pathways. | [122] |
| Epigallocatechin gallate | Drosophila melanogaster | Aging | 10 mg/mL | Significant increase in lifespan and increased survival rate. | [124] |
| Epigallocatechin gallate | Human keratinocyte cells | Aging | 20 µL | Improved anti-aging potential and protection against skin damage. | [128] |
| Caffeic acid | Drosophila melanogaster | Aging | 0.2 mg/mL | Increase in lifespan by delaying the onset of intestinal stem cells and progenitor cells. | [135] |
| Caffeic acid + APPPKK | Fibroblast cells | Skin aging | 100 µM | Reducing stress in cells. | [136] |
| Caffeic acid | Caenorhabditis elegans | Aging | 100 µM | Increase in mean lifespan. | |
| Caffeic acid | Caenorhabditis elegans | Aging | 200 µM | Increase in maximum lifespan. | [138] |
| Lycopene | Mice | Cognitive impairment | 50 mg/kg | Improvement in age-induced neuronal degeneration and MDA and BDNF expression. | [146] |
| Lycopene | Aged chicken | Ovarian aging | 100 ng/mL | Reduce oxidative stress via the activation of the Nrf2/HO-1 pathway. | [21] |
| Lycopene | Oocytes | Post-ovulatory aging | 200 nm | Reduce oxidative stress. | [150] |
| Blakeslea trispora (1.9% Lycopene) | Mice | Aging | 1068 mg/kg | Improved antioxidant activities. | [152] |
| Lutein | Drosophila melanogaster | Aging | 0.1 mg/mL | Increased maximum lifespan via a decrease in malonyldialdehyde levels and the upregulation of antioxidant enzymes. | [157] |
| Lutein | Women | Photoprotection | 20 mg/day | Increase in photoprotective activity marked by a significant increase in MED. | [158] |
| Lutein | ARPE-19 cells | Senescence | 1 µg/mL | Significant reduction in SA-b-gal activity. | [159] |
| Lutein | ARPE-19 cells | Senescence | 20 µM | Decrease in SA-b-gal activity. | [160] |
| Β-sitosterol | Male mice | Age-related muscle loss | 200 mg/kg | Protection from muscle loss and the restoration of grip strength. | [164] |
| Β-sitosterol + vermicularin | Human skin fibroblasts and keratinocytes | Skin aging | 50 µM | Increased expression of hyaluronic acid synthases. | [166] |
| Curcumin | Drosophila melanogaster | Aging | 250 µM | Extension of adult lifespan through the modulation of age-related genes such as mth, thor, INR, and INK | [170] |
| Curcumin | Larval feed | Senescence | 20 µM | Extend the minimum and maximum lifespan of the adult fly by increasing the development time and delaying the onset of senescence. | [171] |
| Curcumin | Caenorhabditis elegans | Aging | 20 µM | Extend the maximum adult lifespan by 17 days and delay the process of aging. | [172] |
| Curcumin | Human Umbilical Vein Endothelial Cells | Senescence | 25 µM | Reduction in hydrogen peroxide-induced apoptosis in the cells. | [173] |
| Curcumin | Albino Wistar rats | Aging | 400 mg/kg | Significant reduction in anti-aging inflammatory markers. | [175] |
| Curcumin | Saccharomyces cerevisiae | Aging | 200 µM | Extension of minimum and maximum lifespan. | [176] |
| Luteolin | Mouse oocytes | Postovulatory oocyte aging | 5 µM | Protection of oocytes from post-ovulatory aging due to the activation of the SIRT1 gene. | [179] |
| Luteolin | Immortalized nucleus pulposus cells | Cellular senescence | 4 µM | Regulation of the SIRT6 gene. | [182] |
| Fisetin | SAMP8 | Neurodegenerative decline | 25 mg/kg | Suppression of changes associated with aging through the alteration of specific brain and plasma metabolites. | [187] |
| Fisetin | Murine and fibroblast cells | Senescence | 5 µM | Suppression of senescent cells. | [188] |
| Fisetin | Vascular cells | Senescence | 1 µM | Reduction in SA-β gal activity. | [189] |
| Fisetin | Sheep | Senescence | 100 mg/kg | Reduction in senescent cells. | [190] |
| Fisetin | MII oocytes | Post-ovulatory oocyte aging | 10 µM | Elevation of SIRT1 expression levels. | [191] |
| Fisetin | Mouse | Premature Aging | 500 mg/kg | Significant upregulation of aging markers and reduction in collagen fiber deposition. | [192] |
| Fisetin | Caenorhabditis elegans | Aging | 0.1 g/L | Increase in both their mean and maximum lifespan. | [193] |
| Kaempferol | Aged rats | Aging | 4 mg/kg | Suppression of the NF-κB cascade through the modulation of nuclear factor-inducing kinase (NIK)/IκB kinase (IKK) and mitogen-activated protein kinases (MAPKs). | [197] |
| β-carotene | Mesenchymal stem cells | Senescence | 5 µM | Decrease the activity of SA-β-Gal, reduce the production of P21, P16, and P53, and lower the levels of pro-inflammatory factors like IL-1β, IL-6, and tumor necrosis factor-β (TNF-β). | [199] |
| Ellagic acid | Mice | Aging | 100 mg/kg | Reduction in ROS levels. | [204] |
| Ellagic acid | SH-SY-5Y | Aging | 10 µM | Significantly increase cell proliferation and GSH levels, while decreasing ROS, MDA, TNF-α, β−GAL, and AGEs levels. | [205] |
| Ellagic acid | Rats | Aging | 100 mg/kg | Improved cognitive impairment and hippocampal damage, increased GABA and 5-HT levels, and suppressed inflammation and oxidative stress. | [206] |
| Ellagic acid | Rats | Aging | 30 mg/kg | Increased the levels of antioxidant enzymes and reduced malondialdehyde concentrations. | [207] |
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Okoro, N.O.; Odiba, A.S.; Osadebe, P.O.; Omeje, E.O.; Liao, G.; Fang, W.; Jin, C.; Wang, B. Bioactive phytochemicals with anti-aging and lifespan extending potentials in Caenorhabditis elegans. Molecules 2021, 26, 7323. [Google Scholar] [CrossRef]
- Song, L.; Zhang, S. Anti-aging activity and modes of action of compounds from natural food sources. Biomolecules 2023, 13, 1600. [Google Scholar] [CrossRef]
- Zhang, L.; Yousefzadeh, M.J.; Suh, Y.; Niedernhofer, L.J.; Robbins, P.D. Signal Transduction, Ageing and Disease, Subcellular Biochemistry; Springer: New York, NY, USA, 2019; Volume 91, pp. 227–247. [Google Scholar] [CrossRef]
- Kuppusamy, U.R.; Arumugam, B. Chapter 18—Novel plant bioactives, their antiaging potencies: Reality and promises. In Plant Bioactives as Natural Panacea against Age-Induced Diseases; Pandey, K.B., Suttajit, M., Eds.; Elsevier: Amsterdam, The Netherlands, 2023; pp. 359–386. [Google Scholar] [CrossRef]
- World Population Prospects. 2022. Available online: https://www.un.org/development/desa/pd/sites/www.un.org.development.desa.pd/files/wpp2022_summary_of_results.pdf (accessed on 16 January 2024).
