

Impact of Drone Disturbances on Wildlife: A Review †
 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
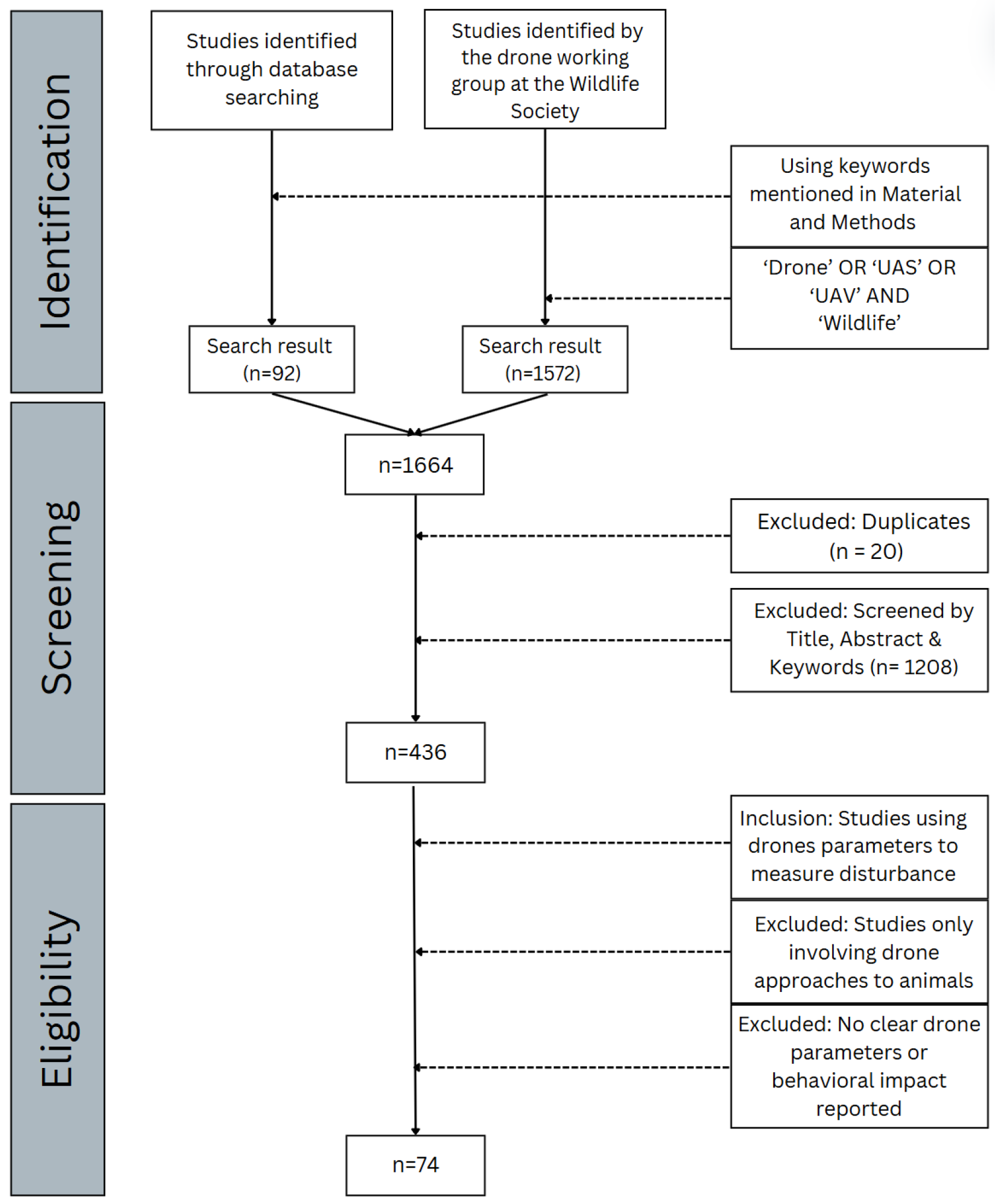
2. Material and Methods
3. Drones and Wildlife
3.1. Operational Factors Influencing Wildlife Responses
3.2. Sensory Stimuli: Noise and Visual Impact


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study | Species Studied | Drone Used | Study Context | UAV Flight Altitude | Observed Impact | Recommendations |
|---|---|---|---|---|---|---|
| Fleeing by Whimbrel in Response to a Recreational Drone (2016) [55] | Whimbrel (Numenius phaeopus) | Recreational drone (Phantom type) | Observing the response of Whimbrel to a recreational drone | Hovered at 5 m and 20 m altitudes | Whimbrels exhibited strong “fleeing” responses | Avoiding low-altitude flights near sensitive bird species. |
| Using Two Drones to Monitor Visual and Acoustic Behaviour of Gray Whales (Eschrichtius robustus) in Baja California, Mexico (2020) [56] | Gray whales (Eschrichtius robustus) | SwellPro SplashDrone 3+ (acoustic) and DJI Phantom 4 (visual) | Testing dual-drone system for simultaneous visual and acoustic monitoring of gray whale behavior | 30 m for visual drone; acoustic drone within 50 m | Minimal disturbance observed. | More research is needed on dual-drone monitoring with varying altitudes. |
| Measuring Disturbance at Swift Breeding Colonies Due to the Visual Aspects of a Drone (2021) [23] | Great dusky swift (Cypseloides senex), White-collared swift (Streptoprocne zonaris) | DJI Mavic Pro | Assessing visual drone disturbance on great dusky swift | 25 m to 64 m | At <40 m, disturbance increased to >60%, leading to temporary colony abandonment. | Recommended flight altitude: >50 m to minimize disturbance. |
| Determination of Optimal Flight Altitude to Minimise Acoustic Drone Disturbance to Wildlife Using Species Audiograms (2021) [44] | Various mammals (20 species) | DJI Inspire 2, Phantom 4, Mavic 2, Mavic Pro, Mavic Pro Platinum, Mavic Mini, Spark | Determining the minimum flight altitude to minimizes UAV noise disturbance | 5 m to 120 m | The optimal altitude varies by species and drone model | Recommended altitudes of 35 m to 120 m depending on species. |
| Behavioral Responses of a Nocturnal Burrowing Marsupial (Lasiorhinus latifrons) to Drone Flight (2021) [57] | Southern Hairy-Nosed Wombat (Lasiorhinus latifrons) | DJI Phantom 4 Pro | Assessing the behavioral responses of southern hairy-nosed wombats during day and night | 100 m, 60 m, 30 m | Night flights triggered stronger retreat responses | Conduct flights outside sensitive hours to reduce disturbance. |
| Drone noise differs by flight maneuver and model: implications for animal surveys (2024) [58] | Not species-specific | DJI Matrice 300, Matrice 200, Phantom 3, Autel Evo II | Evaluating noise emission differences by drone model, flight maneuver, and altitude | 15 m to 120 m | Flyover and turning maneuvers at higher altitudes generated minimal noise | Avoid repetitive sessions and minimize prolonged hovering. |
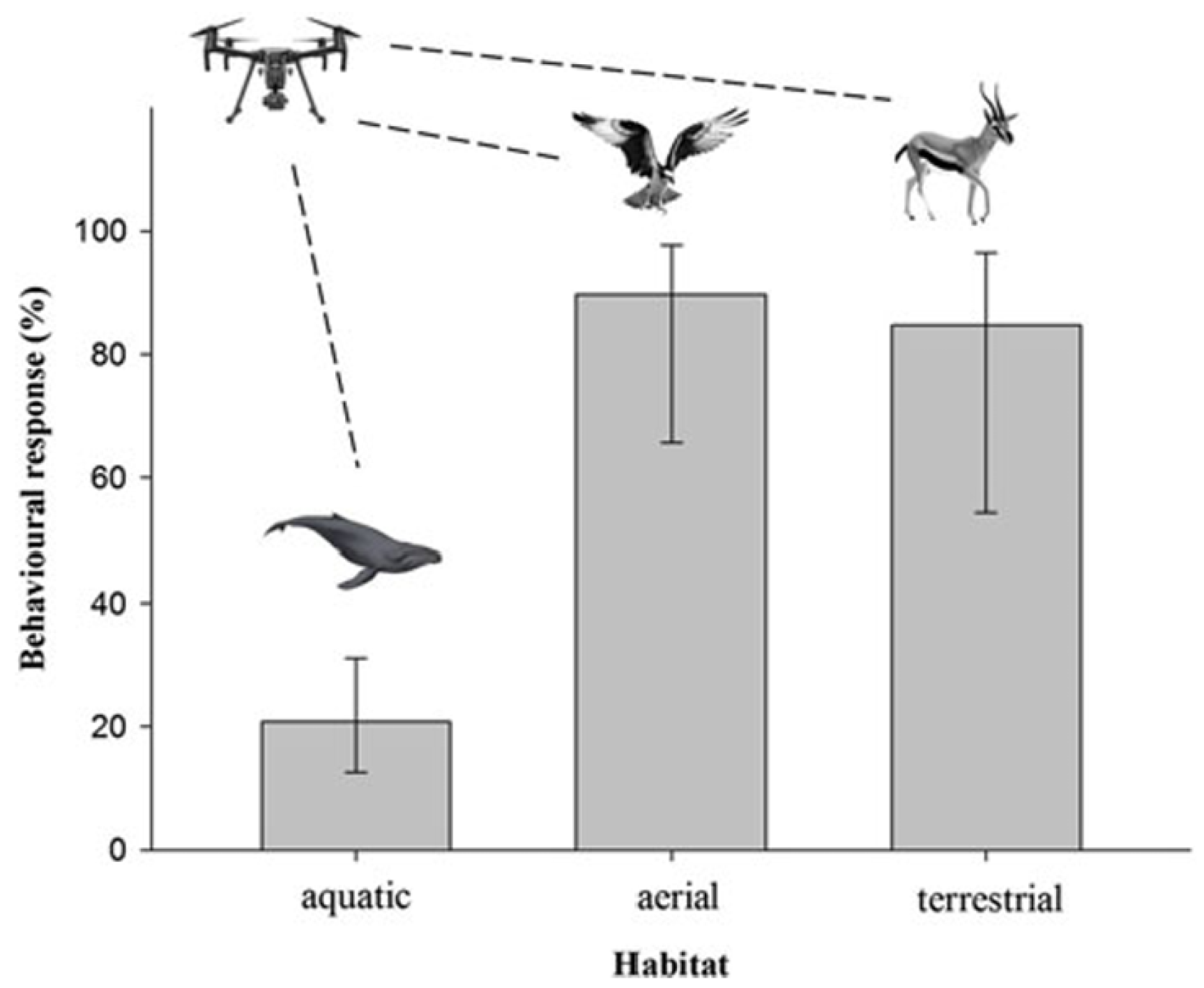
3.3. Species- and Habitat-Specific Sensitivities
| Study | Species Studied | Drone Used | Study Context | UAV Flight Altitude | Observed Impact | Recommendations |
|---|---|---|---|---|---|---|
