Gut Microbiota Is Linked to Physical Health Improvements Resulting from Energy-Restricted Diet and Exercise: A Randomized Controlled Trial in Healthy Adults †


Abstract
:1. Introduction
2. Material and Methods
2.1. Brief Study Design
2.2. Participants
2.3. Body Composition, Diet and Blood Biomarker Assessment Measures
2.4. Fecal Sample Collection and Preparation
2.5. Microbiota Analysis Using 16s rRNA High-Throughput Sequencing
2.6. Statistical Analysis
3. Results and Discussion
3.1. Baseline Characteristics
3.2. Changes in Body Composition, Strength, Blood Biomarkers and Dietary Intake
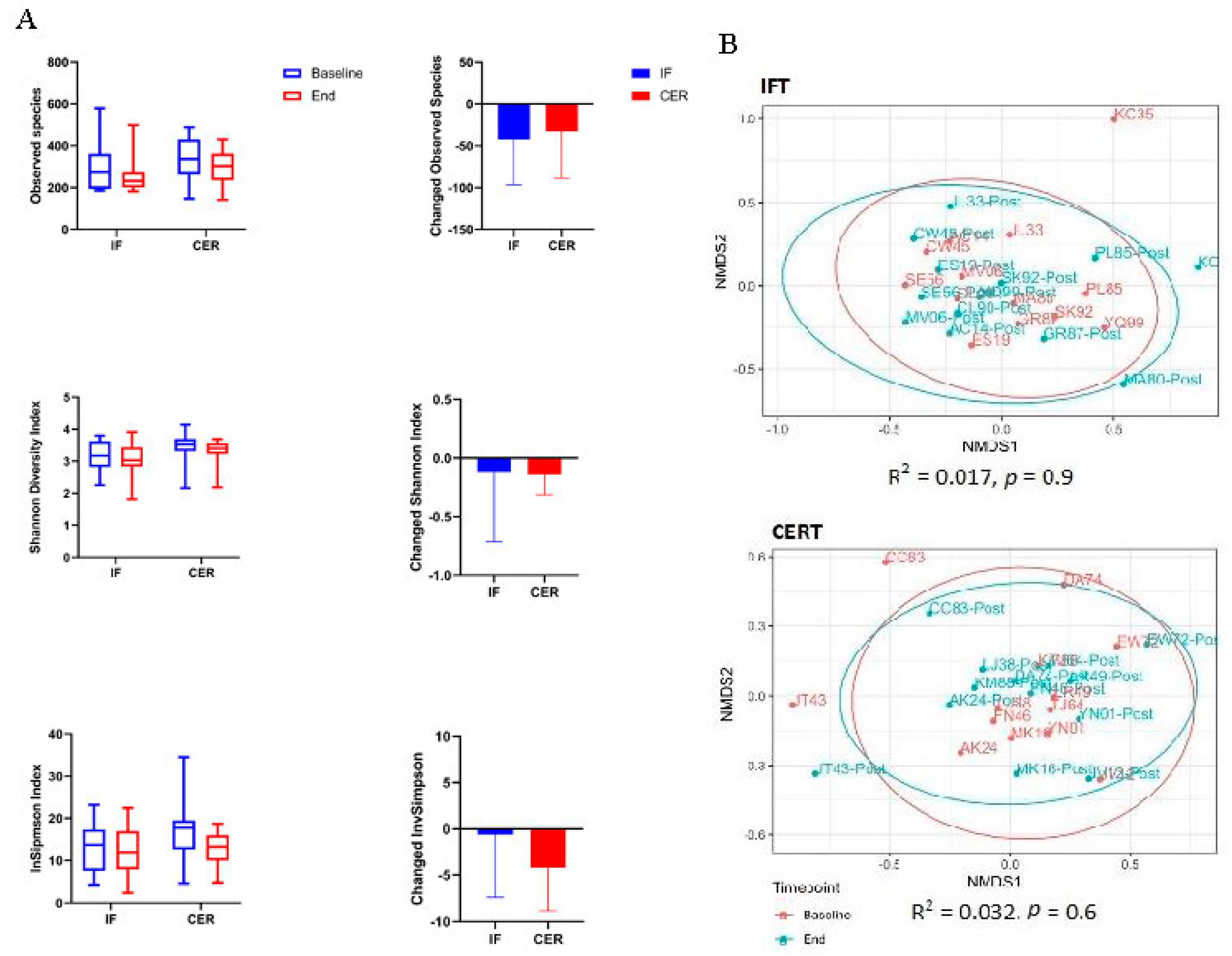
3.3. Effect of Caloric Restriction and Exercise on Fecal Bacterial Diversity
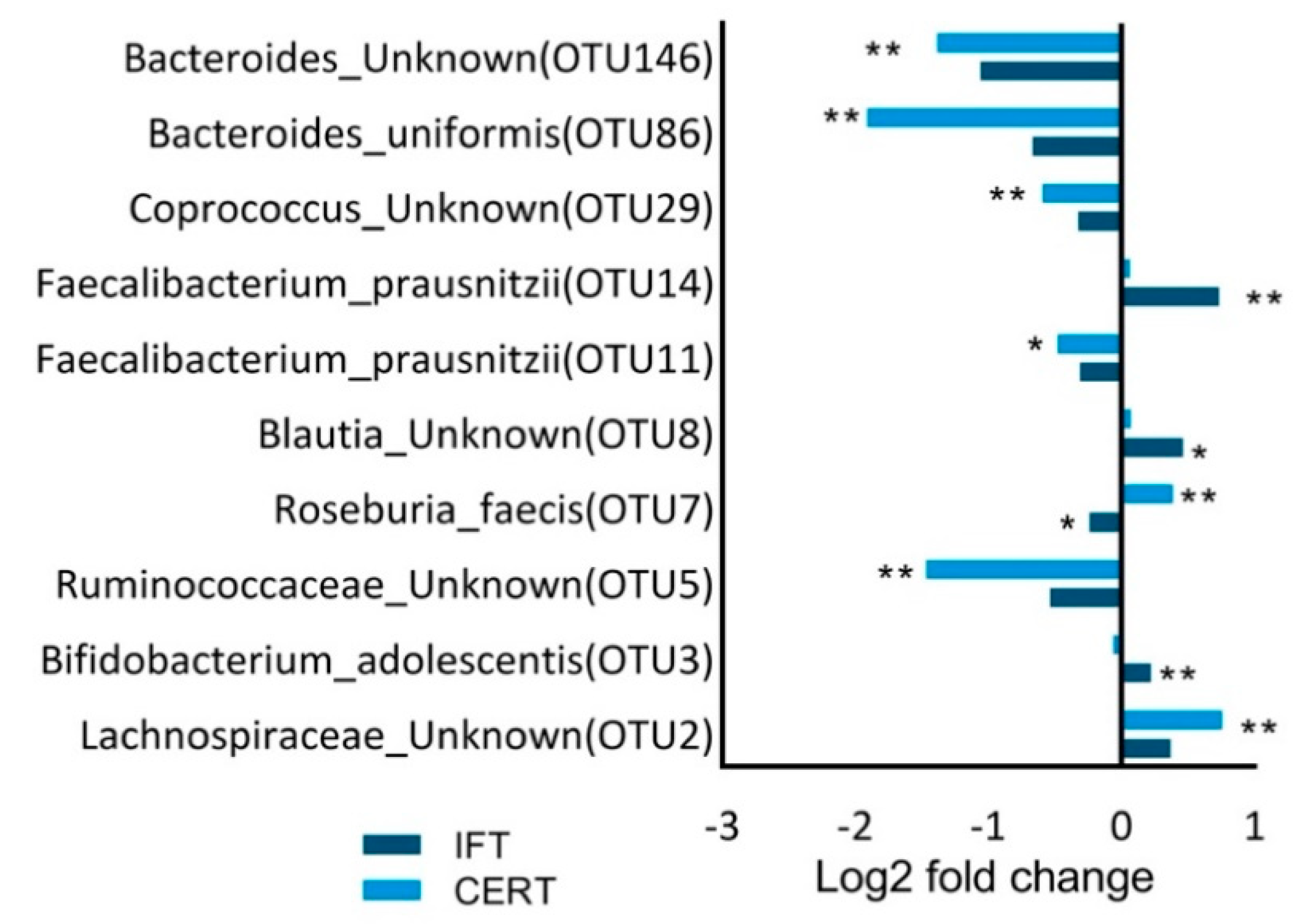
3.4. Taxonomic Composition of the Fecal Microbiota
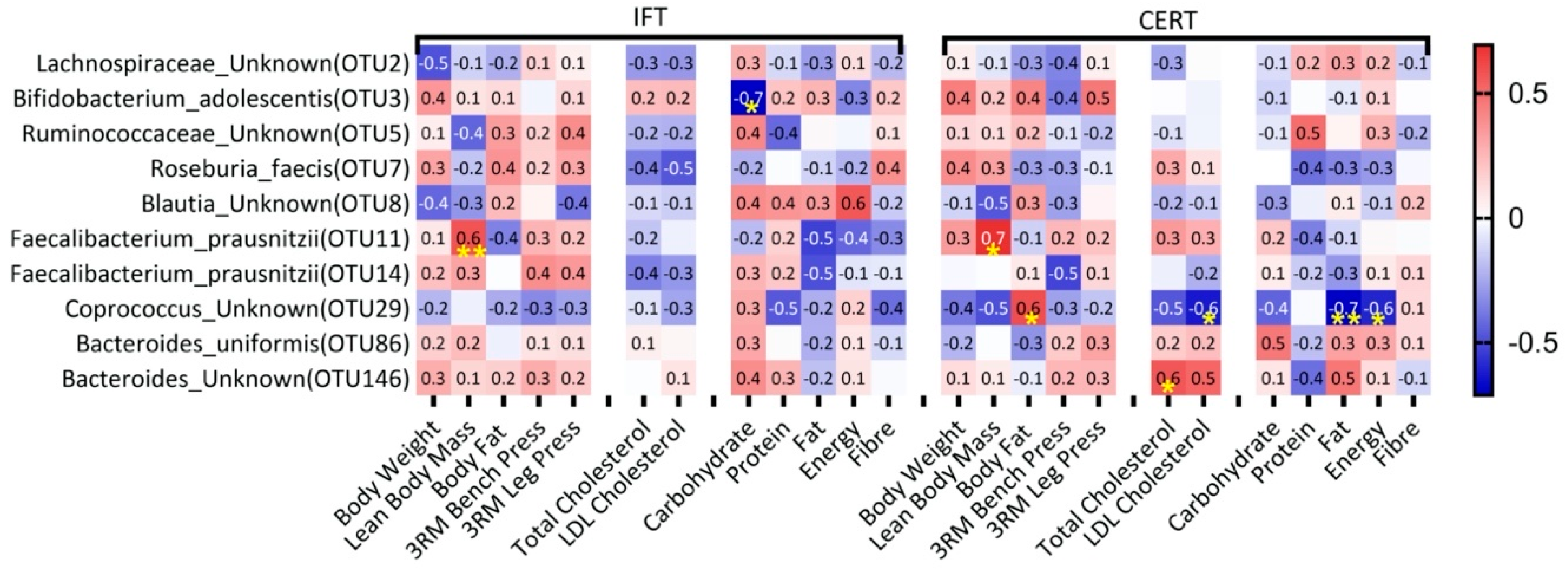
3.5. Correlates between Changes in Diet, Blood Cholesterol and Physical Measures with Changes in Bacterial Abundance
4. Conclusions
Funding
References
- Li, L.; Su, Y.; Li, F.; Wang, Y.; Ma, Z.; Li, Z.; Su, J. The effects of daily fasting hours on shaping gut microbiota in mice. BMC Microbiol. 2020, 20, 65. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Sparks, J.B.; Karyala, S.V.; Settlage, R.; Luo, X.M. Host adaptive immunity alters gut microbiota. ISME J. 2015, 9, 770–781. [Google Scholar] [CrossRef] [PubMed]
