
Nitrogen Metabolism in Non-Nodulated and Nodulated Soybean Plants Related to Ureide Synthesis



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
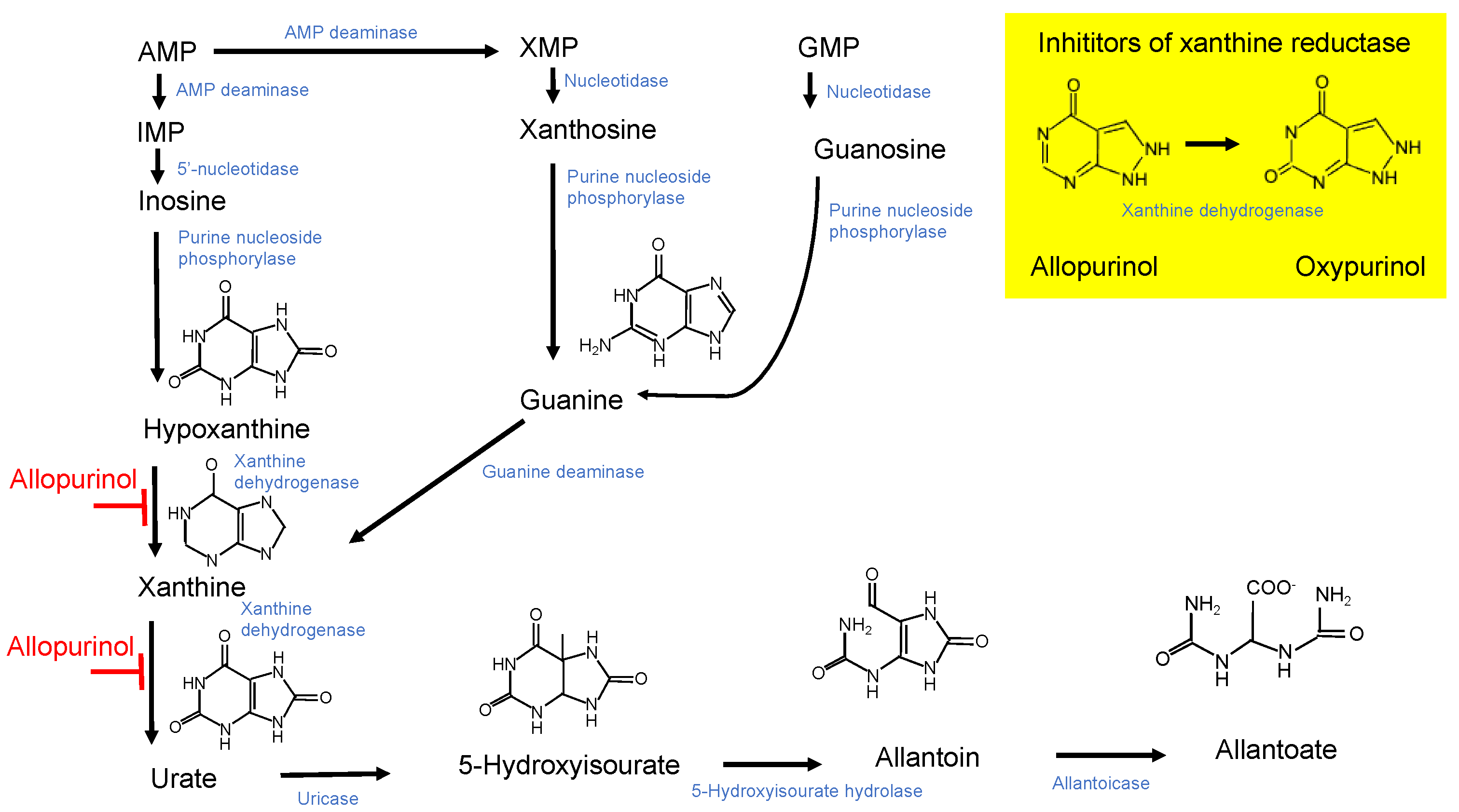
:1. Introduction
2. Materials and Methods
2.1. Plant Cultivation and Inoculation of Rhizobia
2.2. Effect of Urea Concentrations on N Metabolite Concentrations in Non-Nodulated Plants
2.3. Effect of Allopurinol Treatments on N Metabolite Concentrations in Non-Nodulated Plants
2.4. Effect of Allopurinol and Ureide Precursors on N Metabolite Concentrations in Nodulated Plants
2.5. Analysis of the Principal N Metabolites
2.6. Statistics
3. Results
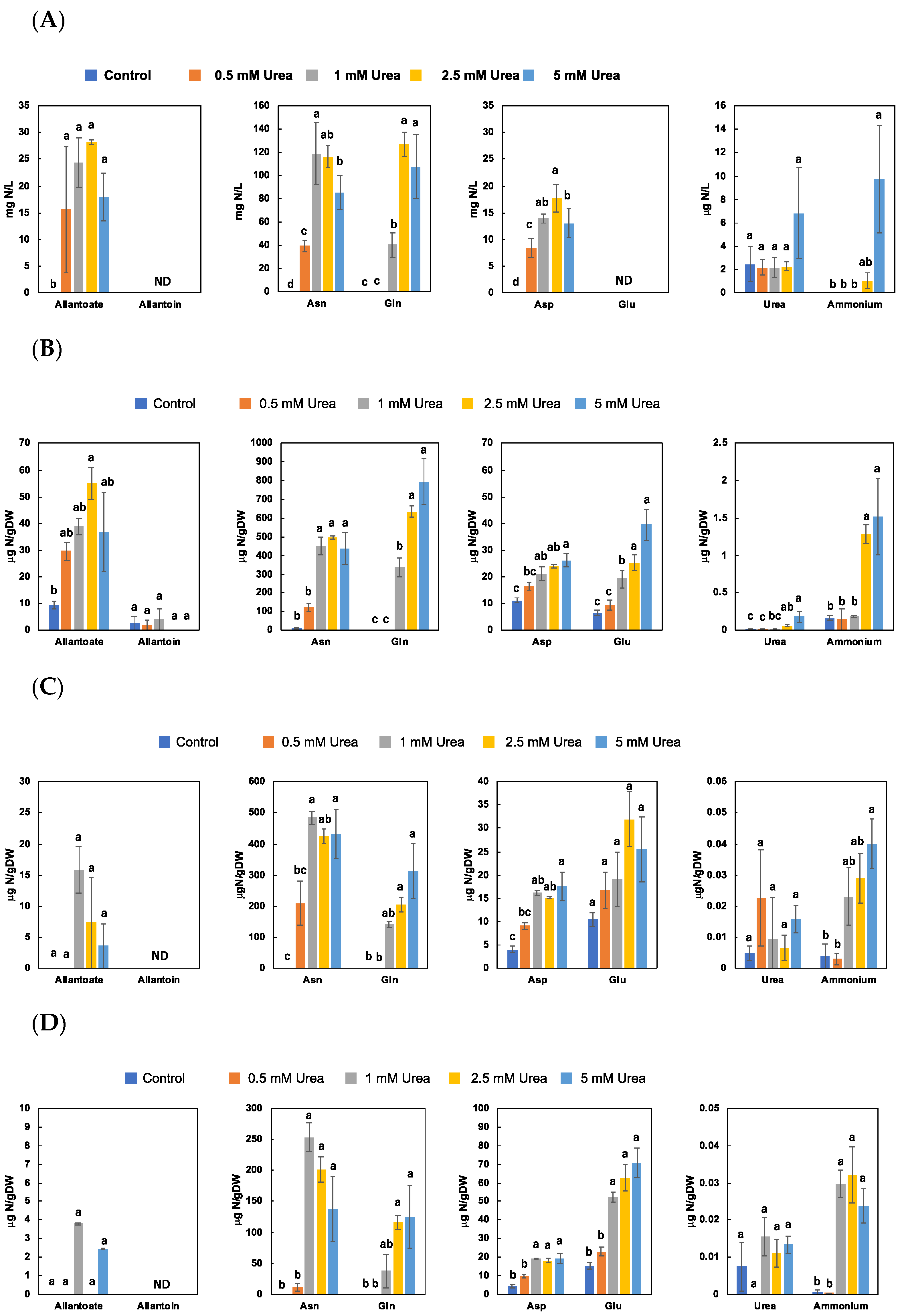
3.1. Effect of Urea Concentrations on N-Metabolite Concentrations in Non-Nodulated Soybean Plants (Experiment 1)
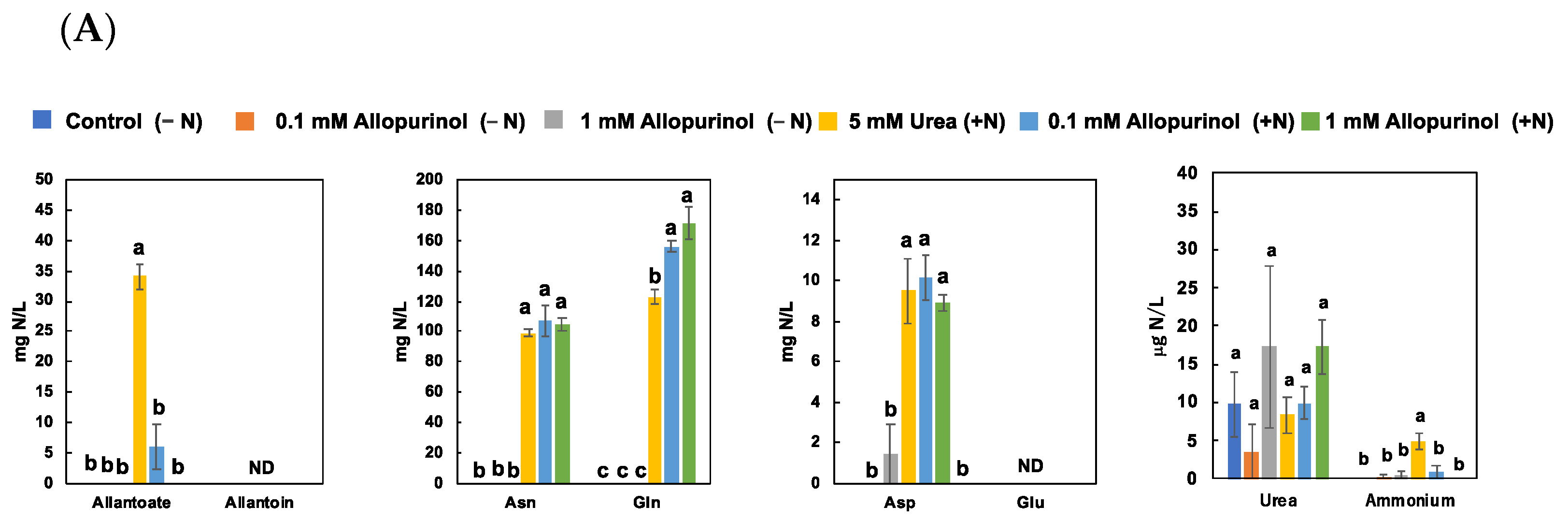
3.2. Effect of Allopurinol Treatments on N Metabolite Concentrations in Non-Nodulated Soybean Plants (Experiment 2)
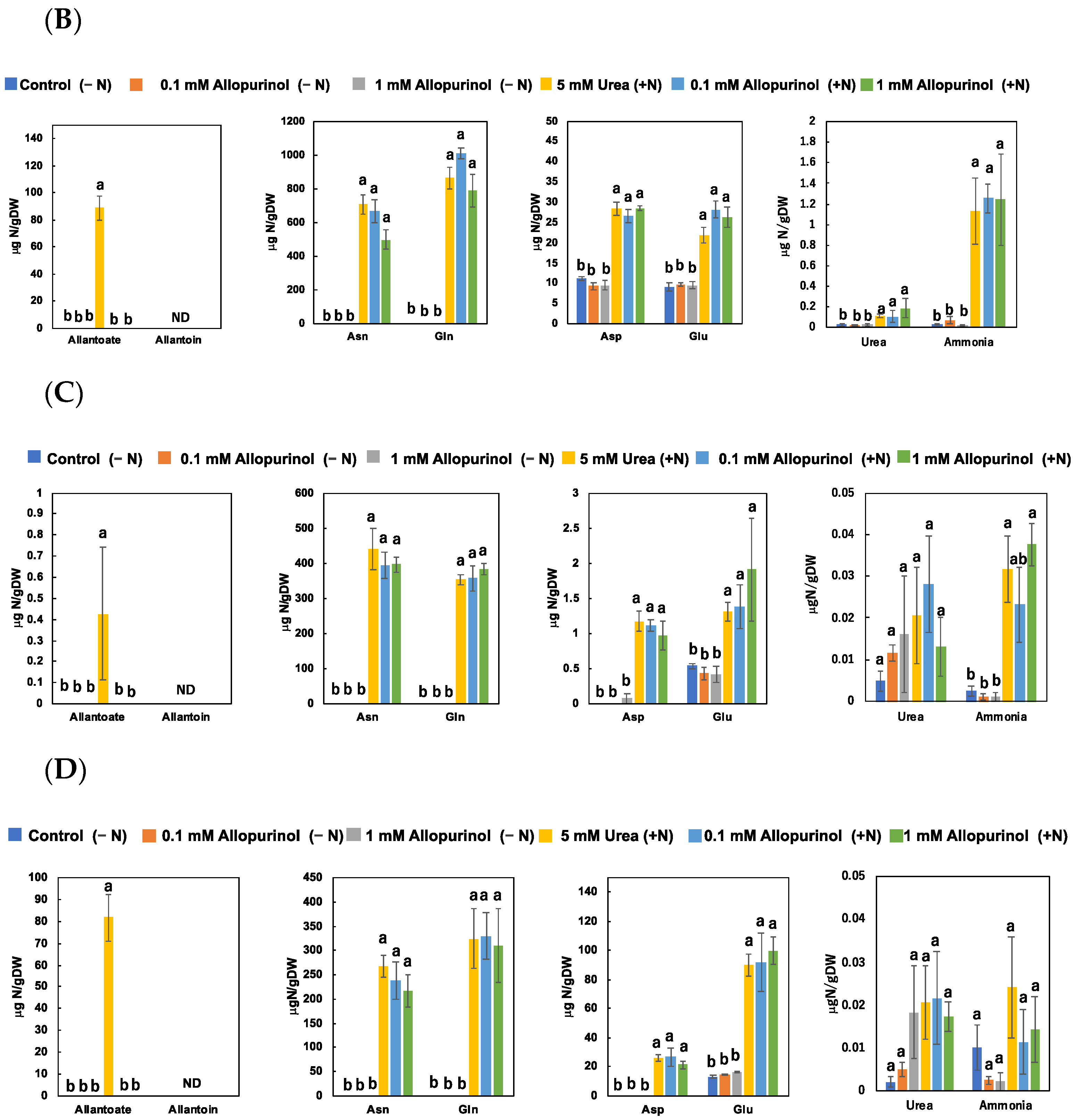
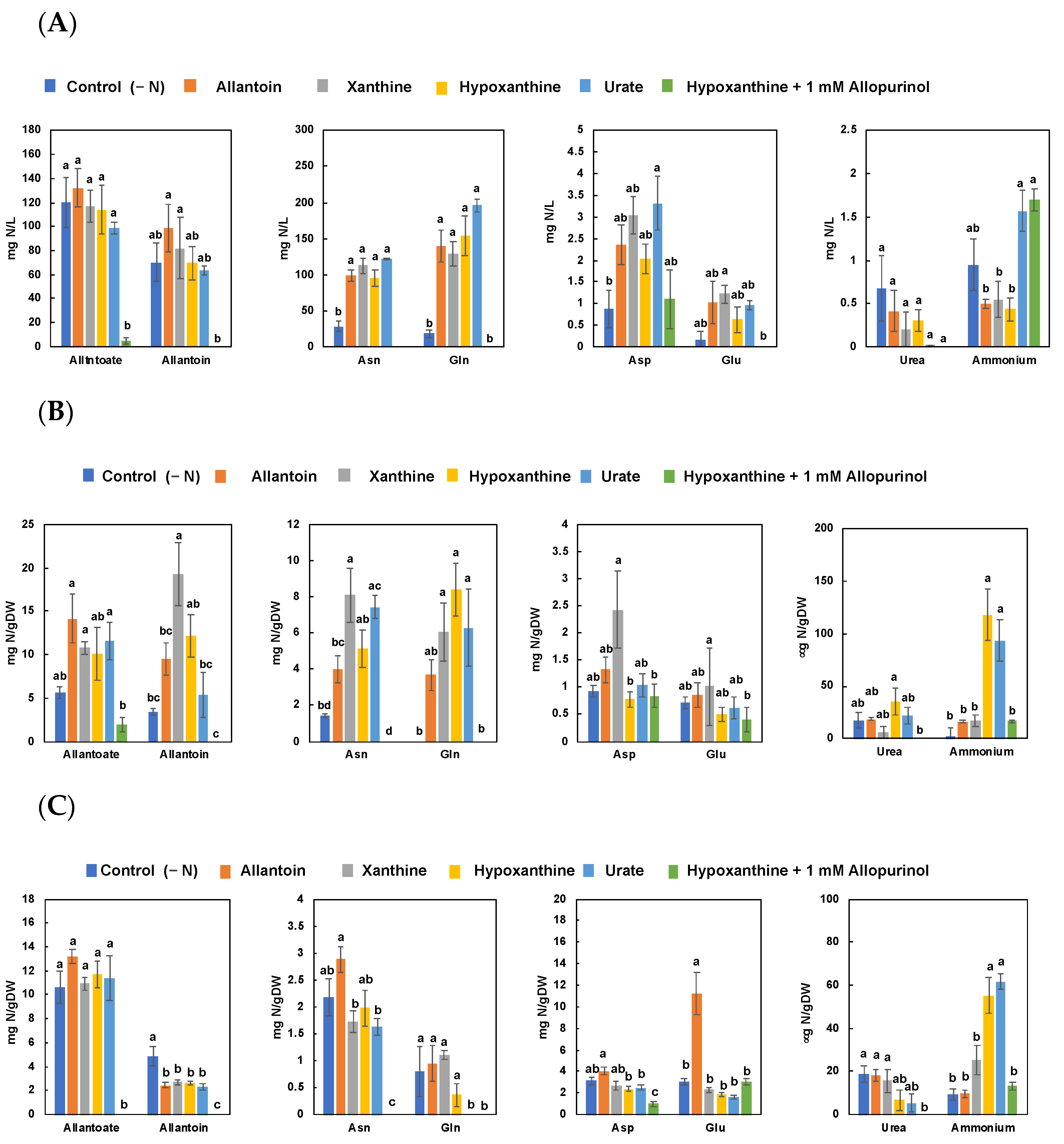
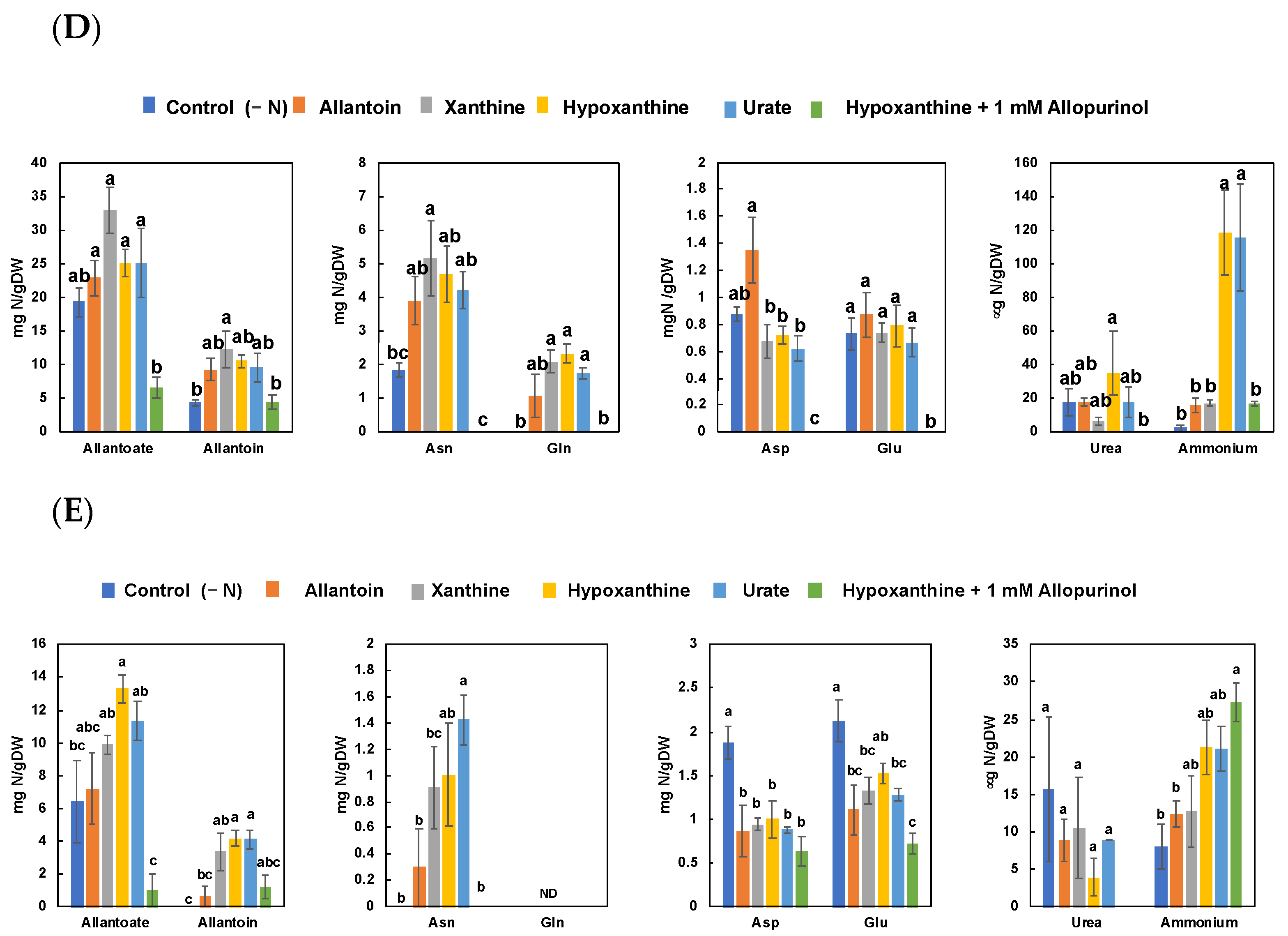
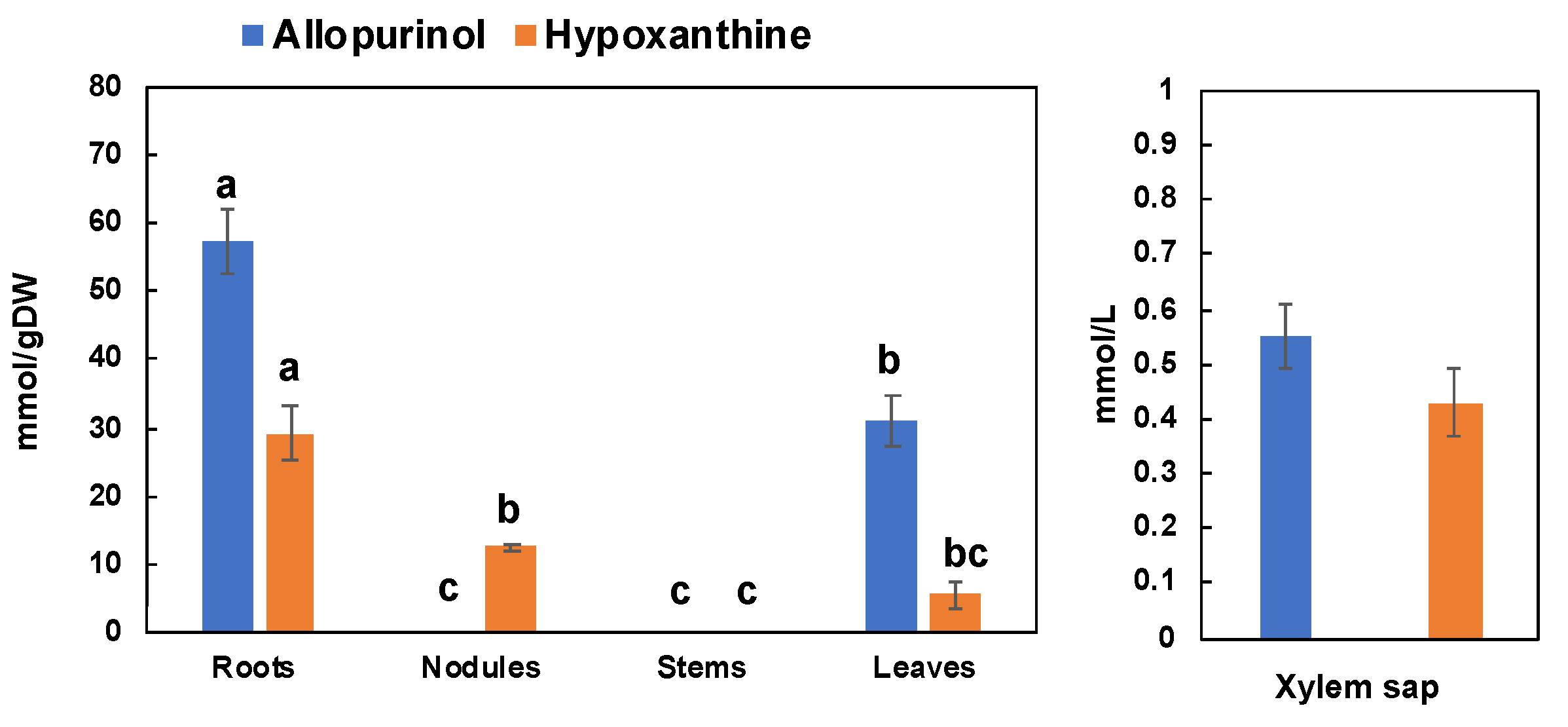
3.3. Effect of Allopurinol and Ureide Precursors on N Metabolite Concentrations in Nodulated Soybean Plants (Experiment 3)
4. Discussion
4.1. Effect of Urea Concentrations on N-Metabolite Concentrations in Non-Nodulated Soybean Plants (Experiment 1)
4.2. Effect of Allopurinol Treatments on N Metabolite Concentrations in Non-Nodulated Plants (Experiment 2)
4.3. Effect of Allopurinol and Ureide Precursors on N-Metabolite Concentrations in Nodulated Plants (Experiment 3)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ohyama, T.; Saito, K.; Kato, N. Assimilation and transport of nitrate, nitrite, and ammonia absorbed by nodulated soybean plants. Soil Sci. Plant Nutr. 1989, 35, 9–20. [Google Scholar] [CrossRef]