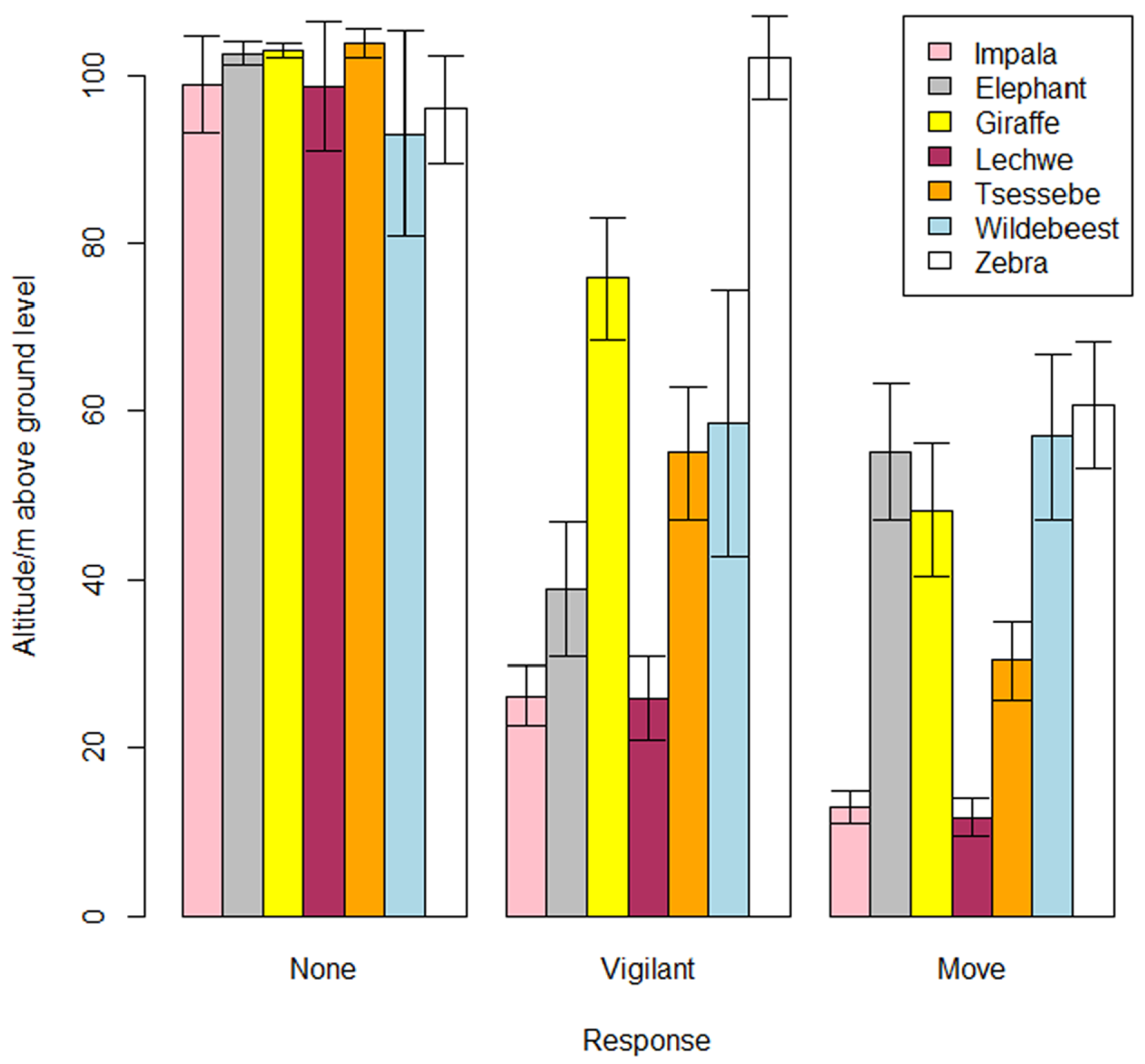
| Terrestrial Mammalian Wildlife Responses to Unmanned Aerial Systems Approaches (2019) [30] | Elephants, Giraffes, Wildebeest, Zebras, Impala, Lechwe, Tsessebe | DJI Phantom 3, DJI Inspire 1 | Assessing vertical and horizontal UAS approaches | 10 m to 100 m | Horizontal approaches triggered fewer reactions than vertical ones | Recommended minimum altitude of 60 m and horizontal distance of 100 m. |
| Responses of Bottlenose Dolphins (Tursiops spp.) to Small Drones (2020) [14] | Bottlenose dolphins (Tursiops spp.) | DJI Phantom 4 | Examining drone altitudes and observation durations influence | 5 m to 60 m | Groups exhibited more reactions at <30 m altitude. Longer hovering times increased the probability of behavioral responses | Recommended flight altitude: ≥30 m to minimize disturbance. |
| Ungulate responses and habituation to unmanned aerial vehicles in Africa’s savanna (2023) [75] | Oryx, Kudu, Springbok, Giraffe, Eland, Hartebeest, Plains Zebra, Impala | DJI Phantom 3, DJI Mavic Pro, Custom X8 (Octocopter), Sky Eye | Assessing the behavioral responses of ungulates to UAV flights | 15 m to 55 m | Species-specific responses varied by altitude and UAV type | Opting for species-specific flight altitudes to reduce disturbance impact. |
| Estuary Stingray (Dasyatis fluviorum) Behavior Does Not Change in Response to Drone Altitude (2023) [76] | Estuary stingray (Dasyatis fluviorum) | DJI Mavic Platinum Pro | Assessing if drone altitude influences the behavior of estuary stingrays | 5 m to 30 m, reducing by 5 m intervals | No significant changes in swimming, foraging, or resting behavior | Further research required into physiological and long-term impact of drone disturbances. |
| Assessing the Behavioral Responses of Small Cetaceans to Unmanned Aerial Vehicles (2021) [77] | Common dolphins (Delphinus delphis), Bottlenose dolphins (Tursiops truncatus) | DJI Phantom 2 | Evaluating the behavioral responses to drones at different altitudes | 5 m to 70 m (descending in 5 m intervals) | No significant responses in diving or swimming speed for either species | Observing animals’ reaction and adjusting flight according is essential for minimizing disturbance. |
| Sociability Strongly Affects the Behavioral Responses of Wild Guanacos to Drones (2021) [78] | Guanacos (Lama guanicoe) | DJI Phantom 4 Advanced | Examining group size, social composition, and flight characteristics on guanaco | 60 m and 180 m with low (2–4 m/s) and high (8–10 m/s) speed | Groups exhibited greater reaction probabilities. Low altitudes (<60 m) increased reactions | To consider collective responses versus those of lone individuals when utilizing drones in ecological studies. |
4. Best Practices and Recommendations
4.1. Flight Parameters
4.2. Species-Specific and Contextual Sensitivity
4.3. Environmental and Temporal Considerations
4.4. Behavioral Monitoring and Adaptive Drone Management
4.5. Data Collection and Standardized Reporting
4.6. Regulatory Compliance
5. Key Challenges in Wildlife Responses to Drones
5.1. Species Responses and Long-Term Impacts of Drones
5.2. Limitations in Multi-Species Risk Assessment
5.3. Ethical and Conservation Considerations
5.4. Technological Constraints and Flight Precision