- Anhê, F.F.; Roy, D.; Pilon, G.; Dudonné, S.; Matamoros, S.; Varin, T.V.; Garofalo, C.; Moine, Q.; Desjardins, Y.; Levy, E.; et al. A polyphenol-rich cranberry extract protects from diet-induced obesity, insulin resistance and intestinal inflammation in association with increased Akkermansia spp. population in the gut microbiota of mice. Gut 2015, 64, 872–883. [Google Scholar] [CrossRef] [PubMed]
- Wiley, N.C.; Dinan, T.G.; Ross, R.P.; Stanton, C.; Clarke, G.; Cryan, J.F. The microbiota-gut-brain axis as a key regulator of neural function and the stress response: Implications for human and animal health. J. Anim. Sci. 2017, 95, 3225–3246. [Google Scholar] [CrossRef] [PubMed]
- Hasan, N.; Yang, H. Factors affecting the composition of the gut microbiota, and its modulation. PeerJ 2019, 7, e7502. [Google Scholar] [CrossRef] [PubMed]
- Mohr, A.E.; Jäger, R.; Carpenter, K.C.; Kerksick, C.M.; Purpura, M.; Townsend, J.R.; West, N.P.; Black, K.; Gleeson, M.; Pyne, D.B.; et al. The athletic gut microbiota. J. Int. Soc. Sports Nutr. 2020, 17, 24. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.M.; Berg Miller, M.E.; Pence, B.D.; Whitlock, K.; Nehra, V.; Gaskins, H.R.; White, B.A.; Fryer, J.D.; Woods, J.A. Voluntary and forced exercise differentially alters the gut microbiome in C57BL/6J mice. J. Appl. Physiol. (1985) 2015, 118, 1059–1066. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.M.; Morris, L.S.; Marchesi, J.R. The gut microbiome: The role of a virtual organ in the endocrinology of the host. J. Endocrinol. 2013, 218, R37–R47. [Google Scholar] [CrossRef] [PubMed]
