
Sigma-Class Glutathione Transferases (GSTσ): A New Target with Potential for Helminth Control


 ,
,
Abstract
1. Introduction
2. Glutathione Transferases (GSTs)
2.1. Cytosolic GST (cGST)
2.1.1. Glutathione (GSH)
2.1.2. Catalytic Mechanism of the Conjugation Reaction
2.1.3. Substrates and Specificity
3. Structural and Functional Generalities of GSTσ
3.1. Role of GSTσ in Helminths
GSTσ as Prostaglandin Synthase in Helminths
3.2. GSTσ of Helminths as Vaccine Candidate
3.3. GSTσ of Helminths as Drug Target
4. Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Stanley, L.A. Drug Metabolism. In Pharmacognosy Fundamentals, Applications and Strategies; Badal, S., Delgoda, R., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 527–545. [Google Scholar]
- Tsuchida, S.; Yamada, T. Glutathione Transferases. In Reference Module in Biomedical Research; Elsevier: Amsterdam, The Netherlands, 2014; pp. 1–13. [Google Scholar]
- Wu, B.; Dong, D. Human cytosolic glutathione transferases: Structure, function, and proteins: A nexus of glutathione transferases and epoxide hydrolases in a metalloenzyme superfamily. Trends Pharmacol. Sci. 2012, 33, 656–668. [Google Scholar] [CrossRef] [PubMed]
- Fabrini, R.; Bocedi, A.; Dawood, K.F.; Turella, P.; Stella, L.; Parker, M.W.; Pedersen, J.Z.; Federici, G.; Antonini, G.; Ricci, G. The extended catalysis of glutathione transferase. FEBS Lett. 2010, 585, 341–345. [Google Scholar] [CrossRef] [PubMed]
- Wilce, M.C.J.; Parker, M.W. Structure and function of glutathione S-transferases. Biochim. Biophys. Acta 1994, 1205, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Deponte, M. Glutathione catalysis and the reaction mechanisms of glutathione-dependent enzymes. Biochim. Biophys. Acta 2013, 1830, 3217–3266. [Google Scholar] [CrossRef]
- Board, P.G.; Menon, D. Glutathione transferases, regulators of cellular metabolism and physiology. Biochim. Biophys. Acta 2013, 1830, 3267–3288. [Google Scholar] [CrossRef]
- Dasaria, S.; Ganjayia, M.S.; Yellanurkondab, P.; Bashac, S.; Meriga, B. Role of glutathione S-transferases in detoxification of a polycyclic aromatic hydrocarbon, methylcholanthrene. Chem. Biol. Interac. 2018, 294, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Flanagan, J.U.; Smythe, M.L. Sigma-class glutathione transferases. Drug Metab. Rev. 2011, 43, 194–214. [Google Scholar] [CrossRef]
- Oakley, A. Glutathione transferases: A structural perspective. Drug Metab. Rev. 2011, 43, 38–151. [Google Scholar] [CrossRef]
- Sun, H.D.; Ru, Y.W.; Zhang, D.J.; Yin, S.Y.; Yin, L.; Xie, Y.Y.; Guan, Y.F.; Liu, S.Q. Proteomic analysis of glutathione S-transferase isoforms in mouse liver mitochondria. World J. Gastroenterol. 2012, 18, 3435–3442. [Google Scholar] [CrossRef]
- Torres-Rivera, A.; Landa, A. Glutathione transferase from parasites: A biochemical view. Acta Trop. 2008, 105, 99–112. [Google Scholar] [CrossRef]
- Sheehan, D.; GMeade, G.; Foley, V.M.; Dowd, C.A. Structure, Function and Evolution of Glutathione Transferases: Implications for Classification of Non-Mammalian Members of An Ancient Enzyme Superfamily. Biochem. J. 2001, 15, 1–16. [Google Scholar] [CrossRef]