- Yamashita, N.; Tanabata, S.; Ohtake, N.; Sueyoshi, K.; Sato, T.; Higuchi, K.; Saito, A.; Ohyama, T. Effects of different chemical forms of nitrogen on the quick and reversible inhibition of soybean nodule growth and nitrogen fixation activity. Front. Plant Sci. 2019, 10, 131. [Google Scholar] [CrossRef] [PubMed]
- Ono, Y.; Fukasawa, M.; Sueyoshi, K.; Ohtake, N.; Sato, T.; Tanabata, S.; Toyota, R.; Higuchi, K.; Saito, A.; Ohyama, T. Application of nitrate, ammonium, or urea changes the concentrations of ureides, urea, amino acids and other metabolites in xylem sap and in the organs of soybean plants (Glycine max (L.) Merr.). Int. J. Mol. Sci. 2021, 22, 4573. [Google Scholar] [CrossRef] [PubMed]
- McClure, P.R.; Israel, D.W. Transport of nitrogen in the xylem of soybean plants. Plant Physiol. 1979, 64, 411–416. [Google Scholar] [CrossRef] [PubMed]
- Tajima, S.; Nomura, M.; Kouchi, H. Ureide Biosynthesis in legume nodules. Front. BioSci. 2004, 9, 1374–1381. [Google Scholar] [CrossRef] [PubMed]
- Ishizuka, J. Physiological roles of soluble nitrogenous compounds on vegetative growth and seed protein formation of soybean plants in Hokkaido. Res. Bull. Hokkaido Natl. Agric. Exp. Stn. 1970, 101, 51–121. [Google Scholar]
- Kushizaki, M.; Ishizuka, J.; Akamatsu, F. Physiological studies on the nutrition of soybean plants. 2. Effect of nodule formation on nitrogenous constituents of soybeans. J. Sci. Soil Manure Jpn. 1964, 35, 323–327. [Google Scholar]
- Matsumoto, T.; Yatazawa, M.; Yamamoto, Y. Incorporation of 15N into allantoin in nodulated soybean plants supplied with 15N2. Plant Cell Physiol. 1977, 18, 459–462. [Google Scholar] [CrossRef]
- Ohyama, T.; Kumazawa, K. Incorporation of 15N into various nitrogenous compounds in intact soybean nodules after exposure to 15N2 gas. Soil Sci. Plant Nutr. 1978, 24, 525–533. [Google Scholar] [CrossRef]
