
Turtle Origins: Chinlechelys tenertesta and Convergence in Modern Cladistic Analysis †



{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
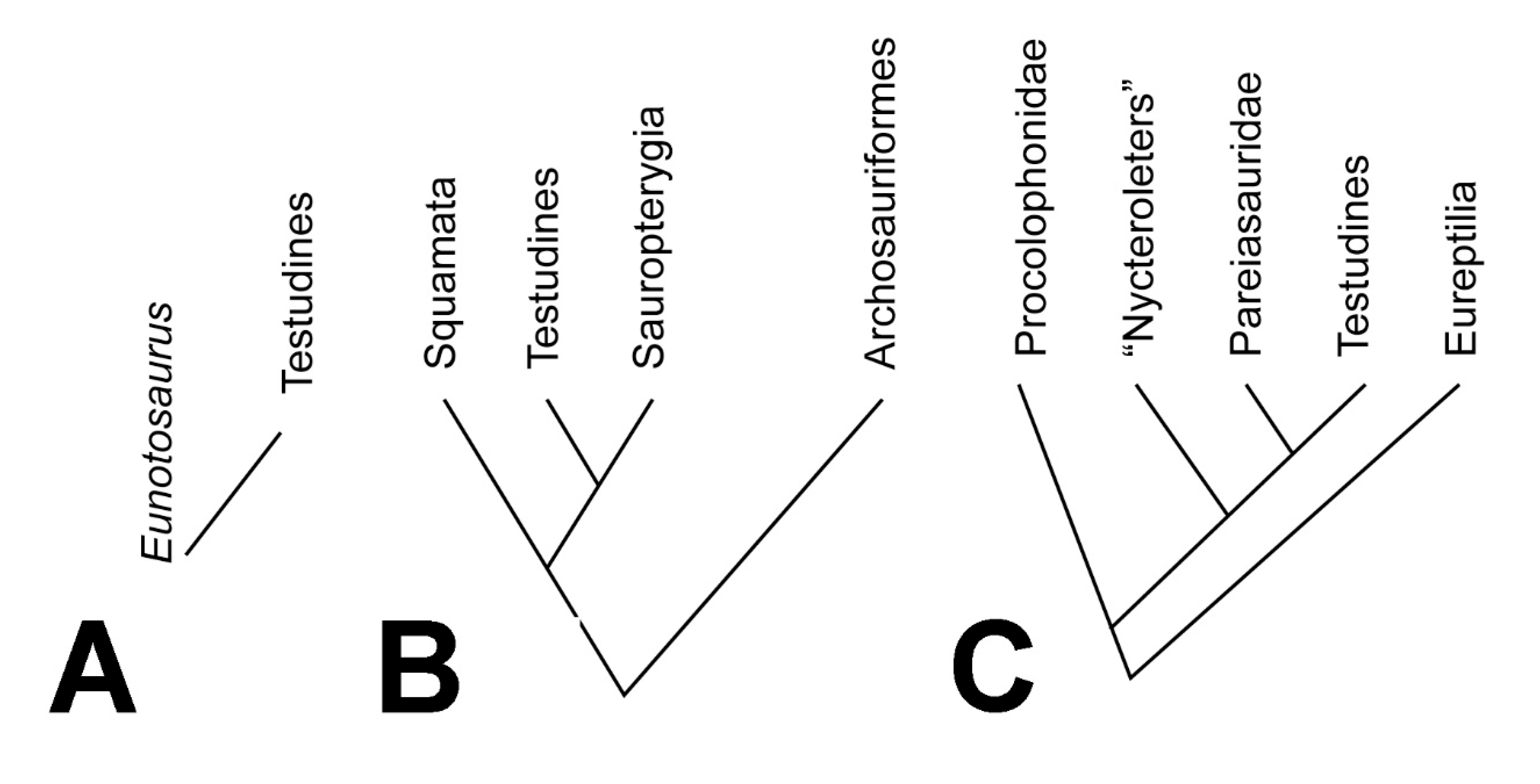
2. Cladistic Solution 1: Turtles as Sauropterygians
3. Cladistic Solution 2: Turtles and Eunotosaurus
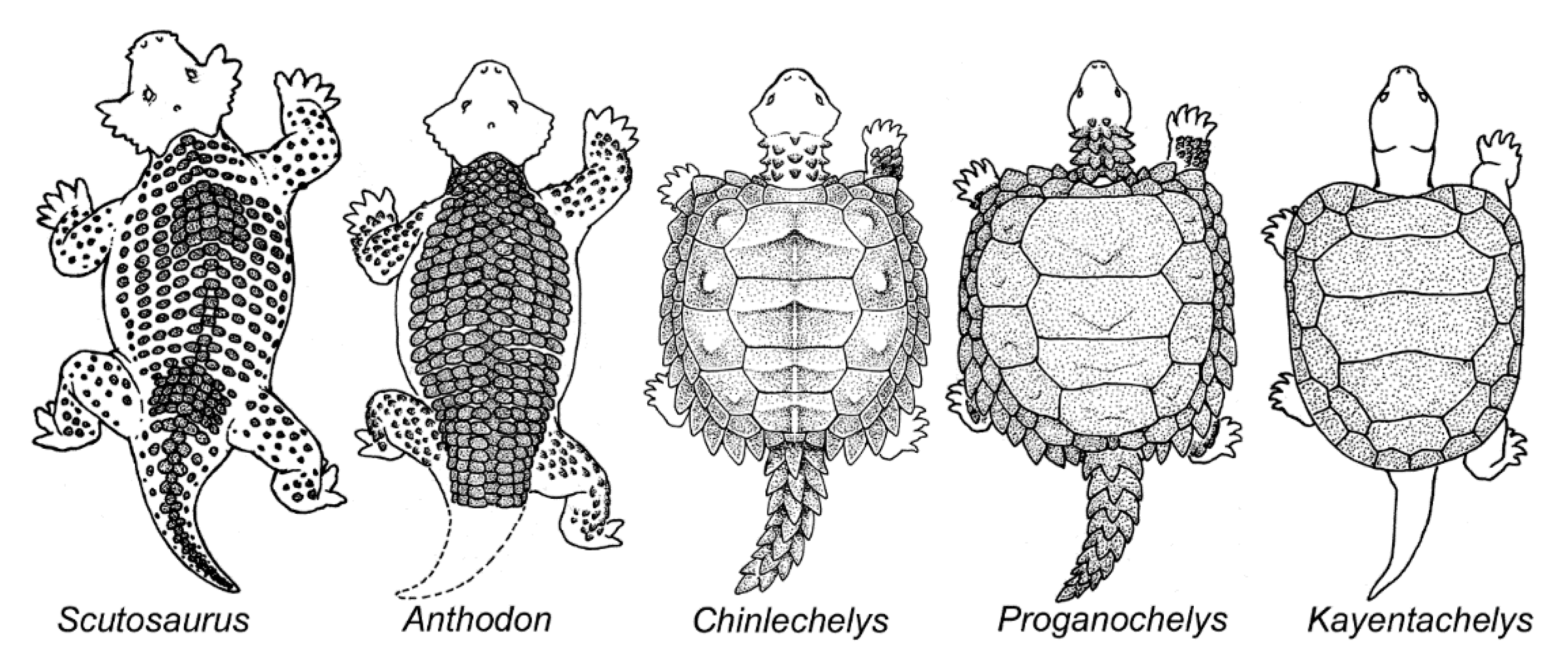
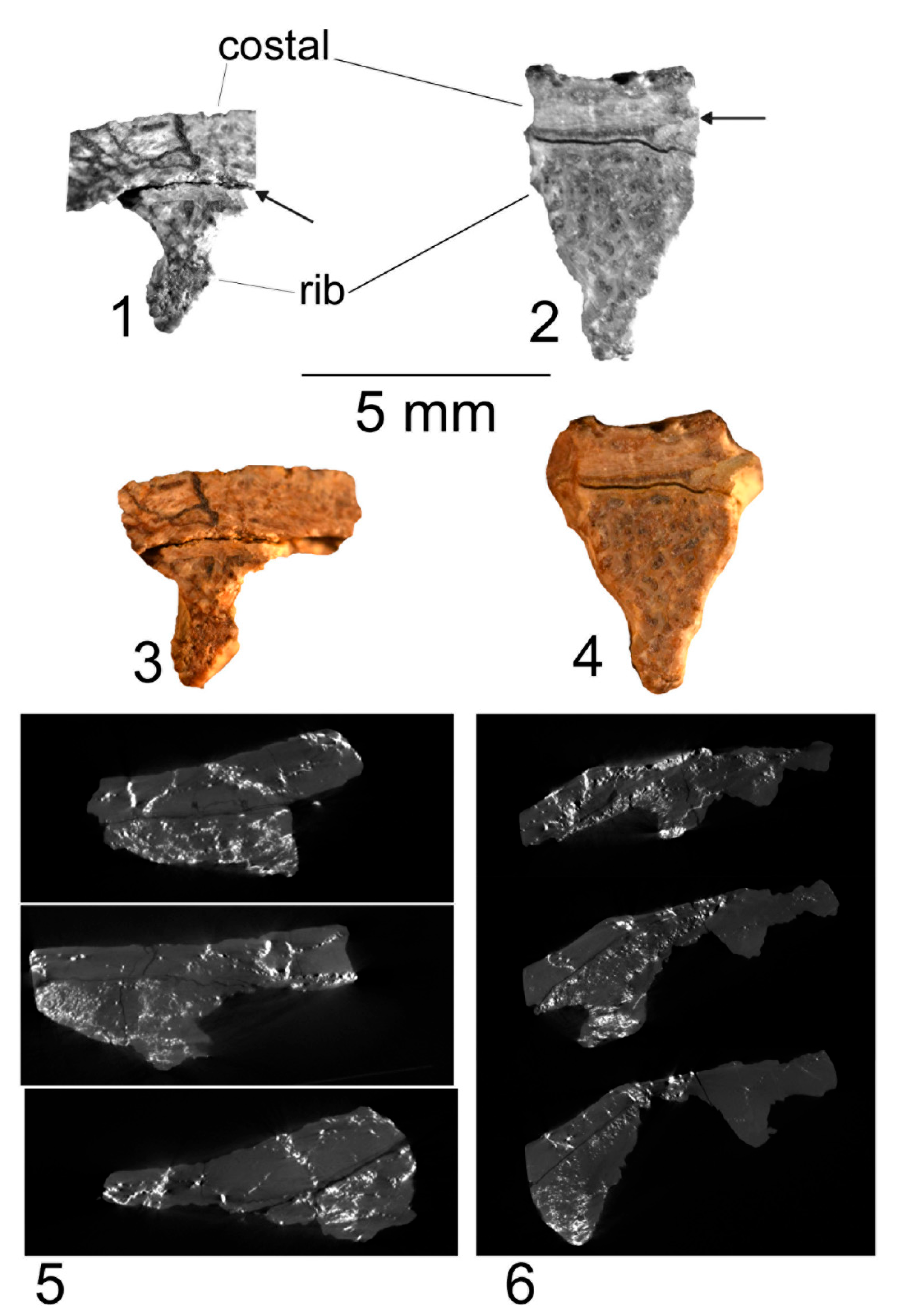
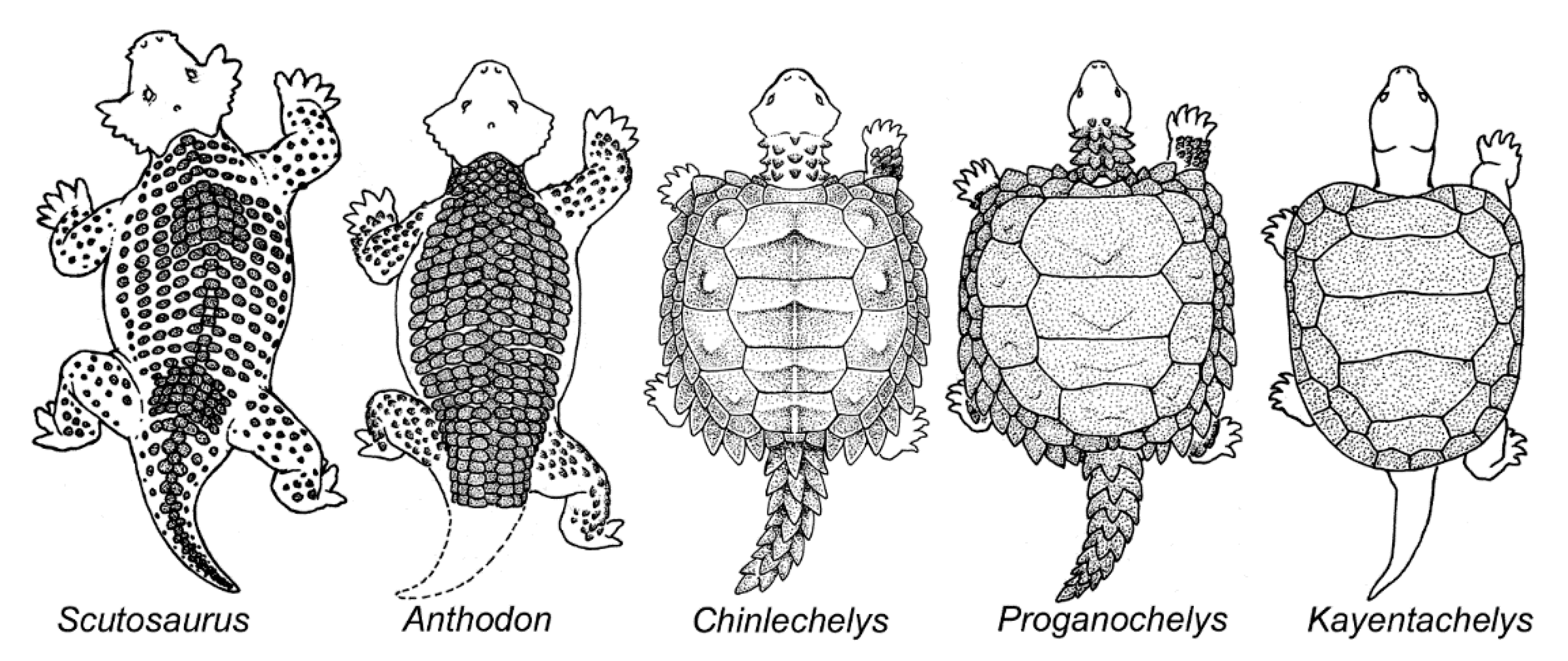
4. Origin of the Turtle Carapace
5. Convergence Is Rampant
6. Integrating Other Data
7. Turtle Tracks
8. Pareiasaurs and Turtles
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lichtig, A.J.; Lucas, S.G. Chinlechelys from the Upper Triassic of New Mexico, USA, and the origin of turtles. Palaeontol. Electron. 2021, 24, a13. [Google Scholar] [CrossRef] [PubMed]
- Rieppel, O.; Reisz, R.R. The origin and early evolution of turtles. Annu. Rev. Ecol. Syst. 1999, 30, 1–22. [Google Scholar] [CrossRef]
- Scheyer, T.M. Comparative Bone Histology of the Turtle Shell (Carapace and Plastron) Implications for Turtle Systematics, Functional Morphology and Turtle Origins. Unpublished. Ph.D. Thesis, University of Bonn, Bonn, Germany, 2007. [Google Scholar]