- Mannervik, B.; Alin, P.T.; Guthenberg, C.; Jensson, H.; Tahir, M.K.; Warholm, M.; Jornvall, H. Identification of three classes of cytosolic glutathione transferase common to several mammalian species: Correlation between structural data and enzymatic properties. Proc. Natl. Acad. Sci. USA 1985, 82, 7202–7206. [Google Scholar] [CrossRef] [PubMed]
- Allocati, N.; Luca, F.; Masulli, M.; Di Ilio, C. Glutathione transferases in bacteria. FEBS J. 2008, 276, 58–75. [Google Scholar] [CrossRef]
- Nava, G.; Robert, L.; Plancarte, A. Characterization of Taenia solium cysticerci microsomal glutathione S-transferase activity. Parasitol. Res. 2007, 101, 1373–1381. [Google Scholar] [CrossRef] [PubMed]
- Rigsby, R.E.; Fillgrove, K.L.; Beihoffer, L.A.; Armstrong, R.N. Fosfomycin resistance drug discovery. Methods Enzymol. 2005, 401, 367–379. [Google Scholar] [PubMed]
- Armstrong, R.N.; Morgenstern, R.; Board, P.G. 10.16 Glutathione Transferases. In Comprehensive Toxicology, 3rd ed.; McQueen, C.A., Ed.; Elsevier Ltd.: Amsterdam, The Netherlands, 2018; pp. 326–362. [Google Scholar]
- Armstrong, R.N. Mechanistic Diversity in a Metalloenzyme Superfamily. Biochemistry 2000, 39, 13625–13632. [Google Scholar] [CrossRef] [PubMed]
- Hayes, J.D.; Strange, R.C. Glutathione S-Transferase Polymorphisms and Their Biological Consequences. Pharmacology 2000, 61, 154–166. [Google Scholar] [CrossRef] [PubMed]
- Ladner, J.E.; Parsons, J.F.; Rife, C.L.; Gilliland, G.L.; Armstrong, R.N. Parallel evolutionary pathways for glutathione transferases: Structure and mechanism of the mitochondrial class kappa enzyme rGSTK1-1. Biochemistry 2004, 43, 352–361. [Google Scholar] [CrossRef] [PubMed]
- Ranson, H.; Hemingway, J. Mosquito glutathione transferases. Methods Enzymol. 2005, 401, 226–241. [Google Scholar]
- Song, X.-W.; Zhong, Q.-S.; Ji, Y.-H.; Zhang, Y.-M.; Tang, J.; Feng, F.; Bi, J.-X.; Xie, J.; Li, B. Characterization of a sigma class GST (GST6) required for cellular detoxification and embryogenesis in Tribolium Castaneum. Insect Sci 2021, 29, 215–229. [Google Scholar] [CrossRef]
- Pearson, W.R. Phylogenies of Glutathione Transferase Families. In: Phase ii: Glutathione transferases. Methods Enzymol. 2005, 401, 186–204. [Google Scholar]
- Mashiyama, S.T.; Malabanan, M.M.; Akiva, E.; Bhosle, R.; Branch, M.C.; Hillerich, B.; Jagessar, K.; Kim, J.; Patskovsky, Y.; Seidel, R.D.; et al. Large-scale determination of sequence, structure, and function relationships in cytosolic glutathione transferases across the biosphere. PLoS Biol. 2014, 12, e1001843. [Google Scholar] [CrossRef] [PubMed]
- Chelvanayagam, G.; Parker, M.W.; Board, P.G. Fly fishing for GST: A unified nomenclature for mammalian and insect glutathione transferases. Chem. Biol. Interact. 2001, 133, 256–260. [Google Scholar]
- Atkinson, H.J.; Babbitt, P.C. Glutathione Transferases Are Structural and Functional Outliers in the Thioredoxin Fold. Biochemistry 2009, 48, 11108–11116. [Google Scholar] [CrossRef] [PubMed]
- Buetler, T.M.; Eaton, D.L. Glutathione S-transferases: Amino acid sequence comparison, classification and phylogenetic relationship. J. Environ. Sci. Health C Toxicol. 1992, 10, 181–203. [Google Scholar] [CrossRef]
