
A Case Report on a Human Bite Contact with a Rabid Honey Badger Mellivora capensis (Kromdraai Area, Cradle of Humankind, South Africa)

 , ,
, , 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Specimens and Rabies Testing

| Virus # | Laboratory Number | Species | Geographic Location | Accession Number | Reference |
|---|---|---|---|---|---|
| 1 | 137/21 | Black-backed jackal | South Africa, Gauteng Province | OP939356 | This study |
| 2 | 144/21 | Black-backed jackal | South Africa, Gauteng Province | OP939357 | This study |
| 3 | 159/21 | Black-backed jackal | South Africa, Gauteng Province | OP939358 | This study |
| 4 | 169/21 | Black-backed jackal | South Africa, Gauteng Province | OP939359 | This study |
| 5 | 170/21 | Black-backed jackal | South Africa, Gauteng Province | OP939360 | This study |
| 6 | 192/21 | Honey badger | South Africa, Gauteng Province | OP939353 | This study |
| 7 | 217/21 | Domestic dog | South Africa, Gauteng Province | OP939355 | This study |
| 8 | 242/21 | Black-backed jackal | South Africa, Gauteng Province | OP939354 | This study |
| 9 | 115/11 | Domestic dog | South Africa, Gauteng Province | JN227482 | [12] |
| 10 | 157/11 | Domestic dog | South Africa, Gauteng Province | JN227483 | [12] |
| 11 | 288/11 | Domestic dog | South Africa, Gauteng Province | JN227487 | [12] |
| 12 | 343/11 | Domestic dog | South Africa, Gauteng Province | JQ756150 | [12] |
| 13 | 464/10 | Domestic dog | South Africa, Gauteng Province | JF327493 | [12] |
| 14 | 503/10 | Domestic dog | South Africa, Gauteng Province | JF327494 | [12] |
| 15 | 409/16 | Black-backed jackal | South Africa, Gauteng Province | MW413411 | [14] |
| 16 | 428/16 | Black-backed jackal | South Africa, Gauteng Province | MW413414 | [14] |
| 17 | 489/16 | Bovine | South Africa, Gauteng Province | MW413403 | [14] |
| 18 | 527/16 | Bovine | South Africa, Gauteng Province | MW413405 | [14] |