- Bever, G.S.; Lyson, T.R.; Field, D.J.; Bhullar, B.S. Evolutionary origin of the turtle skull. Nature 2015, 525, 239–242. [Google Scholar] [CrossRef] [PubMed]
- Watson, D.M.S. Eunotosaurus africanus Seeley, and the ancestry of the Chelonia. Proc. Zool. Soc. Lond. 1914, 11, 1011–1020. [Google Scholar] [CrossRef]
- Lee, M.S.Y. Evolutionary Morphology of Pareiasaurs. Unpublished. Ph.D. Thesis, Queens’ College, University of Cambridge, Cambridge, UK, 1994. [Google Scholar]
- Szczygielski, T. Homeotic shift at the dawn of the turtle evolution. R. Soc. Open Sci. 2017, 4, 4. [Google Scholar] [CrossRef] [PubMed]
- Rieppel, O. Turtles as Hopeful Monsters: Origins and Evolution; Indiana University Press: Bloomington, IN, USA, 2017. [Google Scholar]
- Losos, J.B.; Glor, R.E.; Kolbe, J.J.; Nicholson, K. Adaptation, speciation, and convergence: A hierarchical analysis of adaptive radiation in Caribbean Anolis lizards1. Ann. Mo. Bot. Gard. 2006, 93, 24–33. [Google Scholar] [CrossRef]