5.5. Regulatory Limitations
6. Future Research Directions
6.1. Innovations in Drone Technology
6.2. Tailoring Guidelines for Diverse Ecosystems
6.3. Global Collaboration and Standardization
6.4. Development of Testing Standards for Drones
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Geoghegan, J.L.; Pirotta, V.; Harvey, E.; Smith, A.; Buchmann, J.P.; Ostrowski, M.; Eden, J.S.; Harcourt, R.; Holmes, E.C. Virological Sampling of Inaccessible Wildlife with Drones. Viruses 2018, 10, 300. [Google Scholar] [CrossRef] [PubMed]
- Burnett, J.D.; Lemos, L.; Barlow, D.; Wing, M.G.; Chandler, T.; Torres, L.G. Estimating morphometric attributes of baleen whales with photogrammetry from small UASs: A case study with blue and gray whales. Mar. Mammal Sci. 2019, 35, 108–139. [Google Scholar] [CrossRef]
- Graving, J.M.; Chae, D.; Naik, H.; Li, L.; Koger, B.; Costelloe, B.R.; Couzin, I.D. DeepPoseKit, a software toolkit for fast and robust animal pose estimation using deep learning. eLife 2019, 8, e47994. [Google Scholar] [CrossRef] [PubMed]
- He, G.; Yang, H.; Pan, R.; Sun, Y.; Zheng, P.; Wang, J.; Jin, X.; Zhang, J.; Li, B.; Guo, S. Using unmanned aerial vehicles with thermal-image acquisition cameras for animal surveys: A case study on the Sichuan snub-nosed monkey in the Qinling Mountains. Integr. Zool. 2020, 15, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Linchant, J.; Lhoest, S.; Quevauvillers, S.; Lejeune, P.; Vermeulen, C.; Semeki Ngabinzeke, J.; Luse Belanganayi, B.; Delvingt, W.; Bouché, P. UAS imagery reveals new survey opportunities for counting hippos. PLoS ONE 2018, 13, e0206413. [Google Scholar] [CrossRef]
- Mulero-Pázmány, M.; Stolper, R.; Van Essen, L.; Negro, J.J.; Sassen, T. Remotely Piloted Aircraft Systems as a Rhinoceros Anti-Poaching Tool in Africa. PLoS ONE 2014, 9, e83873. [Google Scholar] [CrossRef]
- Szantoi, Z.; Smith, S.E.; Strona, G.; Koh, L.P.; Wich, S.A. Mapping orangutan habitat and agricultural areas using Landsat OLI imagery augmented with unmanned aircraft system aerial photography. Int. J. Remote Sens. 2017, 38, 2231–2245. [Google Scholar] [CrossRef]
- Fiori, L.; Martinez, E.; Bader, M.K.F.; Orams, M.B.; Bollard, B. Insights into the use of an unmanned aerial vehicle (UAV) to investigate the behavior of humpback whales (Megaptera novaeangliae) in Vava’u, Kingdom of Tonga. Mar. Mammal Sci. 2020, 36, 209–223. [Google Scholar] [CrossRef]
- Gray, P.C.; Fleishman, A.B.; Klein, D.J.; McKown, M.W.; Bézy, V.S.; Lohmann, K.J.; Johnston, D.W. A convolutional neural network for detecting sea turtles in drone imagery. Methods Ecol. Evol. 2019, 10, 345–355. [Google Scholar] [CrossRef]
- Colefax, A.P.; Butcher, P.A.; Pagendam, D.E.; Kelaher, B.P. Reliability of marine faunal detections in drone-based monitoring. Ocean Coast. Manag. 2019, 174, 108–115. [Google Scholar] [CrossRef]
- Rebolo-Ifrán, N.; Grilli, M.G.; Lambertucci, S.A. Drones as a Threat to Wildlife: YouTube Complements Science in Providing Evidence about Their Effect. Environ. Conserv. 2019, 46, 205–210. [Google Scholar] [CrossRef]
- Baltrus, A. Drone Harasses Bighorn Sheep at Zion National Park. Available online: https://www.nationalparkstraveler.org/2014/05/drones-seen-harassing-wildlife-zion-national-park25034 (accessed on 2 December 2024).