- McHugh, D.; Gil, J. Senescence and aging: Causes, consequences, and therapeutic avenues. J. Cell Biol. 2018, 217, 65–77. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, Z.; Ren, Y.; Wang, Y.; Fang, J.; Yue, H.; Ma, S.; Guan, F. Aging and age-related diseases: From mechanisms to therapeutic strategies. Biogerontology 2021, 22, 165–187. [Google Scholar] [CrossRef]
- Dhalaria, R.; Verma, R.; Kumar, D.; Puri, S.; Tapwal, A.; Kumar, V.; Nepovimova, E.; Kuca, K. Bioactive compounds of edible fruits with their anti-aging properties: A comprehensive review to prolong human life. Antioxidants 2020, 9, 1123. [Google Scholar] [CrossRef]
- Mechchate, H.; El Allam, A.; El Omari, N.; El Hachlafi, N.; Shariati, M.A.; Wilairatana, P.; Mubarak, M.S.; Bouyahya, A. Vegetables and Their Bioactive Compounds as Anti-Aging Drugs. Molecules 2022, 27, 2316. [Google Scholar] [CrossRef]
- Russo, G.L.; Spagnuolo, C.; Russo, M.; Tedesco, I.; Moccia, S.; Cervellera, C. Mechanisms of aging and potential role of selected polyphenols in extending healthspan. Biochem. Pharmacol. 2020, 173, 113719. [Google Scholar] [CrossRef]
- Sripanidkulchai, B.; Suttajit, M.; Ratanavalachai, T. Chapter 2—Anti-aging strategies, plant bioactives, and drug development: Current insights. In Plant Bioactives as Natural Panacea against Age-Induced Diseases; Pandey, K.B., Suttajit, M., Eds.; Elsevier: Amsterdam, The Netherlands, 2023; pp. 23–48. [Google Scholar]
- Bjørklund, G.; Shanaida, M.; Lysiuk, R.; Butnariu, M.; Peana, M.; Sarac, I.; Strus, O.; Smetanina, K.; Chirumbolo, S. Natural compounds and products from an anti-aging perspective. Molecules 2022, 27, 7084. [Google Scholar] [CrossRef]
- Ukpai, O.M.; Ijioma, S.N.; Kanu, K.; Orieke, D.; Chinedu-Ndukwe, P.A.; Ugwuanyi, K.C.; Ugbogu, E.A. Phytochemical composition, toxicological profiling and effect on pup birth weight of Corchorus olitorius leaf extract in rats: Implications for fetal macrosomia control. J. Ethnopharmacol. 2024, 319, 117170. [Google Scholar] [CrossRef] [PubMed]
- Ugbogu, E.A.; Okoro, H.; Emmanuel, O.; Ugbogu, O.C.; Ekweogu, C.N.; Uche, M.; Dike, E.D.; Ijioma, S.N. Phytochemical characterization, anti-diarrhoeal, analgesic, anti-inflammatory activities and toxicity profile of Ananas comosus (L.) Merr (pineapple) leaf in albino rats. J. Ethnopharmacol. 2024, 319, 117224. [Google Scholar] [CrossRef] [PubMed]
- Ekweogu, C.N.; Akubugwo, E.I.; Emmanuel, O.; Nosiri, C.I.; Uche, M.E.; Adurosakin, O.E.; Ijioma, S.N.; Ugbogu, E.A. Phytochemical profiling, toxicity studies, wound healing, analgesic and anti-inflammatory activities of Musa paradisiaca L. Musaceae (Plantain) stem extract in rats. J. Ethnopharmacol. 2024, 322, 117639. [Google Scholar] [CrossRef]
- Luo, J.; Si, H.; Jia, Z.; Liu, D. Dietary Anti-aging polyphenols and potential mechanisms. Antioxidants 2021, 10, 283. [Google Scholar] [CrossRef]
- Maoka, T. Carotenoids as natural functional pigments. J. Natural. Med. 2020, 74, 1–16. [Google Scholar] [CrossRef]
- Moreau, R.A.; Whitaker, B.D.; Hicks, K.B. Phytosterols, phytostanols, and their conjugates in foods: Structural diversity, quantitative analysis, and health-promoting uses. Progress Lipid Res. 2002, 41, 457–500. [Google Scholar] [CrossRef]
- Huang, M.; Lu, J.J.; Huang, M.Q.; Bao, J.L.; Chen, X.P.; Wang, Y.T. Terpenoids: Natural products for cancer therapy. Expert Opin. Investigat. Drugs 2012, 21, 1801–1818. [Google Scholar] [CrossRef]
- Kim, S.G.; Sung, J.Y.; Kim, J.R.; Choi, H.C. Quercetin-induced apoptosis ameliorates vascular smooth muscle cell senescence through AMP-activated protein kinase signaling pathway. Korean J. Physiol. Pharmacol. 2020, 24, 69–79. [Google Scholar] [CrossRef]
- Liu, X.; Lin, X.; Zhang, S.; Guo, C.; Li, J.; Mi, Y.; Zhang, C. Lycopene ameliorates oxidative stress in the aging chicken ovary via activation of Nrf2/HO-1 pathway. Aging 2018, 10, 2016–2036. [Google Scholar] [CrossRef]
- Ikonne, E.U.; Ikpeazu, V.O.; Ugbogu, E.A. The potential health benefits of dietary natural plant products in age related eye diseases. Heliyon 2020, 6, e04408. [Google Scholar] [CrossRef]
- Iweala, E.J.; Oluwapelumi, A.E.; Dania, O.E.; Ugbogu, E.A. Bioactive phytoconstituents and their therapeutic potentials in the treatment of haematological cancers: A Review. Life 2023, 13, 1422. [Google Scholar] [CrossRef]
- Murphy, C.T.; McCarroll, S.A.; Bargmann, C.I.; Fraser, A.; Kamath, R.S.; Ahringer, J.; Li, H.; Kenyon, C. Genes that act downstream of DAF-16 to influence the lifespan of Caenorhabditis elegans. Nature 2003, 424, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Murphy, C.T. The search for DAF-16/FOXO transcriptional targets: Approaches and discoveries. Exp. Gerontol. 2006, 41, 910–921. [Google Scholar] [CrossRef]
- Morris, B.J.; Willcox, D.C.; Donlon, T.A.; Willcox, B.J. FOXO3: A major gene for human longevity—A mini-review. Gerontology 2015, 61, 515–525. [Google Scholar] [CrossRef]
- Willcox, B.J.; Donlon, T.A.; He, Q.; Chen, R.; Grove, J.S.; Yano, K.; Masaki, K.H.; Willcox, D.C.; Rodriguez, B.; Curb, J.D. FOXO3A genotype is strongly associated with human longevity. Proc. Natl. Acad. Sci. USA 2008, 105, 13987–13992. [Google Scholar] [CrossRef]
- Flachsbart, F.; Caliebe, A.; Kleindorp, R. Association of FOXO3A variation with human longevity confirmed in German centenarians. Proc. Natl. Acad. Sci. USA 2009, 106, 2700–2705. [Google Scholar] [CrossRef]
- Pietsch, K.; Saul, N.; Menzel, R.; Stürzenbaum, S.R.; Steinberg, C.E.W. Quercetin mediated lifespan extension in Caenorhabditis elegans is modulated by age-1, daf-2, sek-1 and unc-43. Biogerontology 2009, 10, 565–578. [Google Scholar] [CrossRef]
- Baur, J.A.; Pearson, K.J.; Price, N.L.; Jamieson, H.A.; Lerin, C.; Kalra, A.; Prabhu, V.V.; Allard, J.S.; Lopez-Lluch, G.; Lewis, K.; et al. Resveratrol improves health and survival of mice on a high-calorie diet. Nature 2006, 444, 337–342. [Google Scholar] [CrossRef]
- Pearson, K.J.; Baur, J.A.; Lewis, K.N.; Peshkin, L.; Price, N.L.; Labinskyy, N.; Swindell, W.R.; Kamara, D.; Minor, R.K.; Perez, E.; et al. Resveratrol delays age-related deterioration and mimics transcriptional aspects of dietary restriction without extending life span. Cell Metab. 2008, 8, 157–168. [Google Scholar] [CrossRef]
- Miller, R.A.; Harrison, D.E.; Astle, C.M.; Baur, J.A.; Boyd, A.R.; de Cabo, R.; Fernandez, E.; Flurkey, K.; Javors, M.A.; Nelson, J.F.; et al. Rapamycin, but not resveratrol or simvastatin, extends life span of genetically heterogeneous mice. J. Gerontol. A Biol. Sci. Med. Sci. 2011, 66, 191–201. [Google Scholar] [CrossRef]
- Mao, K.; Quipildor, G.F.; Tabrizian, T.; Novaj, A.; Guan, F.; Walters, R.O.; Delahaye, F.; Hubbard, G.B.; Ikeno, Y.; Ejima, K.; et al. Late-life targeting of the IGF-1 receptor improves healthspan and lifespan in female mice. Nat. Commun. 2018, 9, 2394. [Google Scholar] [CrossRef]
- Johnson, S.C.; Rabinovitch, P.S.; Kaeberlein, M. mTOR is a key modulator of ageing and age-related disease. Nature 2013, 493, 338–345. [Google Scholar] [CrossRef]
