
Reduced Gut Bacterial Diversity in Early Life Predicts Feeding Intolerance in Preterm Neonates

 , , , , and
, , , , and 
Abstract
:1. Introduction
2. Methods
2.1. Population and Sampling
2.2. Feeding Protocol
2.3. Ethics
2.4. Metagenomic DNA Extraction, Illumina MiSeq Sequencing
2.5. Statistical Analysis
3. Results
3.1. The Pielou Evenness Index between Tolerant and Intolerant Neonates
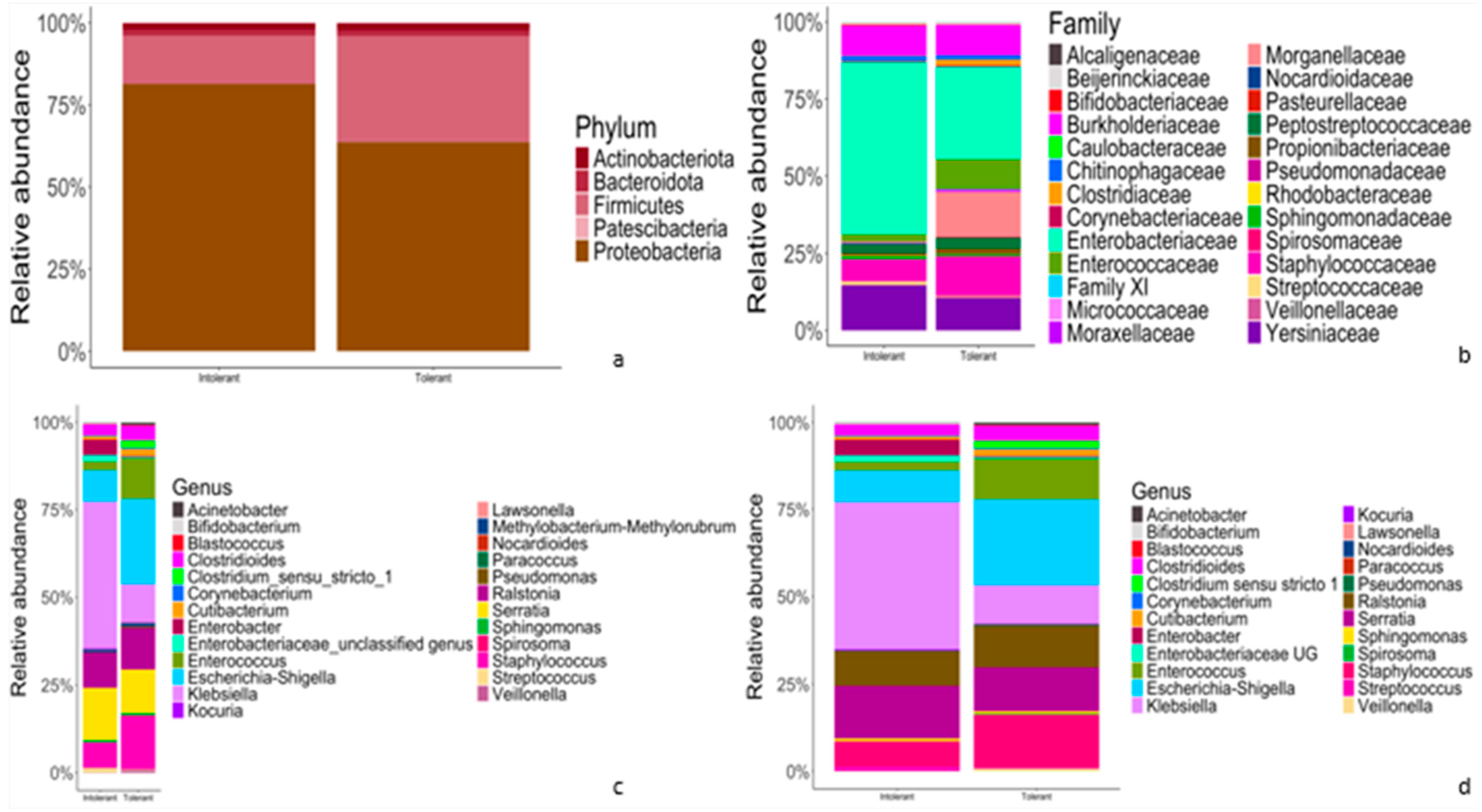
3.2. Relative Abundance of Taxonomic Features of Gut Microbiota between Tolerant and Intolerant Neonates
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fanaro, S.; Chierici, R.; Guerrini, P.; Vigi, V. Intestinal microflora in early infancy: Composition and development. Acta Paediatr. 2007, 92, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Isolauri, E. Development of healthy gut microbiota early in life: Microbiota and health in early life. J. Paediatr. Child Health 2012, 48, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Neu, J.; Mshvildadze, M.; Mai, V. A roadmap for understanding and preventing necrotizing enterocolitis. Curr. Gastroenterol. Rep. 2008, 10, 450–457. [Google Scholar] [CrossRef] [PubMed]
- Saavedra, J.M.; Dattilo, A.M. Early Development of Intestinal Microbiota. Gastroenterol. Clin. N. Am. 2012, 41, 717–731. [Google Scholar] [CrossRef] [PubMed]
- Amir, A.; Erez-Granat, O.; Braun, T.; Sosnovski, K.; Hadar, R.; BenShoshan, M.; Heiman, S.; Abbas-Egbariya, H.; Glick Saar, E.; Efroni, G.; et al. Gut microbiome development in early childhood is affected by day care attendance. NPJ Biofilms Microbiomes 2022, 8, 2. [Google Scholar] [CrossRef] [PubMed]
- Di Mauro, A.; Neu, J.; Riezzo, G.; Raimondi, F.; Martinelli, D.; Francavilla, R.; Indrio, F. Gastrointestinal function development and microbiota. Ital. J. Pediatr. 2013, 39, 15. [Google Scholar] [CrossRef] [PubMed]
- Walls Castellanos, M.; Claud, E.C. The microbiome, guard or threat to infant health. Trends Mol. Med. 2021, 27, 1175–1186. [Google Scholar] [CrossRef] [PubMed]
- Heida, F.H.; Van Zoonen, A.G.J.F.; Hulscher, J.B.F.; Te Kiefte, B.J.C.; Wessels, R.; Kooi, E.M.W.; Bos, A.F.; Harmsen, H.J.M.; De Goffau, M.C. A Necrotizing Enterocolitis-Associated Gut Microbiota Is Present in the Meconium: Results of a Prospective Study. Clin. Infect. Dis. 2016, 62, 863–870. [Google Scholar] [CrossRef] [PubMed]