- Mesquita, G.P.; Mulero-Pázmány, M.; Wich, S.A.; Rodríguez-Teijeiro, J.D. Terrestrial Megafauna Response to Drone Noise Levels in Ex Situ Areas. Drones 2022, 6, 333. [Google Scholar] [CrossRef]
- Giles, A.B.; Butcher, P.A.; Colefax, A.P.; Pagendam, D.E.; Mayjor, M.; Kelaher, B.P. Responses of bottlenose dolphins (Tursiops spp.) to small drones. Aquat. Conserv. Mar. Freshw. Ecosyst. 2021, 31, 677–684. [Google Scholar] [CrossRef]
- Schiffman, R. Drones Flying High as New Tool for Field Biologists. Science 2014, 344, 459. [Google Scholar] [CrossRef]
- Smith, C.E.; Sykora-Bodie, S.T.; Bloodworth, B.; Pack, S.M.; Spradlin, T.R.; LeBoeuf, N.R. Assessment of known impacts of unmanned aerial systems (UAS) on marine mammals: Data gaps and recommendations for researchers in the United States. J. Unmanned Veh. Syst. 2016, 4, 31–44. [Google Scholar] [CrossRef]
- Weston, M.A.; O’Brien, C.; Kostoglou, K.N.; Symonds, M.R.E. Escape responses of terrestrial and aquatic birds to drones: Towards a code of practice to minimize disturbance. J. Appl. Ecol. 2020, 57, 777–785. [Google Scholar] [CrossRef]
- Vas, E.; Lescroël, A.; Duriez, O.; Boguszewski, G.; Grémillet, D. Approaching birds with drones: First experiments and ethical guidelines. Biol. Lett. 2015, 11, 20140754. [Google Scholar] [CrossRef]
- Bech-Hansen, M.; Kallehauge, R.; Lauritzen, J.; Sørensen, M.; Laubek, B.; Jensen, L.; Pertoldi, C.; Bruhn, D. Evaluation of disturbance effect on geese caused by an approaching unmanned aerial vehicle. Bird Conserv. Int. 2020, 30, 169–175. [Google Scholar] [CrossRef]
- Rümmler, M.C.; Mustafa, O.; Maercker, J.; Peter, H.U.; Esefeld, J. Sensitivity of Adélie and Gentoo penguins to various flight activities of a micro UAV. Polar Biol. 2018, 41, 2481–2493. [Google Scholar] [CrossRef]
- Erbe, C.; Dent, M.L.; Gannon, W.L.; McCauley, R.D.; Römer, H.; Southall, B.L.; Stansbury, A.L.; Stoeger, A.S.; Thomas, J.A. The Effects of Noise on Animals. In Exploring Animal Behavior Through Sound: Volume 1: Methods; Erbe, C., Thomas, J.A., Eds.; Springer International Publishing: Cham, Switzerland, 2022; pp. 459–506. [Google Scholar] [CrossRef]
- Scobie, C.A.; Hugenholtz, C.H. Wildlife monitoring with unmanned aerial vehicles: Quantifying distance to auditory detection. Wildl. Soc. Bull. 2016, 40, 781–785. [Google Scholar] [CrossRef]
- Mesquita, G.P.; Rodríguez-Teijeiro, J.D.; Wich, S.A.; Mulero-Pázmány, M. Measuring disturbance at swift breeding colonies due to the visual aspects of a drone: A quasi-experiment study. Curr. Zool. 2020, 67, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Afridi, S.; Hlebowicz, K.; Cawthorne, D.; Lundquist, U.P.S. Unveiling the Impact of Drone Noise on Wildlife: A Crucial Research Imperative. In Proceedings of the 2024 International Conference on Unmanned Aircraft Systems (ICUAS), Chania, Greece, 4–7 June 2024; pp. 1409–1416. [Google Scholar] [CrossRef]
- Caughley, G. Bias in Aerial Survey. J. Wildl. Manag. 1974, 38, 921–933. [Google Scholar] [CrossRef]
- Andersen, D.E.; Rongstad, O.J.; Mytton, W.R. Response of Nesting Red-Tailed Hawks to Helicopter Overflights. Condor 1989, 91, 296–299. [Google Scholar] [CrossRef]
- Patenaude, N.J.; Richardson, W.J.; Smultea, M.A.; Koski, W.R.; Miller, G.W.; Würsig, B.; GReene, C.R., Jr. Aircraft sound and sisturbance to bowhead and beluga whalees during spring migration in the Alaskan Beaufort sea. Mar. Mammal Sci. 2002, 18, 309–335. [Google Scholar] [CrossRef]
- Weimerskirch, H.; Prudor, A.; Schull, Q. Flights of drones over sub-Antarctic seabirds show species-and status-specific behavioural and physiological responses. Polar Biol. 2018, 41, 259–266. [Google Scholar] [CrossRef]
- Brunton, E.; Bolin, J.; Leon, J.; Burnett, S. Fright or Flight? Behavioural Responses of Kangaroos to Drone-Based Monitoring. Drones 2019, 3, 41. [Google Scholar] [CrossRef]
- Bennitt, E.; Bartlam-Brooks, H.L.; Hubel, T.Y.; Wilson, A.M. Terrestrial mammalian wildlife responses to Unmanned Aerial Systems approaches. Sci. Rep. 2019, 9, 2142. [Google Scholar] [CrossRef]
- Dundas, S.J.; Vardanega, M.; O’Brien, P.; McLeod, S.R. Quantifying Waterfowl Numbers: Comparison of Drone and Ground-Based Survey Methods for Surveying Waterfowl on Artificial Waterbodies. Drones 2021, 5, 5. [Google Scholar] [CrossRef]
- Arona, L.; Dale, J.; Heaslip, S.G.; Hammill, M.O.; Johnston, D.W. Assessing the disturbance potential of small unoccupied aircraft systems (UAS) on gray seals (Halichoerus grypus) at breeding colonies in Nova Scotia, Canada. PeerJ 2018, 6, e4467. [Google Scholar] [CrossRef]
- Hartmann, W.L.; Fishlock, V.; Leslie, A. First guidelines and suggested best protocol for surveying African elephants (Loxodonta africana) using a drone. Koedoe 2021, 63, a1687. [Google Scholar] [CrossRef]
- Lu, V.; Xu, F.; Turghan, M.A. Przewalski’s Horses (Equus ferus przewalskii) Responses to Unmanned Aerial Vehicles Flights under Semireserve Conditions: Conservation Implication. Int. J. Zool. 2021, 2021, 6687505. [Google Scholar] [CrossRef]
- Damiano, S. Dolphin Behavioral Responses to Uncrewed Aerial Systems as a Function of Exposure, Height, and Type. Master’s Thesis, Stephen F Austin State University, Nacogdoches, TX, USA, 2023. [Google Scholar]
- Aubin, J.A.; Mikus, M.A.; Michaud, R.; Mennill, D.; Vergara, V. Fly with care: Belugas show evasive responses to low altitude drone flights. Mar. Mammal Sci. 2023, 39, 718–739. [Google Scholar] [CrossRef]
- Orange, J.P.; Bielefeld, R.R.; Cox, W.A.; Sylvia, A.L. Impacts of Drone Flight Altitude on Behaviors and Species Identification of Marsh Birds in Florida. Drones 2023, 7, 584. [Google Scholar] [CrossRef]
- Stepien, E.N.; Khan, J.; Galatius, A.; Teilmann, J. How low can you go? Exploring impact of drones on haul out behaviour of harbour-and grey seals. Front. Mar. Sci. 2024, 11, 1411292. [Google Scholar] [CrossRef]
- Herman, L.M.; Peacock, M.F.; Yunker, M.P.; Madsen, C.J. Bottle-Nosed Dolphin: Double-Slit Pupil Yields Equivalent Aerial and Underwater Diurnal Acuity. Science 1975, 189, 650–652. [Google Scholar] [CrossRef]
