Lagos Bat Virus Infection Dynamics in Free-Ranging Straw-Colored Fruit Bats (Eidolon helvum)

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Material and Methods
2.1. Ethics Statement
2.2. Study Sites
2.3. Study Design
Sera Collection for Lagos Bat Virus Antibody Prevalence in Wild Eidolon helvum
2.4. Collection and Examination of Dead Bats
2.5. Laboratory Investigations
2.5.1. Antigen Detection Using the Fluorescent Antibody Test (FAT)
2.5.2. Detection of LBV-Neutralizing Antibodies (mFAVN)
2.5.3. Detection of LBV RNA Using Molecular Methods
3. Statistical Analysis
4. Results
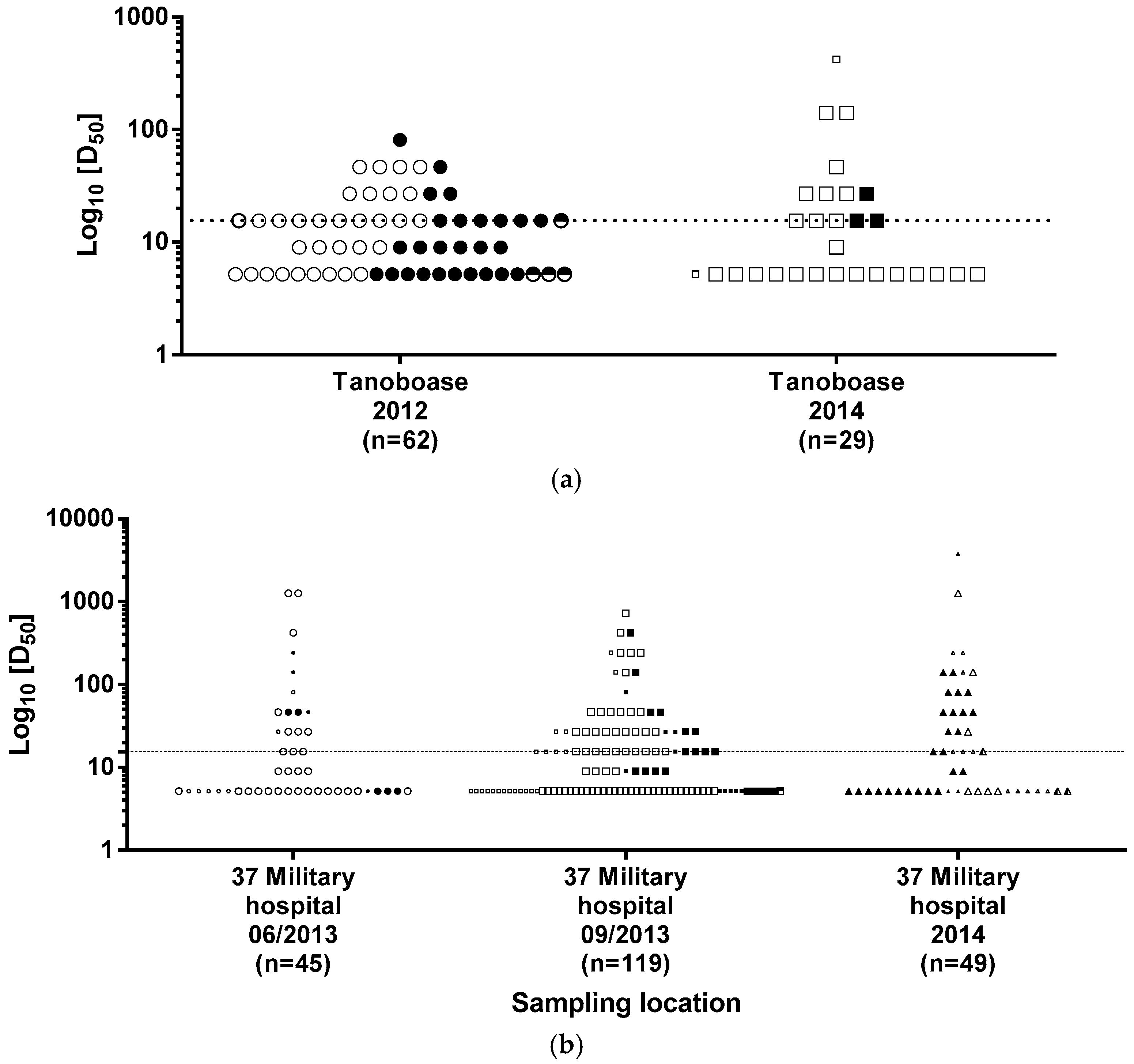
4.1. LBV Seroprevalence
4.2. Necropsy Findings in E. helvum Bats
5. Discussion
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Daszak, P.; Cunningham, A.A.; Hyatt, A.D. Anthropogenic environmental change and the emergence of infectious diseases in wildlife. Acta Trop. 2001, 78, 103–116. [Google Scholar] [CrossRef]
- Hayman, D.T.; Fooks, A.R.; Rowcliffe, J.M.; McCrea, R.; Restif, O.; Baker, K.S.; Horton, D.L.; Suu-Ire, R.D.; Cunningham, A.A.; Wood, J.L.N. Endemic Lagos bat virus infection in Eidolon helvum. Epidemiol. Infect. 2012, 140, 2163–2171. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.E.; Patel, N.G.; Levy, M.A.; Storeygard, A.; Balk, D.; Gittleman, J.L.; Daszak, P. Global trends in emerging infectious diseases. Nat. Biotechnol. 2008, 451, 990–994. [Google Scholar] [CrossRef] [PubMed]
- Calisher, C.H.; Childs, J.E.; Field, H.E.; Holmes, K.V.; Schountz, T. Bats: Important reservoir hosts of emerging viruses. Clin. Microbiol. Rev. 2006, 19, 531–545. [Google Scholar] [CrossRef] [PubMed]
- Taylor, D.A.R.; Kankam, B.O.; Wagner, M.R. The role of fruit bat, Eidolon helvum, in seed dispersal, survival, and germination in Milicia excelsa, a threatened West African hardwood. In Research Advances in Restoration of Iroko as a Commercial Species in West Africa; Cobbinah, J.R., Wagner, M.R., Eds.; Forestry Research Institute of Ghana (FORIG): Kumasi, Ghana, 2000; pp. 29–39. [Google Scholar]
- Luis, A.D.; Hayman, D.T.; O’Shea, T.J.; Cryan, P.M.; Gilbert, A.T.; Pulliam, J.R.; Mills, J.N.; Timonin, M.E.; Willis, C.K.; Cunningham, A.A.; et al. A comparison of bats and rodents as reservoirs of zoonotic viruses: Are bats special? Proc. Biol. Sci. 2013, 280, 20122753. [Google Scholar] [CrossRef] [PubMed]