- Ohyama, T.; Kumazawa, K. Nitrogen assimilation in soybean nodules I. The role of GS/GOGAT system in the assimilation of ammonia produced by N2 fixation. Soil Sci. Plant Nutr. 1980, 26, 109–115. [Google Scholar] [CrossRef]
- Ohyama, T.; Kumazawa, K. Nitrogen assimilation in soybean nodules II. 15N2 assimilation in bacteroid and cytosol fractions of soybean nodules. Soil Sci. Plant Nutr. 1980, 26, 205–213. [Google Scholar] [CrossRef]
- Thomas, R.J.; Schrader, L.S. Ureide Metabolism in higher plants. Phytochemistry 1981, 20, 361–371. [Google Scholar] [CrossRef]
- Ohyama, T.; Kumazawa, K. Assimilation and transport of nitrogenous compounds originated from 15N2 fixation and 15NO3 absorption. Soil Sci. Plant Nutr. 1979, 25, 9–19. [Google Scholar] [CrossRef]
- Ohyama, T. Comparative studies on the distribution of nitrogen in soybean plants supplied with N2 and NO3− at the pod filling stage. II. Assimilation and transport of nitrogenous constituents. Soil Sci. Plant Nutr. 1983, 30, 219–229. [Google Scholar] [CrossRef]
- Ohyama, T.; Kato, N.; Saito, K. Nitrogen transport in xylem of soybean plant supplied with 15NO3−. Soil Sci. Plant Nutr. 1989, 35, 131–137. [Google Scholar] [CrossRef]
- McNeil, D.L.; LaRue, T.A. Effect of nitrogen source on ureides in soybean. Plant Physiol. 1984, 74, 227–232. [Google Scholar] [CrossRef]
- Todd, C.D.; Peter, A.; Tipton, P.A.; Blevins, D.G.; Piedras, P.; Pineda, M.; Polacco, J.C. Update on ureide degradation in legumes. J. Exp. Bot. 2006, 57, 5–12. [Google Scholar] [CrossRef]
- Werner, A.K.; Witte, C.P. The biochemistry of nitrogen mobilization: Purine ring catabolism. Trends Plant Sci. 2011, 16, 381–387. [Google Scholar] [CrossRef]
- Atkins, C.A.; Beevers, L. Synthesis, transport and utilization of translocated solutes of nitrogen. In Nitrogen in Higher Plants; Abrol, Y.P., Ed.; Research Studies Press: Somerset, UK, 1990; pp. 223–295. [Google Scholar]