- Abdalla, A.M.; Bruns, C.M.; Tainer, J.A.; Mannervik, B.; Stenberg, G. Design of a monomeric human glutathione transferase GSTP1, a structurally stable but catalytically inactive protein. Protein Eng. 2002, 15, 827–834. [Google Scholar] [CrossRef] [PubMed]
- Oakley, A.J.; Lo Bello, M.; Mazzetti, A.P.; Federici, G.; Parker, M.W. The glutathione conjugate of ethacrynic acid can bind to human pi class glutathione transferase P1-1 in two different modes. FEBS Lett. 1997, 419, 32–36. [Google Scholar] [CrossRef] [PubMed]
- Denzoin, V.L.A.; Soraci, A.L.; Tapia, M.O. Homeostasis del glutatión. Acta Bioquim. Clin. Latinoam. 2013, 47, 529–539. [Google Scholar]
- Singhal, S.S.; Singh, P.S.; Singhal, P.; Horne, D.; Singhal, J.; Awasthi, S. Antioxidant Role of Glutathione S-Transferases: 4-Hydroxynonenal, a Key Molecule in Stress-Mediated Signaling. Toxicol. Appl. Pharmacol. 2015, 289, 361–370. [Google Scholar] [CrossRef]
- Board, P.G. Transport of Glutathione S-Conjugate from human erythrocytes. FEBS Lett. 1981, 124, 163–165. [Google Scholar] [CrossRef]
- Wheeler, J.B.; Stourman, N.V.; Their, R.; Dommermuth, A.; Vuilleumier, S.; Rose, J.A.; Armstrong, R.N.; Guengerich, F.P. Conjugation of haloalkanes by bacterial and mammalian glutathione transferases: Mono-and dihalomethanes. Chem. Res. Toxicol. 2001, 14, 1118–1127. [Google Scholar] [CrossRef] [PubMed]
- Danielson, U.H.; Esterbauer, H.; Mannervik, B. Structure activity relationships of 4-hydroxyalkenals in the conjugation catalyzed by mammalian glutathione transferases. Biochem. J. 1987, 247, 707–713. [Google Scholar] [CrossRef] [PubMed]
- Labrou, N.E.; Karavangeli, M.; Tsaftaris, A.; Clonis, Y.D. Kinetic analysis of maize glutathione S-transferase I catalyzing the detoxification from chloroacetanilide herbicides. Planta 2005, 222, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.; Crowley, P.; Moreman, A.D.; Barrett, J. Biochemical properties of cloned glutathione S-transferases from Schistosoma mansoni and Schistosoma japonicum. Mol. Biochem. Parasitol. 1993, 61, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Arbildi, P.; La-Rocca, S.; Lopez, V.; Da-Costa, N.; Fernandez, V. Echinococcus granulosus: Evidence of a heterodimeric glutathione transferase built up by phylogenetically distant subunits. Mol. Biochem. Parasitol. 2017, 211, 26–30. [Google Scholar] [CrossRef] [PubMed]
- Meyer, D.J.; Muimo, R.; Thomas, M.; Coates, D.; Isaac, R.E. Purification and characterization of prostaglandin-H E-isomerase, a sigma-class glutathione S-transferase, from Ascaridia galli. Biochem. J. 1996, 313, 223–227. [Google Scholar] [CrossRef] [PubMed]
- LaCourse, E.J.; Perally, S.; Morphew, R.M.; Moxon, J.V.; Prescott, M.; Dowling, D.J.; O’Neill, S.M.; Kipar, A.; Hetzel, U.; Hoey, E.; et al. The Sigma Class Glutathione Transferase from the Liver Fluke Fasciola hepatica. PLoS Negl. Trop. Dis. 2012, 6, e1666. [Google Scholar] [CrossRef] [PubMed]
- Duncan, J.; Cutress, D.; Morphew, R.M.; Brophy, P.M. Purification of native sigma Class Glutathione Transferase from Fasciola hepatica. Mol. Biochem. Parasitol. 2018, 222, 45–50. [Google Scholar] [CrossRef]