- Fooks, A.R.; Banyard, A.C.; Horton, D.L.; Johnson, N.; McElhinney, L.M.; Jackson, A.C. Current status of rabies and prospects for elimination. Lancet 2014, 384, 1389–1399. [Google Scholar] [CrossRef]
- Banyard, A.C.; Hayman, D.; Johnson, N.; McElhinney, L.; Fooks, A.R. Bats and lyssaviruses. Adv. Virus Res. 2011, 79, 239–289. [Google Scholar] [PubMed]
- Banyard, A.C.; Fooks, A.R. The impact of novel lyssavirus discovery. Microbiol. Aust. 2017, 38, 18–21. [Google Scholar] [CrossRef]
- Badrane, H.; Tordo, N. Host switching in lyssavirus history from the Chiroptera to the Carnivora orders. J. Virol. 2001, 75, 8096–8104. [Google Scholar] [CrossRef] [PubMed]
- Markotter, W.; Kuzmin, I.; Rupprecht, C.E.; Randles, J.; Sabeta, C.T.; Wandeler, A.I.; Nel, L.H. Isolation of Lagos bat virus from water mongoose. Emerg. Infect. Dis. 2006, 12, 1913–1918. [Google Scholar] [CrossRef] [PubMed]
- Foggin, C.M. Atypical rabies virus in cats and a dog in Zimbabwe. Vet. Rec. 1982, 110, 338. [Google Scholar] [CrossRef] [PubMed]
- Mebatsion, T.; Cox, J.H.; Frost, J.W. Isolation and characterization of 115 street rabies virus isolates from Ethiopia by using monoclonal antibodies: identification of 2 isolates as Mokola and Lagos bat viruses. J. Infect. Dis. 1992, 166, 972–977. [Google Scholar] [CrossRef] [PubMed]
- King, A.; Crick, J. Rabies-related viruses. In Rabies; Campbell, J.B., Charlton, K.M., Eds.; Kluwer Academic Publishers: Boston, MA, USA, 1988; pp. 177–200. [Google Scholar]
- Hayman, D.T.; Fooks, A.R.; Horton, D.; Suu-Ire, R.; Breed, A.C.; Cunningham, A.A.; Wood, J.L. Antibodies against Lagos bat virus in megachiroptera from West Africa. Emerg. Infect. Dis. 2008, 14, 926–928. [Google Scholar] [CrossRef] [PubMed]
- Dean, D.J.; Abelseth, M.K. Laboratory techniques in rabies: The fluorescent antibody test. Monogr. Ser. World Health Organ. 1973, 23, 73–84. [Google Scholar]
- Boulger, L.R.; Porterfield, J.S. Isolation of a virus from Nigerian fruit bats. Trans. R. Soc. Trop. Med. Hyg. 1958, 52, 421–424. [Google Scholar] [CrossRef]
- Hayman, D.T.; Johnson, N.; Horton, D.L.; Hedge, J.; Wakeley, P.R.; Banyard, A.C.; Zhang, S.; Alhassan, A.; Fooks, A.R. Evolutionary history of rabies in Ghana. PLoS Negl. Trop. Dis. 2011, 5, e1001. [Google Scholar] [CrossRef] [PubMed]
- Heaton, P.R.; McElhinney, L.M.; Lowings, J.P. Detection and identification of rabies and rabies-related viruses using rapid-cycle PCR. J. Virol. Methods 1999, 81, 63–69. [Google Scholar] [CrossRef]
