
Seasonal Phenological Patterns and Flavivirus Vectorial Capacity of Medically Important Mosquito Species in a Wetland and an Urban Area of Attica, Greece



 ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
- (i)
- Monitoring and recording of mosquitoes’ species and population densities in the RUs under study; and
- (ii)
- detection and monitoring of the circulation of WNV and USUV in collected Cx. pipiens s.l. for possible co-circulation.
2. Materials and Methods
2.1. Study Area
2.2. Mosquito Collection and Identification
2.3. Flaviviruses Survey in Culex pipiens Pools
2.4. WNV and USUV Detection
2.5. Infection Rates
3. Molecular Methods for Identification of Anopheles Mosquitoes
Data Analysis
4. Results
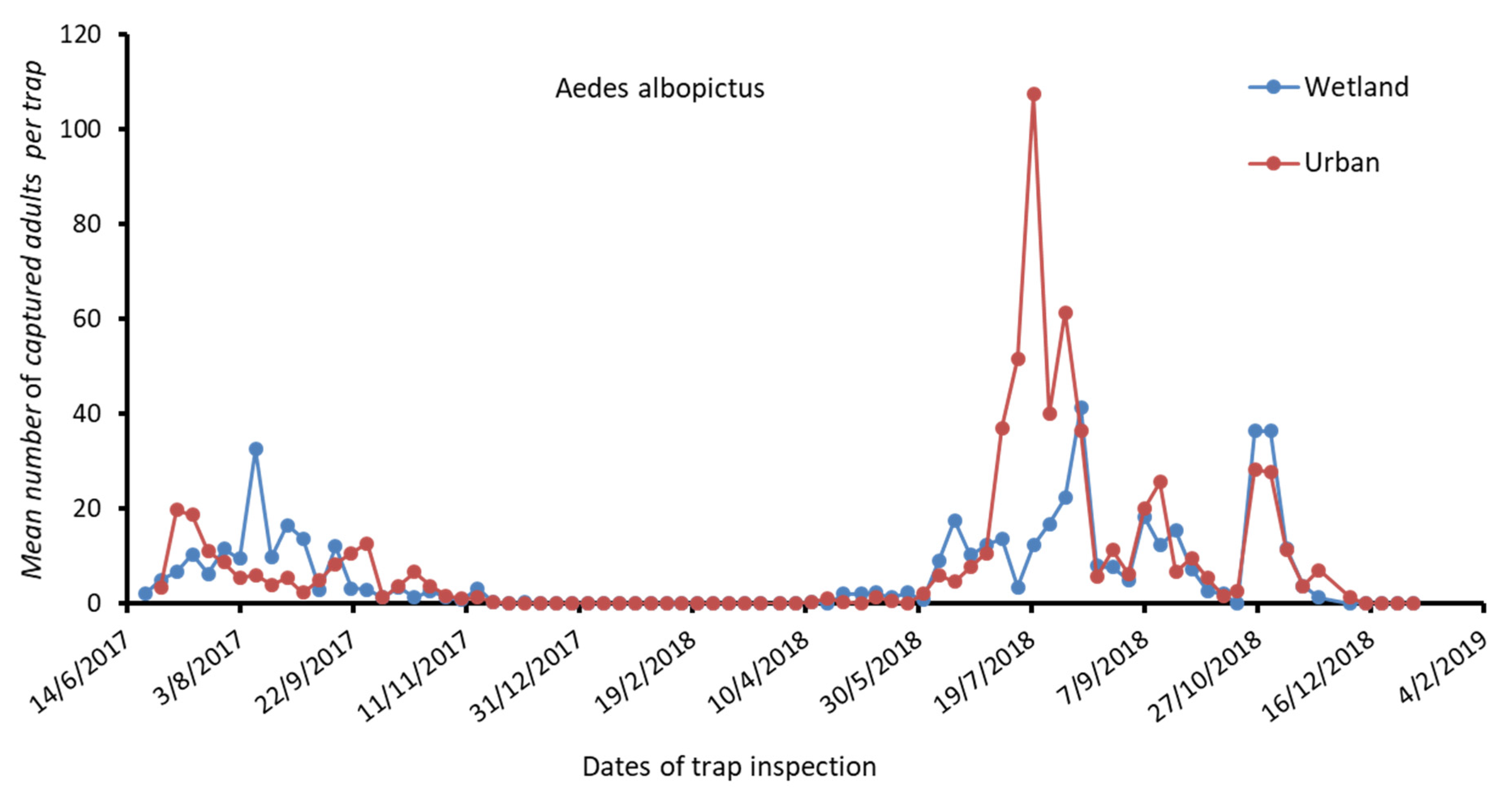
4.1. Mosquito Fauna Identification
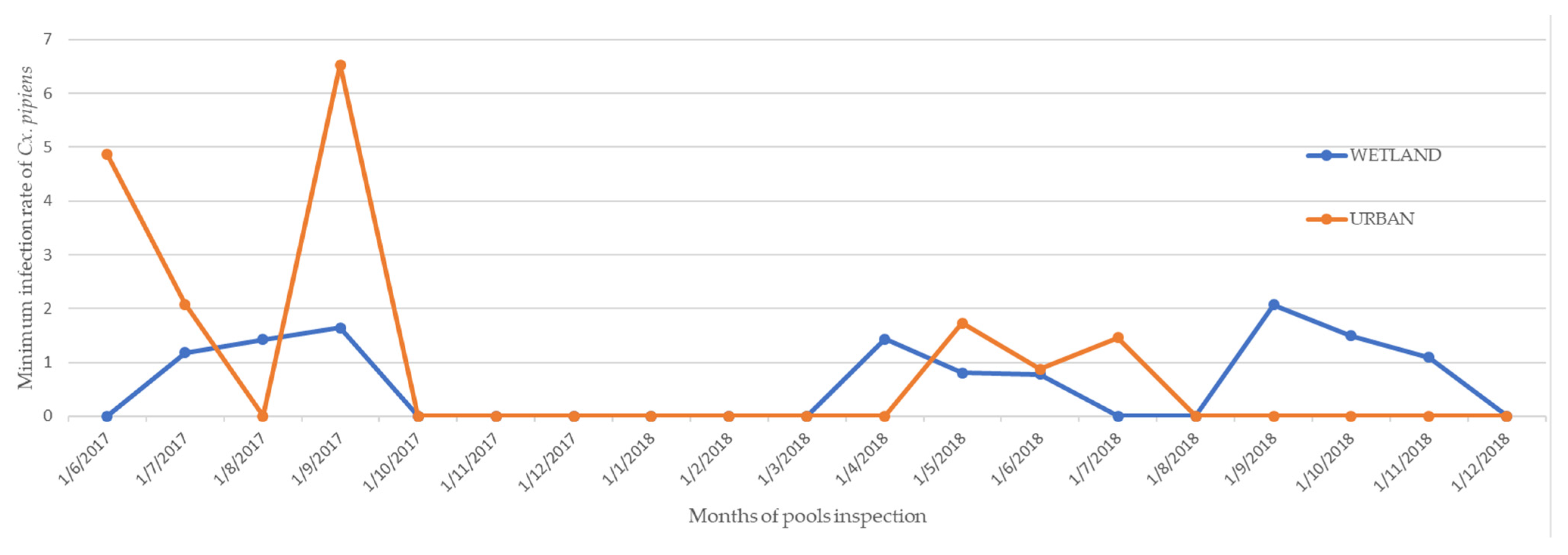
4.2. Flaviviruses Detection
Infection Rates
4.3. Anopheles Specimens’ Molecular Identification
5. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- European Centre for Disease Prevention and Control (ECDC). Guidelines for the Surveillance of Native Mosquitoes in Europe. Stockholm: ECDC. 2014. Available online: https://ecdc.europa.eu/sites/portal/files/media/en/publications/Publications/surveillance-of%20native-mosquitoes%20-guidelines.pdf (accessed on 20 March 2020).