- Penders, J.; Thijs, C.; Vink, C.; Stelma, F.F.; Snijders, B.; Kummeling, I.; van den Brandt, P.A.; Stobberingh, E.E. Factors influencing the composition of the intestinal microbiota in early infancy. Pediatrics 2006, 118, 511–521. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Ao, D.; Cai, X.; Huang, P.; Cai, N.; Lin, S.; Wu, B. Early gut microbiota in very low and extremely low birth weight preterm infants with feeding intolerance: A prospective case-control study. J. Microbiol. 2022, 60, 1021–1031. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.-C.; Sun, Q.; Ji, Y.-C.; Fu, L.-Z.; Wang, Z.-L.; He, Y.; Li, L.-Q. Differences in the Gut Microbiota Composition and Metabolites Associated With Feeding Intolerance in VLBW Infants With a Gestational Age of ≤30 Weeks: A Pilot Study. Front. Cell Infect. Microbiol. 2022, 12, 726322. [Google Scholar] [CrossRef] [PubMed]
- Ruoss, J.L.; Bazacliu, C.; Russell, J.T.; de la Cruz, D.; Li, N.; Gurka, M.J.; Filipp, S.L.; Polin, R.A.; Triplett, E.W.; Neu, J. Routine Early Antibiotic Use in SymptOmatic Preterm Neonates: A Pilot Randomized Controlled Trial. J. Pediatr. 2021, 229, 294–298.e3. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Yan, J.; Wen, H.; Deng, X.; Li, X.; Su, S. Feeding intolerance alters the gut microbiota of preterm infants. PLoS ONE 2019, 14, e0210609. [Google Scholar] [CrossRef] [PubMed]
- Fanaro, S. Feeding intolerance in the preterm infant. Early Hum. Dev. 2013, 89, S13–S20. [Google Scholar] [CrossRef] [PubMed]
- Vitali, R.; Terrin, G.; Palone, F.; Laudadio, I.; Cucchiara, S.; Boscarino, G.; Di Chiara, M.; Stronati, L. Fecal High-Mobility Group Box 1 as a Marker of Early Stage of Necrotizing Enterocolitis in Preterm Neonates. Front. Pediatr. 2021, 9, 672131. [Google Scholar] [CrossRef] [PubMed]
- Terrin, G.; Stronati, L.; Cucchiara, S.; De Curtis, M. Serum Markers of Necrotizing Enterocolitis: A Systematic Review. J. Pediatr. Gastroenterol. Nutr. 2017, 65, e120–e132. [Google Scholar] [CrossRef] [PubMed]
- Qiao, L.-X.; Zhu, W.-Y.; Zhang, H.-Y.; Wang, H. Effect of early administration of probiotics on gut microflora and feeding in pre-term infants: A randomized controlled trial. J. Matern.-Fetal Neonatal Med. 2017, 30, 13–16. [Google Scholar] [CrossRef] [PubMed]
- Patel, R.M.; Ferguson, J.; McElroy, S.J.; Khashu, M.; Caplan, M.S. Defining Necrotizing Enterocolitis: Current Difficulties and Future Opportunities. Pediatr. Res. 2020, 88, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Peddada, S.D. Analysis of compositions of microbiomes with bias correction. Nat. Commun. 2020, 11, 3514. [Google Scholar] [CrossRef] [PubMed]
- Björkström, M.V.; Hall, L.; Söderlund, S.; Håkansson, E.G.; Håkansson, S.; Domellöf, M. Intestinal flora in very low-birth weight infants. Acta Paediatr. 2009, 98, 1762–1767. [Google Scholar] [CrossRef] [PubMed]