- Cuervo, A.M.; Bergamini, E.; Brunk, U.T.; Dröge, W.; Ffrench, M.; Terman, A. Autophagy and aging: The importance of maintaining “clean” cells. Autophagy 2005, 1, 131–140. [Google Scholar] [CrossRef]
- Rubinsztein, D.C.; Mariño, G.; Kroemer, G. Autophagy and aging. Cell 2011, 146, 682–695. [Google Scholar] [CrossRef]
- Witczak, C.A.; Sharoff, C.G.; Goodyear, L.J. AMP-activated protein kinase in skeletal muscle: From structure and localization to its role as a master regulator of cellular metabolism. Cell Mol. Life Sci. 2008, 65, 3737–3755. [Google Scholar] [CrossRef]
- Hwang, J.T.; Park, I.J.; Shin, J.I. Genistein, EGCG, and capsaicin inhibit adipocyte differentiation process via activating AMP-activated protein kinase. Biochem. Biophys. Res. Commun. 2005, 338, 694–699. [Google Scholar] [CrossRef]
- Kim, T.; Davis, J.; Zhang, A.J.; He, X.; Mathews, S.T. Curcumin activates AMPK and suppresses gluconeogenic gene expression in hepatoma cells. Biochem. Biophys. Res. Commun. 2009, 388, 377–382. [Google Scholar] [CrossRef]
- Bruning, A. Inhibition of mTOR signaling by quercetin in cancer treatment and prevention. Anticancer Agents Med. Chem. 2013, 13, 1025–1031. [Google Scholar] [CrossRef]
- Huang, S. Inhibition of PI3K/Akt/mTOR signaling by natural products. Anticancer Agents Med. Chem. 2013, 13, 967–970. [Google Scholar] [CrossRef]
- Syed, D.; Adhami, V.; Khan, M.; Mukhtar, H. Inhibition of Akt/mTOR signaling by the dietary flavonoid fisetin. Anticancer Agents Med. Chem. 2013, 13, 995–1001. [Google Scholar] [CrossRef]
- Rodgers, J.T.; Lerin, C.; Gerhart-Hines, Z.; Puigserver, P. Metabolic adaptations through the PGC-1 alpha and SIRT1 pathways. FEBS Lett. 2008, 582, 46–53. [Google Scholar] [CrossRef]
- Brunet, A.; Sweeney, L.B.; Sturgill, J.F.; Chua, K.F.; Greer, P.L.; Lin, Y.; Tran, H.; Ross, S.E.; Mostoslavsky, R.; Cohen, H.Y.; et al. Stress-dependent regulation of FOXO transcription factors by the SIRT1 deacetylase. Science 2004, 303, 2011–2015. [Google Scholar] [CrossRef]
- Martel, J.; Ojcius, D.M.; Ko, Y.F.; Chang, C.J.; Young, J.D. Antiaging effects of bioactive molecules isolated from plants and fungi. Med. Res. Rev. 2019, 39, 1515–1552. [Google Scholar] [CrossRef]
- Vatner, S.F.; Zhang, J.; Oydanich, M.; Berkman, T.; Naftalovich, R.; Vatner, D.E. Healthful aging mediated by inhibition of oxidative stress. Ageing Res. Rev. 2020, 64, 101194. [Google Scholar] [CrossRef] [PubMed]
- Butt, M.S.; Sultan, M.T. Coffee and its consumption: Benefits and risks. Crit. Rev. Food Sci. Nutr. 2011, 51, 363–373. [Google Scholar] [CrossRef]
- Yu, Y.; Zhang, Z.; Chang, C. Chlorogenic acid intake guidance: Sources, health benefits, and safety. Asia Pac. J. Clin. Nutr. 2022, 31, 602–610. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Singla, R.K.; Pandey, A.K. Chlorogenic acid: A dietary phenolic acid with promising pharmacotherapeutic potential. Cur. Med. Chem. 2023, 30, 3905–3926. [Google Scholar] [CrossRef] [PubMed]
- Liang, N.; Kitts, D.D. Role of chlorogenic acids in controlling oxidative and inflammatory stress conditions. Nutrients 2015, 8, 16. [Google Scholar] [CrossRef]
- Heitman, E.; Ingram, D.K. Cognitive and neuroprotective effects of chlorogenic acid. Nutr. Neurosci. 2017, 20, 32–39. [Google Scholar] [CrossRef]
- Zheng, S.Q.; Huang, X.B.; Xing, T.K.; Ding, A.J.; Wu, G.S.; Luo, H.R. Chlorogenic acid extends the lifespan of Caenorhabditis elegans via insulin/IGF-1 signaling pathway. J. Gerontol. Ser. A Biomed. Sci. Med. Sci. 2017, 72, 464–472. [Google Scholar]
- Son, H.G.; Altintas, O.; Kim, E.J.E.; Kwon, S.; Lee, S.J.V. Age-dependent changes and biomarkers of aging in Caenorhabditis elegans. Aging Cell 2019, 18, e12853. [Google Scholar] [CrossRef]
- Siswanto, F.M.; Sakuma, R.; Oguro, A.; Imaoka, S. Chlorogenic acid activates Nrf2/SKN-1 and prolongs the lifespan of Caenorhabditis elegans via the Akt-FOXO3/DAF16a-DDB1 pathway and activation of DAF16f. J. Gerontol. Ser. A. 2022, 77, 1503–1516. [Google Scholar] [CrossRef]
- Tullet, J.M.; Hertweck, M.; An, J.H.; Baker, J.; Hwang, J.Y.; Liu, S.; Oliveira, R.P.; Baumeister, R.; Blackwell, T.K. Direct inhibition of the longevity-promoting factor SKN-1 by insulin-like signaling in C. elegans. Cell 2008, 132, 1025–1038. [Google Scholar] [CrossRef]
- Tungmunnithum, D.; Abid, M.; Elamrani, A.; Drouet, S.; Addi, M.; Hano, C. Almond skin extracts and chlorogenic acid delay chronological aging and enhanced oxidative stress response in yeast. Life 2020, 10, 80. [Google Scholar] [CrossRef]
- Hada, Y.; Uchida, H.A.; Otaka, N.; Onishi, Y.; Okamoto, S.; Nishiwaki, M.; Takemoto, R.; Takeuchi, H.; Wada, J. The protective effect of chlorogenic acid on vascular senescence via the Nrf2/HO-1 pathway. Int. J. Mol. Sci. 2020, 21, 4527. [Google Scholar] [CrossRef]
- Kato, M.; Ochiai, R.; Kozuma, K.; Sato, H.; Katsuragi, Y. Effect of chlorogenic acid intake on cognitive function in the elderly: A pilot study. Evid. Based Complement. Alternat. Med. 2018, 2018, 8608497. [Google Scholar] [CrossRef]
- Suzukamo, C.; Ochiai, R.; Mitsui, Y.; Osaki, N.; Ono, T. Short-Term Intake of Chlorogenic Acids Improves Psychomotor Speed and Motor Speed in Adults: A Randomized Crossover Trial. Brain Sci. 2022, 12, 370. [Google Scholar] [CrossRef]
- Saitou, K.; Ochiai, R.; Kozuma, K.; Sato, H.; Koikeda, T.; Osaki, N.; Katsuragi, Y. Effect of chlorogenic acids on cognitive function: A randomized, double-blind, placebo-controlled trial. Nutrients 2018, 10, 1337. [Google Scholar] [CrossRef]
- Girsang, E.; Ginting, C.N.; Lister, I.N.E.; yashfa Gunawan, K.; Widowati, W. Anti-inflammatory and antiaging properties of chlorogenic acid on UV-induced fibroblast cell. Peer J. 2021, 9, e11419. [Google Scholar] [CrossRef]
- Tanigawa, T.; Kanazawa, S.; Ichibori, R.; Fujiwara, T.; Magome, T.; Shingaki, K.; Miyata, S.; Hata, Y.; Tomita, K.; Matsuda, K.; et al. (+)-Catechin protects dermal fibroblasts against oxidative stress-induced apoptosis. BMC Complement. Alternat. Med. 2014, 14, 1–7. [Google Scholar] [CrossRef]
- Ribitsch, I.; Gueltekin, S.; Keith, M.F.; Minichmair, K.; Peham, C.; Jenner, F.; Egerbacher, M. Age-related changes of tendon fibril micro-morphology and gene expression. J. Anat. 2020, 236, 688–700. [Google Scholar] [CrossRef]
- Alves, G.D.A.D.; Oliveira de Souza, R.; Ghislain Rogez, H.L.; Masaki, H.; Fonseca, M.J.V. Cecropia obtusa extract and chlorogenic acid exhibit antiaging effect in human fibroblasts and keratinocytes cells exposed to UV radiation. PLoS ONE 2019, 14, e0216501. [Google Scholar] [CrossRef]
- Singh, M.; Kapoor, A.; Bhatnagar, A. Oxidative and reductive metabolism of lipid-peroxidation derived carbonyls. Chemico-Biol. Interact. 2015, 234, 261–273. [Google Scholar] [CrossRef]
- Kim, J.; Lee, Y.M.; Jung, W.; Park, S.B.; Kim, C.S.; Kim, J.S. Aster koraiensis extract and chlorogenic acid inhibit retinal angiogenesis in a mouse model of oxygen-induced retinopathy. Evid. Based Complement. Alternat. Med. 2018, 2018, 6402650. [Google Scholar] [CrossRef]
- Apte, R.S. Age-related macular degeneration. N. Engl. J. Med. 2021, 385, 539–547. [Google Scholar] [CrossRef]