- Kuzmin, I.V.; Niezgoda, M.; Franka, R.; Agwanda, B.; Markotter, W.; Beagley, J.C.; Urazova, O.Y.; Breiman, R.F.; Rupprecht, C.E. Lagos bat virus in Kenya. J. Clin. Microbiol. 2008, 46, 1451–1461. [Google Scholar] [CrossRef] [PubMed]
- Fahr, J.; Abedi-Lartey, M.; Esch, T.; Machwitz, M.; Suu-Ire, R.; Wikelski, M.; Dechmann, D.K.N. Pronounced seasonal changes in the movement ecology of a highly gregarious central-place forager, the African straw-coloured fruit bat (Eidolon helvum). PLoS ONE 2015, 10, e0138985. [Google Scholar] [CrossRef] [PubMed]
- O’Shea, T.J.; Shankar, V.; Bowen, R.A.; Rupprecht, C.E.; Wimsatt, J.H. Do bats acquire immunity to rabies? Evidence from the field. Bat Res. News 2003, 44, 161. [Google Scholar]
- Bowen, R.A.; O’Shea, T.J.; Shankar, V.; Neubaum, M.A.; Neubaum, D.J.; Rupprecht, C. Prevalence of neutralizing antibodies to rabies virus in serum of seven species of insectivorous bats from Colorado and New Mexico. J. Wildl. Dis. 2013, 49, 367–374. [Google Scholar] [CrossRef] [PubMed]
- Hayman, D.T.S.; Bowen, A.R.; Cryan, P.M.; McCracken, G.F.; O’Shea, T.J.; Peel, A.J.; Gilbert, A.; Webb, C.T.; Wood, J.L.N. Ecology of zoonotic infectious diseases in bats: Current knowledge and future directions. Zoonoses Public Health 2013, 60, 2–21. [Google Scholar] [CrossRef] [PubMed]
- Amengual, B.; Bourhy, H.; Lopez-Roig, M.; Serra-Cobo, J. Temporal dynamics of European bat lyssavirus type 1 and survival of Myotis myotis bats in natural colonies. PLoS ONE 2007, 2, e566. [Google Scholar] [CrossRef] [PubMed]
- Turmelle, A.S.; Jackson, F.R.; Green, D.; McCracken, G.F.; Rupprecht, C.E. Host immunity to repeated rabies virus infection in big brown bats. J. Gen. Virol. 2010, 91, 2360–2366. [Google Scholar] [CrossRef] [PubMed]
- Steece, R.; Altenbach, J.S. Prevalence of rabies specific antibodies in the Mexican free-tailed bat (Tadarida brasiliensis mexicana) at Lava Cave, New Mexico. J. Wildl. Dis. 1989, 25, 490–496. [Google Scholar] [CrossRef] [PubMed]
- Kamins, A.O.; Rowcliffe, J.M.; Ntiamoa-Baidu, Y.; Cunningham, A.A.; Wood, J.L.N.; Restif, O. Characteristics and risk perceptions of Ghanaians potentially exposed to bat-borne zoonoses through bushmeat. Ecohealth 2015, 12, 104–120. [Google Scholar] [CrossRef] [PubMed]
