
Who Killed the Small Mammals of Ittenheim (Northeastern France)? An Integrative Approach and New Taphonomic Data for Investigating Bone Assemblages Accumulated by Small Carnivores


Abstract
:1. Introduction
2. Material and Methods
2.1. Description of Sites
2.2. Taxonomic Identification
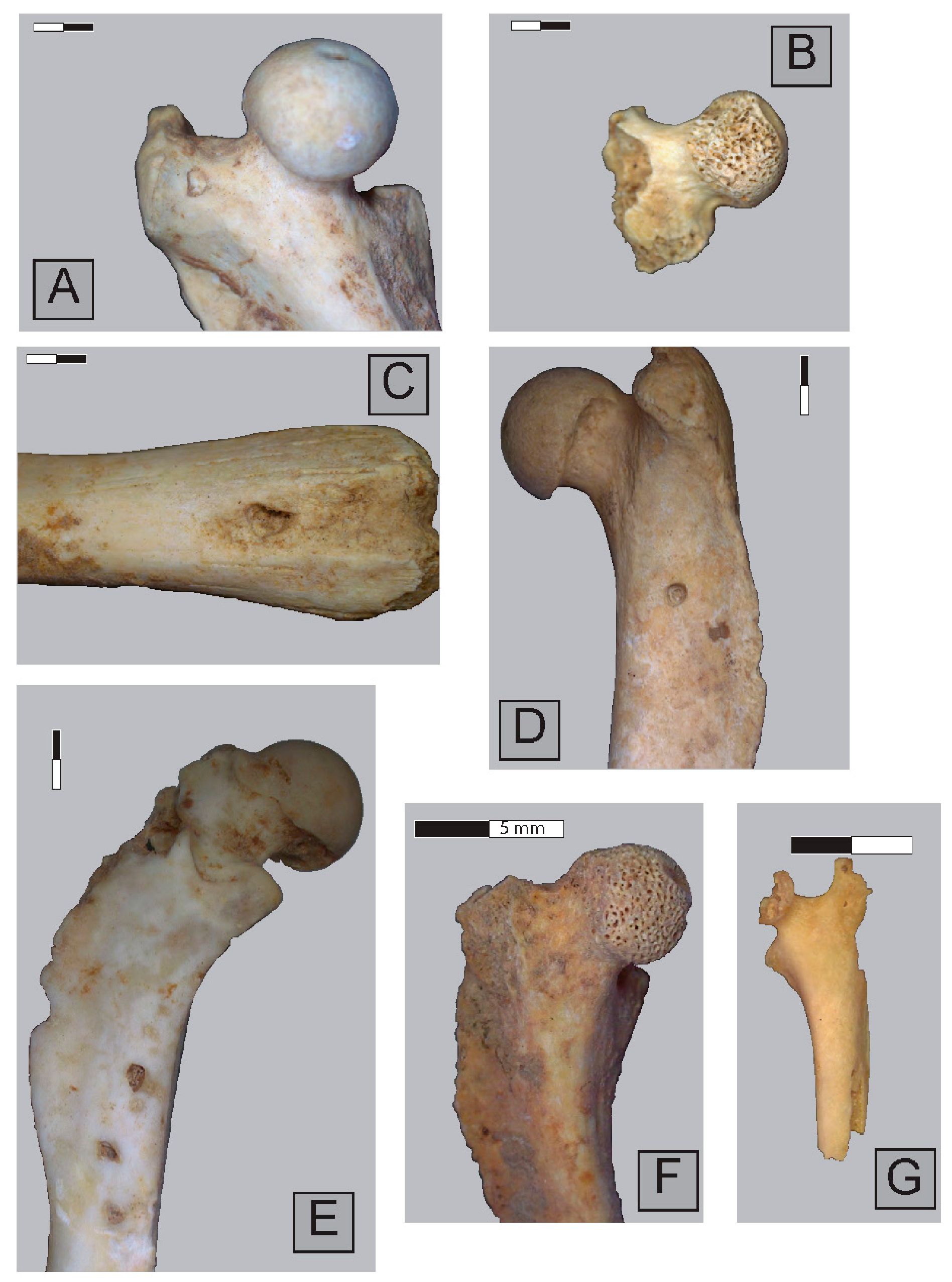
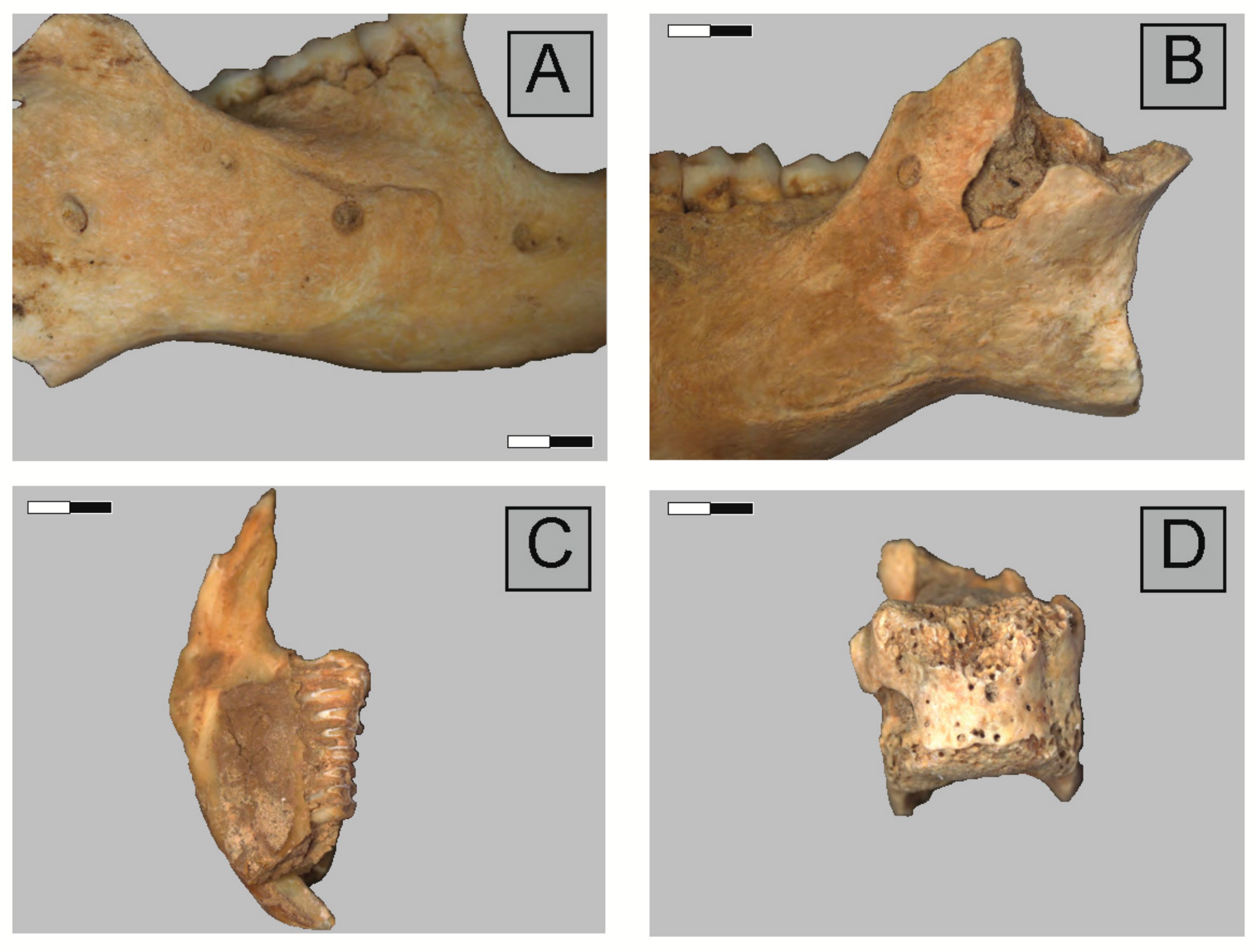
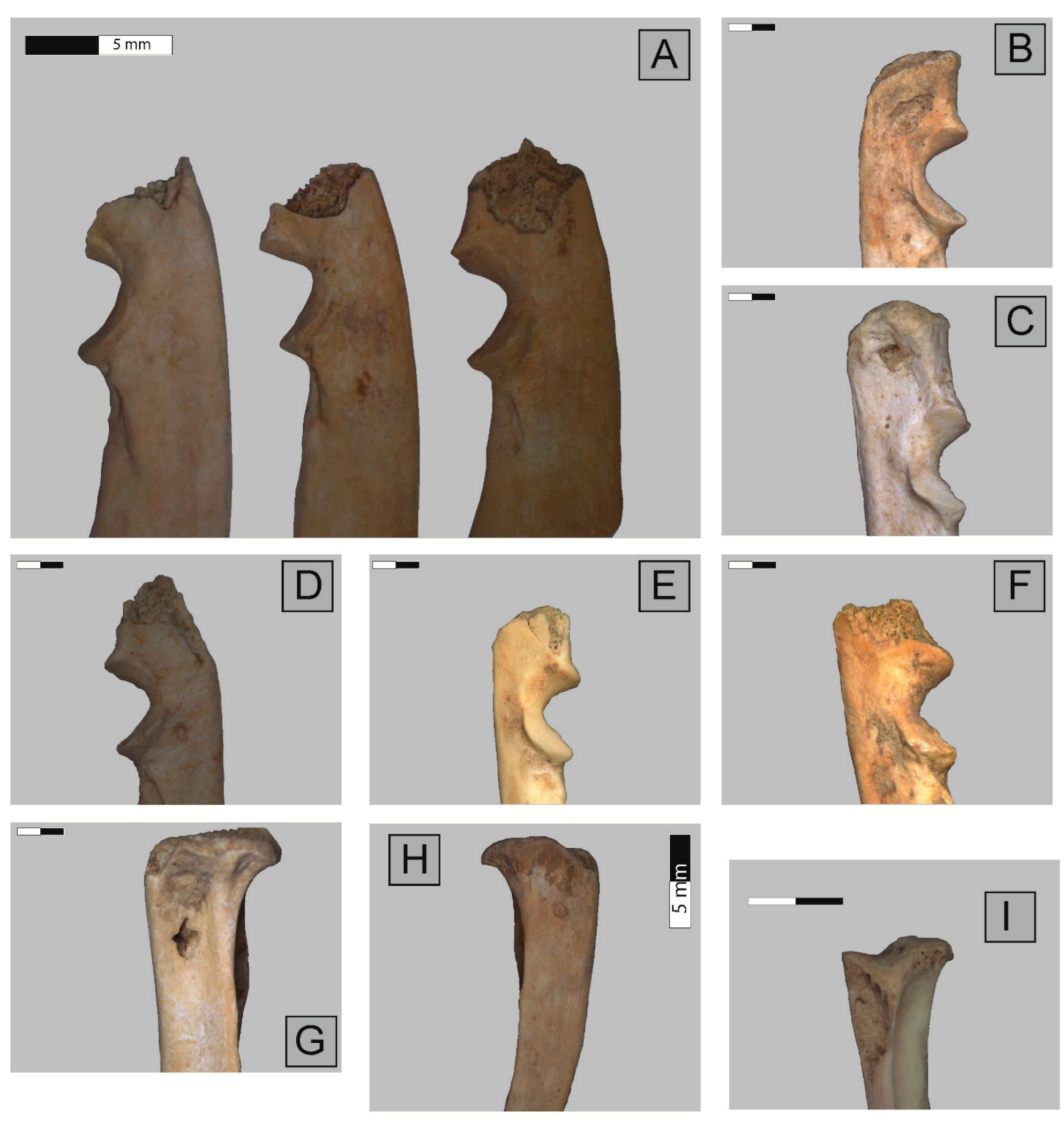