| 19 | 769/16 | Black-backed jackal | South Africa, Gauteng Province | MK103257 | [15] |
| 20 | 784/16 | Black-backed jackal | South Africa, Gauteng Province | MK103273 | [15] |
| 21 | 454/17 | Black-backed jackal | South Africa, North-West Province | MT454646 | [16] |
| 22 | 460/17 | Black-backed jackal | South Africa, North-West Province | MT454647 | [16] |
| 23 | 466/17 | Black-backed jackal | South Africa, North-West Province | MT454648 | [16] |
| 24 | 474/17 | Black-backed jackal | South Africa, North-West Province | MT454649 | [16] |
| 25 | 480/17 | Black-backed jackal | South Africa, North-West Province | MT454651 | [16] |
| 26 | 14/377 | Domestic dog | South Africa, Eastern Cape Province | MF197279 | [17] |
| 27 | 14/392 | Domestic dog | South Africa, Eastern Cape Province | MF197280 | [17] |
| 28 | 14/430 | Domestic dog | South Africa, Eastern Cape Province | MF197227 | [17] |
| 29 | 15/474 | Domestic dog | South Africa, Eastern Cape Province | MF197283 | [17] |
| 30 | 16/318 | Domestic dog | South Africa, Eastern Cape Province | MF197284 | [17] |
| 31 | 16/388 | Domestic dog | South Africa, Eastern Cape Province | MF197285 | [17] |
| 32 | 10/299 | Domestic dog | South Africa, KwaZulu Natal Province | KC660351 | [18] |
| 33 | 10/479 | Domestic dog | South Africa, KwaZulu Natal Province | KC660348 | [18] |
| 34 | 88/08 | Domestic dog | South Africa, Limpopo Province | GU808517 | [19] |
| 35 | 169/09 | Domestic dog | South Africa, Limpopo Province | GU808515 | [19] |
| 36 | 460/09 | Domestic dog | South Africa, Limpopo Province | GU808514 | [19] |
| 37 | 572/09 | Domestic dog | South Africa, Limpopo Province | GU808513 | [19] |
| 38 | 1305/09 | Domestic dog | South Africa, Limpopo Province | GU808512 | [19] |
| 39 | 201/12 | Bovine | Lesotho, Maseru | MF197287 | [17] |
| 40 | 60/14 | Bovine | Lesotho, Maseru | MF197297 | [17] |
| 41 | 195/14 | Bovine | Lesotho, Berea | MF197299 | [17] |