- Dalton, A.; Mermier, C.; Zuhl, M. Exercise influence on the microbiome-gut-brain axis. Gut Microbes 2019, 10, 555–568. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, C.M.; Davy, B.M.; Hulver, M.W.; Neilson, A.P.; Bennett, B.J.; Davy, K.P. Does Exercise Alter Gut Microbial Composition? A Systematic Review. Med. Sci. Sports. Exerc. 2019, 51, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Beli, E.; Yan, Y.; Moldovan, L.; Vieira, C.P.; Gao, R.; Duan, Y.; Prasad, R.; Bhatwadekar, A.; White, F.A.; Townsend, S.D.; et al. Restructuring of the Gut Microbiome by Intermittent Fasting Prevents Retinopathy and Prolongs Survival in. Diabetes 2018, 67, 1867–1879. [Google Scholar] [CrossRef] [PubMed]
- Rinninella, E.; Cintoni, M.; Raoul, P.; Ianiro, G.; Laterza, L.; Lopetuso, L.R.; Ponziani, F.R.; Gasbarrini, A.; Mele, M.C. Gut Microbiota during Dietary Restrictions: New Insights in Non-Communicable Diseases. Microorganisms 2020, 8, 1140. [Google Scholar] [CrossRef] [PubMed]
- McMurdie, P.J.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- He, Y.; Yin, J.; Lei, J.; Liu, F.; Zheng, H.; Wang, S.; Wu, S.; Sheng, H.; McGovern, E.; Zhou, H. Fasting challenges human gut microbiome resilience and reduces Fusobacterium. Med. Microecol. 2019, 1, 100003. [Google Scholar] [CrossRef]
- Ott, B.; Skurk, T.; Hastreiter, L.; Lagkouvardos, I.; Fischer, S.; Büttner, J.; Kellerer, T.; Clavel, T.; Rychlik, M.; Haller, D. Effect of caloric restriction on gut permeability, inflammation markers, and fecal microbiota in obese women. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zou, H.; Wang, D.; Ren, H.; Cai, K.; Chen, P.; Fang, C.; Shi, Z.; Zhang, P.; Wang, J.; Yang, H. Effect of Caloric Restriction on BMI, Gut Microbiota, and Blood Amino Acid Levels in Non-Obese Adults. Nutrients 2020, 12, 631. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Alvarez, L.; Xu, H.; Martinez-Tellez, B. Influence of Exercise on the Human Gut Microbiota of Healthy Adults: A Systematic Review. Clin. Transl. Gastroenterol. 2020, 11, e00126. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Xie, C.; Lu, S.; Nichols, R.G.; Tian, Y.; Li, L.; Patel, D.; Ma, Y.; Brocker, C.N.; Yan, T. Intermittent fasting promotes white adipose browning and decreases obesity by shaping the gut microbiota. Cell Metab. 2017, 26, 672–685.e4. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Baseline Variable | IFT (n = 12) Mean ± SD | CERT(n = 13) Mean ± SD | IFT vs. CERT (p) |
|---|---|---|---|
| Male/Female | 6/6 | 5/8 | |
| Age (years) | 25.8 ± 5.1 | 23.6 ± 4.1 | 0.24 |
| Weight | 80.2 ± 11.4 | 78.2 ± 13.8 | 0.70 |
| BMI (kg/m2) | 27.3 ± 2.7 | 27.2 ± 3.2 | 0.97 |
| LBM (kg) | 54.0 ± 12.1 | 51.4 ± 12.0 | 0.59 |
| Body fat (kg) | 30.4 ± 7.9 | 30.6 ± 8.3 | 0.96 |
| Bench Press (3RM) | 42.5 ± 19.2 | 37.6 ± 20.2 | 0.54 |
| Leg Press (3RM) | 111.7 ± 57.1 | 96.7 ± 64.4 | 0.55 |
| TC * | 4.7 ± 1.0 | 4.2 ± 0.6 | 0.19 |
| LDL-C * | 2.7 ± 0.8 | 2.3 ± 0.5 | 0.09 |
| HDL-C * | 1.5 ± 0.5 | 1.5 ± 0.4 | 0.81 |