- Cutress, D.J. Towards Validation of the Sigma Class GST from the Liver Fluke Fasciola hepatica as Chemotherapeutic Targets. Ph.D. Dissertation, Aberystwyth University, Department of Life Sciences, Aberystwyth, UK, 2019. [Google Scholar]
- Sommer, A.; Rickert, R.; Fischer, P.; Steinhart, H.; Walter, R.D.; Liebau, E. A dominant role for extracellular glutathione S-transferase from Onchocerca volvulus is the production of prostaglandin D2. Infect. Immun. 2003, 71, 3603–3606. [Google Scholar] [CrossRef]
- Nguyen, H.A.; Bae, Y.A.; Lee, E.G.; Kim, S.H.; Diaz, S.; Camacho, P.; Nawa, Y.; Kang, I.; Kong, Y. A novel Sigma-like glutathione transferase of Taenia solium metacestode. Int. J. Parasitol. 2010, 40, 1097–1106. [Google Scholar] [CrossRef]
- Hirowatari, A.; Nagaoka, S.; Yamada, N.; Yamamoto, K. Identifying a sigma class glutathione S-transferase 2 from the silkworm Bombyx mori. J. Insect Biotechnol. Sericol. 2017, 9, 1_001–1_007. [Google Scholar]
- Singh, S.P.; Coronella, J.A.; Benes, H.; Cochrane, B.J.; Zimniak, P. Catalytic function of Drosophila melanogaster glutathione S-transferase DmGST1-1 (GST-2) in conjugation of lipid peroxidation end products. Eur. J. Biochem. 2001, 268, 2912–2923. [Google Scholar] [CrossRef]
- Yamamoto, K.; Fujii, Y.H.; Aso, Y.; Banno, Y.; Koga, K. Expression and Characterization of a Sigma-Class Glutathione S-Transferase of the Fall Webworm, Hyphantria cunea. Biosci. Biotechnol. Biochem. 2007, 71, 553–560. [Google Scholar] [CrossRef]
- Qin, G.; Jia, M.; Liu, T.; Zhang, X.; Guo, Y.; Zhu, K.Y.; Ma, E.; Zhang, J. Characterization and Functional Analysis of four Glutathione S-Transferases from the Migratory Locust, Locusta migratoria. PLoS ONE 2013, 8, e58410. [Google Scholar] [CrossRef]
- Hassan, F.; Singh, K.P.; Ali, V.; Behera, S.; Shivam, P.; Das, P.; Dinesh, D.S. Detection and functional characterization of sigma class GST in Phlebotomus argentipes and its role in stress tolerance and DDT resistance. Sci. Rep. 2019, 9, 19636. [Google Scholar] [CrossRef] [PubMed]
- Valles, S.M.; Perer, P.O.; Strong, A.C. Purification, biochemical characterization, and cDNA cloning of a glutathione S-transferase from the red imported fire ant, Solenopsis invicta. Insect Biochem. Mol. Biol. 2003, 33, 981–988. [Google Scholar] [CrossRef] [PubMed]
- Abdalla, A.M.; Ghada, S.A.; Karim, S.A.A. Biochemical characterization and peptide mass fingerprinting of two glutathione transferases from Biomphalaria alexandrina snails (Gastropoda: Planorbidae). J. Genet. Eng. Biotechnol. 2022, 20, 99. [Google Scholar] [CrossRef]
- Li, H.; Yang, Z.; Huang, Q.; Li, Y. Molecular cloning and characterization of a sigma-class glutathione S-Transferase from the freshwater mussel Hyriopsis cumingii. Microbiol. Immunol. 2015, 59, 219–230. [Google Scholar] [CrossRef]
- Harris, J.; Coles, B.; Meyer, D.J.; Kettere, B. The Isolation and Characterization of The Major Glutathione S-Transferase from The Squid Loligo vulgaris. Comp. Biochem. Physiol. Part-B Biochem. Mol. Biol. 1991, 98, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Umasuthan, N.; Revathy, K.S.; Lee, Y.; Whang, I.; Choi, C.Y.; Lee, J. A novel molluscan sigma-like glutathione S-transferase from Manila clam, Ruditapes philippinarum: Cloning, characterization and transcriptional profiling. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2012, 155, 539–550. [Google Scholar] [CrossRef]