- Ferraguti, M.; Martínez-de la Puente, J.; Roiz, D.; Ruiz, S.; Soriguer, R.; Figuerola, J. Effects of landscape anthropization on mosquito community composition and abundance. Sci. Rep. 2016, 6, 29002. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Arrondo, I.; McMahon, B.J.; Hernández-Triana, L.M.; Santibañez, P.; Portillo, A.; Oteo, J.A. Surveillance of mosquitoes (Diptera, Culicidae) in a northern central region of Spain: Implications for the medical community. Front. Vet. Sci. 2019, 6, 86. [Google Scholar] [CrossRef]
- Török, E.; Tomazatos, A.; Cadar, D.; Horváth, C.; Keresztes, L.; Jansen, S.; Becker, N.; Kaiser, A.; Popescu, O.; Schmidt-Chanasit, J.; et al. Pilot longitudinal mosquito surveillance study in the Danube Delta Biosphere Reserve and the first reports ofAnopheles algeriensis Theobald, 1903 and Aedes hungaricus Mihalyi, 1955 for Romania. Parasit. Vectors 2016, 9, 196. [Google Scholar] [CrossRef] [Green Version]
- Barzon, L. Ongoing and emerging arbovirus threats in Europe. J. Clin. Virol. 2018, 107, 38–47. [Google Scholar] [CrossRef]
- Martina, B.E.; Barzon, L.; Pijlman, G.P.; de la Fuente, J.; Rizzoli, A.; Wammes, L.J.; Takken, W.; van Rij, R.P.; Papa, A. Human to human transmission of arthropod-borne pathogens. Curr. Opin. Virol. 2017, 22, 13–21. [Google Scholar] [CrossRef]
- Weaver, S.C.; Reisen, W.K. Present and future arboviral threats. Antivir. Res. 2010, 85, 328–345. [Google Scholar] [CrossRef] [Green Version]
- Chaskopoulou, A.; Dovas, C.I.; Chaintoutis, S.C.; Kashefi, J.; Koehler, P.; Papanastassopoulou, M. Detection and Early Warning of West Nile Virus Circulation in Central Macedonia, Greece, Using Sentinel Chickens and Mosquitoes. Vector-Borne Zoonotic Dis. 2013, 13, 723–732. [Google Scholar] [CrossRef]
- Mavridis, K.; Fotakis, E.A.; Kioulos, I.; Mpellou, S.; Konstantas, S.; Varela, E.; Gewehr, S.; Diamantopoulos, V.; Vontas, J. Detection of West Nile Virus—Lineage 2 in Culex pipiens mosquitoes, associated with disease outbreak in Greece, 2017. Acta Trop. 2018, 182, 64–68. [Google Scholar] [CrossRef]
- Papa, A.; Gewehr, S.; Tsioka, K.; Kalaitzopoulou, S.; Pappa, S.; Mourelatos, S. Detection of flaviviruses and alphaviruses in mosquitoes in Central Macedonia, Greece, 2018. Acta Trop. 2020, 202, 105278. [Google Scholar] [CrossRef]
- Papa, A.; Tsioka, K.; Gewehr, S.; Kalaitzopoulou, S.; Pappa, S.; Mourelatos, S. West Nile virus lineage 2 in Culex mosquitoes in Thessaly, Greece, 2019. Acta Trop. 2020, 208, 105514. [Google Scholar] [CrossRef]
- Papa, A.; Tsioka, K.; Gewehr, S.; Kalaitzopouou, S.; Pervanidou, D.; Vakali, A.; Kefaloudi, C.; Pappa, S.; Louka, X.; Mourelatos, S. West Nile fever upsurge in a Greek regional unit, 2020. Acta Trop. 2021, 221, 106010. [Google Scholar] [CrossRef] [PubMed]
- Patsoula, E.; Vakali, A.; Balatsos, G.; Pervanidou, D.; Beleri, S.; Tegos, N.; Baka, A.; Spanakos, G.; Georgakopoulou, T.; Tserkezou, P.; et al. West Nile Virus Circulation in Mosquitoes in Greece (2010–2013). BioMed Res. Int. 2016, 2016, 2450682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patsoula, E.; Beleri, S.; Tegos, N.; Mkrtsian, R.; Vakali, A.; Pervanidou, D. Entomological Data and Detection of West Nile Virus in Mosquitoes in Greece (2014–2016), Before Disease Re-Emergence in 2017. Vector-Borne Zoonotic Dis. 2020, 20, 60–70. [Google Scholar] [CrossRef]
- Čabanová, V.; Šikutová, S.; Straková, P.; Šebesta, O.; Vichová, B.; Zubríková, D.; Miterpakova, M.; Mendel, J.; Hurníková, Z.; Hubálek, Z.; et al. Co-Circulation of West Nile and Usutu Flaviviruses in Mosquitoes in Slovakia, 2018. Viruses 2019, 11, 639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christova, I.; Papa, A.; Trifonova, I.; Panayotova, E.; Pappa, S.; Mikov, O. West Nile virus lineage 2 in humans and mosquitoes in Bulgaria, 2018–2019. J. Clin. Virol. 2020, 127, 104365. [Google Scholar] [CrossRef] [PubMed]
- Fros, J.; Miesen, P.; Vogels, C.; Gaibani, P.; Sambri, V.; Martina, B.E.; Koenraadt, C.J.; van Rij, R.P.; Vlak, J.M.; Takken, W.; et al. Comparative Usutu and West Nile virus transmission potential by local Culex pipiens mosquitoes in north-western Europe. One Health 2015, 1, 31–36. [Google Scholar] [CrossRef] [Green Version]
- Bisia, M.; Jeffries, C.L.; Lytra, I.; Michaelakis, A.; Walker, T. A Comparison of Adult Mosquito Trapping Methods to Assess Potential West Nile Virus Mosquito Vectors in Greece during the Onset of the 2018 Transmission Season. Insects 2020, 11, 329. [Google Scholar] [CrossRef]