- Huie, J.M.; Prates, I.; Bell, R.C.; de Queiroz, K. Convergent patterns of adaptive radiation between island and mainland Anolis lizards. Biol. J. Linn. Soc. 2021, 134, 85–110. [Google Scholar] [CrossRef]
- Gaffney, E.S.; Jenkins, F.A., Jr. The cranial morphology of Kayentachelys, an Early Jurassic cryptodire, and the early history of turtles. Acta Zool. 2010, 91, 335–368. [Google Scholar]
- Tokita, M.; Watanabe, T.; Sato, H.; Kondo, S.; Kitayama, C. A comparative study of cranial osteogenesis in turtles: Implications for the diversification of skull morphology. Zoomorphology 2021, 140, 539–554. [Google Scholar] [CrossRef]
- Lichtig, A.J.; Lucas, S.G.; Klein, H.; Lovelace, D.M. Triassic turtle tracks and the origin of turtles: Hist. Biol. 2018, 30, 1112–1122. [Google Scholar] [CrossRef]
- Avanzini, M.; García-Ramos, J.C.; Lires, J.; Menegon, M.; Pinuela, L.A.U.R.A.; Fernández, L.A. Turtle tracks from the late Jurassic of Asturias, Spain. Acta Palaeontol. Pol. 2005, 50, 743–755. [Google Scholar]