- Fiorani, M.; Guidarelli, A.; Blasa, M.; Azzolini, C.; Candiracci, M.; Piatti, E.; Cantoni, O. Mitochondria accumulate large amounts of quercetin: Prevention of mitochondrial damage and release upon oxidation of the extramitochondrial fraction of the flavonoid. J. Nutr. Biochem. 2010, 21, 397–404. [Google Scholar] [CrossRef]
- Deepika; Maurya, P.K. Health benefits of quercetin in age-related diseases. Molecules 2022, 27, 2498. [Google Scholar] [CrossRef]
- Choi, S.M.; Kim, B.C.; Cho, Y.H.; Choi, K.H.; Chang, J.; Park, M.S.; Kim, M.K.; Cho, K.H.; Kim, J.K. Effects of flavonoid compounds on β-amyloid-peptide-induced neuronal death in cultured mouse cortical neurons. Chonnam Med. J. 2014, 50, 45–51. [Google Scholar] [CrossRef]
- Nguyen, T.L.A.; Bhattacharya, D. Antimicrobial activity of quercetin: An approach to its mechanistic principle. Molecules 2022, 27, 2494. [Google Scholar] [CrossRef]
- Liu, R.; Zhang, T.T.; Zhou, D.; Bai, X.Y.; Zhou, W.L.; Huang, C. Quercetin protects against the Aβ25–35-induced amnesic injury: Involvement of inactivation of RAGE-mediated pathway and conservation of the NVU. Neuropharmacology 2013, 67, 419–431. [Google Scholar] [CrossRef]
- Yang, Y.; Li, Y.; Du, X.; Liu, Z.; Zhu, C.; Mao, W.; Liu, G.; Jiang, Q. Anti-aging effects of quercetin in Cladocera Simocephalus vetulus using proteomics. ACS Omega 2023, 8, 17609–17619. [Google Scholar] [CrossRef] [PubMed]
- Remigante, A.; Spinelli, S.; Trichilo, V.; Loddo, S.; Sarikas, A.; Pusch, M.; Dossena, S.; Marino, A.; Morabito, R. D-Galactose induced early aging in human erythrocytes: Role of band 3 protein. J. Cell. Physiol. 2022, 237, 1586–1596. [Google Scholar] [CrossRef]
- Remigante, A.; Spinelli, S.; Basile, N.; Caruso, D.; Falliti, G.; Dossena, S.; Marino, A.; Morabito, R. Oxidation stress as a mechanism of aging in human erythrocytes: Protective effect of quercetin. Int. J. Mol. Sci. 2022, 23, 7781. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Cai, J.; Fang, Z.; Li, S.; Huang, Z.; Tang, Z.; Luo, Q.; Chen, H. The composition and anti-aging activities of polyphenol extract from Phyllanthus emblica L. fruit. Nutrients 2022, 14, 857. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.; Zhao, X.; Amevor, F.K.; Du, X.; Wang, Y.; Li, D.; Shu, G.; Tian, Y.; Zhao, X. Therapeutic application of quercetin in aging-related diseases: SIRT1 as a potential mechanism. Front. Immunol. 2022, 13, 943321. [Google Scholar] [CrossRef] [PubMed]
- Abharzanjani, F.; Afshar, M.; Hemmati, M.; Moossavi, M. Short-term High Dose of Quercetin and Resveratrol Alters Aging Markers in Human Kidney Cells. Int. J. Prev. Med. 2017, 8, 64. [Google Scholar] [CrossRef] [PubMed]
- Forney, L.A.; Lenard, N.R.; Stewart, L.K.; Henagan, T.M. Dietary quercetin attenuates adipose tissue expansion and inflammation and alters adipocyte morphology in a tissue-specific manner. Int. J. Mol. Sci. 2018, 19, 895. [Google Scholar] [CrossRef] [PubMed]
- Kobori, M.; Takahashi, Y.; Sakurai, M.; Akimoto, Y.; Tsushida, T.; Oike, H.; Ippoushi, K. Quercetin suppresses immune cell accumulation and improves mitochondrial gene expression in adipose tissue of diet-induced obese mice. Mol. Nutr. Food Res. 2016, 60, 300–312. [Google Scholar] [CrossRef] [PubMed]
- Zoico, E.; Nori, N.; Darra, E.; Tebon, M.; Rizzatti, V.; Policastro, G.; De Caro, A.; Rossi, A.P.; Fantin, F.; Zamboni, M. Senolytic effects of quercetin in an in vitro model of pre-adipocytes and adipocytes induced senescence. Sci. Rep. 2021, 11, 23237. [Google Scholar] [CrossRef] [PubMed]
- Shin, E.J.; Lee, J.S.; Hong, S.; Lim, T.G.; Byun, S. Quercetin directly targets JAK2 and PKCδ and prevents UV-induced photoaging in human skin. Int. J. Mol. Sci. 2019, 20, 5262. [Google Scholar] [CrossRef] [PubMed]
- Ganeshpurkar, A.; Saluja, A.K. The pharmacological potential of rutin. Saudi Pharma. J. 2017, 25, 149–164. [Google Scholar] [CrossRef]
- Li, T.; Chen, S.; Feng, T.; Dong, J.; Li, Y.; Li, H. Rutin protects against aging-related metabolic dysfunction. Food Funct. 2016, 7, 1147–1154. [Google Scholar] [CrossRef]
- Makino, T.; Kanemaru, M.; Okuyama, S.; Shimizu, R.; Tanaka, H.; Mizukami, H. Anti-allergic effects of enzymatically modified isoquercitrin (α-oligoglucosyl quercetin 3-O-glucoside), quercetin 3-O-glucoside, α-oligoglucosyl rutin, and quercetin, when administered orally to mice. J. Nat. Med. 2013, 67, 881–886. [Google Scholar] [CrossRef]
- Han, Y.; Ding, Y.; Xie, D.; Hu, D.; Li, P.; Li, X.; Xue, W.; Jin, L.; Song, B. Design, synthesis, and antiviral activity of novel rutin derivatives containing 1, 4-pentadien-3-one moiety. Europ. J. Med. Chem. 2015, 92, 732–737. [Google Scholar] [CrossRef]
- Perk, A.A.; Shatynska-Mytsyk, I.; Gerçek, Y.C.; Boztaş, K.; Yazgan, M.; Fayyaz, S.; Farooqi, A.A. Rutin mediated targeting of signaling machinery in cancer cells. Cancer Cell Int. 2014, 14, 1–5. [Google Scholar] [CrossRef]
- Choi, S.J.; Lee, S.N.; Kim, K.; Joo, D.H.; Shin, S.; Lee, J. Biological effects of rutin on skin aging. Int. J. Mol. Med. 2016, 38, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Xu, Q.; Wufuer, H.; Li, Z.; Sun, R.; Jiang, Z.; Dou, X.; Fu, Q.; Campisi, J.; Sun, Y. Rutin is a potent senomorphic agent to target senescent cells and can improve chemotherapeutic efficacy. Aging Cell 2024, 23, e13921. [Google Scholar] [CrossRef] [PubMed]
- Chattopadhyay, D.; Chitnis, A.; Talekar, A.; Mulay, P.; Makkar, M.; James, J.; Thirumurugan, K. Hormetic efficacy of rutin to promote longevity in Drosophila melanogaster. Biogerontology 2017, 18, 397–411. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Li, J.; Pan, R.; Cheng, J.; Cui, Q.; Chen, J.; Yuan, Z. Sodium rutin extends lifespan and health span in mice including positive impacts on liver health. Br. J. Pharmacol. 2022, 179, 1825–1838. [Google Scholar] [CrossRef]
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and anthocyanins: Colored pigments as food, pharmaceutical ingredients, and the potential health benefits. Food Nutr. Res. 2017, 61, 1361779. [Google Scholar] [CrossRef]
- Speer, H.; D’Cunha, N.M.; Alexopoulos, N.I.; McKune, A.J.; Naumovski, N. Anthocyanins and human health—A focus on oxidative stress, inflammation and disease. Antioxidants 2020, 9, 366. [Google Scholar] [CrossRef]
- D’Cunha, N.M.; Georgousopoulou, E.N.; Dadigamuwage, L.; Kellett, J.; Panagiotakos, D.B.; Thomas, J.; McKune, A.J.; Mellor, D.D.; Naumovski, N. Effect of long-term nutraceutical and dietary supplement use on cognition in the elderly: A 10-year systematic review of randomised controlled trials. Br. J. Nutr. 2018, 119, 280–298. [Google Scholar] [CrossRef]
- Ullah, R.; Khan, M.; Shah, S.A.; Saeed, K.; Kim, M.O. Natural antioxidant anthocyanins—A hidden therapeutic candidate in metabolic disorders with major focus in neurodegeneration. Nutrients 2019, 11, 1195. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, H.; Han, Y.; Pei, Y.; Guo, Y.; Cui, S.W. Purple sweet potato extract maintains intestinal homeostasis and extend lifespan through increasing autophagy in female Drosophila melanogaster. J. Food Biochem. 2021, 45, e13861. [Google Scholar] [CrossRef]
- Buszczak, M.; Krämer, H. Autophagy keeps the balance in tissue homeostasis. Dev. Cell 2019, 49, 499–500. [Google Scholar] [CrossRef]