- Marka, A.; Diamantidis, A.; Papa, A.; Valiakos, G.; Chaintoutis, S.C.; Doukas, D.; Tserkezou, P.; Giannakopoulos, A.; Papaspyropoulos, K.; Patsoula, E.; et al. West Nile Virus State of the Art Report of MALWEST Project. Int. J. Environ. Res. Public Health 2013, 10, 6534–6610. [Google Scholar] [CrossRef]
- Calzolari, M.; Monaco, F.; Montarsi, F.; Bonilauri, P.; Ravagnan, S.; Bellini, R.; Cattoli, G.; Cordioli, P.; Cazzin, S.; Pinoni, C.; et al. New incursions of West Nile virus lineage 2 in Italy in 2013: The value of the entomological surveillance as early warning system. Vet. Ital. 2013, 49, 315–319. [Google Scholar]
- Riccardo, F.; Monaco, F.; Bella, A.; Savini, G.; Russo, F.; Cagarelli, R.; Dottori, M.; Rizzo, C.; Venturi, G.; Di Luca, M.; et al. An early start of West Nile virus seasonal transmission: The added value of One Heath surveillance in detecting early circulation and triggering timely response in Italy, June to July 2018. Eurosurveill 2018, 23, 1800427. [Google Scholar] [CrossRef] [Green Version]
- Bellini, R.; Zeller, H.; Van Bortel, W. A review of the vector management methods to prevent and control outbreaks of West Nile virus infection and the challenge for Europe. Parasites Vectors 2014, 7, 323. [Google Scholar] [CrossRef] [Green Version]
- Gossner, C.M.; Marrama, L.; Carson, M.; Allerberger, F.; Calistri, P.; Dilaveris, D.; Lecollinet, S.; Morgan, D.; Nowotny, N.; Paty, M.C.; et al. West Nile virus surveillance in Europe: Moving towards an integrated animal-human-vector approach. Eurosurveill 2017, 22, 30526. [Google Scholar] [CrossRef] [PubMed]
- Jourdain, F.; Samy, A.M.; Hamidi, A.; Bouattour, A.; Alten, B.; Faraj, C.; Roiz, D.; Petrić, D.; Pérez-Ramírez, E.; Velo, E.; et al. Towards harmonisation of entomological surveillance in the Mediterranean area. PLoS Negl. Trop. Dis. 2019, 13, e0007314. [Google Scholar] [CrossRef] [PubMed]
- Calisher, C.H.; Gould, E.A. Taxonomy of the virus family Flaviviridae. Adv. Virus Res. 2003, 59, 1–19. [Google Scholar] [PubMed]
- Calzolari, M.; Bonilauri, P.; Bellini, R.; Albieri, A.; Defilippo, F.; Maioli, G.; Galletti, G.; Gelati, A.; Barbieri, I.; Tamba, M.; et al. Evidence of Simultaneous Circulation of West Nile and Usutu Viruses in Mosquitoes Sampled in Emilia-Romagna Region (Italy) in 2009. PLoS ONE 2010, 5, e14324. [Google Scholar] [CrossRef] [Green Version]
- Papa, A. Emerging arboviruses of medical importance in the Mediterranean region. J. Clin. Virol. 2019, 115, 5–10. [Google Scholar] [CrossRef]
- Zannoli, S.; Sambri, V. West Nile Virus and Usutu Virus Co-Circulation in Europe: Epidemiology and Implications. Microorganisms 2019, 7, 184. [Google Scholar] [CrossRef] [Green Version]
- Hubálek, Z.; Halouzka, J. West Nile Fever–a Reemerging Mosquito-Borne Viral Disease in Europe. Emerg. Infect. Dis. 1999, 5, 643–650. [Google Scholar] [CrossRef]
- Chancey, C.; Grinev, A.; Volkova, E.; Rios, M. The Global Ecology and Epidemiology of West Nile Virus. BioMed Res. Int. 2015, 2015, 376230. [Google Scholar] [CrossRef] [Green Version]
- Haussig, J.M.; Young, J.J.; Gossner, C.M.; Mezei, E.; Bella, A.; Sirbu, A.; Pervanidou, D.; Drakulovic, M.B.; Sudre, B. Early start of the West Nile fever transmission season 2018 in Europe. Eurosurveill 2018, 23, 1800428. [Google Scholar] [CrossRef] [Green Version]
- Martínez-de la Puente, J.; Ferraguti, M.; Ruiz, S.; Roiz, D.; Llorente, F.; Pérez-Ramírez, E.; Jiménez-Clavero, M.A.; Soriguer, R.; Figuerola, J. Mosquito community influences West Nile virus seroprevalence in wild birds: Implications for the risk of spillover into human populations. Sci. Rep. 2018, 8, 2599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaintoutis, S.C.; Chaskopoulou, A.; Chassalevris, T.; Koehler, P.G.; Papanastassopoulou, M.; Dovas, C.I. West Nile Virus Lineage 2 Strain in Greece, 2012. Emerg. Infect. Dis. 2013, 19, 827–829. [Google Scholar] [CrossRef] [PubMed]
- Danis, K.; Papa, A.; Theocharopoulos, G.; Dougas, G.; Athanasiou, M.; Detsis, M.; Baka, A.; Lytras, T.; Mellou, K.; Bonovas, S.; et al. Outbreak of West Nile virus infection in Greece, 2010. Emerg. Infect. Dis. 2011, 17, 1868–1872. [Google Scholar] [CrossRef]
- Hadjichristodoulou, C.; Pournaras, S.; Mavrouli, M.; Marka, A.; Tserkezou, P.; Baka, A.; Billinis, C.; Katsioulis, A.; Psaroulaki, A.; Papa, A.; et al. West Nile Virus Seroprevalence in the Greek Population in 2013: A Nationwide Cross-Sectional Survey. PLoS ONE 2015, 10, e0143803. [Google Scholar] [CrossRef] [Green Version]
- MALWEST. West Nile Virus, Epidemiology. Available online: http://www.malwest.gr/en-us/westnilevirus/informationforhealthcareprofessionals/epidemiology.aspx (accessed on 27 May 2020).