- Freuling, C.M.; Binger, T.; Beer, M.; Adu-Sarkodie, Y.; Schatz, J.; Fischer, M.; Hanke, D.; Hoffmann, B.; Hoper, D.; Mettenleiter, T.C.; et al. Lagos bat virus transmission in an Eidolon helvum bat colony, Ghana. Virus Res. 2015, 210, 42–45. [Google Scholar] [CrossRef] [PubMed]
- Hayman, D.T.; Emmerich, P.; Yu, M.; Wang, L.F.; Suu-Ire, R.; Fooks, A.R.; Cunningham, A.A.; Wood, J.L. Long-term survival of an urban fruit bat seropositive for Ebola and Lagos bat viruses. PLoS ONE 2010, 5, e11978. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.S.; Horton, D.L.; Easton, A.J.; Fooks, A.R.; Banyard, A.C. Rabies virus vaccines: Is there a need for a pan-lyssavirus vaccine? Vaccine 2012, 30, 7447–7454. [Google Scholar] [CrossRef] [PubMed]
- Badrane, H.; Bahloul, C.; Perrin, P.; Tordo, N. Evidence of two Lyssavirus phylogroups with distinct pathogenicity and immunogenicity. J. Virol. 2001, 75, 3268–3276. [Google Scholar] [CrossRef] [PubMed]
- Hanlon, C.A.; Kuzmin, I.V.; Blanton, J.D.; Weldon, W.C.; Manangan, J.S.; Rupprecht, C.E. Efficacy of rabies biologics against new lyssaviruses from Eurasia. Virus Res. 2005, 111, 44–54. [Google Scholar] [CrossRef] [PubMed]
- Horton, D.L.; Banyard, A.C.; Marston, D.A.; Wise, E.; Selden, D.; Nunez, A.; Hicks, D.; Lembo, T.; Cleaveland, S.; Peel, A.J.; et al. Antigenic and genetic characterization of a divergent African virus, Ikoma lyssavirus. J. Gen. Virol. 2014, 95, 1025–1032. [Google Scholar] [CrossRef] [PubMed]
- Nolden, T.; Banyard, A.C.; Finke, S.; Fooks, A.R.; Hanke, D.; Hoper, D.; Horton, D.L.; Mettenleiter, T.C.; Muller, T.; Teifke, J.P.; et al. Comparative studies on the genetic, antigenic and pathogenic characteristics of Bokeloh bat lyssavirus. J. Gen. Virol. 2014, 95, 1647–1653. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| 37-Military Hospital | Tanoboase | Totals | |||||
|---|---|---|---|---|---|---|---|
| Sampling Date | 10-06-2013 | 04-09-2013 | 14-07-2014 | 17-12-2012 | 01-07-2014 | ||
| Age | Adult | 34 | 87 | 32 | 57 | 26 | 236 |
| Juvenile | 11 | 6 | 3 | 0 | 2 | 22 | |
| Sub-adult | 0 | 25 | 11 | 0 | 0 | 36 | |
| Not determined | 0 | 1 | 3 | 5 | 1 | 10 | |
| Sex | Male | 34 | 88 | 30 | 33 | 25 | 210 |
| Female | 11 | 30 | 16 | 24 | 3 | 84 | |
| Unsexed | 1 | 3 | 5 | 1 | 10 | ||
| Serostatus | Positive | 17 | 53 | 24 | 29 | 13 | 136 |
| Negative | 28 | 66 | 25 | 33 | 16 | 168 | |