| 42 | 17/15 | Domestic dog | Lesotho, Maseru | MF197305 | [17] |
| 43 | 21/15 | Bovine | Lesotho, Maseru | MF197303 | [17] |
| 44 | 24/15 | Domestic dog | Lesotho, Maseru | MF197304 | [17] |
| 45 | 572/99 | Domestic dog | Mozambique, Gaza Province | KM262039 | [17] |
| 46 | 558/05 | Domestic dog | Mozambique, Inhambane Province | KM262043 | [17] |
| 47 | 482/12 | Domestic dog | Mozambique, Maputo Province | KM262047 | [17] |
| 48 | 233/13 | Domestic dog | Mozambique, Maputo Province | KM262049 | [20] |
| 49 | 804/06 | Domestic dog | Mozambique, Manica | EU123934 | [20] |
| 50 | 187/14 | Domestic dog | Zimbabwe, Harare | MF425794 | [20] |
| 51 | 200/14 | Domestic dog | Zimbabwe, Harare | MF425796 | [20] |
| 52 | 355/14 | Domestic dog | Zimbabwe, Harare | MF425800 | [20] |
| 53 | 239/15 | Domestic dog | Zimbabwe, Harare | MF425805 | [19] |
| 54 | 406/15 | Domestic dog | Zimbabwe, Harare | MF425808 | [19] |
| 55 | 464/15 | Domestic dog | Zimbabwe, Harare | MF425811 | [19] |

{kind=link}
{kind=link}
{kind=link}
2.2. Antigenic Typing
2.3. Total RNA Extractions, Reverse Transcription PCR (RT-PCR), Nucleotide Sequencing and Phylogenetic Reconstruction
3. Results
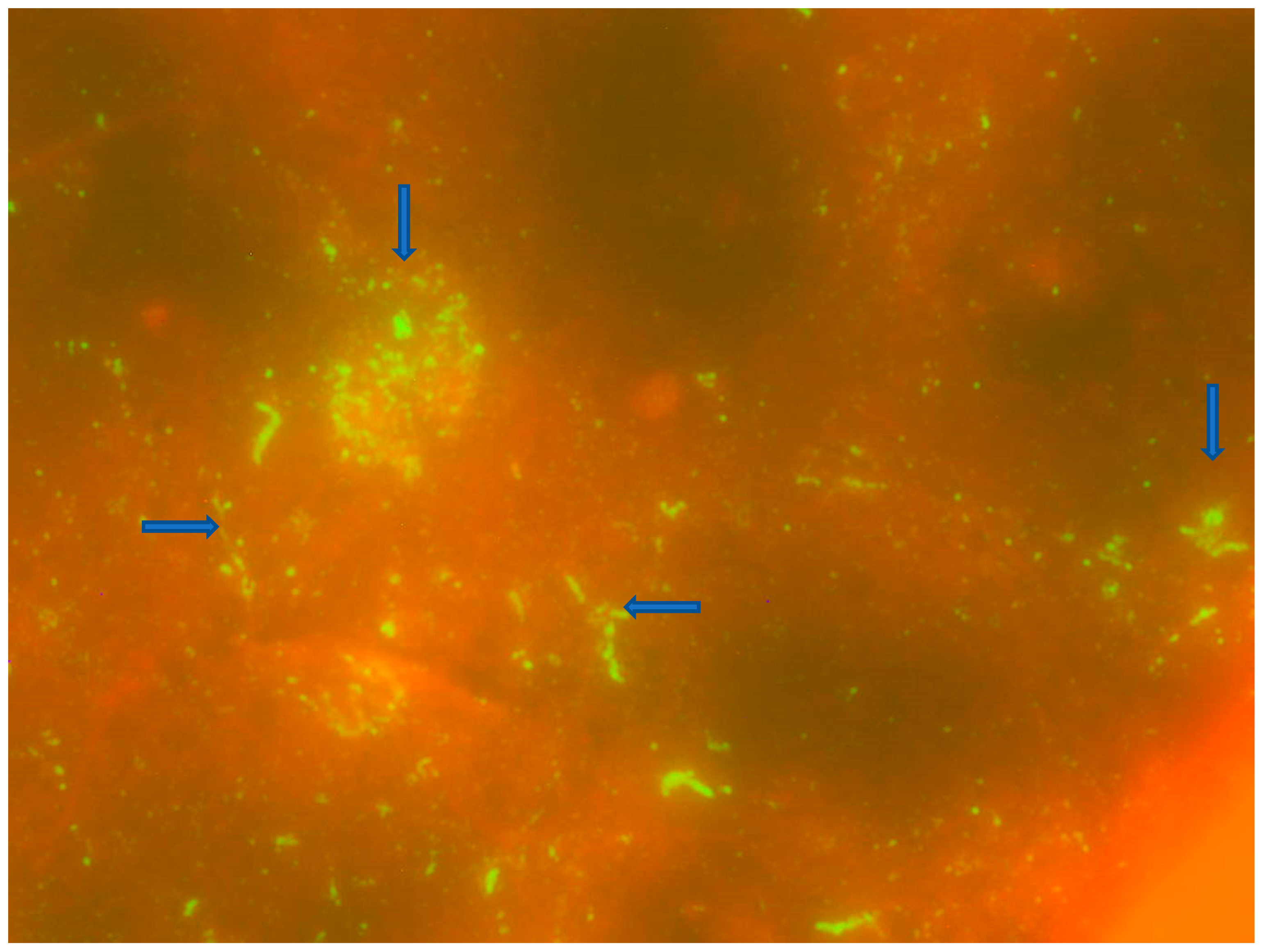
3.1. Direct Fluorescent Antibody Testing and Monoclonal Antibody Typing
3.2. RT-PCR, Sequencing and Phylogenetic Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hampson, K.; Coudeville, L.; Lembo, T.; Sambo, M.; Kieffer, A.; Attlan, M.; Barrat, J.; Blanton, J.D.; Briggs, D.J.; Cleaveland, S.; et al. Estimating the global burden of endemic canine rabies. PLoS Negl. Trop. Dis. 2015, 9, e0003709. [Google Scholar]
- WHO. Human rabies transmitted by dogs: Current status of global data 2015. Wkly. Epidemiol. Rec. 2016, 2, 13–22. [Google Scholar]
- International Committee on the Taxonomy of Viruses. The ICTV Online (10th) Report on Virus Taxonomy. 2017. Available online: https://talk.ictvonline.org/taxonomy (accessed on 7 December 2020).