- Bauer, G.B.; Colbert, D.E.; Gaspard, J.C.; Littlefield, B.L.; Fellner, W. Underwater Visual Acuity of Florida Manatees (Trichechus manatus latirostris). Int. J. Comp. Psychol. 2003, 16, 130–142. [Google Scholar] [CrossRef]
- Ramos, E.A.; Maloney, B.; Magnasco, M.O.; Reiss, D. Bottlenose Dolphins and Antillean Manatees Respond to Small Multi-Rotor Unmanned Aerial Systems. Front. Mar. Sci. 2018, 5, 316. [Google Scholar] [CrossRef]
- Moss, C.J.; Croze, H.; Lee, P.C. The Amboseli Elephants: A Long-Term Perspective on a Long-Lived Mammal; University of Chicago Press: Chicago, IL, USA, 2011. [Google Scholar]
- Penny, S.G.; White, R.L.; Scott, D.M.; MacTavish, L.; Pernetta, A.P. Using drones and sirens to elicit avoidance behaviour in white rhinoceros as an anti-poaching tactic. Proc. R. Soc. B Biol. Sci. 2019, 286, 20191135. [Google Scholar] [CrossRef]
- Duporge, I.; Spiegel, M.P.; Thomson, E.R.; Chapman, T.; Lamberth, C.; Pond, C.; Macdonald, D.W.; Wang, T.; Klinck, H. Determination of optimal flight altitude to minimise acoustic drone disturbance to wildlife using species audiograms. Methods Ecol. Evol. 2021, 12, 2196–2207. [Google Scholar] [CrossRef]
- Nowak, R.M. Walker’s Mammals of the World; The Johns Hopkins University Press: Baltimore, MD, USA, 1999; Volume 1. [Google Scholar]
- Egan, C.C.; Blackwell, B.F.; Fernández-Juricic, E.; Klug, P.E. Testing a key assumption of using drones as frightening devices: Do birds perceive drones as risky? Condor 2020, 122, duaa014. [Google Scholar] [CrossRef]
- Wang, Z.; Griffin, A.S.; Lucas, A.; Wong, K. Psychological warfare in vineyard: Using drones and bird psychology to control bird damage to wine grapes. Crop Prot. 2019, 120, 163–170. [Google Scholar] [CrossRef]
- Storms, R.F.; Carere, C.; Musters, R.; van Gasteren, H.; Verhulst, S.; Hemelrijk, C.K. Deterrence of birds with an artificial predator, the RobotFalcon. J. R. Soc. Interface 2022, 19, 20220497. [Google Scholar] [CrossRef] [PubMed]
- McEvoy, J.F.; Hall, G.P.; McDonald, P.G. Evaluation of unmanned aerial vehicle shape, flight path and camera type for waterfowl surveys: Disturbance effects and species recognition. PeerJ 2016, 4, e1831. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulou, M.; Hildenbrandt, H.; Sankey, D.W.E.; Portugal, S.J.; Hemelrijk, C.K. Self-organization of collective escape in pigeon flocks. PLoS Comput. Biol. 2022, 18, e1009772. [Google Scholar] [CrossRef]
- Augustine, J.K.; Burchfield, D. Evaluation of unmanned aerial vehicles for surveys of lek-mating grouse. Wildl. Soc. Bull. 2022, 46, e1333. [Google Scholar] [CrossRef]
- Wolfe, D.H.; Patten, M.A.; Shochat, E.; Pruett, C.L.; Sherrod, S.K. Causes and Patterns of Mortality in Lesser Prairie-chickens Tympanuchus pallidicinctus and Implications for Management. Wildl. Biol. 2007, 13, 95–104. [Google Scholar] [CrossRef]
- Brisson-Curadeau, É.; Bird, D.; Burke, C.; Fifield, D.A.; Pace, P.; Sherley, R.B.; Elliott, K.H. Seabird species vary in behavioural response to drone census. Sci. Rep. 2017, 7, 17884. [Google Scholar] [CrossRef]
- ABC News. Video: Raven Attacks Drone Delivering Coffee. Available online: https://abcnews.go.com/International/video/raven-attacks-drone-delivering-coffee-80189172 (accessed on 14 December 2024).
- Allport, G. Fleeing by Whimbrel (Numenius phaeopus) in response to a recreational drone in Maputo Bay, Mozambique. Biodivers. Obs. 2016, 7, 1–5. [Google Scholar]
- Frouin-Mouy, H.; Tenorio-Hallé, L.; Thode, A.; Swartz, S.; Urbán, J. Using two drones to simultaneously monitor visual and acoustic behaviour of gray whales (Eschrichtius robustus) in Baja California, Mexico. J. Exp. Mar. Biol. Ecol. 2020, 525, 151321. [Google Scholar] [CrossRef]
- Headland, T.; Ostendorf, B.; Taggart, D. The behavioral responses of a nocturnal burrowing marsupial (Lasiorhinus latifrons) to drone flight. Ecol. Evol. 2021, 11, 12173–12181. [Google Scholar] [CrossRef]
- Macke, E.N.; Jones, L.R.; Iglay, R.B.; Elmore, J.A. Drone noise differs by flight maneuver and model: Implications for animal surveys. Drone Syst. Appl. 2024, 12, 1–5. [Google Scholar] [CrossRef]
- Harris, C.M. Absorption of sound in air versus humidity and temperature. J. Acoust. Soc. Am. 1966, 40, 148–159. [Google Scholar] [CrossRef]
- Franchini, M.; Atzeni, L.; Lovari, S.; Nasanbat, B.; Ravchig, S.; Herrador, F.C.; Bombieri, G.; Augugliaro, C. Spatiotemporal behavior of predators and prey in an arid environment of Central Asia. Curr. Zool. 2023, 69, 670–681. [Google Scholar] [CrossRef]
- Preisser, E.L.; Bolnick, D.I.; Benard, M.F. Scared to death? The effects of intimidation and consumption in predator–prey interactions. Ecology 2005, 86, 501–509. [Google Scholar] [CrossRef]
- Brown, J.S.; Laundré, J.W.; Gurung, M. The ecology of fear: Optimal foraging, game theory, and trophic interactions. J. Mammal. 1999, 80, 385–399. [Google Scholar] [CrossRef]
- Schroeder, N.M.; Panebianco, A.; Gonzalez Musso, R.; Carmanchahi, P. An experimental approach to evaluate the potential of drones in terrestrial mammal research: A gregarious ungulate as a study model. R. Soc. Open Sci. 2020, 7, 191482. [Google Scholar] [CrossRef]
- Moreland, E.E.; Cameron, M.F.; Angliss, R.P.; Boveng, P.L. Evaluation of a ship-based unoccupied aircraft system (UAS) for surveys of spotted and ribbon seals in the Bering Sea pack ice. J. Unmanned Veh. Syst. 2015, 3, 114–122. [Google Scholar] [CrossRef]
- Christiansen, F.; Rojano-Doñate, L.; Madsen, P.T.; Bejder, L. Noise Levels of Multi-Rotor Unmanned Aerial Vehicles with Implications for Potential Underwater Impacts on Marine Mammals. Front. Mar. Sci. 2016, 3. [Google Scholar] [CrossRef]
- Erbe, C.; Parsons, M.; Duncan, A.; Osterrieder, S.K.; Allen, K. Aerial and underwater sound of unmanned aerial vehicles (UAV). J. Unmanned Veh. Syst. 2017, 5, 92–101. [Google Scholar] [CrossRef]
- Frontiers. Are Drones Disturbing Marine Mammals? Available online: https://www.frontiersin.org/news/2017/02/15/are-drones-disturbing-marine-mammals (accessed on 12 December 2024).