- European Centre for Disease Prevention and Control (ECDC). Historical Data by Year—West Nile Fever Seasonal Surveillance. Available online: https://www.ecdc.europa.eu/en/west-nile-fever/surveillance-and-disease-data/historical (accessed on 20 March 2020).
- Pervanidou, D.; Vakali, A.; Georgakopoulou, T.; Panagiotopoulos, T.; Patsoula, E.; Koliopoulos, G.; Politis, C.; Stamoulis, K.; Gavana, E.; Pappa, S.; et al. West Nile virus in humans, Greece, 2018: The largest seasonal number of cases, 9 years after its emergence in the country. Eurosurveillance 2020, 25, 1900543. [Google Scholar] [CrossRef] [PubMed]
- Papa, A.; Papadopoulou, E.; Chatzixanthouliou, C.; Glouftsios, P.; Pappa, S.; Pervanidou, D.; Georgiou, L. Emergence of West Nile virus lineage 2 belongin g to the Eastern European subclade, Greece. Arch Virol. 2019, 164, 1673–1675. [Google Scholar] [CrossRef]
- Gaibani, P.; Rossini, G. An overview of Usutu virus. Microbes Infect. 2017, 19, 382–387. [Google Scholar] [CrossRef]
- Nikolay, B.; Diallo, M.C.; Boye, S.; Sall, A.A. Usutu virus in Africa. Vector Borne Zoonotic Dis. 2011, 11, 1417–1423. [Google Scholar] [CrossRef]
- Vázquez, A.; Jimenez-Clavero, M.; Franco, L.; Donoso-Mantke, O.; Sambri, V.; Niedrig, M.; Zeller, H.; Tenorio, A. Usutu virus: Potential risk of human disease in Europe. Eurosurveill 2011, 16, 19935. [Google Scholar] [CrossRef]
- Clé, M.; Beck, C.; Salinas, S.; Lecollinet, S.; Gutierrez, S.; Van de Perre, P.; Baldet, T.; Foulongne, V.; Simonin, Y. Usutu virus: A new threat? Epidemiol. Infect. 2019, 147, e232. [Google Scholar] [CrossRef] [Green Version]
- Roesch, F.; Fajardo, A.; Moratorio, G.; Vignuzzi, M. Usutu Virus: An Arbovirus on the Rise. Viruses 2019, 11, 640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michel, F.; Sieg, M.; Fischer, D.; Keller, M.; Eiden, M.; Reuschel, M.; Schmidt, V.; Schwehn, R.; Rinder, M.; Urbaniak, S.; et al. Evidence for West Nile Virus and Usutu Virus Infections in Wild and Resident Birds in Germany, 2017 and 2018. Viruses 2019, 11, 674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weissenböck, H.; Bakonyi, T.; Rossi, G.; Mani, P.; Nowotny, N. Usutu virus, Italy, 1996. Emerg. Infect. Dis. 2013, 19, 274–277. [Google Scholar] [CrossRef] [PubMed]
- Vilibic-Cavlek, T.; Petrovic, T.; Savic, V.; Barbic, L.; Tabain, I.; Stevanovic, V.; Klobucar, A.; Mrzljak, A.; Ilic, M.; Bogdanic, M.; et al. Epidemiology of Usutu Virus: The European Scenario. Pathogens 2020, 9, 699. [Google Scholar] [CrossRef] [PubMed]
- Lühken, R.; Jöst, H.; Cadar, D.; Thomas, S.M.; Bosch, S.; Tannich, E.; Becker, N.; Ziegler, U.; Lachmann, L.; Schmidt-Chanasit, J. Distribution of Usutu Virus in Germany and Its Effect on Breeding Bird Populations. Emerg. Infect. Dis. 2017, 23, 1994–2001. [Google Scholar] [CrossRef] [Green Version]
- Oude Munnink, B.B.; Münger, E.; Nieuwenhuijse, D.F.; Kohl, R.; van der Linden, A.; Schapendonk, C.M.E.; van der Jeugd, H.; Kik, M.; Rijks, J.M.; Reusken, C.B.E.M.; et al. Genomic monitoring to understand the emergence and spread of Usutu virus in the Netherlands, 2016–2018. Sci. Rep. 2020, 10, 2798. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, U.; Ye, J.; Ruan, X.; Wan, S.; Zhu, B.; Cao, S. Usutu Virus: An Emerging Flavivirus in Europe. Viruses 2015, 7, 219–238. [Google Scholar] [CrossRef] [Green Version]
- Nikolay, B. A review of West Nile and Usutu virus co-circulation in Europe: How much do transmission cycles overlap? Trans. R. Soc. Trop. Med. Hyg. 2015, 109, 609–618. [Google Scholar] [CrossRef] [Green Version]
- Bakonyi, T.; Erdélyi, K.; Brunthaler, R.; Dán, Á.; Weissenböck, H.; Nowotny, N. Usutu virus, Austria and Hungary, 2010–2016. Emerg. Microbes Infect. 2017, 6, e85. [Google Scholar] [CrossRef] [Green Version]
- Benzarti, E.; Sarlet, M.; Franssen, M.; Cadar, D.; Schmidt-Chanasit, J.; Rivas, J.F.; Linden, A.; Desmecht, D.; Garigliany, M. Usutu Virus Epizootic in Belgium in 2017 and 2018: Evidence of Virus Endemization and Ongoing Introduction Events. Vector-Borne Zoonotic Dis. 2020, 20, 43–50. [Google Scholar] [CrossRef]
- Hönig, V.; Palus, M.; Kaspar, T.; Zemanova, M.; Majerova, K.; Hofmannova, L.; Papezik, P.; Sikutova, S.; Rettich, F.; Hubalek, Z.; et al. Multiple Lineages of Usutu Virus (Flaviviridae, Flavivirus) in Blackbirds (Turdus merula) and Mosquitoes (Culex pipiens, Cx. modestus) in the Czech Republic (2016–2019). Microorganisms 2019, 7, 568. [Google Scholar] [CrossRef] [Green Version]
- Aberle, S.W.; Kolodziejek, J.; Jungbauer, C.; Stiasny, K.; Aberle, J.H.; Zoufaly, A.; Hourfar, M.K.; Weidner, L.; Nowotny, N. Increase in human West Nile and Usutu virus infections, Austria, 2018. Eurosurveill 2018, 23, 1800545. [Google Scholar] [CrossRef] [Green Version]
- Cadar, D.; Lühken, R.; van der Jeugd, H.; Garigliany, M.; Ziegler, U.; Keller, M.; Lahoreau, J.; Lachmann, L.; Becker, N.; Kik, M.; et al. Widespread activity of multiple lineages of Usutu virus, western Europe, 2016. Eurosurveill 2017, 22, 30452. [Google Scholar] [CrossRef] [PubMed]
- Cavrini, F.; Gaibani, P.; Longo, G.; Pierro, A.M.; Rossini, G.; Bonilauri, P.; Gerunda, G.E.; Di Benedetto, F.; Pasetto, A.; Girardis, M.; et al. Usutu virus infection in a patient who underwent orthotropic liver transplantation, Italy, August–September 2009. Eurosurveill 2009, 14, 19448. [Google Scholar] [CrossRef]