| Variable | Group | Baseline Mean ± SD | Post-Intervention Mean ± SD | p (Group) | p (Time) | p (I) |
|---|---|---|---|---|---|---|
| Weight | IFT | 80.2 ± 11.4 | 76.0 ± 10.3 | 0.54 | <0.001 | 0.73 |
| CERT | 78.2 ± 13.8 | 74.5 ± 12.0 | ||||
| BMI (kg/m2) | IFT | 27.3 ± 2.7 | 25.9 ± 2.7 | 0.97 | <0.001 | 0.73 |
| CERT | 27.2 ± 3.2 | 26.0 ± 3.0 | ||||
| LBM (kg) | IFT | 54.0 ± 12.1 | 55.8 ± 11.4 | 0.005 | <0.001 | 0.97 |
| CERT | 51.4 ± 12.0 | 53.2 ± 11.4 | ||||
| Body fat (kg) | IFT | 30.4 ± 7.9 | 22.7 ± 5.9 | 0.07 | <0.001 | 0.73 |
| CERT | 30.6 ± 8.3 | 23.4 ± 8.0 | ||||
| Bench Press (3RM) | IFT | 42.5 ± 19.2 | 46.3 ± 17.8 | 0.001 | <0.001 | 0.17 |
| CERT | 37.6 ± 20.2 | 43.5 ± 19.3 | ||||
| Leg Press (3RM) | IFT | 111.7 ± 57.1 | 140.3 ± 60.0 | 0.001 | <0.001 | 0.96 |
| CERT | 96.7 ± 64.4 | 125.8 ± 58.1 | ||||
| TC * | IFT | 4.7 ± 1.0 | 4.1 ± 0.7 | 0.30 | 0.001 | 0.19 |
| CERT | 4.2 ± 0.6 | 3.9 ± 0.8 | ||||
| LDL-C * | IFT | 2.7 ± 0.8 | 2.2 ± 0.5 | 0.15 | 0.001 | 0.15 |
| CERT | 2.3 ± 0.5 | 2.1 ± 0.5 | ||||
| HDL-C * | IFT | 1.5 ± 0.5 | 1.4 ± 0.5 | 0.37 | 0.12 | 0.41 |
| CERT | 1.5 ± 0.4 | 1.4 ± 0.4 | ||||
| Dietary Variable | Baseline Mean ± SD | During-Intervention Mean ± SD | p (Group) | p (Time) | p (I) | |
| Energy (kJ) | IFT | 7344 ± 1453 | 6581 ± 1336 | 0.20 | 0.07 | 0.99 |
| CERT | 7511 ± 1763 | 6742 ± 1411 | ||||
| Protein (grams/day) | IFT | 89.7 ± 31.0 | 111.9 ± 24.0 | 0.05 | <0.001 | 0.64 |
| CERT | 86.4 ± 22.5 | 114.2 ± 26.3 | ||||
| Carbohydrates (grams/day) | IFT | 179.6 ± 52.6 | 148.6 ± 35.9 | 0.04 | 0.007 | 0.83 |