- Christiansen, F.; Nielsen, M.L.K.; Charlton, C.; Bejder, L.; Madsen, P.T. Southern right whales show no behavioral response to low noise levels from a nearby unmanned aerial vehicle. Mar. Mammal Sci. 2020, 36, 953–963. [Google Scholar] [CrossRef]
- Torres, L.G.; Nieukirk, S.L.; Lemos, L.; Chandler, T.E. Drone Up! Quantifying Whale Behavior From a New Perspective Improves Observational Capacity. Front. Mar. Sci. 2018, 5, 319. [Google Scholar] [CrossRef]
- Pirotta, V.; Smith, A.; Ostrowski, M.; Russell, D.; Jonsen, I.D.; Grech, A.; Harcourt, R. An Economical Custom-Built Drone for Assessing Whale Health. Front. Mar. Sci. 2017, 4, 425. [Google Scholar] [CrossRef]
- Koski, W.R.; Gamage, G.; Davis, A.R.; Mathews, T.; LeBlanc, B.; Ferguson, S.H. Evaluation of UAS for photographic re-identification of bowhead whales, Balaena mysticetus. J. Unmanned Veh. Syst. 2015, 3, 22–29. [Google Scholar] [CrossRef]
- Domínguez-Sánchez, C.A.; Acevedo-Whitehouse, K.A.; Gendron, D. Effect of drone-based blow sampling on blue whale (Balaenoptera musculus) behavior. Mar. Mammal Sci. 2018, 34, 841–850. [Google Scholar] [CrossRef]
- Fettermann, T.; Fiori, L.; Bader, M.; Doshi, A.; Breen, D.; Stockin, K.A.; Bollard, B. Behaviour reactions of bottlenose dolphins (Tursiops truncatus) to multirotor Unmanned Aerial Vehicles (UAVs). Sci. Rep. 2019, 9, 8558. [Google Scholar] [CrossRef]
- Moleón, M.; Sánchez-Zapata, J.A.; Donázar, J.A.; Revilla, E.; Martín-López, B.; Gutiérrez-Cánovas, C.; Getz, W.M.; Morales-Reyes, Z.; Campos-Arceiz, A.; Crowder, L.B.; et al. Rethinking megafauna. Proc. R. Soc. B Biol. Sci. 2020, 287, 20192643. [Google Scholar] [CrossRef]
- vanVuuren, M.; vanVuuren, R.; Silverberg, L.M.; Manning, J.; Pacifici, K.; Dorgeloh, W.; Campbell, J. Ungulate responses and habituation to unmanned aerial vehicles in Africa’s savanna. PLoS ONE 2023, 18, e0288975. [Google Scholar] [CrossRef] [PubMed]
- Bourke, E.; Raoult, V.; Williamson, J.E.; Gaston, T.F. Estuary Stingray (Dasyatis fluviorum) Behaviour Does Not Change in Response to Drone Altitude. Drones 2023, 7, 164. [Google Scholar] [CrossRef]
- Castro, J.; Borges, F.O.; Cid, A.; Laborde, M.I.; Rosa, R.; Pearson, H.C. Assessing the Behavioural Responses of Small Cetaceans to Unmanned Aerial Vehicles. Remote Sens. 2021, 13, 156. [Google Scholar] [CrossRef]
- Schroeder, N.M.; Panebianco, A. Sociability strongly affects the behavioural responses of wild guanacos to drones. Sci. Rep. 2021, 11, 20901. [Google Scholar] [CrossRef]
- Ditmer, M.A.; Vincent, J.B.; Werden, L.K.; Tanner, J.C.; Laske, T.G.; Iaizzo, P.A.; Garshelis, D.L.; Fieberg, J.R. Bears Show a Physiological but Limited Behavioral Response to Unmanned Aerial Vehicles. Curr. Biol. CB 2015, 25, 2278–2283. [Google Scholar] [CrossRef] [PubMed]
- Palomino-González, A.; Kovacs, K.M.; Lydersen, C.; Ims, R.A.; Lowther, A.D. Drones and marine mammals in Svalbard, Norway. Mar. Mammal Sci. 2021, 37, 1212–1229. [Google Scholar] [CrossRef]
- Bevan, E.; Whiting, S.; Tucker, T.; Guinea, M.; Raith, A.; Douglas, R. Measuring behavioral responses of sea turtles, saltwater crocodiles, and crested terns to drone disturbance to define ethical operating thresholds. PLoS ONE 2018, 13, e0194460. [Google Scholar] [CrossRef] [PubMed]
- Junda, J.; Greene, E.; Bird, D.M. Proper flight technique for using a small rotary-winged drone aircraft to safely, quickly, and accurately survey raptor nests. J. Unmanned Veh. Syst. 2015, 3, 222–236. [Google Scholar] [CrossRef]
- Barr, J.R.; Green, M.C.; DeMaso, S.J.; Hardy, T.B. Drone surveys do not increase colony-wide flight behaviour at waterbird nesting sites, but sensitivity varies among species. Sci. Rep. 2020, 10, 3781. [Google Scholar] [CrossRef]
- Culik, B.; Adelung, D.; Woakes, A.J. The Effect of Disturbance on the Heart Rate and Behaviour of Adélie Penguins (Pygoscelis adeliae) during the Breeding Season. In Proceedings of the Antarctic Ecosystems; Kerry, K.R., Hempel, G., Eds.; Springer Nature: Berlin/Heidelberg, Germany, 1990; pp. 177–182. [Google Scholar] [CrossRef]
- Bonter, D.N.; Zuckerberg, B.; Sedgwick, C.W.; Hochachka, W.M. Daily foraging patterns in free-living birds: Exploring the predation–starvation trade-off. Proc. R. Soc. B Biol. Sci. 2013, 280, 20123087. [Google Scholar] [CrossRef]
- Junda, J.H.; Greene, E.; Zazelenchuk, D.; Bird, D.M. Nest defense behaviour of four raptor species (osprey, bald eagle, ferruginous hawk, and red-tailed hawk) to a novel aerial intruder—A small rotary-winged drone. J. Unmanned Veh. Syst. 2016, 4, 217–227. [Google Scholar] [CrossRef]
- Turney, S.; Godin, J.G.J. To forage or hide? Threat-sensitive foraging behaviour in wild, non-reproductive passerine birds. Curr. Zool. 2014, 60, 719–728. [Google Scholar] [CrossRef]
- Gremm. Belugas and Drones: A Call for Caution. Available online: https://baleinesendirect.org/en/les-belugas-et-les-drones-un-appel-a-la-prudence/ (accessed on 11 December 2024).
- Pomeroy, P.; O’Connor, L.; Davies, P. Assessing use of and reaction to unmanned aerial systems in gray and harbor seals during breeding and molt in the UK. J. Unmanned Veh. Syst. 2015, 3, 102–113. [Google Scholar] [CrossRef]
- Colombelli-Négrel, D.; Sach, I.Z.; Hough, I.; Hodgson, J.C.; Daniels, C.B.; Kleindorfer, S. Koalas showed limited behavioural response and no physiological response to drones. Appl. Anim. Behav. Sci. 2023, 264, 105963. [Google Scholar] [CrossRef]
- Barnas, A.; Newman, R.; Felege, C.J.; Corcoran, M.P.; Hervey, S.D.; Stechmann, T.J.; Rockwell, R.F.; Ellis-Felege, S.N. Evaluating behavioral responses of nesting lesser snow geese to unmanned aircraft surveys. Ecol. Evol. 2018, 8, 1328–1338. [Google Scholar] [CrossRef] [PubMed]
- Borrelle, S.B.; Fletcher, A.T. Will drones reduce investigator disturbance to surface-nesting seabirds? Mar. Ornithol. 2017, 45, 89–94. [Google Scholar] [CrossRef]
- Pinel-Ramos, E.J.; Aureli, F.; Wich, S.; Petersen, M.F.; Dias, P.A.D.; Spaan, D. The Behavioral Responses of Geoffroy’s Spider Monkeys to Drone Flights. Drones 2024, 8, 500. [Google Scholar] [CrossRef]
- Mo, M.; Bonatakis, K. Approaching wildlife with drones: Using scientific literature to identify factors to consider for minimising disturbance. Aust. Zool. 2021, 42, 1–29. [Google Scholar] [CrossRef]
- Smith, C.A.; Tantillo, J.A.; Hale, B.; Decker, D.J.; Forstchen, A.B.; Pomeranz, E.F.; Lauber, T.B.; Schiavone, M.V.; Frohlich, K.; Lederle, P.E.; et al. A practical framework for ethics assessment in wildlife management decision-making. J. Wildl. Manag. 2024, 88, e22502. [Google Scholar] [CrossRef]
- Hodgson, J.C.; Koh, L.P. Best practice for minimising unmanned aerial vehicle disturbance to wildlife in biological field research. Curr. Biol. 2016, 26, R404–R405. [Google Scholar] [CrossRef]
- Fenwick, N.; Griffin, G.; Gauthier, C. The welfare of animals used in science: How the “Three Rs” ethic guides improvements. Can. Vet. J. 2009, 50, 523–530. [Google Scholar] [PubMed Central]
- Fernandes, M.R.; Pedroso, A.R. Animal experimentation: A look into ethics, welfare and alternative methods. Rev. Assoc. Medica Bras. (1992) 2017, 63, 923–928. [Google Scholar] [CrossRef]
- Bartlett, B.; Santos, M.; Dorian, T.; Moreno, M.; Trslic, P.; Dooly, G. Real-Time UAV Surveys with the Modular Detection and Targeting System: Balancing Wide-Area Coverage and High-Resolution Precision in Wildlife Monitoring. Remote Sens. 2025, 17, 879. [Google Scholar] [CrossRef]
- Barnas, A.F.; Chabot, D.; Hodgson, A.J.; Johnston, D.W.; Bird, D.M.; Ellis-Felege, S.N. A standardized protocol for reporting methods when using drones for wildlife research. J. Unmanned Veh. Syst. 2020, 8, 89–98. [Google Scholar] [CrossRef]
- Hvala, A.; Rogers, R.M.; Alazab, M.; Campbell, H.A. Supplementing aerial drone surveys with biotelemetry data validates wildlife detection probabilities. Front. Conserv. Sci. 2023, 4. [Google Scholar] [CrossRef]
- Costello, M.J.; Wieczorek, J. Best practice for biodiversity data management and publication. Biol. Conserv. 2014, 173, 68–73. [Google Scholar] [CrossRef]
- Binley, A.D.; Vincent, J.G.; Rytwinski, T.; Soroye, P.; Bennett, J.R. Making the most of existing data in conservation research. Perspect. Ecol. Conserv. 2024, 22, 122–128. [Google Scholar] [CrossRef]
- Maalouf, G.; Meier, K.; Richardson, T.; Guerin, D.; Watson, M.; Schultz Lundquist, U.P.; Afridi, S.; Rolland, E.G.; Hundevadt Jepsen, J.; Njoroge, W.; et al. Insights into Safe and Scalable BVLOS UAS Operations from Kenya’s Ol Pejeta Conservancy. In Proceedings of the 2025 International Conference on Unmanned Aircraft Systems (ICUAS), Charlotte, NC, USA, 14–17 May 2025. [Google Scholar]
- JARUS—Joint Authorities for Rulemaking on Unmanned Systems. Available online: http://jarus-rpas.org/ (accessed on 11 March 2025).