- Cook, C.L.; Huang, Y.-J.S.; Lyons, A.C.; Alto, B.W.; Unlu, I.; Higgs, S.; VanLandingham, D.L. North American Culex pipiens and Culex quinquefasciatus are competent vectors for Usutu virus. PLoS Negl. Trop. Dis. 2018, 12, e0006732. [Google Scholar] [CrossRef] [Green Version]
- Grottola, A.; Marcacci, M.; Tagliazucchi, S.; Gennari, W.; Di Gennaro, A.; Orsini, M.; Monaco, F.; Marchegiano, P.; Marini, V.; Meacci, M.; et al. Usutu virus infections in humans: A retrospective analysis in the municipality of Modena, Italy. Clin. Microbiol. Infect. 2017, 23, 33–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santini, M.; Vilibic-Cavlek, T.; Barsic, B.; Barbic, L.; Savic, V.; Stevanovic, V.; Listes, E.; Di Gennaro, A.; Savini, G. First cases of human Usutu virus neuroinvasive infection in Croatia, August-September 2013: Clinical and laboratory features. J. Neurovirol. 2015, 21, 92–97. [Google Scholar] [CrossRef]
- Simonin, Y.; Sillam, O.; Carles, M.J.; Gutierrez, S.; Gil, P.; Constant, O.; Martin, M.F.; Girard, G.; Van de Perre, P.; Salinas, S.; et al. Human Usutu Virus Infection with Atypical Neurologic Presentation, Montpellier, France, 2016. Emerg. Infect. Dis. 2018, 24, 875–878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaintoutis, S.C.; Dovas, C.L.; Papanastassopoulou, M.; Gewehr, S.; Danis, K.; Beck, C.; Lecollinet, S.; Vasilis, A.; Kalaitzopoulou, S.; Panagiotopoulos, T.; et al. Evaluation of a West Nile virus surveillance and early warning system in Greece, based on domestic pigeons. Comp. Immunol. Microbiol. Infect. Dis. 2014, 37, 131–141. [Google Scholar] [CrossRef]
- National Public Health Organization (NPHO). Malaria/Annual Epidemiological Data. Available online: https://eody.gov.gr/en/epidemiological-statistical-data/ (accessed on 3 April 2020).
- Hellenic Statistical Authority (HSA). Population-Housing Census 2011. Available online: https://www.statistics.gr/en/2011-census-pop-hous (accessed on 15 July 2020).
- Region of Attiki–European Commission. Available online: https://ec.europa.eu/growth/tools-databases/regional-innovation-monitor/base-profile/region-attiki (accessed on 31 January 2020).
- Gaitanis, A.; Kalogeropoulos, K.; Detsis, V.; Chalkias, C. Monitoring 60 Years of Land Cover Change in the Marathon Area, Greece. Land 2015, 4, 337–354. [Google Scholar] [CrossRef] [Green Version]
- MALWEST 2013. Study on Presence, Seasonal Variation and Spatial Distribution of Mosquitoes and Design of an Integrated Mosquito Management Plan. Report Regarding Mosquito Species and Geographical Distribution. Available online: http://www.malwest.gr/en-us/deliverables.aspx (accessed on 27 July 2020).
- Stefopoulou, A.; Balatsos, G.; Petraki, A.; LaDeau, S.L.; Papachristos, D.; Michaelakis, A. Reducing Aedes albopictus breeding sites through education: A study in urban area. PLoS ONE 2018, 13, e0202451. [Google Scholar] [CrossRef] [Green Version]
- Biogents, A.G. The BG-Sentinel: Biogent’s Mosquito Trap for Researchers. Available online: http://www.bg-sentinel.com/ (accessed on 29 September 2020).
- Becker, N.; Petrić, D.; Zgomba, M.; Boase, C.; Madon, M.; Dahl, C.; Kaiser, A. Mosquitoes and Their Control, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2010. [Google Scholar]
- Darsie, R.F.J.; Samanidou-Voyadjoglou, A. Keys for the identification of the mosquitoes of Greece. J. Am. Mosq. Control Assoc. 1997, 13, 247–254. [Google Scholar]
- Samanidou–Voyadjoglou, A.; Harbach, R.E. Keys to the adult female mosquitoes (Culicidae) of Greece. Eur. Mosq. Bull. 2001, 10, 13–20. [Google Scholar]
- Wilkerson, R.C.; Linton, Y.-M.; Fonseca, D.; Schultz, T.R.; Price, D.C.; Strickman, D.A. Making Mosquito Taxonomy Useful: A Stable Classification of Tribe Aedini that Balances Utility with Current Knowledge of Evolutionary Relationships. PLoS ONE 2015, 10, e0133602. [Google Scholar] [CrossRef]
- Beleri, S.; Chatzinikolaou, S.; Nearchou, A.; Patsoula, E. Entomological study of the mosquito fauna in the regional unit of Drama, region of East Macedonia-Thrace, Greece (2015 to 2016). Vector Borne Zoonotic Dis 2017, 17, 665–671. [Google Scholar] [CrossRef] [PubMed]
- Patsoula, E.; Samanidou-Voyadjoglou, A.; Spanakos, G.; Kremastinou, J.; Nasioulas, G.; Vakalis, N.C. Molecular characterization of the Anopheles maculipennis complex during surveillance for the 2004 Olympic Games in Athens. Med. Vet. Entomol. 2007, 21, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Hapip, A.C.; Liu, B.; Fang, C.T. Highly sensitive TaqManRT-PCR assay for detection and quantification of both lineages of West Nile virus RNA. J. Clin. Virol. 2006, 36, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Cavrini, F.; Della Pepa, M.E.; Gaibani, P.; Pierro, A.M.; Rossini, G.; Landini, M.P.; Sambri, V. A rapid and specific real-time RT-PCR assay to identify Usutu virus in human plasma, serum, and cerebrospinal fluid. J. Clin. Virol. Off. Publ. Pan Am. Soc. Clin. Virol. 2011, 50, 221–223. [Google Scholar] [CrossRef]
- Weissenböck, H.; Bakonyi, Τ.; Chvala, S.; Nowotny, N. Experimental Usutu virus infection of suckling mice causes neuronal and glial cell apoptosis and demyelination. Acta Neuropathol. 2004, 108, 453–460. [Google Scholar] [CrossRef]
- Biggerstaff Brad, J. PooledInfRate, Version 4.0: A Microsoft® Office Excel© Add-In to Compute Prevalence Estimates from Pooled Samples; Centers for Disease Control and Prevention: Fort Collins, CO, USA, 2009.