- Han, Y.; Guo, Y.; Cui, S.W.; Li, H.; Shan, Y.; Wang, H. Purple sweet potato extract extends lifespan by activating autophagy pathway in male Drosophila melanogaster. Experiment. Gerontol. 2021, 144, 111190. [Google Scholar] [CrossRef]
- Wang, L.; Li, Y.M.; Lei, L.; Liu, Y.; Wang, X.; Ma, K.Y.; Chen, Z.Y. Cranberry anthocyanin extract prolongs lifespan of fruit flies. Experiment. Gerontol. 2015, 69, 189–195. [Google Scholar] [CrossRef]
- Katewa, S.D.; Kapahi, P. Role of TOR signaling in aging and related biological processes in Drosophila melanogaster. Experiment. Gerontol. 2011, 46, 382–390. [Google Scholar] [CrossRef]
- Peng, C.; Zuo, Y.; Kwan, K.M.; Liang, Y.; Ma, K.Y.; Chan, H.Y.E.; Huang, Y.; Yu, H.; Chen, Z.Y. Blueberry extract prolongs lifespan of Drosophila melanogaster. Experiment. Gerontol. 2012, 47, 170–178. [Google Scholar] [CrossRef]
- Sun, X.; Komatsu, T.; Lim, J.; Laslo, M.; Yolitz, J.; Wang, C.; Poirier, L.; Alberico, T.; Zou, S. Nutrient-dependent requirement for SOD1 in lifespan extension by protein restriction in Drosophila melanogaster. Aging Cell 2012, 11, 783–793. [Google Scholar] [CrossRef]
- Yan, F.; Chen, Y.; Azat, R.; Zheng, X. Mulberry anthocyanin extract ameliorates oxidative damage in hepg2 cells and prolongs the lifespan of Caenorhabditis elegans through MAPK and Nrf2 pathways. Oxid. Med. Cell Longev. 2017, 2017, 7956158. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wang, J.; Zhang, L.; Zheng, Y.; Ma, G.; Sun, X.; Yuan, J. Preparation of Dendrobium officinale flower anthocyanin and extended lifespan in Caenorhabditis elegans. Molecules 2022, 27, 8608. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Song, X.; Zhang, D.; Zhou, F.; Wang, D.; Wei, Y.; Gao, F.; Xie, L.; Jia, G.; Wu, W.; et al. Blueberry anthocyanins: Protection against ageing and light-induced damage in retinal pigment epithelial cells. Br. J. Nutr. 2012, 108, 16–27. [Google Scholar] [CrossRef]
- Zhang, L.X.; Li, C.X.; Kakar, M.U.; Khan, M.S.; Wu, P.F.; Amir, R.M.; Dai, D.F.; Naveed, M.; Li, Q.Y.; Saeed, M.; et al. Resveratrol (RV): A pharmacological review and call for further research. Biomed. Pharmacother. 2021, 143, 112164. [Google Scholar] [CrossRef]
- Porquet, D.; Casadesús, G.; Bayod, S.; Vicente, A.; Canudas, A.M.; Vilaplana, J.; Pelegrí, C.; Sanfeliu, C.; Camins, A.; Pallàs, M.; et al. Dietary resveratrol prevents Alzheimer’s markers and increases life span in SAMP8. Age 2013, 35, 1851–1865. [Google Scholar] [CrossRef]
- D’Souza, Y.; Elharram, A.; Soon-Shiong, R.; Andrew, R.D.; Bennett, B.M. Characterization of Aldh2-/-mice as an age-related model of cognitive impairment and Alzheimer’s disease. Mol. Brain 2015, 8, 1–16. [Google Scholar] [CrossRef]
- Bayod, S.; Del Valle, J.; Pelegri, C.; Vilaplana, J.; Canudas, A.M.; Camins, A.; Jimenez, A.; Sanchez-Roige, S.; Lalanza, J.F.; Escorihuela, R.M.; et al. Macroautophagic process was differentially modulated by long-term moderate exercise in rat brain and peripheral tissues. J. Physiol. Pharmacol. 2014, 65, 229–239. [Google Scholar]
- Donmez, G.; Wang, D.; Cohen, D.E.; Guarente, L. Retraction notice to: SIRT1 suppresses β-amyloid production by activating the α-secretase gene ADAM10. Cell 2014, 158, 959. [Google Scholar] [CrossRef]
- Song, J.; Liu, L.; Hao, K.; Mao, S.; Tang, Y.; Tong, X.; Dai, F. Resveratrol elongates the lifespan and improves antioxidant activity in the silkworm Bombyx mori. J. Pharmaceut. Anal. 2021, 11, 374–382. [Google Scholar] [CrossRef]
- Abolaji, A.O.; Adedara, A.O.; Adie, M.A.; Vicente-Crespo, M.; Farombi, E.O. Resveratrol prolongs lifespan and improves 1-methyl-4-phenyl-1, 2, 3, 6-tetrahydropyridine-induced oxidative damage and behavioural deficits in Drosophila melanogaster. Biochem. Biophys. Res. Commun. 2018, 503, 1042–1048. [Google Scholar] [CrossRef]
- Takihara, Y.; Inatani, M.; Eto, K.; Inoue, T.; Kreymerman, A.; Miyake, S.; Ueno, S.; Nagaya, M.; Nakanishi, A.; Iwao, K.; et al. In vivo imaging of axonal transport of mitochondria in the diseased and aged mammalian CNS. Proc. Natl. Acad. Sci. USA 2015, 112, 10515–10520. [Google Scholar] [CrossRef] [PubMed]
- Reddy, P.H.; Reddy, T.P. Mitochondria as a therapeutic target for aging and neurodegenerative diseases. Curr. Alzheimer Res. 2011, 8, 393–409. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Luo, Z.; Jin, M.; Sheng, W.; Wang, H.T.; Long, X.; Wu, Y.; Hu, P.; Xu, H.; Zhang, X. Exploration of age-related mitochondrial dysfunction and the anti-aging effects of resveratrol in zebrafish retina. Aging 2019, 11, 3117. [Google Scholar] [CrossRef]
- Baek, S.H.; Shin, W.C.; Ryu, H.S.; Lee, D.W.; Moon, E.; Seo, C.S.; Hwang, E.; Lee, H.S.; Ahn, M.H.; Jeon, Y.; et al. Creation of resveratrol-enriched rice for the treatment of metabolic syndrome and related diseases. PLoS ONE 2013, 8, e57930. [Google Scholar] [CrossRef]
- Islam, M.S.; Jin, Y.Y.; Chung, H.J.; Kim, H.J.; Baek, S.H.; Hong, S.T. Effect of the resveratrol rice DJ526 on longevity. Nutrients 2019, 11, 1804. [Google Scholar] [CrossRef]
- Khan, M.; Park, S.; Kim, H.J.; Lee, K.J.; Kim, D.H.; Baek, S.H.; Hong, S.T. The resveratrol rice DJ526 callus significantly increases the lifespan of drosophila (resveratrol rice DJ526 callus for longevity). Nutrients 2019, 11, 983. [Google Scholar] [CrossRef]
- Kim, H.S.; Quon, M.J.; Kim, J.A. New insights into the mechanisms of polyphenols beyond antioxidant properties; lessons from the green tea polyphenol, epigallocatechin 3-gallate. Redox Biol. 2014, 2, 187–195. [Google Scholar] [CrossRef]
- Mehmood, S.; Maqsood, M.; Mahtab, N.; Khan, M.I.; Sahar, A.; Zaib, S.; Gul, S. Epigallocatechin gallate: Phytochemistry, bioavailability, utilization challenges, and strategies. J. Food Biochem. 2022, 46, e14189. [Google Scholar] [CrossRef]
- Cai, S.X.; Huang, J.A.; Wang, L.L.; Dong, X.R.; Gong, Y.S.; Li, J.; Li, Q.; Liu, Z.; Luo, G. Inhibiting effects of epigallocatechin gallate (EGCG) on the formation of age pigment in vitro and in vivo. J. Med. Plants Res. 2011, 5, 5470–5478. [Google Scholar]
- Kumar, R.; Sharma, A.; Kumari, A.; Gulati, A.; Padwad, Y.; Sharma, R. Epigallocatechin gallate suppresses premature senescence of preadipocytes by inhibition of PI3K/Akt/mTOR pathway and induces senescent cell death by regulation of Bax/Bcl-2 pathway. Biogerontology 2019, 20, 171–189. [Google Scholar] [CrossRef]
- Lilja, S.; Oldenburg, J.; Pointner, A.; Dewald, L.; Lerch, M.; Hippe, B.; Switzeny, O.; Haslberger, A. Epigallocatechin gallate effectively affects senescence and anti-SASP via SIRT3 in 3T3-L1 preadipocytes in comparison with other bioactive substances. Oxid. Med. Cell Longev. 2020, 2020, 4793125. [Google Scholar] [CrossRef] [PubMed]