- Schwiertz, A.; Gruhl, B.; Löbnitz, M.; Michel, P.; Radke, M.; Blaut, M. Development of the Intestinal Bacterial Composition in Hospitalized Preterm Infants in Comparison with Breast-Fed, Full-Term Infants. Pediatr. Res. 2003, 54, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.; Liao, X.-S.; Yin, T.; Liu, X.-C.; Bao, L.; Li, L.-Q. Alterations of the gut microbiota and short chain fatty acids in necrotizing enterocolitis and food protein-induced allergic protocolitis infants: A prospective cohort study. Front. Cell Infect. Microbiol. 2022, 12, 1030588. [Google Scholar] [CrossRef] [PubMed]
- Pammi, M.; Cope, J.; Tarr, P.I.; Warner, B.B.; Morrow, A.L.; Mai, V.; Gregory, K.E.; Kroll, J.S.; McMurtry, V.; Ferris, M.J.; et al. Intestinal dysbiosis in preterm infants preceding necrotizing enterocolitis: A systematic review and meta-analysis. Microbiome 2017, 5, 31. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Chang, Y.; Wang, Z.; Bao, W.; Li, Z. Altered gut microbiota is associated with feeding intolerance in preterm infants. Turk. J. Pediatr. 2021, 63, 206. [Google Scholar] [CrossRef] [PubMed]
- La Rosa, P.S.; Warner, B.B.; Zhou, Y.; Weinstock, G.M.; Sodergren, E.; Hall-Moore, C.M.; Stevens, H.J.; Bennett, W.E.; Shaikh, N.; Linneman, L.A.; et al. Patterned progression of bacterial populations in the premature infant gut. Proc. Natl. Acad. Sci. USA 2014, 111, 12522–12527. [Google Scholar] [CrossRef] [PubMed]
- Millar, M.R.; Linton, C.J.; Cade, A.; Glancy, D.; Hall, M.; Jalal, H. Application of 16S rRNA gene PCR to study bowel flora of preterm infants with and without necrotizing enterocolitis. J. Clin. Microbiol. 1996, 34, 2506–2510. [Google Scholar] [CrossRef] [PubMed]
- Smith, B.; Bodé, S.; Skov, T.H.; Mirsepasi, H.; Greisen, G.; Krogfelt, K.A. Investigation of the early intestinal microflora in premature infants with/without necrotizing enterocolitis using two different methods. Pediatr. Res. 2012, 71, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Shan, G.; Sodergren, E.; Weinstock, G.; Walker, W.A.; Gregory, K.E. Longitudinal Analysis of the Premature Infant Intestinal Microbiome Prior to Necrotizing Enterocolitis: A Case-Control Study. PLoS ONE 2015, 10, e0118632. [Google Scholar] [CrossRef] [PubMed]
- Hagerty, S.L.; Hutchison, K.E.; Lowry, C.A.; Bryan, A.D. An empirically derived method for measuring human gut microbiome alpha diversity: Demonstrated utility in predicting health-related outcomes among a human clinical sample. PLoS ONE 2020, 15, e0229204. [Google Scholar] [CrossRef] [PubMed]
- Young, V.B.; Schmidt, T.M. Overview of the Gastrointestinal Microbiota. Adv. Exp. Med. Biol. 2008, 635, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Arboleya, S.; Binetti, A.; Salazar, N.; Fernández, N.; Solís, G.; Hernández-Barranco, A.; Margolles, A.; los Reyes-Gavilán, C.G.; Gueimonde, M. Establishment and development of intestinal microbiota in preterm neonates. FEMS Microbiol. Ecol. 2012, 79, 763–772. [Google Scholar] [CrossRef] [PubMed]