- Sinka, M.E.; Banga, M.J.; Manguin, S.; Coetzee, M.; Mbogo, C.M.; Hemingway, J.; Patil, A.P.; Temperley, W.H.; Getheing, P.W.; Kabari, C.W.; et al. The dominant Anopheles vectors of human malaria in Africa, Europe and middle East: Occurrence data, distribution maps and bionomic précis. Parasites Vectors 2010, 3, 117. [Google Scholar] [CrossRef] [Green Version]
- Kavran, M.; Zgomba, M.; Weitzel, T.; Petric, D.; Manz, C.; Becker, N. Distribution of Anopheles daciae and other Anopheles maculipennis complex species in Serbia. Parasitol. Res. 2018, 117, 3277–3287. [Google Scholar] [CrossRef] [Green Version]
- Naddaf, S.R.; Oshaghi, M.A.; Vatandoost, H. Confirmation of Two Sibling Species among Anopheles fluviatilis Mosquitoes in South and Southeastern Iran by Analysis of Cytochrome Oxidase I Gene. J. Arthropod Borne Dis. 2012, 6, 144–150. [Google Scholar]
- Linton, Y.M.; Smith, L.; Koliopoulos, G.; Zounos, A.K.; Samanidou-Voyadjoglou, A.; Patsoula, E.; Harbach, R. The Anopheles (Anopheles) maculipennis complex (Diptera: Culicidae) in Greece. J. Nat. Hist. 2007, 41, 2683–2699. [Google Scholar] [CrossRef]
- Halekoh, U.; Højsgaard, S.; Yan, J. The R package geepack for generalized estimating equations. J. Stat. Softw. 2006, 15, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Yan, J. Geepack: Yet another package for generalized estimating equations. R-News 2002, 2, 12–14. [Google Scholar]
- Barzon, L.; Papa, A.; Lavezzo, E.; Franchin, E.; Pacenti, M.; Sinigaglia, A.; Masi, G.; Trevisan, M.; Squarzon, L.; Toppo, S.; et al. Phylogenetic characterization of Central/Southern European lineage 2 West Nile virus: Analysis of human outbreaks in Italy and Greece, 2013–2014. Clin. Microbiol. Infect. 2015, 21, 1122.e1–1122.e10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engler, O.; Savini, G.; Papa, A.; Figuerola, J.; Groschup, M.H.; Kampen, H.; Medlock, J.; Vaux, A.; Wilson, A.J.; Werner, D.; et al. European Surveillance for West Nile Virus in Mosquito Populations. Int. J. Environ. Res. Public Health 2013, 10, 4869–4895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilibic-Cavlek, T.; Savic, V.; Petrovic, T.; Toplak, I.; Barbic, L.; Petric, D.; Tabain, I.; Hrnjakovic-Cvjetkovic, I.; Bogdanic, M.; Klobucar, A.; et al. Emerging Trends in the Epidemiology of West Nile and Usutu Virus Infections in Southern Europe. Front. Vet.-Sci. 2019, 6, 437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilibic-Cavlek, T.; Savic, V.; Sabadi, D.; Peric, L.; Barbic, L.; Klobucar, A.; Miklausic, B.; Tabain, I.; Santini, M.; Vucelja, M.; et al. Prevalence and molecular epidemiology of West Nile and Usutu virus infections in Croatia in the “One health” context, 2018. Transbound. Emerg. Dis. 2019, 66, 1946–1957. [Google Scholar] [CrossRef] [PubMed]
- Calzolari, M.; Chiapponi, C.; Bonilauri, P.; Lelli, D.; Baioni, L.; Barbieri, I.; Lavazza, A.; Pongolini, S.; Dottori, M.; Moreno, A. Co-circulation of two Usutu virus strains in Northern Italy between 2009 and 2014. Infect. Genet. Evol. 2017, 51, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Engel, D.; Jöst, H.; Wink, M.; Börstler, J.; Bosch, S.; Garigliany, M.-M.; Jöst, A.; Czajka, C.; Lühken, R.; Ziegler, U.; et al. Reconstruction of the Evolutionary History and Dispersal of Usutu Virus, a Neglected Emerging Arbovirus in Europe and Africa. mBio 2016, 7, e01938-15. [Google Scholar] [CrossRef] [Green Version]
- Blagrove, M.S.C.; Sherlock, K.; Chapman, G.E.; Impoinvil, D.E.; McCall, P.J.; Medlock, J.M.; Lycett, G.; Solomon, T.; Baylis, M. Evaluation of the vector competence of a native UK mosquito Ochlerotatus detritus (Aedes detritus) for dengue, chikungunya and West Nile viruses. Parasites Vectors 2016, 9, 452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camp, J.V.; Kolodziejek, J.; Nowotny, N. Targeted surveillance reveals native and invasive mosquito species infected with Usutu virus. Parasites Vectors 2019, 12, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mancini, G.; Montarsi, F.; Calzolari, M.; Capelli, G.; Dottori, M.; Ravagnan, S.; Lelli, D.; Chiari, M.; Santilli, A.; Quaglia, M.; et al. Mosquito species involved in the circulation of West Nile and Usutu viruses in Italy. Vet. Ital. 2017, 53, 97–110. [Google Scholar] [PubMed]
- Puggioli, A.; Bonilauri, P.; Calzolari, M.; Lelli, D.; Carrieri, M.; Urbanelli, S.; Pudar, D.; Bellini, R. Does Aedes albopictus (Diptera: Culicidae) play any role in Usutu virus transmission in Northern Italy? Experimental oral infection and field evidences. Acta Trop. 2017, 172, 192–196. [Google Scholar] [CrossRef]
- Gomes, B.; Kioulos, E.; Papa, A.; Almeida, A.P.; Vontas, J.; Pinto, J. Distribution and hybridization of Culex pipiens forms in Greece during the West Nile virus outbreak of 2010. Infect. Genet. Evol. 2013, 16, 218–225. [Google Scholar] [CrossRef]
- Vakalis, N.; Patsoula, E.; Samanidou-Voyadjoglou, A. Mosquito surveillance and control (Chapter 11). In Mass Gathering and Public Health. The Experience of the Athens 2004 Olympic Games; Tsouros, A.D., Efstathiou, P.A., Eds.; WHO Europe: Geneva, Switzerland, 2007; pp. 193–208. [Google Scholar]
- Ministry of Rural Development and Food. Available online: http://www.minagric.gr/index.php/el/for-citizen-2/nosimata-zoon/602-progrepitdneilou17 (accessed on 30 July 2021).