- Wagner, A.E.; Piegholdt, S.; Rabe, D.; Baenas, N.; Schloesser, A.; Eggersdorfer, M.; Stocker, A.; Rimbach, G. Epigallocatechin gallate affects glucose metabolism and increases fitness and lifespan in Drosophila melanogaster. Oncotarget 2015, 6, 30568. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Na, L.; Feng, R.; Gong, L.; Zhao, Y.; Li, Q.; Li, Y.; Sun, C. The phytochemical, EGCG, extends lifespan by reducing liver and kidney function damage and improving age-associated inflammation and oxidative stress in healthy rats. Aging Cell 2013, 12, 1041–1049. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Li, Y.; Ling, F.; Guan, Y.; Zhang, D.; Zhu, Q.; Liu, J.; Wu, Y.; Niu, Y. The phytochemical epigallocatechin gallate prolongs the lifespan by improving lipid metabolism, reducing inflammation and oxidative stress in high-fat diet-fed obese rats. Aging Cell 2020, 19, e13199. [Google Scholar] [CrossRef]
- Shoko, T.; Maharaj, V.J.; Naidoo, D.; Tselanyane, M.; Nthambeleni, R.; Khorombi, E.; Apostolides, Z. Anti-aging potential of extracts from Sclerocarya birrea (A. Rich.) Hochst and its chemical profiling by UPLC-Q-TOF-MS. BMC Complement. Alternat. Med. 2018, 18, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Avadhani, K.S.; Manikkath, J.; Tiwari, M.; Chandrasekhar, M.; Godavarthi, A.; Vidya, S.M.; Hariharapura, R.C.; Kalthur, G.; Udupa, N.; Mutalik, S. Skin delivery of epigallocatechin-3-gallate (EGCG) and hyaluronic acid loaded nano-transfersomes for antioxidant and anti-aging effects in UV radiation induced skin damage. Drug Deliv. 2017, 24, 61–74. [Google Scholar] [CrossRef] [PubMed]
- Khan, F.A.; Maalik, A.; Murtaza, G. Inhibitory mechanism against oxidative stress of caffeic acid. J. Food Drug Anal. 2016, 24, 695–702. [Google Scholar] [CrossRef] [PubMed]
- Rusmana, D.; Wahyudianingsih, R.; Elisabeth, M.; Balqis, B.; Maesaroh, M.; Widowati, W. Antioxidant activity of Phyllanthus niruri extract, rutin and quercetin. Indones. Biomed. J. 2017, 9, 84–90. [Google Scholar] [CrossRef]
- Pavlíková, N. Caffeic acid and diseases-mechanisms of action. Int. J. Mol. Sci. 2022, 24, 588. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, M.O.; Christensen, L.P.; Grevsen, K. Harvest strategies for optimization of the content of bioactive alkamides and caffeic acid derivatives in aerial parts and in roots of Echinacea purpurea. J. Agric. Food Chem. 2018, 66, 11630–11639. [Google Scholar] [CrossRef] [PubMed]
- Girsang, E.; Lister, I.N.E.; Ginting, C.N.; Sholihah, I.A.; Raif, M.A.; Kunardi, S.; Million, H.; Widowati, W. Antioxidant and antiaging activity of rutin and caffeic acid. Pharmaciana 2020, 10, 147–156. [Google Scholar] [CrossRef]
- Bastianini, M.; Faffa, C.; Sisani, M.; Petracci, A. Caffeic acid-layered double hydroxide hybrid: A new raw material for cosmetic applications. Cosmetics 2018, 5, 51. [Google Scholar] [CrossRef]
- Sheng, X.; Zhu, Y.; Zhou, J.; Yan, L.; Du, G.; Liu, Z.; Chen, H. Antioxidant effects of caffeic acid lead to protection of drosophila intestinal stem cell aging. Front. Cell Develop. Biol. 2021, 9, 735483. [Google Scholar] [CrossRef]
- Lee, H.; Vilian, A.E.; Kim, J.Y.; Chun, M.H.; Suh, J.S.; Seo, H.H.; Cho, S.H.; Shin, I.S.; Kim, S.J.; Park, S.H.; et al. Design and development of caffeic acid conjugated with Bombyx mori derived peptide biomaterials for anti-aging skin care applications. RSC Adv. 2017, 7, 30205–30213. [Google Scholar] [CrossRef]
- Okada, Y.; Okada, M. Quercetin, caffeic acid and resveratrol regulate circadian clock genes and aging-related genes in young and old human lung fibroblast cells. Mol. Biol Rep. 2020, 47, 1021–1032. [Google Scholar] [CrossRef]
- Gutierrez-Zetina, S.M.; González-Manzano, S.; Ayuda-Durán, B.; Santos-Buelga, C.; González-Paramás, A.M. Caffeic and dihydrocaffeic acids promote longevity and increase stress resistance in Caenorhabditis elegans by modulating expression of stress-related genes. Molecules 2021, 26, 1517. [Google Scholar] [CrossRef]
- Ademosun, O.T.; Adebayo, A.H.; Ajanaku, K.O. Solanum lycopersicum and Daucus carota: Effective anticancer agents (a mini review). J. Phys. Conf. Ser. 2021, 1943, 012169. [Google Scholar] [CrossRef]
- Khan, U.M.; Sevindik, M.; Zarrabi, A.; Nami, M.; Ozdemir, B.; Kaplan, D.N.; Selamoglu, Z.; Hasan, M.; Kumar, M.; Alshehri, M.M.; et al. Lycopene: Food sources, biological activities, and human health benefits. Oxid. Med. Cell. Longev. 2021, 2021, 2713511. [Google Scholar] [CrossRef]
- Riccioni, G.; Mancini, B.; Di Ilio, E.; Bucciarelli, T.; D’orazio, N. Protective effect of lycopene in cardiovascular disease. Europ. Rev. Med. Pharmacol. Sci. 2008, 12, 183–190. [Google Scholar]
- Imran, M.; Ghorat, F.; Ul-Haq, I.; Ur-Rehman, H.; Aslam, F.; Heydari, M.; Shariati, M.A.; Okuskhanova, E.; Yessimbekov, Z.; Thiruvengadam, M.; et al. Lycopene as a natural antioxidant used to prevent human health disorders. Antioxidants 2020, 9, 706. [Google Scholar] [CrossRef]
- Bao, Q.; Pan, J.; Qi, H.; Wang, L.; Qian, H.; Jiang, F.; Hu, X. Aging and age-related diseases–from endocrine therapy to target therapy. Mol. Cell. Endocrinol. 2014, 394, 115–118. [Google Scholar] [CrossRef]
- Zhou, Y.; Dong, Y.; Xu, Q.; He, Y.; Tian, S.; Zhu, S.; Zhu, Y.; Dong, X. Mussel oligopeptides ameliorate cognition deficit and attenuate brain senescence in D-galactose-induced aging mice. Food Chem. Toxicol. 2013, 59, 412–420. [Google Scholar] [CrossRef]
- Petyaev, I.M. Lycopene deficiency in ageing and cardiovascular disease. Oxid. Med. Cell. Longev. 2016, 2016, 3218605. [Google Scholar] [CrossRef]
- Zhao, B.; Liu, H.; Wang, J.; Liu, P.; Tan, X.; Ren, B.; Liu, Z.; Liu, X. Lycopene supplementation attenuates oxidative stress, neuroinflammation, and cognitive impairment in aged CD-1 mice. J. Agric. Food Chem. 2018, 66, 3127–3136. [Google Scholar] [CrossRef]
- Lee, W.; Ku, S.K.; Bae, J.W.; Bae, J.S. Inhibitory effects of lycopene on HMGB1-mediated pro-inflammatory responses in both cellular and animal models. Food Chem. Toxicol. 2012, 50, 1826–1833. [Google Scholar] [CrossRef]
- Min, J.Y.; Min, K.B. Serum lycopene, lutein and zeaxanthin, and the risk of Alzheimer’s disease mortality in older adults. Dement. Geriatr. Cogn. Dis. 2014, 37, 246–256. [Google Scholar] [CrossRef]
- Abir, M.H.; Mahamud, A.S.U.; Tonny, S.H.; Anu, M.S.; Hossain, K.S.; Protic, I.A.; Khan, M.S.U.; Baroi, A.; Moni, A.; Uddin, M.J. Pharmacological potentials of lycopene against aging and aging-related disorders: A review. Food Sci. Nutr. 2023, 11, 5701–5735. [Google Scholar] [CrossRef]
- Rakha, S.I.; Elmetwally, M.A.; El-Sheikh Ali, H.; Balboula, A.Z.; Mahmoud, A.M.; Zaabel, S.M. Lycopene reduces the in vitro aging phenotypes of mouse oocytes by improving their oxidative status. Vet. Sci. 2022, 9, 336. [Google Scholar] [CrossRef]
- López-Nieto, M.J.; Costa, J.; Peiro, E.; Méndez, E.; Rodríguez-Sáiz, M.; De la Fuente, J.L.; Cabri, W.; Barredo, J.L. Biotechnological lycopene production by mated fermentation of Blakeslea trispora. Appl. Microbiol. Biotechnol. 2004, 66, 153–159. [Google Scholar] [CrossRef]
- Hu, W.; Dai, D.; Li, W. Anti-aging effect of Blakeslea trispora powder on adult mice. Biotechnol. Lett. 2013, 35, 1309–1315. [Google Scholar] [CrossRef]
- Liu, X.; Dilxat, T.; Shi, Q.; Qiu, T.; Lin, J. The combination of nicotinamide mononucleotide and lycopene prevents cognitive impairment and attenuates oxidative damage in D-galactose induced aging models via Keap1-Nrf2 signaling. Gene 2022, 822, 146348. [Google Scholar] [CrossRef]