- Palmer, C.; Bik, E.M.; DiGiulio, D.B.; Relman, D.A.; Brown, P.O. Development of the Human Infant Intestinal Microbiota. PLoS Biol. 2007, 5, e177. [Google Scholar] [CrossRef] [PubMed]
- York, A. Microbiota Succession in Early Life. Nat. Res. 2019. Available online: https://www.nature.com/articles/d42859-019-00010-6 (accessed on 11 July 2024).
- Kamada, N.; Chen, G.Y.; Inohara, N.; Núñez, G. Control of pathogens and pathobionts by the gut microbiota. Nat. Immunol. 2013, 14, 685–690. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Li, H.; Jia, J.; Liu, Y.; Zhang, N.; Wang, R.; Qu, W.; Liu, Y.; Jia, L. Mechanisms by which the intestinal microbiota affects gastrointestinal tumours and therapeutic effects. Mol. Biomed. 2023, 4, 45. [Google Scholar] [CrossRef] [PubMed]
- Shin, N.-R.; Whon, T.W.; Bae, J.-W. Proteobacteria: Microbial signature of dysbiosis in gut microbiota. Trends Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef] [PubMed]
- Jost, T.; Lacroix, C.; Braegger, C.P.; Chassard, C. New insights in gut microbiota establishment in healthy breast fed neonates. PLoS ONE 2012, 7, e44595. [Google Scholar] [CrossRef] [PubMed]
- Torow, N.; Hornef, M.W. The Neonatal Window of Opportunity: Setting the Stage for Life-Long Host-Microbial Interaction and Immune Homeostasis. J. Immunol. 2017, 198, 557–563. [Google Scholar] [CrossRef]
- Walker, W.A.; Iyengar, R.S. Breast milk, microbiota, and intestinal immune homeostasis. Pediatr. Res. 2015, 77, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Cassir, N.; Simeoni, U.; La Scola, B. Gut microbiota and the pathogenesis of necrotizing enterocolitis in preterm neonates. Future Microbiol. 2016, 11, 273–292. [Google Scholar] [CrossRef] [PubMed]
- Be’er, A.; Ariel, G.; Kalisman, O.; Helman, Y.; Sirota-Madi, A.; Zhang, H.P.; Florin, E.-L.; Payne, S.M.; Ben-Jacob, E.; Swinney, H.L. Lethal protein produced in response to competition between sibling bacterial colonies. Proc. Natl. Acad. Sci. USA 2010, 107, 6258–6263. [Google Scholar] [CrossRef] [PubMed]
- Be’er, A.; Zhang, H.P.; Florin, E.-L.; Payne, S.M.; Ben-Jacob, E.; Swinney, H.L. Deadly competition between sibling bacterial colonies. Proc. Natl. Acad. Sci. USA 2009, 106, 428–433. [Google Scholar] [CrossRef] [PubMed]
- Lozupone, C.A.; Stombaugh, J.I.; Gordon, J.I.; Jansson, J.K.; Knight, R. Diversity, stability and resilience of the human gut microbiota. Nature 2012, 489, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Zhao, F. Microbial transmission, colonisation and succession: From pregnancy to infancy. Gut 2023, 72, 772–786. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Intolerant n = 14 | Tolerant n = 13 | p | |