- Polekhina, G.; Board, P.G.; Blackburn, A.C.; Parker, M.W. Crystal Structure of Maleylacetoacetate Isomerase/Glutathione Transferase Zeta Reveals the Molecular Basis for Its Remarkable Catalytic Promiscuity. Biochemistry 2001, 40, 1567–1576. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.; von Rosenvinge, E.C.; Johnson, W.W.; Armstrongw, R.N.; Gilliland, G.L. Location of a potential transport binding site in a sigma class glutathione transferase by x-ray crystallography. Proc. Natl. Acad. Sci. USA 1996, 93, 8208–8213. [Google Scholar] [CrossRef] [PubMed]
- Line, K.; Isupov, M.N.; LaCourse, E.J.; Cutress, D.J.; Morphew, R.M.; Brophy, P.M.; Littlechild, J.A. X-ray structure of Fasciola hepatica sigma class glutathione transferase 1 reveals a disulfide bond to support stability in gastro-intestinal environment. Sci. Rep. 2019, 9, 902. [Google Scholar] [CrossRef] [PubMed]
- Kanaoka, Y.; Ago, H.; Inagaki, E.; Nanayama, T.; Miyano, M.; Kikuno, R.; Fujii, Y.; Eguchi, N.; Toh, H.; Urade, Y.; et al. Cloning and Crystal Structure of Hematopoietic Prostaglandin D Synthase. Cell 1997, 90, 1085–1095. [Google Scholar] [CrossRef] [PubMed]
- Kanaoka, Y.; Fujimori, K.; Kikuno, R.; Sakaguchi, Y.; Urade, Y.; Hayaishi, O. Structure and chromosomal localisation of human and mouse genes for haematopoietic prostaglandin D synthase. Eur. J. Biochem. 2000, 267, 3315–3322. [Google Scholar] [CrossRef] [PubMed]
- Hervé, M.; Angeli, V.; Pinzar, E.; Wintjens, R.; Faveeuw, C.; Narumiya, S.; Capron, A.; Urade, Y.; Capron, M.; Riveau, G.; et al. Pivotal roles of the parasite PGD2 synthase and of the host D prostanoid receptor 1 in schistosome immune evasión. Eur. J. Inmunol. 2003, 33, 2764–2772. [Google Scholar] [CrossRef]
- Axarli, I.; Dhavala, P.; Papageorgiou, A.C.; Labrou, N.E. Crystallographic and Functional Characterization of the Fluorodifen-inducible Glutathione Transferase from Glycine max Reveals an Active Site Topography Suited for Diphenylether Herbicides and a Novel L-site. J. Mol. Biol. 2009, 385, 984–1002. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K.; Higashiura, A.; Suzuki, M.; Aritake, K.; Urade, Y.; Uodome, N.; Nakagawa, A. Crystal structure of a Bombyx mori sigma-class glutathione transferase exhibiting prostaglandin E synthase activity. Biochim. Biophys. Acta 2013, 1830, 3711–3718. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.; von Rosenvinge, E.C.; Johnson, W.W.; Tomarev, S.I.; Piatigorsky, J.; Armstrong, R.N.; Gilliland, G.L. Three-dimensional structure, catalytic properties, and evolution of a sigma class glutathione transferase from squid, a progenitor of the lens S-crystallins of cephalopods. Biochemistry 1995, 34, 5317–5328. [Google Scholar] [CrossRef]
- Geerts, S.; Gryseels, B. Drug resistance in human helminths: Current situation and lessons from livestock. Clin. Microbiol. Rev. 2000, 13, 207–222. [Google Scholar] [CrossRef]