| Total Sampled | Total Positive (% Positive) | |
|---|---|---|
| Adult | 136 | 109 (80.15%) |
| Juvenile | 58 | 25 (43.10%) |
| Total | 294 | 134 (45.57%) |
| Positive (+ve) | Negative (−ve) | Total Tested | |
|---|---|---|---|
| Dry Season | 29 (21%) | 33 (20%) | 62 (20%) |
| Rainy Season | 107 (79%) | 135 (80%) | 242 (80%) |
| Totals | 136 (45%) | 168 (55%) | 304 |
| Bat ID | Location | Date Found | Condition | Comments |
|---|---|---|---|---|
| A1 | 37-Hospital | 20/01/2013 | Found dead | N.A.D. * |
| A2 | 37-Hospital | 20/02/2013 | Found dead | N.A.D. |
| A3 | 37-Hospital | 20/02/2013 | Found dead | N.A.D. |
| A4 | 37-Hospital | 20/02/2013 | Found dead | N.A.D. |
| A6 | 37-Hospital | 28/10/2013 | Found dead | Very enlarged spleen |
| A5 | Accra Ridge | 29/09/2013 | Found dead | Found dead |
| A12 | 37-Hospital | 27/11/2013 | Found sick | Paralysed |
| A13 | 37-Hospital | 27/11/2013 | Found dead | Fractured right arm |
| A7 | 37-Hospital | 07/12/2013 | Found sick | Paralysis |
| A8 | 37-Hospital | 07/12/2013 | Found sick | Paralysis |
| A9 | 37-Hospital | 07/12/2013 | Found dead | Abscess on neck |
| A10 | 37-Hospital | 10/12/2013 | Found dead | N.A.D. |
| A11 | 37-Hospital | 08/12/2013 | Found dead | Fractured forearm |
| A14 | 37-Hospital | 11/12/2013 | Found dead | Fractured forearm |
| A15 | 37-Hospital | 11/12/2013 | Found dead | Severe intestinal, abdominal haemorrhages |
| A16 | 37-Hospital | 11/12/2013 | Found dead | Fractured head |
| A17 | 37-Hospital | 13/12/2013 | Found sick | Paralysis |
| A18 | 37-Hospital | 13/12/2013 | Found dead | N.A.D. |
| A19 | 37-Hospital | 13/12/2013 | Found dead | Punctured wing |
| A22 | 37-Hospital | 22/12/2013 | Found sick | Paralysis, severe haemorrhages on head |
| A23 | 37-Hospital | 22/12/2013 | Found dead | N.A.D. |
| A24 | 37-Hospital | 22/12/2013 | Found dead | N.A.D. |
| A20 | 37-Hospital | 03/01/2014 | Found dead | N.A.D. |
| A21 | 37-Hospital | 03/01/2014 | Found dead | Emaciated carcass |
| A25 | 37-Hospital | 22/01/2014 | Found dead | Fractured forearm |
| A26 | 37-Hospital | 22/01/2014 | Found dead | Decomposed carcass |
| A27 | 37-Hospital | 22/01/2014 | Found dead | N.A.D. |
| A28 | 37-Hospital | 29/01/2014 | Found dead | N.A.D. |
| A29 | 37-Hospital | 05/02/2014 | Found dead | N.A.D. |
| A30 | 37-Hospital | 12/03/2014 | Found dead | N.A.D. |
| A31 | 37-Hospital | 12/03/2014 | Found dead | N.A.D. |