- Sies, H.; Stahl, W. Non-nutritive bioactive constituents of plants: Lycopene, lutein and zeaxanthin. Int. J. Vit. Nutr. Res. 2003, 73, 95–100. [Google Scholar] [CrossRef]
- Zhao, C.; Cheng, H.; Jiang, P.; Yao, Y.; Han, J. Preparation of lutein-loaded particles for improving solubility and stability by Polyvinylpyrrolidone (PVP) as an emulsion-stabilizer. Food Chem. 2014, 156, 123–128. [Google Scholar] [CrossRef]
- Liu, R.; Wang, T.; Zhang, B.; Qin, L.; Wu, C.; Li, Q.; Ma, L. Lutein and zeaxanthin supplementation and association with visual function in age-related macular degeneration. Investigat. Ophthalmol. Vis. Sci. 2015, 56, 252–258. [Google Scholar] [CrossRef]
- Zhang, Z.; Han, S.; Wang, H.; Wang, T. Lutein extends the lifespan of Drosophila melanogaster. Arch. Gerontol. Geriatr. 2014, 58, 153–159. [Google Scholar] [CrossRef]
- Žmitek, K.; Žmitek, J.; Butina, M.R.; Hristov, H.; Pogačnik, T.; Pravst, I. Dietary lutein supplementation protects against ultraviolet-radiation-induced erythema: Results of a randomized double-blind placebo-controlled study. J. Funct. Foods 2020, 75, 104265. [Google Scholar] [CrossRef]
- Hwang, J.S.; Han, S.G.; Lee, C.H.; Seo, H.G. Lutein suppresses hyperglycemia-induced premature senescence of retinal pigment epithelial cells by upregulating SIRT1. J. Food Biochem. 2018, 42, e12495. [Google Scholar] [CrossRef]
- Chae, S.Y.; Park, S.Y.; Park, G. Lutein protects human retinal pigment epithelial cells from oxidative stress induced cellular senescence. Mol. Med. Rep. 2018, 18, 5182–5190. [Google Scholar] [CrossRef]
- Bin Sayeed, M.S.; Karim, S.M.R.; Sharmin, T.; Morshed, M.M. Critical analysis on characterization, systemic effect, and therapeutic potential of beta-sitosterol: A plant-derived orphan phytosterol. Medicines 2016, 3, 29. [Google Scholar] [CrossRef]
- Gupta, R.; Sharma, A.K.; Dobhal, M.P.; Sharma, M.C.; Gupta, R.S. Antidiabetic and antioxidant potential of β-sitosterol in streptozotocin-induced experimental hyperglycemia. J. Diabetes 2011, 3, 29–37. [Google Scholar] [CrossRef]
- Sun, Y.; Gao, L.; Hou, W.; Wu, J. β-Sitosterol alleviates inflammatory response via inhibiting the activation of ERK/p38 and NF-κB pathways in LPS-exposed BV2 cells. Biomed. Res. Int. 2020, 2020, 7532306. [Google Scholar] [CrossRef]
- Hah, Y.S.; Lee, W.K.; Lee, S.; Kim, E.J.; Lee, J.H.; Lee, S.J.; Ji, Y.H.; Kim, S.G.; Lee, H.H.; Hong, S.Y.; et al. β-Sitosterol attenuates dexamethasone-induced muscle atrophy via regulating FoxO1-dependent signaling in C2C12 cell and mice model. Nutrients 2022, 14, 2894. [Google Scholar] [CrossRef]
- Lomenick, B.; Shi, H.; Huang, J.; Chen, C. Identification and characterization of β-sitosterol target proteins. Bioorganic Med. Chem. Lett. 2015, 25, 4976–4979. [Google Scholar] [CrossRef]
- Haiyuan, Y.U.; Shen, X.; Liu, D.; Hong, M.; Lu, Y. The protective effects of β-sitosterol and vermicularin from Thamnolia vermicularis (Sw.) Ach. against skin aging in vitro. An. Acad. Bras. Cienc. 2019, 91, e20181088. [Google Scholar] [CrossRef]
- Ajanaku, C.O.; Ademosun, O.T.; Atohengbe, P.O.; Ajayi, S.O.; Obafemi, Y.D.; Owolabi, O.A.; Akinduti, P.A.; Ajanaku, K.O. Functional bioactive compounds in ginger, turmeric, and garlic. Front. Nutr. 2022, 9, 1012023. [Google Scholar] [CrossRef]
- Bielak-Zmijewska, A.; Grabowska, W.; Ciolko, A.; Bojko, A.; Mosieniak, G.; Bijoch, Ł.; Sikora, E. The role of curcumin in the modulation of ageing. Int. J. Mol. Sci. 2019, 20, 1239. [Google Scholar] [CrossRef]
- Zia, A.; Farkhondeh, T.; Pourbagher-Shahri, A.M.; Samarghandian, S. The role of curcumin in aging and senescence: Molecular mechanisms. Biomed. Pharmacother. 2021, 134, 111119. [Google Scholar] [CrossRef]
- Lee, K.S.; Lee, B.S.; Semnani, S.; Avanesian, A.; Um, C.Y.; Jeon, H.J.; Seong, K.M.; Yu, K.; Min, K.J.; Jafari, M. Curcumin extends life span, improves health span, and modulates the expression of age-associated aging genes in Drosophila melanogaster. Rejuvenation Res. 2010, 13, 561–570. [Google Scholar] [CrossRef]
- Soh, J.W.; Marowsky, N.; Nichols, T.J.; Rahman, A.M.; Miah, T.; Sarao, P.; Khasawneh, R.; Unnikrishnan, A.; Heydari, A.R.; Silver, R.B.; et al. Curcumin is an early-acting stage-specific inducer of extended functional longevity in Drosophila. Experiment. Gerontol. 2013, 48, 229–239. [Google Scholar] [CrossRef]
- Liao, V.H.C.; Yu, C.W.; Chu, Y.J.; Li, W.H.; Hsieh, Y.C.; Wang, T.T. Curcumin-mediated lifespan extension in Caenorhabditis elegans. Mech. Ageing Dev. 2011, 132, 480–487. [Google Scholar] [CrossRef]
- Sun, Y.; Hu, X.; Hu, G.; Xu, C.; Jiang, H. Curcumin attenuates hydrogen peroxide-induced premature senescence via the activation of SIRT1 in human umbilical vein endothelial cells. Biol. Pharmaceut. Bull. 2015, 38, 1134–1141. [Google Scholar] [CrossRef]
- Grabowska, W.; Suszek, M.; Wnuk, M.; Lewinska, A.; Wasiak, E.; Sikora, E.; Bielak-Zmijewska, A. Curcumin elevates sirtuin level but does not postpone in vitro senescence of human cells building the vasculature. Oncotarget 2016, 7, 19201. [Google Scholar] [CrossRef]
- Shailaja, M.; Gowda, K.D.; Vishakh, K.; Kumari, N.S. Anti-aging role of curcumin by modulating the inflammatory markers in albino wistar rats. J. Natl. Med. Assoc. 2017, 109, 9–13. [Google Scholar] [CrossRef]
- Stępień, K.; Wojdyła, D.; Nowak, K.; Mołoń, M. Impact of curcumin on replicative and chronological aging in the Saccharomyces cerevisiae yeast. Biogerontology 2020, 21, 109–123. [Google Scholar] [CrossRef]
- Caporali, S.; De Stefano, A.; Calabrese, C.; Giovannelli, A.; Pieri, M.; Savini, I.; Tesauro, M.; Bernardini, S.; Minieri, M.; Terrinoni, A. Anti-inflammatory and active biological properties of the plant-derived bioactive compounds luteolin and luteolin 7-glucoside. Nutrients 2022, 14, 1155. [Google Scholar] [CrossRef]
- Çetinkaya, M.; Baran, Y. Therapeutic potential of luteolin on cancer. Vaccines 2023, 11, 554. [Google Scholar] [CrossRef]
- Xing, X.; Peng, J.; Zhao, J.; Shi, R.; Wang, C.; Zhang, Z.; Wang, Z.; Li, Z.; Wu, Z. Luteolin regulates the distribution and function of organelles by controlling SIRT1 activity during postovulatory oocyte aging. Front. Nutr. 2023, 10, 1192758. [Google Scholar] [CrossRef]
- Hu, L.L.; Li, H.G.; Liao, B.Y.; Xu, Y.; Sun, S.C.; Wang, J.L. Exposure to nonylphenol impairs oocyte quality via the induction of organelle defects in mice. Ecotoxicol. Environ. Saf. 2022, 230, 113136. [Google Scholar] [CrossRef]
- Chen, C.; Zhou, M.; Ge, Y.; Wang, X. SIRT1 and aging related signaling pathways. Mech. Ageing Dev. 2020, 187, 111215. [Google Scholar] [CrossRef]
- Xie, T.; Yuan, J.; Mei, L.; Li, P.; Pan, R. Luteolin suppresses TNF α induced inflammatory injury and senescence of nucleus pulposus cells via the Sirt6/NF κB pathway. Experiment. Ther. Med. 2022, 24, 1–11. [Google Scholar]
- Zhu, R.Z.; Li, B.S.; Gao, S.S.; Seo, J.H.; Choi, B.M. Luteolin inhibits H2O2-induced cellular senescence via modulation of SIRT1 and p53. Korean J. Physiol. Pharmacol. 2021, 25, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Syed, D.N.; Ahmad, N.; Mukhtar, H. Fisetin: A dietary antioxidant for health promotion. Antioxid. Redox Signal. 2013, 19, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Naeimi, A.F.; Alizadeh, M. Antioxidant properties of the flavonoid fisetin: An updated review of in vivo and in vitro studies. Trends Food Sci. Technol. 2017, 70, 34–44. [Google Scholar] [CrossRef]