| CERT | 182.8 ± 40.5 | 155.9 ± 34.4 | ||||
| Fat (grams/day) | IFT | 69.6 ± 16.7 | 56.1 ± 11.0 | 0.43 | 0.004 | 0.89 |
| CERT | 74.6 ± 26.0 | 58.1 ± 11.9 | ||||
| Fiber (grams/day) | IFT | 18.3 ± 7.1 | 23.8 ± 6.1 | 0.30 | 0.04 | 0.27 |
| CERT | 18.2 ± 6.4 | 19.9 ± 6.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gondalia, S.; Cooke, M.; Keenan, S.; Belski, R. Gut Microbiota Is Linked to Physical Health Improvements Resulting from Energy-Restricted Diet and Exercise: A Randomized Controlled Trial in Healthy Adults. Proceedings 2020, 61, 8. https://doi.org/10.3390/IECN2020-06992
Gondalia S, Cooke M, Keenan S, Belski R. Gut Microbiota Is Linked to Physical Health Improvements Resulting from Energy-Restricted Diet and Exercise: A Randomized Controlled Trial in Healthy Adults. Proceedings. 2020; 61(1):8. https://doi.org/10.3390/IECN2020-06992
Chicago/Turabian StyleGondalia, Shakuntla, Matthew Cooke, Stephen Keenan, and Regina Belski. 2020. "Gut Microbiota Is Linked to Physical Health Improvements Resulting from Energy-Restricted Diet and Exercise: A Randomized Controlled Trial in Healthy Adults" Proceedings 61, no. 1: 8. https://doi.org/10.3390/IECN2020-06992
APA StyleGondalia, S., Cooke, M., Keenan, S., & Belski, R. (2020). Gut Microbiota Is Linked to Physical Health Improvements Resulting from Energy-Restricted Diet and Exercise: A Randomized Controlled Trial in Healthy Adults. Proceedings, 61(1), 8. https://doi.org/10.3390/IECN2020-06992