| A32 | 37-Hospital | 12/03/2014 | Found dead | N.A.D. |
| A33 | 37-Hospital | 12/03/2014 | Found dead | N.A.D. |
| A34 | 37-Hospital | 12/03/2014 | Found dead | N.A.D. |
| A35 | 37-Hospital | 12/03/2014 | Found dead | N.A.D. |
| A36 | 37-Hospital | 06/04/2014 | Found sick | Paralysis, emaciated |
| A37 | 37-Hospital | 06/04/2014 | Found sick | Paralysis |
| A38 | Achimota | 09/07/2014 | Found dead | N.A.D. |
| T1 | Tanoboase | 22/01/2013 | Found dead | N.A.D. |
| T2 | Tanoboase | 22/01/2013 | Found sick | Fractured forearm |
| T3 | Tanoboase | 22/01/2013 | Found sick | Fractured forearm |
| T4 | Tanoboase | 23/01/2013 | Found dead | Multiple body injury |
| T5 | Tanoboase | 23/01/2013 | Found sick | Fractured forearm |
| T6 | Tanoboase | 07/03/2013 | Found sick | Weak |
| T7 | Tanoboase | 07/03/2013 | Found sick | Multiple body injury |
| T8 | Tanoboase | 07/03/2013 | Found sick | Weak |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suu-Ire, R.D.; Fooks, A.R.; Banyard, A.C.; Selden, D.; Amponsah-Mensah, K.; Riesle, S.; Ziekah, M.Y.; Ntiamoa-Baidu, Y.; Wood, J.L.N.; Cunningham, A.A. Lagos Bat Virus Infection Dynamics in Free-Ranging Straw-Colored Fruit Bats (Eidolon helvum). Trop. Med. Infect. Dis. 2017, 2, 25. https://doi.org/10.3390/tropicalmed2030025
Suu-Ire RD, Fooks AR, Banyard AC, Selden D, Amponsah-Mensah K, Riesle S, Ziekah MY, Ntiamoa-Baidu Y, Wood JLN, Cunningham AA. Lagos Bat Virus Infection Dynamics in Free-Ranging Straw-Colored Fruit Bats (Eidolon helvum). Tropical Medicine and Infectious Disease. 2017; 2(3):25. https://doi.org/10.3390/tropicalmed2030025
Chicago/Turabian StyleSuu-Ire, Richard D., Anthony R. Fooks, Ashley C. Banyard, David Selden, Kofi Amponsah-Mensah, Silke Riesle, Meyir Y. Ziekah, Yaa Ntiamoa-Baidu, James L. N. Wood, and Andrew A. Cunningham. 2017. "Lagos Bat Virus Infection Dynamics in Free-Ranging Straw-Colored Fruit Bats (Eidolon helvum)" Tropical Medicine and Infectious Disease 2, no. 3: 25. https://doi.org/10.3390/tropicalmed2030025
APA StyleSuu-Ire, R. D., Fooks, A. R., Banyard, A. C., Selden, D., Amponsah-Mensah, K., Riesle, S., Ziekah, M. Y., Ntiamoa-Baidu, Y., Wood, J. L. N., & Cunningham, A. A. (2017). Lagos Bat Virus Infection Dynamics in Free-Ranging Straw-Colored Fruit Bats (Eidolon helvum). Tropical Medicine and Infectious Disease, 2(3), 25. https://doi.org/10.3390/tropicalmed2030025