- Atkins, C.A.; Smith, P.M. Translocation in legumes: Assimilates, nutrients, and signaling molecules. Plant Physiol. 2007, 144, 550–561. [Google Scholar] [CrossRef]
- Atkins, C.A.; Storer, P.J.; Pate, J.S. Pathways of nitrogen assimilation in cowpea nodules studied using 15N2 and allopurinol. Plant Physiol. 1988, 86, 204–207. [Google Scholar] [CrossRef]
- Pate, J.S.; Sharkey, P.J.; Lewis, O.A. Xylem to phloem transfer of solutes in fruiting shoots of legumes, studied by a phloem bleeding technique. Planta 1975, 122, 11–26. [Google Scholar] [CrossRef] [PubMed]
- Ohyama, T. Comparative studies on the distribution of nitrogen in soybean plants supplied with N2 and NO3− at the pod filling stage. Soil Sci. Plant Nutr. 1983, 29, 133–145. [Google Scholar] [CrossRef]
- Ohyama, T.; Kawai, S. Nitrogen assimilation and transport in soybean leaves: Investigation by petiole girdling treatment. Soil Sci. Plant Nutr. 1983, 29, 227–231. [Google Scholar] [CrossRef]
- Winkler, R.; Blevins, D.; Polacco, J.C.; Randall, D. Ureide catabolism of soybeans: II. Pathway of catabolism in intact leaf tissue. Plant Physiol. 1987, 83, 585–591. [Google Scholar] [CrossRef]
- Herridge, D.F.; Atkins, C.A.; Pate, J.S.; Rainbird, R.M. Allantoin and allantoic acid in the nitrogen economy of the cowpea (Vigna unguiculata [L.] Walp.). Plant Physiol. 1978, 62, 495–498. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, V.R.; Wittenbach, V.A.; Giaquinta, R.T. Paraveinal mesophyll of soybean leaves in relation to assimilate transfer and compartmentation: III. Immunohistochemical localization of specific glycopeptides in the vacuole after depodding. Plant Physiol. 1983, 72, 586–589. [Google Scholar] [CrossRef] [PubMed]
- Pélissier, H.C.; Frerich, A.; Desimone, M.; Schumacher, K.; Tegeder, M. PvUPS1, an allantoin transporter in nodulated roots of French bean. Plant Physiol. 2004, 134, 664–675. [Google Scholar] [CrossRef] [PubMed]