- Rühle v. Lilienstern, H. Fährten und Spüren im Chirotheriem−Sandstein von Südthüringen. Fortschr. Der Geol. Und Paläontolog. 1939, 12, 293–387. [Google Scholar]
- Haubold, H. Die tetrapodenfährten des Buntsandsteins in der Deutschen Demokratischen Republik und in Westdeutschland und ihre Äquivalente in der gesamten Trias. Paläont Abh 1971, 4, 395–660. [Google Scholar]
- Lovelace, D.M.; Lovelace, S.D. Paleoenvironments and paleoecology of a Lower Triassic invertebrate and vertebrate ichnoassemblage from the Red Peak Formation (Chugwater Group), central Wyoming. Palaios 2012, 27, 636–657. [Google Scholar] [CrossRef]
- Storrs, G.W. Anatomy and relationships of Corosaurus alcovensis (Diapsida: Sauropterygia) and the Triassic Alcova Limestone of Wyoming. Peabody Mus. Nat. Hist. 1991, 44, 1–163. [Google Scholar]
- Zhang, Q.; Wen, W.; Hu, S.; Benton, M.J.; Zhou, C.; Xie, T.; Zhang, Q. Nothosaur foraging tracks from the Middle Triassic of southwestern China. Nat. Commun. 2014, 5, 3973. [Google Scholar] [CrossRef] [PubMed]