- Badrane, H.; Bahloul, C.; Perrin, P.; Tordo, N. Evidence of Two Lyssavirus Phylogroups with Distinct Pathogenicity and Immunogenicity. J. Virol. 2001, 75, 3268–3276. [Google Scholar] [CrossRef] [Green Version]
- Kuzmin, I.V. Chapter One-Basic Facts about Lyssaviruses. In Current Laboratory Techniques in Rabies Diagnosis, Research and Prevention; Rupprecht, C., Nagarajan, T., Eds.; Academic Press: Amsterdam, The Netherlands, 2014; Volume 1, pp. 3–21. ISBN 978-0-12-800014-4. [Google Scholar]
- Coertse, J.; Grobler, C.S.; Sabeta, C.T.; Seamark, E.C.J.; Kearney, T.; Paweska, J.T.; Markotter, W. Lyssaviruses in Insectivorous Bats, South Africa, 2003–2018. Emerg. Infect. Dis. 2020, 26, 3056–3060. [Google Scholar] [CrossRef]
- Von Teichman, B.F.; Thomson, G.R.; Meredith, C.D.; Nel, L.H. Molecular epidemiology of rabies virus in South Africa: Evidence for two distinct virus groups. J. Gen. Virol. 1995, 76, 73–82. [Google Scholar] [CrossRef]
- Sabeta, C.T.; Mansfield, K.L.; McElhinney, L.M.; Fooks, A.R.; Nel, L.H. Molecular epidemiology of rabies in bat-eared foxes (Otocyon megalotis) in South Africa. Virus Res. 2007, 129, 1–10. [Google Scholar] [CrossRef]
- Zulu, G.C.; Sabeta, C.T.; Nel, L.H. Molecular epidemiology of rabies: Focus on domestic dogs (Canis familiaris) and black-backed jackals (Canis mesomelas) from northern South Africa. Virus Res. 2009, 140, 71–78. [Google Scholar] [CrossRef]
- Bingham, J. Canine rabies ecology in southern Africa. Emerg. Infect. Dis. 2005, 11, 1337–1342. [Google Scholar] [CrossRef]
- Weyer, J.; Dermaux-Msimang, V.; Grobbelaar, A.; Le Roux, C.; Moolla, N.; Paweska, J.; Blumberg, L. Epidemiology of human rabies in South Africa, 2008–2018. S. Afr. Med. J. 2020, 110, 877–881. [Google Scholar] [CrossRef]
- Sabeta, C.T.; Weyer, J.; Geertsma, P.; Mohale, D.; Miyen, J.; Blumberg, L.H.; Leman, P.A.; Phahladira, B.; Shumba, W.; Walters, J.; et al. Emergence of rabies in the Gauteng Province, South Africa: 2010–2011. J. S. Afr. Vet. Assoc. 2013, 84, E1–E5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manual of Diagnostic Tests and Vaccines for Terrestrial Animals. Rabies (Infection with Rabies virus and other lyssaviruses). Chapter 3.1.18. Available online: https://www.woah.org/en/what-we-do/standards/codes-and-manuals/terrestrial-manual-online-access/ (accessed on 23 February 2023).
- Ngoepe, E.; Chirima, J.G.; Mohale, D.; Mogano, K.; Suzuki, T.; Makita, K.; Sabeta, C.T. Rabies outbreak in black-backed jackals (Canis mesomelas), South Africa, 2016. Epidemiol. Infect. 2022, 150, e137. [Google Scholar] [CrossRef]