- Collier, R.; Tegeder, M. Soybean ureide transporters play a critical role in nodule development, function and nitrogen export. Plant J. 2012, 72, 355–367. [Google Scholar] [CrossRef]
- Reinbothe, H.; Mothes, K. Urea, ureides, and guanidines in plants. Annu. Rev. Plant Physiol. 1962, 13, 129–149. [Google Scholar] [CrossRef]
- Bollard, E.G. Nitrogenous compounds in plant xylem sap. Nature 1956, 178, 1189–1190. [Google Scholar] [CrossRef]
- Ashihara, H.; Ludwig, I.A. Degradation of purine nucleotides. In Plant Nucletide Metabolism, Biosynthesis, Degradation, and Alkaloid Formation, 1st ed.; Ashihara, H., Ludwig, I.A., Crozier, A., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2020. [Google Scholar]
- Schubert, K.R.; Boland, M.J. The ureides. In The Biochemistry of Plants. A Comprehensive Treatise, Vol 16: Intermediary Nitrogen Metabolism; Miflin, B.J., Lea, P.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 197–282. [Google Scholar]
- Krupka, R.M.; Towers, G.H.N. Studies of the metabolic relations of allantoin in wheat. Can. J. Bot. 1959, 37, 539–545. [Google Scholar] [CrossRef]
- Ohyama, T.; Kumazawa, K. Nitrogen assimilation in soybean nodules V. Possible pathway of allantoin synthesis in soybean noudles. Soil Sci. Plant Nutr. 1981, 27, 111–114. [Google Scholar] [CrossRef]
- Mothes, K. The metabolism of urea and ureides. Can. J. Bot. 1961, 39, 1785–1807. [Google Scholar] [CrossRef]
- Friberg, S.R.; Bollard, E.G.; Megarty, M.P. The natural occurrence of urea and ureides in the soluble nitrogen of the banana plant. In Plant Physiology, Proceedings of the Plant Physiology Meetings, Stanford University, CA, USA, 25–29 August 1957; American Society of Plant Biologist, Oxford Academic: Oxford, England, 1957; Volume 52, p. lii. [Google Scholar]
- Brunel, A.; Bruneo-Capelle, G. Synthese de lacide allantoique chez les Champignons Basidiomycetes. CR Acad. Sci. 1951, 232, 1130–1132. [Google Scholar]