- Xing, L.; Klein, H.; Lockley, M.G.; Wu, X.C.; Benton, M.J.; Zeng, R.; Romilio, A. Footprints of marine reptiles from the Middle Triassic (Anisian-Ladinian) Guanling Formation of Guizhou Province, southwestern China: The earliest evidence of synchronous style of swimming. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2020, 558, 109943. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lichtig, A.J.; Lucas, S.G. Turtle Origins: Chinlechelys tenertesta and Convergence in Modern Cladistic Analysis. Proceedings 2023, 87, 4. https://doi.org/10.3390/IECG2022-14068
Lichtig AJ, Lucas SG. Turtle Origins: Chinlechelys tenertesta and Convergence in Modern Cladistic Analysis. Proceedings. 2023; 87(1):4. https://doi.org/10.3390/IECG2022-14068
Chicago/Turabian StyleLichtig, Asher J., and Spencer G. Lucas. 2023. "Turtle Origins: Chinlechelys tenertesta and Convergence in Modern Cladistic Analysis" Proceedings 87, no. 1: 4. https://doi.org/10.3390/IECG2022-14068
APA StyleLichtig, A. J., & Lucas, S. G. (2023). Turtle Origins: Chinlechelys tenertesta and Convergence in Modern Cladistic Analysis. Proceedings, 87(1), 4. https://doi.org/10.3390/IECG2022-14068