- Schepers, C. Molecular epidemiology of rabies in domestic and wildlife in South Africa. Unpublished.
- Sabeta, C.; Marston, D.; McElhinney, L.; Horton, D.; Phahladira, B.; Fooks, A.R. Rabies in the African civet: An incidental host for lyssaviruses? Viruses 2020, 12, 368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coetzer, A.; Coertse, J.; Makalo, M.J.; Molomo, M.; Markotter, W.; Nel, L.H. Epidemiology of Rabies in Lesotho: The Importance of Routine Surveillance and Virus Characterization. Trop. Med. Infect. Dis. 2017, 2, 30. [Google Scholar] [CrossRef] [Green Version]
- Mollentze, N.; Nel, L.H.; Townsend, S.; Roux, K.; Hampson, K.; Haydon, D.T.; Soubeyrand, S.A. A Bayesian approach for inferring the dynamics of partially observed endemic infectious diseases from space-time-genetic data. Proc. Biol. Sci. 2014, 281, 20133251. [Google Scholar] [CrossRef] [Green Version]
- Sabeta, C.T.; Mkhize, G.C.; Ngoepe, E.C. An evaluation of rabies control in northern Limpopo province, South Africa. Epidemiol. Infect. 2011, 139, 1470–1475. [Google Scholar] [CrossRef]
- Coetzer, A.; Gwenhure, L.; Makaya, P.; Markotter, W.; Nel, L. Epidemiological aspects of the persistent transmission of rabies during an outbreak (2010–2017) in Harare, Zimbabwe. PLoS ONE 2019, 14, e0210018. [Google Scholar] [CrossRef]
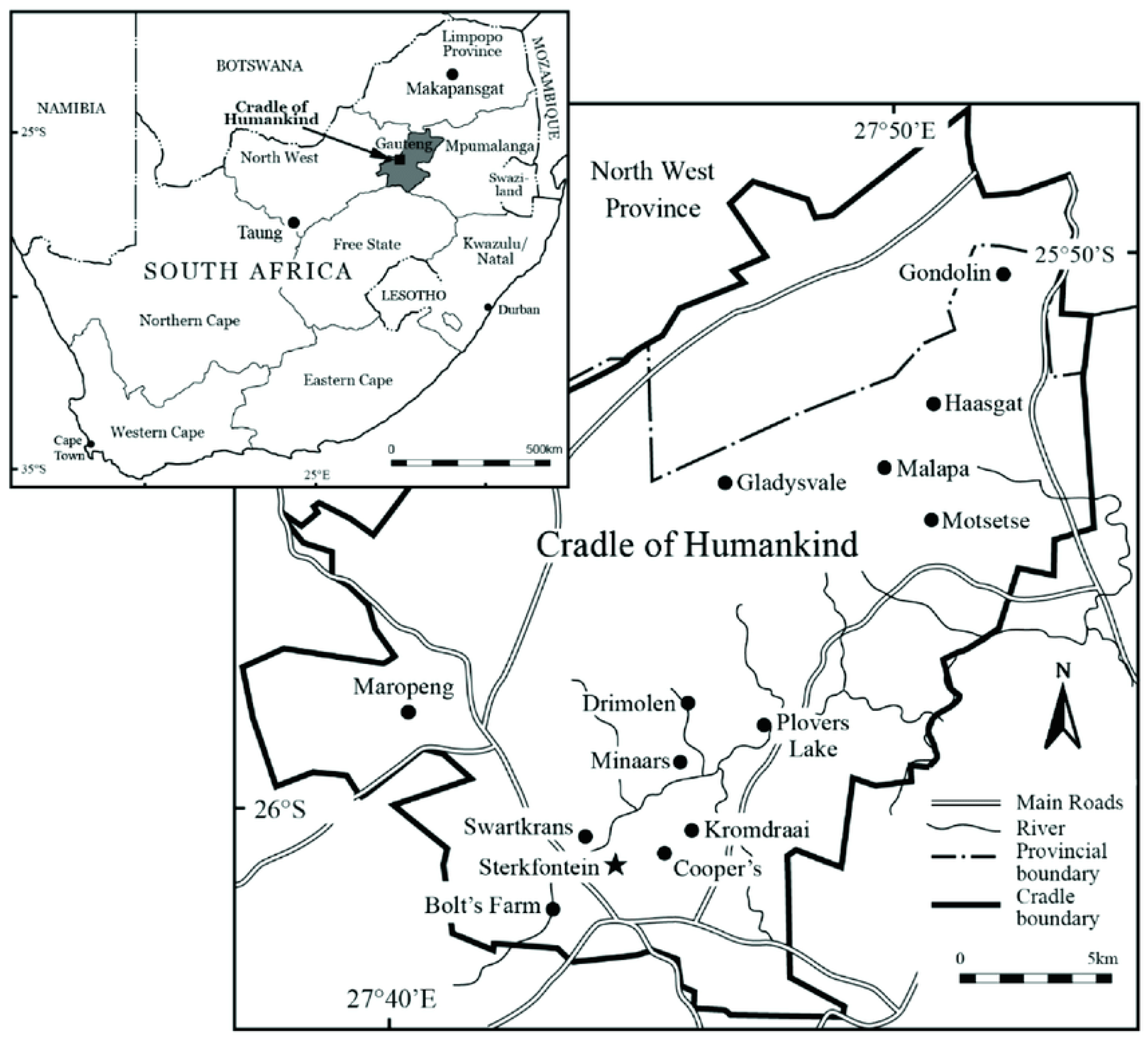
- Caruana, M.; Stratford, D.J. Historical perspectives on the significance of Archaeology in the Cradle of Humakind, South Africa. S. Afr. Archaeol. Soc. Goodwin Ser. 2019, 12, 44–55. [Google Scholar]
- Ngoepe, E.; Fehlner-Gardiner, C.; Wandeler, A.I.; Sabeta, C.T. Antigenic characterisation of lyssaviruses in South Africa. Onderstepoort J. Vet. Res. 2014, 81, 1–9. [Google Scholar] [CrossRef]
- Sacramento, D.; Bourhy, H.; Tordo, N. PCR technique as an alternative method for diagnosis and molecular epidemiology of rabies virus. Mol. Cell. Probes. 1991, 5, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Tordo, N.; Poch, O.; Ermine, A.; Keith, G.; Rougeon, F. Completion of the rabies virus genome sequence determination: Highly conserved domains among the L (polymerase) proteins of unsegmented negative-strand RNA viruses. Virology 1988, 165, 565–576. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTALW: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Tamura, K.; Nei, M. MEGA3: Integrated software for Molecular Evolutionary Genetics Analysis and sequence alignment. Brief. Bioinform. 2004, 5, 150–163. [Google Scholar] [CrossRef] [Green Version]