- Bortolotti, M.; Polito, L.; Battelli, M.G.; Bolognesi, A. Xanthine oxidoreductase: One enzyme for multiple physiological tasks. Redox Biol. 2021, 41, 101882. [Google Scholar] [CrossRef]
- Enroth, C.; Bryan, T.; Eger, B.T.; Okamoto, K.; Nishinoi, T.; Nishino, T.; Pai, F. Crystal structures of bovine milk xanthine dehydrogenase and xanthine oxidase: Structure-based mechanism of conversion. Proc. Natl. Acad. Sci. USA 2000, 97, 10723–10728. [Google Scholar] [CrossRef]
- Triplett, E.W.; Blevins, D.G.; Randall, D.D. Purification and properties of soybean nodule xanthine dehydrogenase. Arch. Biochem. Biophys. 1982, 219, 39–46. [Google Scholar] [CrossRef]
- Hafez, R.M.; Abdel-Rahman, T.M.; Naguib, R.M. Uric acid in plants and microorganisms: Biological applications and genetics—A review. J. Adv. Res. 2017, 8, 475–486. [Google Scholar] [CrossRef]
- Walter-Sack, I.; de Vries, J.X.; Kreinerl, C.; Ittensohn, A.; Stenzhorn, G.; Voss, A.; Weber, E. Bioequivalence of allopurinol preparations: To be assessed by the parent drug or the active metabolite? Clin. Investig. 1993, 71, 240–246. [Google Scholar] [CrossRef]
- Nishino, T.; Okamoto, K. Mechanistic insights into xanthine oxidoreductase from development studies of candidate drugs to treat hyperuricemia and gout. J. Biol. Inorg. Chem. 2015, 20, 195–207. [Google Scholar] [CrossRef]
- Fujihara, S.; Yamaguchi, M. Effect of allopurinol [4-hydroxypyrazolo(3,4-d)pyrimidine] on the metabolism of allantoin in soybean plants. Plant Physiol. 1978, 62, 134–138. [Google Scholar] [CrossRef] [PubMed]
- Ohyama, T.; Isaka, M.; Saito, A.; Higuchi, K. Effects of nodulation on metabolite concentrations in xylem sap and in the organs of soybean plants supplied with different N forms. Metabolites 2023, 13, 319. [Google Scholar] [CrossRef] [PubMed]