- Brophy, P.M.; Barrett, J. Glutathione transferase in helminths. Parasitology 1990, 100, 345–349. [Google Scholar] [CrossRef] [PubMed]
- Cwiklinski, K.; Dalton, J.P.; Dufresne, P.J.; LaCourse, J.; Williams, D.J.L.; Hodgkinson, J.; Paterson, S. The Fasciola hepatica genome: Gene duplication and polymorphism reveal adaptation to the host environment and the capacity for rapid evolution. Genome Biol. 2015, 16, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Eida, A.M. Proteinoids and parasitic diseases. Parasitol. United J. 2015, 8, 38–51. [Google Scholar] [CrossRef]
- Miller, W.D.; Dill, K.D. Ligand binding to proteins: The binding landscape model. Protein Sci. 1997, 6, 2166–2179. [Google Scholar] [CrossRef] [PubMed]
- Angeli, V.; Faveeuw, C.; Roye, O.; Fontaine, J.; Teissier, E.; Capron, A.; Wolowczuk, I.; Capron, M.; Trottein, F. Role of the parasite-derived prostaglandin D2 in the inhibition of epidermal Langerhans cell migration during schistosomiasis infection. J. Exp. Med. 2001, 193, 1135–1147. [Google Scholar] [CrossRef] [PubMed]
- Cervonic, V.; Houston, S.A.; Scott, C.L.; Aumeunier, A.; Yrlid, U.; Mowat, A.M.; Milling SW, F. Intestinal CD103− dendritic cells migrate in lymph and prime effector T cells. Mucosal Immunol. 2013, 6, 104–113. [Google Scholar]
- Rose, C.; Praulins, G.; Armstrong, S.D.; Casas-Sanchez, A.; Davis, J.; Molyneux, G.; Yunta, C.; Stead, Z.; Prescott, M.; Perally, S.; et al. Characterization of a novel glycosylated glutathione transferase of Onchocerca ochengi, closest relative of the human river blindness parasite. Parasitology 2019, 146, 1773–1784. [Google Scholar] [CrossRef] [PubMed]
- Faveeuw, C.; Mallevaey, T.; Paschinger, K.; Fontaine, W.J.; Mollicone, R.; Oriol, R.; Altmann, F.; Lerouge, P.; Capron, M.; Trottei, F. Schistosome N-glycans containing core α3-fucose and core β2-xylose epitopes are strong inducers of Th2 responses in mice. Eur. J. Immunol. 2003, 33, 1271–1281. [Google Scholar] [CrossRef] [PubMed]
- Toet, H.; Piedrafita, D.M.; Spithill, T.W. Liver fluke vaccines in ruminants: Strategies, progress and future opportunities. Int. J. Parasitol. 2014, 44, 915–927. [Google Scholar] [CrossRef]
- Spithill, T.W.; Piedrafita, D.; Smooker, P.M. Immunological approaches for the control of fasciolosis. Int. J. Parasitol. 1997, 27, 1221–1235. [Google Scholar] [CrossRef]
- Diemert, D.J.; Bottazzi, M.E.; Plieskatt, J.; Hotez, P.J.; Bethony, J.M. Lessons along the Critical Path: Developing Vaccines against Human Helminths. Trends Parasitol. 2018, 34, 749–758. [Google Scholar] [CrossRef] [PubMed]
- Riveau, G.; Schacht, A.M.; Dompnier, J.P.; Deplanque, D.; Seck, M.; Waucquier, N.; Senghor, S.; Delcroix-Genete, D.; Hermann, I.E.; Idris-Khodja, I.N.; et al. Safety and efficacy of the rSh28GST urinary schistosomiasis vaccine: A phase 3 randomized, controlled trial in Senegalese children. PLoS Negl. Trop. Dis. 2018, 12, e0006968. [Google Scholar] [CrossRef] [PubMed]