- Hills, D.M.; Bull, J.J. An empirical test of bootstrapping as a method for assessing confidence in phylogenetic analysis. Syst. Biol. 1993, 42, 182–192. [Google Scholar] [CrossRef]
- Nel, L.H.; Thomson, G.R.; Von Teichman, B.F. Molecular epidemiology of rabies virus in South Africa. Onderstepoort J. Vet. Res. 1993, 60, 301–306. [Google Scholar] [PubMed]
- Sabeta, C.T.; Shumba, W.; Mohale, D.; Miyen, J.M.; Wandeler, A.I.; Nel, L.H. Mongoose rabies and the African civet in Zimbabwe. Vet. Record. 2008, 163, 580. [Google Scholar] [CrossRef] [PubMed]
- Faye, M.; Faye, O.; Paola, N.D.; Ndione, M.H.D.; Diagne, M.M.; Diagne, C.T.; Bob, N.S.; Fall, G.; Heraud, J.M.; Sall, A.A. Rabies surveillance in Senegal 2001 to 2015 uncovers first infection of a honey-badger. Transbound Emerg. Dis. 2022, 69, e1350–e1364. [Google Scholar] [CrossRef]
- Sabeta, C.T.; Bingham, J.; Nel, L.H. Molecular epidemiology of canid rabies in Zimbabwe and South Africa. Virus Res. 2003, 91, 203–211. [Google Scholar] [CrossRef]
- Sacramento, D.; Badrane, H.; Bourhy, H.; Tordo, N. Molecular epidemiology of rabies virus in France: Comparison with vaccine strains. J. Gen. Virol. 1992, 73, 1149–1158. [Google Scholar]
- Bingham, J.; Foggin, C.M.; Wandeler, A.I.; Hill, F.W. The epidemiology of rabies in Zimbabwe. 1. Rabies in dogs (Canis familiaris). Onderstepoort J. Vet. Res. 1999, 66, 1–10. [Google Scholar]
- Randall, D.A.; Williams, S.D.; Kuzmin, I.V.; Rupprecht, C.E.; Tallents, L.A.; Tefera, Z.; Argaw, K.; Shiferaw, F.; Knobel, D.L.; Sillero-Zubiri, C.; et al. Rabies in endangered Ethiopian wolves. Emerg. Infect. Dis. 2004, 10, 2214–2217. [Google Scholar] [CrossRef]
- Marston, D.A.; Horton, D.L.; Nunez, J.; Ellis, R.J.; Orton, R.J.; Johnson, N.; Banyard, A.C.; McElhinney, L.M.; Freuling, C.M.; Fırat, M.; et al. Genetic analysis of a rabies virus host shift event reveals within-host viral dynamics in a new host. Virus Evol. 2017, 13, 3. [Google Scholar] [CrossRef] [Green Version]
- Koeppel, K.N.; Kuhn, B.F.; Thompson, P.N. Oral bait preferences for rabies vaccination in free-ranging black-backed jackal (Canis mesomelas) and non-target species in a multi-site field study in a peri-urban protected area in South Africa. Prev. Vet. Med. 2020, 175, 104867. [Google Scholar] [CrossRef] [PubMed]