- Ohyama, T.; Takayama, K.; Akagi, A.; Saito, A.; Higuchi, K.; Sato, T. Development of an N-free culture solution for cultivation of nodulated soybean with less pH fluctuation by the addition of potassium bicarbonate. Agriculture 2023, 13, 739. [Google Scholar] [CrossRef]
- Sakazume, T.; Tanaka, K.; Aida, H.; Ishikawa, S.; Nagumo, Y.; Takahashi, Y.; Ohtake, N.; Sueyoshi, K.; Ohyama, T. Estimation of nitrogen fixation rate of soybean (Glycine max (L.) Merr,) by micro-scale relative ureide analysis using root bleeding xylem sap and apoplast fluid in stem. Bull. Facul. Agric. Niigata Univ. 2014, 67, 27–41. [Google Scholar]
- Doi, M.; Higuchi, K.; Saito, A.; Sato, T.; Ohyama, T. N absorption, transport, and recycling in nodulated soybean plants by split-root experiment using 15N-labeled nitrate. Nitrogen 2022, 3, 636–651. [Google Scholar] [CrossRef]
- Ohyama, T.; Ikebe, K.; Okuoka, S.; Ozawa, T.; Nishiura, T.; Ishiwata, T.; Yamazaki, A.; Tanaka, F.; Takahashi, T.; Umezawa, T.; et al. A deep placement of lime nitrogen reduces the nitrate leaching and promotes soybean growth and seed yield. Crop Environ. 2022, 1, 221–230. [Google Scholar] [CrossRef]
- MEdical and PHarmaceutical Statistics (MEPHAS). Available online: www.gen-info.osaka-u.ac.jp/MEPHAS/mokuji1-e.html (accessed on 24 May 2002).
- Baslam, M.; Mitsui, T.; Sueyoshi, K.; Ohyama, T. Recent advances in carbon and nitrogen metabolism in C3 plants. Int. J. Mol. Sci. 2021, 22, 318. [Google Scholar] [CrossRef]
- Cheng, X.-G.; Nomura, M.; Sato, T.; Fujikake, H.; Ohyama, T.; Tajima, S. Effect of exogenous NH4+-N supply on distribution of ureide content in various tissues of alfalfa plants, Medicago sativa. Soil Sci. Plant Nutr. 1999, 45, 921–927. [Google Scholar] [CrossRef]
- Matsumoto, T.; Yatazawa, M.; Yamamoto, Y. Effects of exogenous nitrogen compounds on the concentrations of allantoin and various constituents in several organs of soybean plants. Plant Cell Physiol. 1977, 18, 613–624. [Google Scholar] [CrossRef]