- European Centre for Disease Prevention and Control (ECDC). West Nile Virus Infection, Annual Epidemiological Report for 2018. Available online: https://www.ecdc.europa.eu/en/publications-data/west-nile-virus-infection-annual-epidemiological-report-2018 (accessed on 25 May 2021).
- Mavrouli, M.; Vrioni, G.; Kapsimali, V.; Tsiamis, C.; Mavroulis, S.; Pervanidou, D.; Billinis, C.; Hadjichristodoulou, C.; Tsakris, A. Reemergence of West Nile virus Infections in Southern Greece, 2017. Am. J. Trop. Med. Hyg. 2019, 100, 420–426. [Google Scholar] [CrossRef] [Green Version]
- Valiakos, G.; Touloudi, A.; Iacovakis, C.; Athanasiou, A.; Birtsas, P.; Spyrou, V.; Billinis, C. Molecular detection and phylogenetic analysis of West Nile virus lineage 2 in sedentary wild birds (Eurasian magpie), Greece, 2010. Eurosurveill 2011, 16, 19862. [Google Scholar] [CrossRef]
- Llorente, F.; García-Irazábal, A.; Pérez-Ramírez, E.; Cano-Gómez, C.; Sarasa, M.; Vázquez, A.; Jiménez-Clavero, M.A. Influence of flavivirus co-circulation in serological diagnostics and surveillance: A model of study using West Nile, Usutu and Bagaza viruses. Transbound. Emerg. Dis. 2019, 66, 2100–2106. [Google Scholar] [CrossRef]
- Calzolari, M.; Gaibani, P.; Bellini, R.; Defilippo, F.; Pierro, A.; Albieri, A.; Maioli, G.; Luppi, A.; Rossini, G.; Balzani, A.; et al. Mosquito, bird and human surveillance of West Nile and Usutu viruses in Emilia-Romagna Region (Italy) in 2010. PLoS ONE 2012, 7, e38058. [Google Scholar] [CrossRef] [Green Version]
- Collins, F.H.; Kamau, L.; Ranson, H.; Vulule, J.M. Molecular entomology and prospects for malaria control. Bull. World Health Organ. 2003, 78, 1412–1423. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Attica Region | Surveyed RU Areas (Abbreviation) and Period | BG-Sentinel Traps (BGs) CO2 + BG-Lure | Sampling Location | GPS Coordinates (Decimal Degrees) | Microenvironment Description of Sampling Site | No. of Weeks | No. of Collections (Field/Problematic Collections) | Sampling Frequency | |
|---|---|---|---|---|---|---|---|---|---|
| Latitude | Longitude | ||||||||
| East Attica (EA) | Wetland Area (WT) Marathonas-Schinias (MS) 15/06/2017–28/12/2018 | MS1 | Schinias | 38.147729 | 24.002428 | Private house/outdoor garden with a large number of trees and large green spaces, agricultural area (semi-urban area) | 80 | 74 (80/6) | Weekly |
| MS2 | Schinias | 38.131439 | 23.995250 | Private house/outdoor garden with a large number of trees and large green spaces, agricultural area (semi-urban area) | 80 | 69 (80/11) | Weekly | ||
| MS3 | Schinias | 38.143819 | 24.030472 | Private house/outdoor garden with a large number of trees and large green spaces, agricultural area (bordered by the marsh) | 59 | 59 (59/0) | Weekly | ||
| MS4 | Kato Souli | 38.166265 | 24.022584 | Private house/outdoor garden with a large number of trees and large green spaces, agricultural area (rural area) | 76 | 75 (76/1) | Weekly | ||
| Total MS | 295 | 277 (295/18) | |||||||
| South Sector (SS) | Urban area (UR) Palaio Faliro (PF) 21/06/2017–27/12/2018 | PF1 | Open Protection Centers for Elderly | 37.931997 | 23.692625 | Municipality building/outdoor garden with a large number of trees and large green spaces, urban area | 79 | 77 (79/2) | Weekly |
| PF2 | City Hall | 37.928111 | 23.699008 | Municipality building/outdoor garden with a large number of trees and large green spaces, urban area | 79 | 77 (79/2) | Weekly | ||
| PF3 | Rema Pikrodafnis | 37.923989 | 23.710129 | Private house/outdoor garden with a large number of trees and large green spaces, urban area | 78 | 64 (78/14) | Weekly | ||
| Total PF | 236 | 218 (236/18) | |||||||
| B (95% CI) | Wald χ2 | p | |
|---|---|---|---|
| Culex | |||
| Intercept | 117.608 (38.523–196.693) | 8.50 | 0.004 |
| Area: Faliro (ref: Marathonas) | −90.582 (−171.388, −776) | 4.83 | 0.028 |
| Aedes | |||
| Intercept | 6.560 (1.965, 11.154) | 7830 | 0.005 |
| Area: Faliro (ref: Marathonas) | 3.038 (−4.685, 10.762) | 0.594 | 0.441 |
| Mosquito Species | Total Number/Species (%) | F ♀ | M ♂ | Marathonas-Schinias (MS) Wetland Area (WT) | Palaio Faliro (PF) Urban Area (UR) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MS1 | MS2 | MS3 | MS4 | Total Number/Species (%) | PF1 | PF2 | PF3 | Total Number/Species (%) | ||||