- SORA Workshop: From Version 2.0 to 2.5—Hybrid|EASA. Available online: https://www.easa.europa.eu/en/newsroom-and-events/events/sora-workshop-version-20-25 (accessed on 11 March 2025).
- Bakirci, M. Enhancing vehicle detection in intelligent transportation systems via autonomous UAV platform and YOLOv8 integration. Appl. Soft Comput. 2024, 164, 112015. [Google Scholar] [CrossRef]
- Rolland, E.; Grøntved, K.; Laporte-Devylder, L.; Kline, J.; Lundquist, U.; Christensen, A. Drone Swarms for Animal Monitoring: A Method for Collecting High-Quality Multi-Perspective Data. In Proceedings of the 15th Annual International Micro Air Vehicle Conference and Competition, Bristol, UK, 16–20 September 2024; Paper no. IMAV2024-38. Richardson, T., Ed.; University of Bristol: Bristol, UK; pp. 316–323. [Google Scholar]
- Rolland, E.; Laporte-Devylder, L.; Christensen, A. Advancing Wildlife Monitoring in Gregarious Species with Drone Swarms. In Proceedings of the Distributed Computing and Artificial Intelligence, 21st International Conference, Jeju, Republic of Korea, 21–23 June 2024; Springer: Berlin/Heidelberg, Germany, 2024. [Google Scholar]
- Bakirci, M. A novel swarm unmanned aerial vehicle system: Incorporating autonomous flight, real-time object detection, and coordinated intelligence for enhanced performance. Trait. Signal 2023, 40, 2063–2078. [Google Scholar] [CrossRef]
- Meier, K.; Richards, A.; Watson, M.; Johnson, C.; Hine, D.; Richardson, T.; Maalouf, G. WildBridge: Conservation Software for Animal Localisation using Commercial Drones. In Proceedings of the 15th Annual International Micro Air Vehicle Conference and Competition, Bristol, UK, 16–20 September 2024; Paper no. IMAV2024-39. Richardson, T., Ed.; University of Bristol: Bristol, UK, 2024; pp. 324–333. [Google Scholar]


| Study | Species Studied | Drone Used | Study Context | UAV Flight Altitude | Observed Impact | Recommendations |
|---|---|---|---|---|---|---|
| Assessing the Disturbance Potential of Small Unoccupied Aircraft Systems (UAS) on Gray Seals at breeding colonies in Nova Scotia, Canada (2018) [32] | Gray seals (Halichoerus grypus) | Fixed-wing UAS (eBee) | To assess the disturbance of UAS during population surveys | 75–85 m | No significant changes in seal behavior (counts, posture, or movements) | Lower acoustics at higher altitudes found suitable for minimal invasive surveys. |
| First Guidelines and Suggested Best Protocol for Surveying African Elephants using a drone (2021) [33] | African elephants (Loxodonta africana) | DJI Mavic Pro Platinum | Investigating behavioral responses to drone speed, angle of approach, and initial altitude | Combination of flights: 35 m, 50 m, and 100 m with 2 m/s, 4 m/s, and 6 m/s, and approach angle of 45° and 90° | 90° approach angle and higher speed reported to influence elephant behavior | Higher speed and high approach angles should be avoided to induce less stress on animals. |
| Przewalski’s Horses Responses to Unmanned Aerial Vehicles Flights under Semireserve Conditions: Conservation Implication (2021) [34] | Przewalski’s horses (Equus ferus przewalskii) | DJI Mavic 2 Zoom | Behavioral responses of Przewalski’s horses to different UAV flight altitudes | 1 m to 52 m | Alert and run-away responses varied by age and gender | Optimizing flights according to the biological state of the animal to minimize disturbance. |
| Dolphin Behavioral Responses to Uncrewed Aerial Systems as a Function of Exposure, Height, and Type (2023) [35] | Bottlenose dolphins (Tursiops truncatus) | DJI Mini 2, DJI Mavic 2 Enterprise, DJI Inspire 2, DJI Mini 3 Pro, DJI Avata, SplashDrone 4, PHASM fixed-wing UAS | Behavioral responses of bottlenose dolphins to different UAS types and heights | Hovering at 300 ft and descending to 20 ft (varied by UAS) | Larger, noisier UAS induced higher responses | Avoid lower drone flight altitudes |
| Fly with Care: Belugas Show Evasive Responses to Low Altitude Drone Flights (2023) [36] | Belugas (Delphinapterus leucas) | DJI Phantom 4, Phantom 4 Pro | Impact of drones on endangered St. Lawrence belugas | 16.9 m to 124.9 m | Evasive reactions occurred at low-altitude flights. Larger groups were more likely to show avoidance responses. | Recommended flight altitude: >30 m. Group size also influences the increased alertness at lower altitudes. |