- Lee, D.K.; Redillas, M.C.F.R.; Jung, H.; Choi, S.; Kim, Y.S.; Kim, J.K. A nitrogen molecular sensing system, comprised of the ALLANTOINASE and UREIDE PERMEASE 1 genes, can be used to monitor N status in rice. Front. Plant Sci. 2018, 9, 444. [Google Scholar] [CrossRef]
- Redillas, M.C.F.R.; Bang, S.W.; Lee, D.K.; Kim, Y.S.; Jung, H.; Chung, P.J. Allantoin accumulation through overexpression of ureide permease1 improves rice growth under limited nitrogen conditions. Plant Biotechnol. J. 2019, 17, 1289–1301. [Google Scholar] [CrossRef] [PubMed]
- Takagi, H.; Watanabe, S.; Tanaka, S.; Matsuura, T.; Mori, I.C.; Hirayama, T.; Shimada, H.; Sakamoto, A. Disruption of ureide degradation affects plant growth and development during and after transition from vegetative to reproductive stages. BMC Plant Biol. 2018, 18, 287. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ohyama, T.; Matsumoto, K.; Goto, H.; Saito, A.; Higuchi, K. Nitrogen Metabolism in Non-Nodulated and Nodulated Soybean Plants Related to Ureide Synthesis. Nitrogen 2023, 4, 209-222. https://doi.org/10.3390/nitrogen4020014
Ohyama T, Matsumoto K, Goto H, Saito A, Higuchi K. Nitrogen Metabolism in Non-Nodulated and Nodulated Soybean Plants Related to Ureide Synthesis. Nitrogen. 2023; 4(2):209-222. https://doi.org/10.3390/nitrogen4020014
Chicago/Turabian StyleOhyama, Takuji, Kahori Matsumoto, Haruka Goto, Akihiro Saito, and Kyoko Higuchi. 2023. "Nitrogen Metabolism in Non-Nodulated and Nodulated Soybean Plants Related to Ureide Synthesis" Nitrogen 4, no. 2: 209-222. https://doi.org/10.3390/nitrogen4020014
APA StyleOhyama, T., Matsumoto, K., Goto, H., Saito, A., & Higuchi, K. (2023). Nitrogen Metabolism in Non-Nodulated and Nodulated Soybean Plants Related to Ureide Synthesis. Nitrogen, 4(2), 209-222. https://doi.org/10.3390/nitrogen4020014