|---|---|---|---|
| Prenatal characteristics | |||
| Antenatal corticosteroids, No. (%) | 11 (84.6) | 10 (76.9) | 0.500 |
| IUGR, No. (%) | 4 (28.6) | 3 (23.1) | 0.546 |
| Pregnancy-induced hypertension, No. (%) | 4 (28.6) | 4 (30.8) | 0.615 |
| Hypothyroidism, No. (%) | 4 (30.8) | 2 (15.4) | 0.322 |
| Twins, No. (%) | 8 (61.5) | 3 (23.1) | 0.055 |
| Gestational Diabetes, No. (%) | 1 (7.1) | 3 (23.1) | 0.269 |
| Mother’s age ≥ 35 years old, No. (%) | 7 (63.6) | 6 (46.2) | 0.329 |
| Cesarean section, No. (%) | 14 (100) | 12 (92.3) | 0.500 |
| Perinatal characteristics | |||
| Gestational age, weeks | 30.2 (28.9 to 31.4) | 29.2 (28.0 to 30.3) | 0.219 |
| Birth weight, g | 1281.7 (1139.7 to 1423.8) | 1280.2 (1044.2 to 1516.1) | 0.990 |
| 5-min Apgar score | 7.86 (7.36 to 8.36) | 8.46 (7.83 to 9.10) | 0.114 |
| pH at birth | 7.25 (7.21 to 7.28) | 7.24 (7.15 to 7.32) | 0.809 |
| Base excess on cord blood, mmol/L | −7.09 (−9.96 to −4.22) | −6.27 (−9.78 to −2.76) | 0.699 |
| CRIB II score | 4.75 (2.00 to 7.50) | 6.83 (5.23 to 8.43) | 0.125 |
| Breast milk, g/kg/1st week | 88.0 (27.4 to 148.7) | 84.7 (13.5 to 155.9) | 0.938 |
| Variables | ß | S.E. | Wald | p Value | Odds Ratio (OR) | 95 C.I. for OR | |
|---|---|---|---|---|---|---|---|
| Lower | Upper | ||||||
| VLBW | 0.348 | 1.217 | 0.082 | 0.775 | 1.416 | 0.130 | 15.392 |
| MEF | 0.128 | 1.103 | 0.014 | 0.907 | 1.137 | 0.131 | 9.879 |
| pH on cord blood ° | 0.515 | 1.310 | 0.154 | 0.694 | 1.673 | 0.128 | 21.79 |
| Pielou-evenness ± | 2.486 | 1.035 | 5.769 | 0.016 * | 12.018 | 1.580 | 91.40 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Chiara, M.; Lazzaro, A.; Scribano, D.; Trancassini, M.; Pietropaolo, V.; Sonnessa, M.; De Luca, C.; Prota, R.; Onestà, E.; Laccetta, G.; et al. Reduced Gut Bacterial Diversity in Early Life Predicts Feeding Intolerance in Preterm Neonates. Trop. Med. Infect. Dis. 2024, 9, 174. https://doi.org/10.3390/tropicalmed9080174
Di Chiara M, Lazzaro A, Scribano D, Trancassini M, Pietropaolo V, Sonnessa M, De Luca C, Prota R, Onestà E, Laccetta G, et al. Reduced Gut Bacterial Diversity in Early Life Predicts Feeding Intolerance in Preterm Neonates. Tropical Medicine and Infectious Disease. 2024; 9(8):174. https://doi.org/10.3390/tropicalmed9080174
Chicago/Turabian StyleDi Chiara, Maria, Alessandro Lazzaro, Daniela Scribano, Maria Trancassini, Valeria Pietropaolo, Michele Sonnessa, Chiara De Luca, Rita Prota, Elisa Onestà, Gianluigi Laccetta, and et al. 2024. "Reduced Gut Bacterial Diversity in Early Life Predicts Feeding Intolerance in Preterm Neonates" Tropical Medicine and Infectious Disease 9, no. 8: 174. https://doi.org/10.3390/tropicalmed9080174