- Miranda-Blancas, R.; Rodríguez-Lima, O.; García-Gutiérrez, P.; Flores-López, R.; Jiménez, L.; Zubillaga, R.A.; Rudiño-Piñera, E.; Landa, A. Biochemical characterization and gene structure analysis of the 24-kDa glutathione transferase sigma from Taenia solium. FEBS Open Bio 2024, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Vibanco-Pérez, N.; Jimenez, L.; Mendoza-Hernandez, G.; Landa, A. Characterization of a recombinant mu-class glutathione S-transferase from Taenia solium. Parasitol. Res. 2002, 88, 398–404. [Google Scholar]
- Jensen, O.N. Interpreting the protein language using proteomics. Nat. Rev. Mol. Cell Biol. 2006, 7, 391–403. [Google Scholar] [CrossRef] [PubMed]
- Molina-Hernández, V.; Mulcahy, G.; Pérez, J.; Martínez-Moreno, A.; Donnelly, S.; O’Neill, S.M.; Dalton, J.P.; Cwiklinski, K. Fasciola hepatica vaccine: We may not be there yet but we’re on the right road. Vet. Parasitol. 2015, 208, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Johnson, K.A.; Angelucci, F.; Bellelli, A.; Hervé, M.; Fontaine, J.; Tsernoglou, D.; Capron, A.; Trottein, F.; Brunori, M. Crystal Structure of the 28 kDa Glutathione S-Transferase from Schistosoma haematobium. Biochemistry 2003, 42, 10084–10094. [Google Scholar] [CrossRef] [PubMed]
- Riveau, G.; Deplanque, D.; Remoue, F.; Schacht, A.M.; Capron, H.V.M.; Thiry, M.; Martial, J.; Libersa, C.; Capron, A. Safety and Immunogenicity of rSh28GST Antigen in Humans: Phase 1 Randomized Clinical Study of a Vaccine Candidate against Urinary Schistosomiasis. PLoS Negl. Trop. Dis. 2012, 6, e1704. [Google Scholar] [CrossRef]
- Zhan, B.; Perally, S.; Brophy, P.M.; Xue, J.; Goud, G.; Liu, S.; Deumic, V.; de Oliveira, L.M.; Bethony, J.; Bottazzi, M.E.; et al. Molecular cloning, biochemical characterization, and partial protective immunity of the heme-binding glutathione S-transferases from the human hookworm Necator americanus. Infect. Immun. 2010, 78, 1552–1563. [Google Scholar] [CrossRef]
- Diemert, D.J.; Freire, J.; Valente, V.; Fraga, C.G.; Talles, F.; Grahek, S.; Campbell, D.; Jariwala, A.; Periago, M.V.; Enk, M.; et al. Safety and immunogenicity of the Na-GST-1 hookworm vaccine in Brazilian and American adults. PLoS Negl. Trop. Dis. 2017, 11, e0005574, Erratum in: PLoS Negl. Trop. Dis. 2020, 14, e0008670. [Google Scholar] [CrossRef]
- Zinsou, J.F.; Diemert, D.J.; Dejon-Agobé, J.C.; Adégbité, B.R.; Honkpehedji, Y.J.; Vodonou, K.G.; Bikangui, R.; Edoa, J.R.; Massinga Loembe, M.; Li, G.; et al. Safety and immunogenicity of the co-administered Na-APR-1 and Na-GST-1 hookworm vaccines in school-aged children in Gabon: A randomised, controlled, observer-blind, phase 1, dose-escalation trial. Lancet Infect. Dis. 2024. [Google Scholar] [CrossRef] [PubMed]
- García-Gutiérrez, P.; Zubillaga, R.A.; Téllez-Plancarte, A.; Flores-López, R.; Camarillo-Cadena, M.; Landa, A. Discovery of a new non-substrate inhibitor of the 26.5 kDa glutathione transferase from Taenia solium by virtual screening. J. Mol. Graph Model. 2020, 100, 107707. [Google Scholar] [CrossRef] [PubMed]


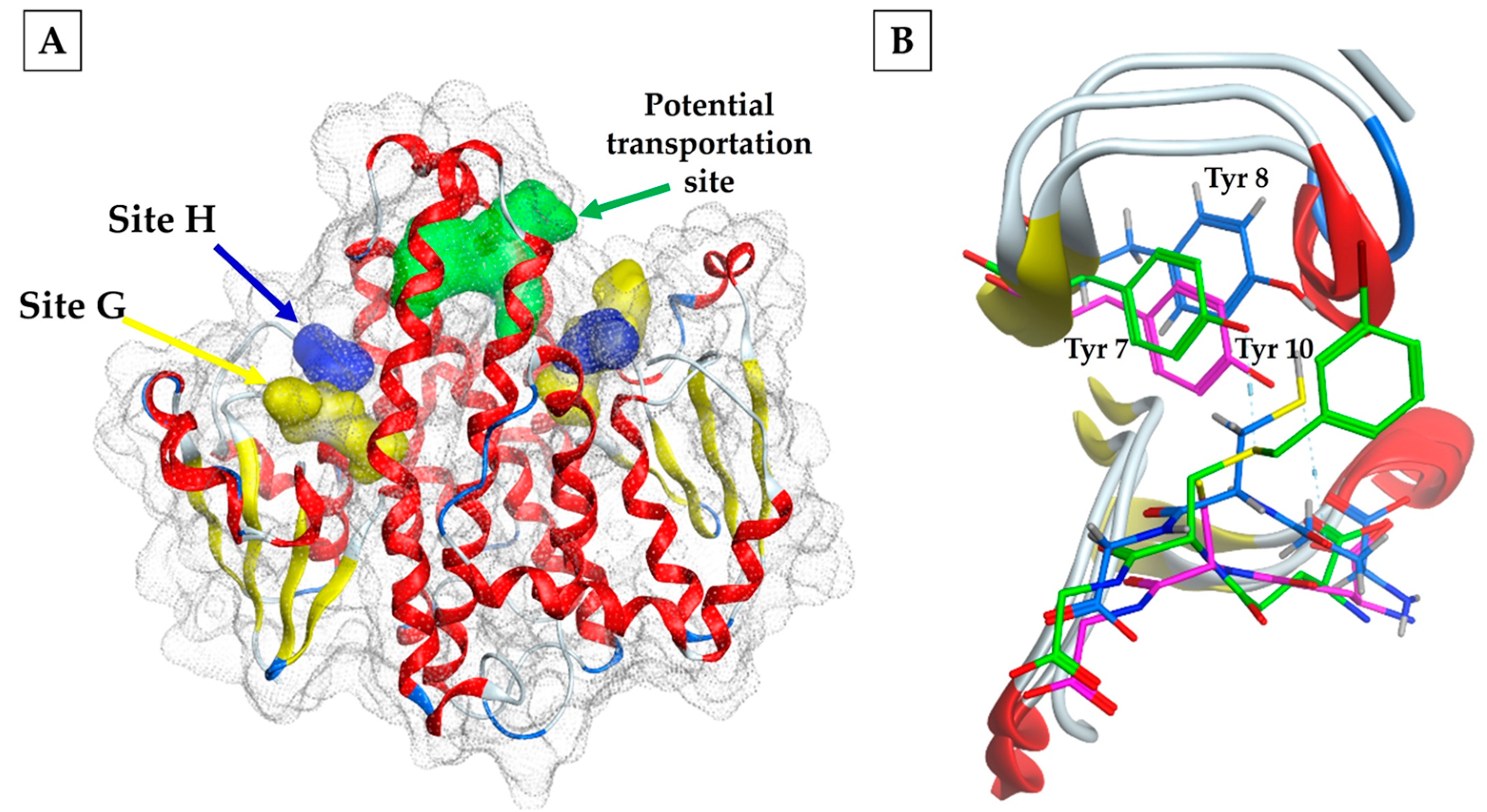{kind=link}
| G S T | Cytosolic | Involved in metabolization of xenobiotics, drugs, and Insecticides and in immunomodulation via the synthesis of prostaglandin D2 | Mammals: Classes: alpha, mu, pi, sigma, theta, zeta, omega Plants: Classes: phi, tau, lambda, iota, theta, sigma, zeta Insects and Helminths Classes: alfa-mu, epsilon, delta, mu, sigma, omega, theta, zeta |