| Impacts of Drone Flight Altitude on Marsh Bird Behaviors and Species Identification of Marsh Birds in Florida (2023) [37] | Marsh birds, including passerines, wading birds, and waterfowl | DJI Mavic 2 Zoom | Evaluate the effects of drone altitude on marsh bird behavior | 12 m, 30 m, 61 m, and 91 m | Minimal behavioral reactions at 12 m and 30 m | Lower altitudes can be opted only if animals shows no disturbance. |
| How low can you go? Exploring impact of drones on haul out behaviour of harbour—and grey seals (2024) [38] | Harbor seals (Phoca vitulina), Grey seals (Halichoerus grypus) | DJI Phantom 4 Pro, Autel EVO II RTK | Impact of varying drone flight altitudes and approaches on harbor and grey seal behavior | 70 m to 10 m, descending by 5–10 m intervals | Increased vigilance and displacement at altitudes < 30 m | Direct overheads path should be avoided with lower speeds to minimize disturbance. |
| Study | Species Studied | Drone Used | Study Context | UAV Flight Altitude | Observed Impact | Recommendations |
|---|---|---|---|---|---|---|
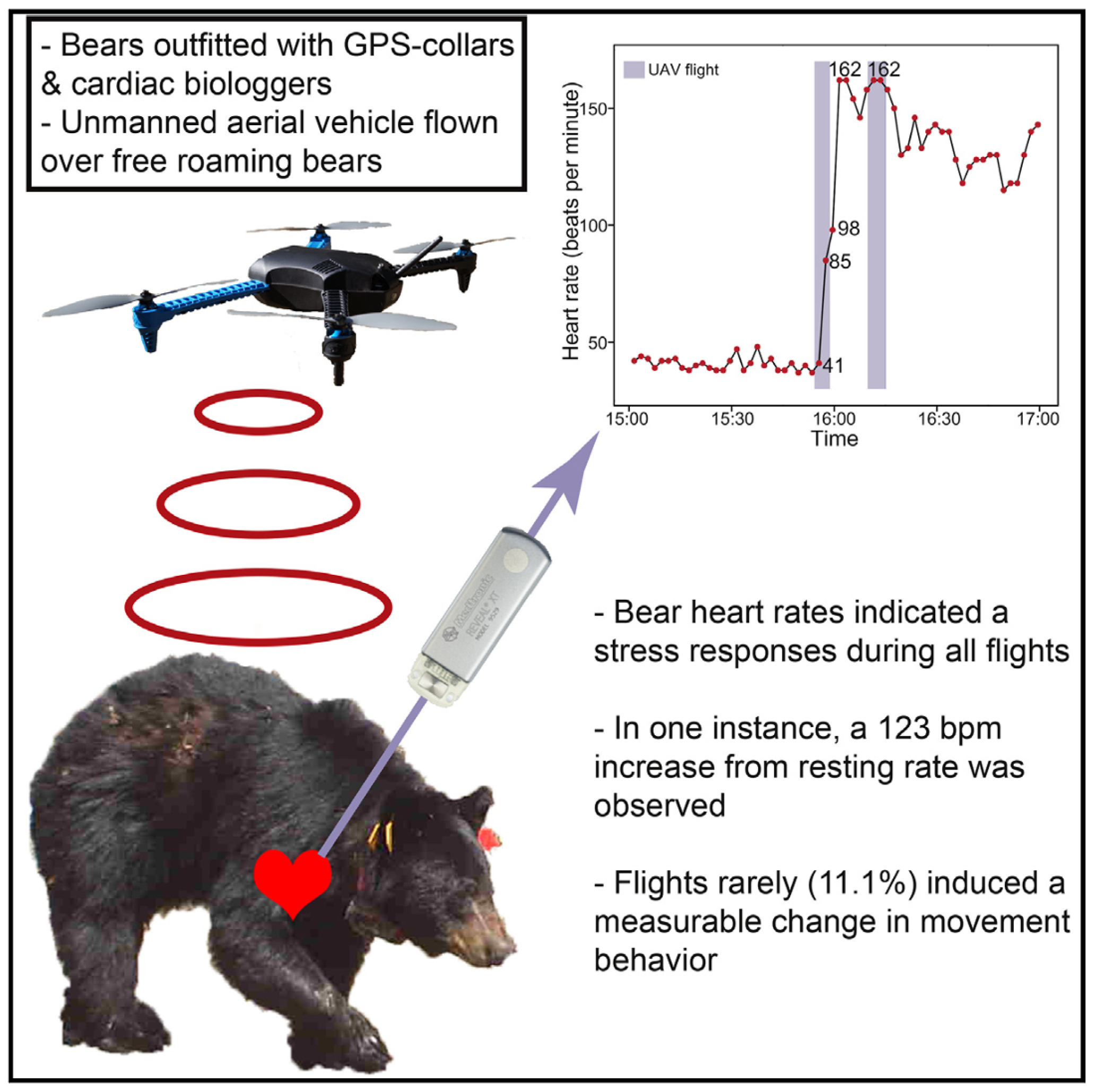
| Bears Show a Physiological but Limited Behavioral Response to Unmanned Aerial Vehicles (2015) [79] | American black bear (Ursus americanus) | 3D Robotics Quadcopter | Assessing physiological and behavioral responses to drone flights | 20 m to 43 m | No behavioral responses (movement or avoidance) but increased stress (heart rate of 123 bpm) | Developing frameworks that consider species’ vulnerability and additional stress. |
| Fright or Flight? Behavioral Responses of Kangaroos to Drone-Based Monitoring (2019) [29] | Eastern grey kangaroo (Macropus giganteus) | DJI Phantom 3 Advanced | Assessing vigilance behavior to drone altitude and flight characteristics | 30 m to 120 m | Flight responses most frequent at 30 m altitude | Minimum flight altitude of 60 m is recommended to minimize disturbance. |
| Koalas Showed Limited Behavioral Response and No Physiological Response to Drones (2023) [90] | Koalas (Phascolarctos cinereus) | DJI Mavic 2 Pro | Assessing behavioral and physiological to drones | 15 m above the enclosure | Short-term increase in vigilance but no significant change in heart rate or breathing rate | Further research required on the behavioral and physiological responses. |
| Evaluating Behavioral Responses of Nesting Lesser Snow Geese to Unmanned Aircraft Surveys (2018) [91] | Lesser snow geese (Anser caerulescens caerulescens) | Fixed-wing Trimble UX5 | Measuring behavioral responses of nesting snow geese | 75 m, 100 m, and 120 m above ground | Increased vigilance (head cocking and scanning) during flights | Closer proximity may be acceptable only if it ensures minimal stress to the species. |
| Will Drones Reduce Investigator Disturbance to Surface-Nesting Birds? (2017) [92] | Various surface-nesting seabirds (e.g., gulls, penguins) | DJI Phantom, Trimble UX5, other off-the-shelf drones | Assessing drone-based monitoring disturbance in surface-nesting seabirds | 50 m to 120 m depending on species | Species-specific responses varied. Visual predator-like flight patterns (e.g., vertical approaches) increased reactions. | Avoiding direct overhead flight patterns. |
| Behavioral Responses of Geoffroy’s Spider Monkeys to Drone Flights (2024) [93] | Geoffroy’s spider monkeys (Ateles geoffroyi) | Mavic 2 Enterprise Advanced | Assessing drone flight influence on spider monkey | 35 m, 50 m flight heights | Minimal changes observed in behaviors | More research is required to explore long-term behavioral impact and habituation to drones. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Afridi, S.; Laporte-Devylder, L.; Maalouf, G.; Kline, J.M.; Penny, S.G.; Hlebowicz, K.; Cawthorne, D.; Lundquist, U.P.S. Impact of Drone Disturbances on Wildlife: A Review. Drones 2025, 9, 311. https://doi.org/10.3390/drones9040311
Afridi S, Laporte-Devylder L, Maalouf G, Kline JM, Penny SG, Hlebowicz K, Cawthorne D, Lundquist UPS. Impact of Drone Disturbances on Wildlife: A Review. Drones. 2025; 9(4):311. https://doi.org/10.3390/drones9040311
Chicago/Turabian StyleAfridi, Saadia, Lucie Laporte-Devylder, Guy Maalouf, Jenna M. Kline, Samuel G. Penny, Kasper Hlebowicz, Dylan Cawthorne, and Ulrik Pagh Schultz Lundquist. 2025. "Impact of Drone Disturbances on Wildlife: A Review" Drones 9, no. 4: 311. https://doi.org/10.3390/drones9040311
APA StyleAfridi, S., Laporte-Devylder, L., Maalouf, G., Kline, J. M., Penny, S. G., Hlebowicz, K., Cawthorne, D., & Lundquist, U. P. S. (2025). Impact of Drone Disturbances on Wildlife: A Review. Drones, 9(4), 311. https://doi.org/10.3390/drones9040311