- Knobel, D.L.; Cleaveland, S.; Coleman, P.G.; Fèvre, E.M.; Meltzer, M.I.; Miranda, M.E.; Shaw, A.; Zinsstag, J.; Meslin, F.X. Re-evaluating the burden of rabies in Africa and Asia. Bull. World Health Organ. 2005, 83, 360–368. [Google Scholar] [PubMed]
- Hofmeyr, M.; Hofmeyr, D.; Nel, L.H.; Bingham, J. A second outbreak of rabies in African wild dogs (Lycaon pictus) in Madikwe Game Reserve, South Africa, demonstrating the efficacy of vaccination against natural rabies challenge. Anim. Conserv. 2006, 7, 193–198. [Google Scholar] [CrossRef]


| Mab | 137/21 | 159/21 | 169/21 | 170/21 | 192/21 | 217/21 |
|---|---|---|---|---|---|---|
| 1C5 | - | - | - | - | - | - |
| 26AB7 | +++ | +++ | +++ | +++ | +++ | +++ |
| 26BE2 | +++ | +++ | +++ | +++ | +++ | +++ |
| 32GD12 | ++ | +++ | +++ | +++ | ++ | +++ |
| 38HF2 | ++ | +++ | +++ | +++ | ++ | +++ |
| M612 | - | - | - | - | - | - |
| M837 | - | - | - | - | - | - |
| M850 | - | - | - | - | - | - |
| M853 | +++ | +++ | +++ | ++ | ++ | +++ |
| M1001 | - | - | - | - | - | - |
| M1335 | - | - | - | - | - | - |
| M1386 | - | - | - | - | - | - |
| M1400 | - | - | - | - | - | - |
| M1407 | ++ | ++ | ++ | ++ | ++ | ++ |
| M1412 | ++ | ++ | ++ | ++ | ++ | +++ |
| M1494 | - | - | - | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohale, D.K.; Ngoepe, E.; Mparamoto, M.; Blumberg, L.; Sabeta, C.T. A Case Report on a Human Bite Contact with a Rabid Honey Badger Mellivora capensis (Kromdraai Area, Cradle of Humankind, South Africa). Trop. Med. Infect. Dis. 2023, 8, 186. https://doi.org/10.3390/tropicalmed8040186
Mohale DK, Ngoepe E, Mparamoto M, Blumberg L, Sabeta CT. A Case Report on a Human Bite Contact with a Rabid Honey Badger Mellivora capensis (Kromdraai Area, Cradle of Humankind, South Africa). Tropical Medicine and Infectious Disease. 2023; 8(4):186. https://doi.org/10.3390/tropicalmed8040186
Chicago/Turabian StyleMohale, Debrah Kgwana, Ernest Ngoepe, Munangatire Mparamoto, Lucille Blumberg, and Claude Taurai Sabeta. 2023. "A Case Report on a Human Bite Contact with a Rabid Honey Badger Mellivora capensis (Kromdraai Area, Cradle of Humankind, South Africa)" Tropical Medicine and Infectious Disease 8, no. 4: 186. https://doi.org/10.3390/tropicalmed8040186
APA StyleMohale, D. K., Ngoepe, E., Mparamoto, M., Blumberg, L., & Sabeta, C. T. (2023). A Case Report on a Human Bite Contact with a Rabid Honey Badger Mellivora capensis (Kromdraai Area, Cradle of Humankind, South Africa). Tropical Medicine and Infectious Disease, 8(4), 186. https://doi.org/10.3390/tropicalmed8040186