| Mitochondrial | They participate in energy and lipid metabolism in the mitochondria | Humans Class: Kappa Helminths Unclassified | |
| MAPEG (Membrane-associated) | Involved in the biosynthesis of eicosanoids, glutathione, and prostaglandin E2 | Humans
|
| Organism | GSTσ | Activity on Different Substrates (µmol/min/mg) | ||||||
|---|---|---|---|---|---|---|---|---|
| CDNB a | DCNB b | EAC c | HEN d | CHP e | PC f | Reference | ||
| Helminths | ||||||||
| Ascaris galli | n | N.D. | N.D. | 260 | 0.5 | 1 | Detected | [39] |
| Fasciola hepatica | rFhGST-S1 | 474 ± 292 | N.D. | 898 ± 204 | 645 ± 129 | 7080 ± 1010 | Detected | [40] |
| rFhGSTs1a | 13,900 ± 907 | [41] | ||||||
| nFhGSTs1b | 7360± 216 | N.D. | 1730 ± 212 | [42] | ||||
| Onchocerca volvulus | nOv-GST1 | 10 | 0.649± 0.015 | N.D. | Detected | [43] | ||
| Schistosoma mansoni | rSm28GST | 7270 ± 25 | 1580 ± 97 | 158 ± 97 | 287 ± 17 | 167 ± 7 | Detected | [37] |
| Taenia solium | rTsMσGST | 1.08 | N.D. | N.D. | 8.4 ± 0.4 | [44] | ||
| Insects | ||||||||
| Bombyx mori | rbmGSTS2 | 0.006 | 0.04 | 5.43 | Detected | [45] | ||
| Drosophila melanogaster | nDmGSTS1-1 | 0.49 ± 0.02 | 0.44 ± 0.01 | 0.24 ± 0.02 | 9.4 ± 0.6 | 0.173 | [46] | |
| Hyphantria cunea | nhcGST | 6.99 ± 0.78 | 0.90 ± 0.09 | [47] | ||||
| Migratory locusta | rLmGSTs5 | 3890 ± 962 | 7.41 ± 0.96 | [48] | ||||
| Phlebotomus argentipes | rPargGSTσ | 8.75 | 204 | 92.5 | [49] | |||
| Solenopsis invicta | nSiGSTS1 | 87.4 ± 5.1 | [50] | |||||
| Tribolium castaneum | rTcGSTS6 | 3.8 | 176 | 66.7 | [23] | |||
| Mollusks | ||||||||
| Biomphalaria alexandrina | nBaGST2 | 30 ± 1.5 | 0.60 ± 0.02 | [51] | ||||
| Hyriopsis cumingii | rHcGSTS | 4.54 ± 0.08 | [52] | |||||
| Loligo vulgaris | nL. vulgarisGST | 273 | 0.6 | N.D. | 17.7 | 1.2 | [53] | |
| Ruditapes philippinarum | rRpGSTσ | 4.6 ± 0.17 | 0.28 ± 0.03 | 0.39 ± 0.02 | N.D. | [54] | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez Pérez, L.d.C.; Zubillaga, R.A.; García-Gutiérrez, P.; Landa, A. Sigma-Class Glutathione Transferases (GSTσ): A New Target with Potential for Helminth Control. Trop. Med. Infect. Dis. 2024, 9, 85. https://doi.org/10.3390/tropicalmed9040085
Sánchez Pérez LdC, Zubillaga RA, García-Gutiérrez P, Landa A. Sigma-Class Glutathione Transferases (GSTσ): A New Target with Potential for Helminth Control. Tropical Medicine and Infectious Disease. 2024; 9(4):85. https://doi.org/10.3390/tropicalmed9040085
Chicago/Turabian StyleSánchez Pérez, Lluvia de Carolina, Rafael A. Zubillaga, Ponciano García-Gutiérrez, and Abraham Landa. 2024. "Sigma-Class Glutathione Transferases (GSTσ): A New Target with Potential for Helminth Control" Tropical Medicine and Infectious Disease 9, no. 4: 85. https://doi.org/10.3390/tropicalmed9040085
APA StyleSánchez Pérez, L. d. C., Zubillaga, R. A., García-Gutiérrez, P., & Landa, A. (2024). Sigma-Class Glutathione Transferases (GSTσ): A New Target with Potential for Helminth Control. Tropical Medicine and Infectious Disease, 9(4), 85. https://doi.org/10.3390/tropicalmed9040085