| Aedes (Stegomyia) albopictus (Skuse) # | 3762 (8.05) | 3041 | 721 | 828 | 685 | 87 | 155 | 1755 (3.75) | 991 | 160 | 856 | 2007 (4.3) |
| Aedes (Ochlerotatus) caspius (Pallas) | 81 (0.173) | 79 | 2 | 21 | 21 | 6 | 27 | 75 (0.16) | 0 | 0 | 6 | 6 (0.013) |
| Aedes (Ochlerotatus) detritus (Haliday) | 588 (1.25) | 580 | 8 | 187 | 105 | 247 | 49 | 588 (1.25) | 0 | 0 | 0 | 0 |
| Anopheles (Anopheles) algeriensis (Theobald) | 209 (0.45) | 209 | 0 | 22 | 2 | 67 | 118 | 209 (0.45) | 0 | 0 | 0 | 0 |
| Anopheles (Anopheles) claviger (Meigen) | 160 (0.34) | 160 | 0 | 39 | 2 | 34 | 85 | 160 (0.34) | 0 | 0 | 0 | 0 |
| Anopheles (Anopheles) maculipennis s.l. (Meigen) | 2 (0.0042) | 2 | 0 | 1 | 0 | 1 | 0 | 2 (0.0042) | 0 | 0 | 0 | 0 |
| Anopheles (Anopheles) sacharovi (Favre) | 93 (0.2) | 93 | 0 | 43 | 0 | 33 | 17 | 93 (0.2) | 0 | 0 | 0 | 0 |
| Coquillettidia (Coquillettidia) richiardii (Ficalbi) | 164 (0.35) | 164 | 0 | 55 | 0 | 41 | 68 | 164 (0.35) | 0 | 0 | 0 | 0 |
| Culex (Culex) pipiens (Linnaeus) | 41,236 (88.25) | 41,050 | 186 | 11,480 | 4989 | 3416 | 14,643 | 34,528 (73.90) | 3150 | 985 | 2573 | 6708 (14.35) |
| Culex (Culex) theileri (Theobald) | 16 (0.034) | 16 | 0 | 0 | 0 | 0 | 16 | 16 (0.034) | 0 | 0 | 0 | 0 |
| Culiseta (Culiseta) annulata (Schrank) | 11 (0.023) | 11 | 0 | 6 | 0 | 3 | 2 | 11 (0.023) | 0 | 0 | 0 | 0 |
| Culiseta (Culicella) fumipennis (Stephens) | 5 (0.010) | 0 | 5 | 0 | 0 | 5 | 0 | 5 (0.010) | 0 | 0 | 0 | 0 |
| Culiseta (Allotheobaldia) longiareolata (Macquart) | 395 (0.84) | 254 | 141 | 34 | 21 | 18 | 127 | 200 (0.43) | 81 | 97 | 17 | 195 (0.41) |
| Culiseta (Culiseta) subochrea (Edwards) | 3 (0.0064) | 3 | 0 | 2 | 0 | 0 | 1 | 3 (0.0064) | 0 | 0 | 0 | 0 |
| Uranotaenia (Pseudoficalbia) unguiculata (Edwards) | 1 (0.002) | 1 | 0 | 0 | 0 | 0 | 1 | 1 (0.002) | 0 | 0 | 0 | 0 |
| Total | 46,726 | 55,307 | 1347 | 12,718 | 5825 | 3958 | 15,309 | 37,810 | 4222 | 1242 | 3452 | 8916 |
| Surveyed RUs | West Nile Virus | Usutu Virus | |||||
|---|---|---|---|---|---|---|---|
| 2017 | 2018 | Total No. of Positive/Tested Pools | MLE | 2017 | 2018 | Total No. of Positive/Tested Pools | |
| Np/Nt * | Np/Nt | Np/Nt | Np/Nt | ||||
| East Attica/Marathonas-Schinias (wetland area) | 18/120 | 12/147 | 30/267 | 0.00010 (95% CL 0.0007–0.0014) | 0/120 | 0/147 | 0/267 |
| South Sector/Palaio Faliro (urban area) | 5/33 | 3/66 | 8/99 | 0.01 (95% CL 0.0006–0.0026) | 0/33 | 0/66 | 0/99 |
| Total no. of positive/tested pools per transmission year | 23/153 | 15/213 | 38/366 | 0/153 | 0/213 | 0/366 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beleri, S.; Balatsos, G.; Karras, V.; Tegos, N.; Sereti, F.; Rachiotis, G.; Hadjichristodoulou, C.; Papadopoulos, N.; Papachristos, D.; Michaelakis, A.; et al. Seasonal Phenological Patterns and Flavivirus Vectorial Capacity of Medically Important Mosquito Species in a Wetland and an Urban Area of Attica, Greece. Trop. Med. Infect. Dis. 2021, 6, 176. https://doi.org/10.3390/tropicalmed6040176
Beleri S, Balatsos G, Karras V, Tegos N, Sereti F, Rachiotis G, Hadjichristodoulou C, Papadopoulos N, Papachristos D, Michaelakis A, et al. Seasonal Phenological Patterns and Flavivirus Vectorial Capacity of Medically Important Mosquito Species in a Wetland and an Urban Area of Attica, Greece. Tropical Medicine and Infectious Disease. 2021; 6(4):176. https://doi.org/10.3390/tropicalmed6040176
Chicago/Turabian StyleBeleri, Stavroula, Georgios Balatsos, Vasilios Karras, Nikolaos Tegos, Fani Sereti, Georgios Rachiotis, Christos Hadjichristodoulou, Nikolaos Papadopoulos, Dimitrios Papachristos, Antonios Michaelakis, and et al. 2021. "Seasonal Phenological Patterns and Flavivirus Vectorial Capacity of Medically Important Mosquito Species in a Wetland and an Urban Area of Attica, Greece" Tropical Medicine and Infectious Disease 6, no. 4: 176. https://doi.org/10.3390/tropicalmed6040176
APA StyleBeleri, S., Balatsos, G., Karras, V., Tegos, N., Sereti, F., Rachiotis, G., Hadjichristodoulou, C., Papadopoulos, N., Papachristos, D., Michaelakis, A., & Patsoula, E. (2021). Seasonal Phenological Patterns and Flavivirus Vectorial Capacity of Medically Important Mosquito Species in a Wetland and an Urban Area of Attica, Greece. Tropical Medicine and Infectious Disease, 6(4), 176. https://doi.org/10.3390/tropicalmed6040176