- Ravula, A.R.; Teegala, S.B.; Kalakotla, S.; Pasangulapati, J.P.; Perumal, V.; Boyina, H.K. Fisetin, potential flavonoid with multifarious targets for treating neurological disorders: An updated review. Eur. J. Pharmacol. 2021, 910, 174492. [Google Scholar] [CrossRef]
- Currais, A.; Farrokhi, C.; Dargusch, R.; Armando, A.; Quehenberger, O.; Schubert, D.; Maher, P. Fisetin reduces the impact of aging on behavior and physiology in the rapidly aging SAMP8 mouse. J. Gerontol. Ser. A 2018, 73, 299–307. [Google Scholar] [CrossRef]
- Yousefzadeh, M.J.; Zhu, Y.I.; McGowan, S.J.; Angelini, L.; Fuhrmann-Stroissnigg, H.; Xu, M.; Ling, Y.Y.; Melos, K.I.; Pirtskhalava, T.; Inman, C.L.; et al. Fisetin is a senotherapeutic that extends health and lifespan. EBioMedicine 2018, 36, 18–28. [Google Scholar] [CrossRef]
- Mahoney, S.A.; Venkatasubramanian, R.; Darrah, M.A.; Ludwig, K.R.; VanDongen, N.S.; Greenberg, N.T.; Longtine, A.G.; Hutton, D.A.; Brunt, V.E.; Campisi, J.; et al. Intermittent supplementation with fisetin improves arterial function in old mice by decreasing cellular senescence. Aging Cell 2023, 23, e14060. [Google Scholar] [CrossRef]
- Huard, C.A.; Gao, X.; Dey Hazra, M.E.; Dey Hazra, R.O.; Lebsock, K.; Easley, J.T.; Millett, P.J.; Huard, J. Effects of Fisetin treatment on cellular senescence of various tissues and organs of old sheep. Antioxidants 2023, 12, 1646. [Google Scholar] [CrossRef]
- Xing, X.; Liang, Y.; Li, Y.; Zhao, Y.; Zhang, Y.; Li, Z.; Li, Z.; Wu, Z. Fisetin delays postovulatory oocyte aging by regulating oxidative stress and mitochondrial function through Sirt1 pathway. Molecules 2023, 28, 5533. [Google Scholar] [CrossRef]
- Zhao, R.; Kou, H.; Jiang, D.; Wang, F. Exploring the anti-aging effects of fisetin in telomerase-deficient progeria mouse model. PeerJ 2023, 11, e16463. [Google Scholar] [CrossRef]
- Park, S.; Kim, B.K.; Park, S.K. Effects of fisetin, a plant-derived flavonoid, on response to oxidative stress, aging, and age-related diseases in Caenorhabditis elegans. Pharmaceuticals 2022, 15, 1528. [Google Scholar] [CrossRef] [PubMed]
- Brahmachari, G. Discovery and Development of Anti-Breast Cancer Agents from Natural Products: An Overview. In Discovery and Development of Anti-Breast Cancer Agents from Natural Products; Brahmachari, G., Ed.; Elsevier: Amsterdam, The Netherlands, 2021; pp. 1–6. [Google Scholar]
- Periferakis, A.; Periferakis, K.; Badarau, I.A.; Petran, E.M.; Popa, D.C.; Caruntu, A.; Costache, R.S.; Scheau, C.; Caruntu, C.; Costache, D.O. Kaempferol: Antimicrobial properties, sources, clinical, and traditional applications. Int. J. Mol. Sci. 2022, 23, 15054. [Google Scholar] [CrossRef] [PubMed]
- Qattan, M.Y.; Khan, M.I.; Alharbi, S.H.; Verma, A.K.; Al-Saeed, F.A.; Abduallah, A.M.; Al Areefy, A.A. therapeutic importance of kaempferol in the treatment of cancer through the modulation of cell signalling pathways. Molecules 2022, 27, 8864. [Google Scholar] [CrossRef]
- Park, M.J.; Lee, E.K.; Heo, H.S.; Kim, M.S.; Sung, B.; Kim, M.K.; Lee, J.; Kim, N.D.; Anton, S.; Choi, J.S.; et al. The anti-inflammatory effect of kaempferol in aged kidney tissues: The involvement of nuclear factor-kappaB via nuclear factor-inducing kinase/IkappaB kinase and mitogen-activated protein kinase pathways. J. Med. Food 2009, 12, 351–358. [Google Scholar] [CrossRef]
- Tan, B.L.; Norhaizan, M.E. Carotenoids: How effective are they to prevent age-related diseases? Molecules 2019, 24, 1801. [Google Scholar] [CrossRef]
- Zheng, W.V.; Xu, W.; Li, Y.; Qin, J.; Zhou, T.; Li, D.; Xu, Y.; Cheng, X.; Xiong, Y.; Chen, Z. Anti-aging effect of β-carotene through regulating the KAT7-P15 signaling axis, inflammation and oxidative stress process. Cell. Mol. Biol. Lett. 2022, 27, 86. [Google Scholar] [CrossRef]
- Jesus, M.; Martins, A.P.; Gallardo, E.; Silvestre, S. Diosgenin: Recent highlights on pharmacology and analytical methodology. J. Anal. Methods Chem. 2016, 2016, 4156293. [Google Scholar] [CrossRef]
- Semwal, P.; Painuli, S.; Abu-Izneid, T.; Rauf, A.; Sharma, A.; Daştan, S.D.; Kumar, M.; Alshehri, M.M.; Taheri, Y.; Das, R.; et al. Diosgenin: An updated pharmacological review and therapeutic perspectives. Oxid. Med. Cell. Longev. 2022, 2022, 1035441. [Google Scholar] [CrossRef]
- Song, L.; Li, C.; Wu, F.; Zhang, S. Dietary intake of diosgenin delays aging of male fish Nothobranchius guentheri through modulation of multiple pathways that play prominent roles in ROS production. Biogerontology 2022, 23, 201–213. [Google Scholar] [CrossRef]
- Wang, W.; Wang, S.; Liu, Y.; Wang, X.; Nie, J.; Meng, X.; Zhang, Y. Ellagic acid: A dietary-derived phenolic compound for drug discovery in mild cognitive impairment. Front. Aging Neurosci. 2022, 14, 925855. [Google Scholar] [CrossRef]
- Rahimi, B.V.; Askari, V.R.; Mousavi, S.H. Ellagic acid dose and time-dependently abrogates d-galactose-induced animal model of aging: Investigating the role of PPAR-γ. Life Sci. 2019, 232, 116595. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, V.B.; Askari, V.R.; Mousavi, S.H. Ellagic acid reveals promising anti-aging effects against d-galactose-induced aging on human neuroblastoma cell line, SH-SY5Y: A mechanistic study. Biomed. Pharmacother. 2018, 108, 1712–1724. [Google Scholar] [CrossRef] [PubMed]
- Xian, W.; Deng, Y.; Yang, Y.; Tan, Z.; Chen, C.; Li, W.; Yang, R. Ameliorative effect of ellagic acid on aging in rats with the potential mechanism relying on the gut microbiota and urolithin a-producing ability. J. Agric. Food Chem. 2023, 71, 7396–7407. [Google Scholar] [CrossRef]
- Naghibi, N.; Sadeghi, A.; Movahedinia, S.; Rahimi Naiini, M.; Rajizadeh, M.A.; Bahri, F.; Nazari-Robati, M. Ellagic acid ameliorates aging-induced renal oxidative damage through upregulating SIRT1 and NRF2. BMC Complement. Med. Ther. 2023, 23, 77. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iweala, E.J.; Adurosakin, O.E.; Innocent, U.; Omonhinmin, C.A.; Dania, O.E.; Ugbogu, E.A. Anti-Aging Potential of Bioactive Phytoconstituents Found in Edible Medicinal Plants: A Review. Sci 2024, 6, 36. https://doi.org/10.3390/sci6020036
Iweala EJ, Adurosakin OE, Innocent U, Omonhinmin CA, Dania OE, Ugbogu EA. Anti-Aging Potential of Bioactive Phytoconstituents Found in Edible Medicinal Plants: A Review. Sci. 2024; 6(2):36. https://doi.org/10.3390/sci6020036
Chicago/Turabian StyleIweala, Emeka Joshua, Oluwapelumi Eniola Adurosakin, Ugochukwu Innocent, Conrad A. Omonhinmin, Omoremime Elizabeth Dania, and Eziuche Amadike Ugbogu. 2024. "Anti-Aging Potential of Bioactive Phytoconstituents Found in Edible Medicinal Plants: A Review" Sci 6, no. 2: 36. https://doi.org/10.3390/sci6020036
APA StyleIweala, E. J., Adurosakin, O. E., Innocent, U., Omonhinmin, C. A., Dania, O. E., & Ugbogu, E. A. (2024). Anti-Aging Potential of Bioactive Phytoconstituents Found in Edible Medicinal Plants: A Review. Sci, 6(2), 36. https://doi.org/10.3390/sci6020036