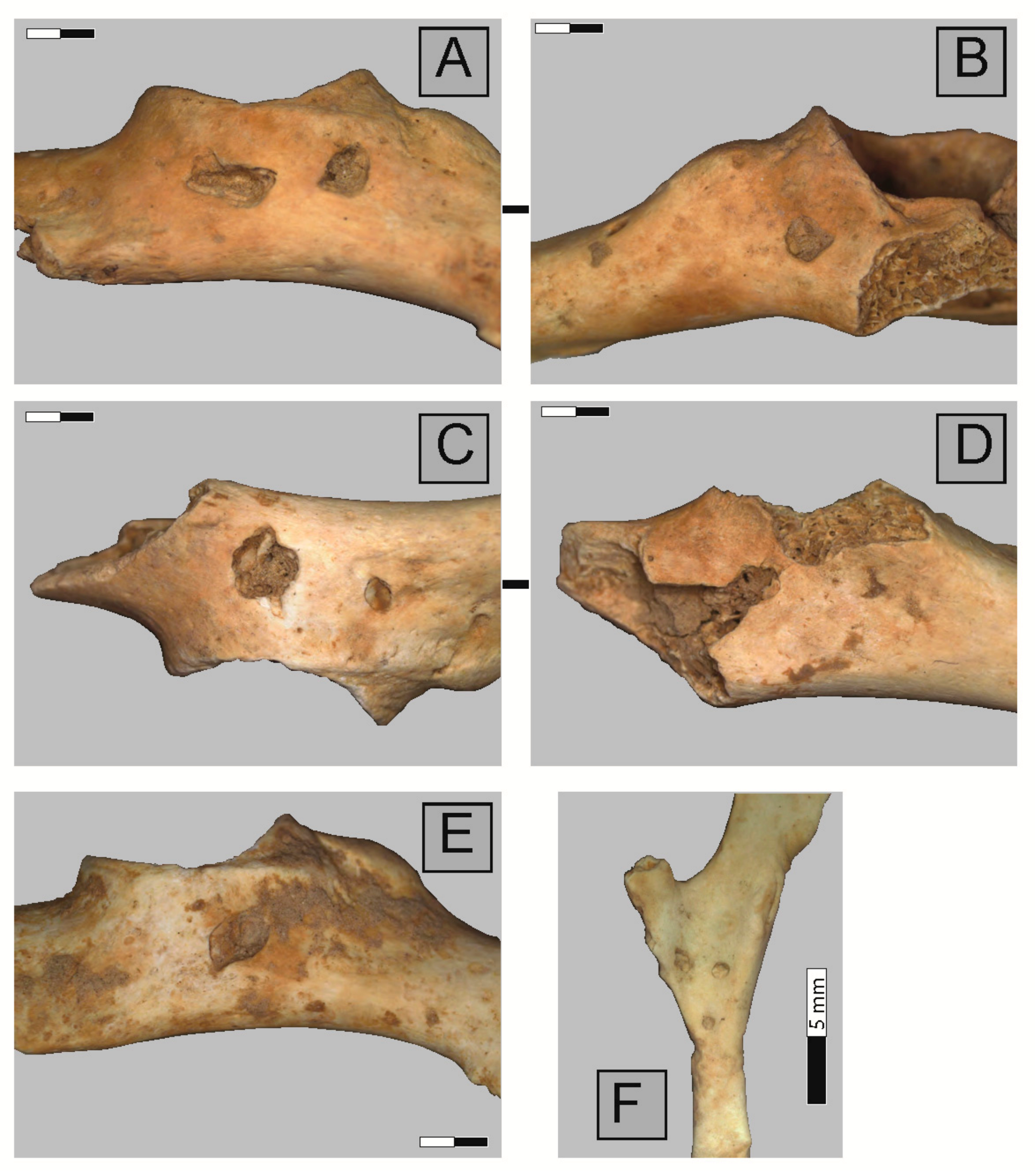
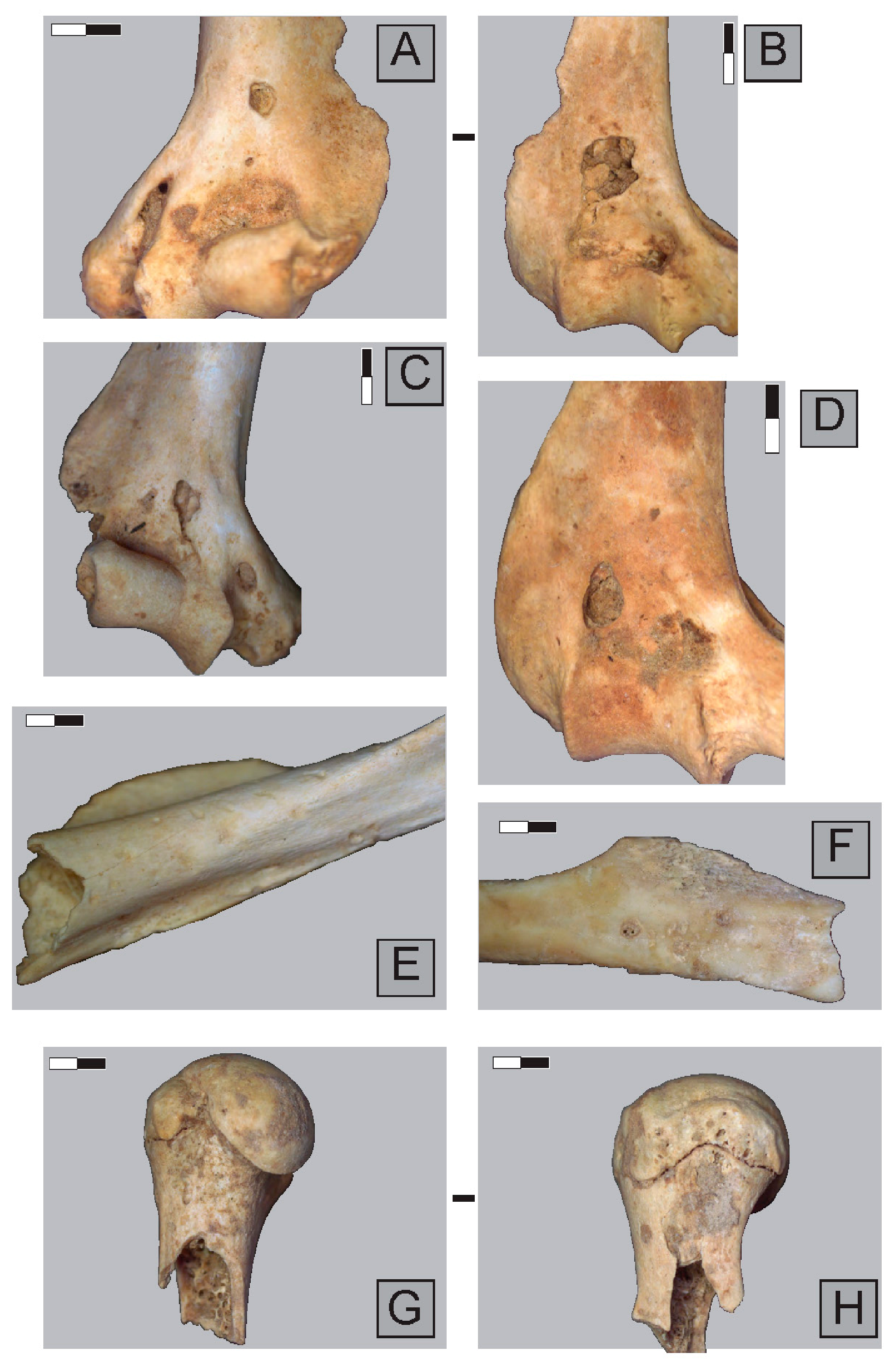
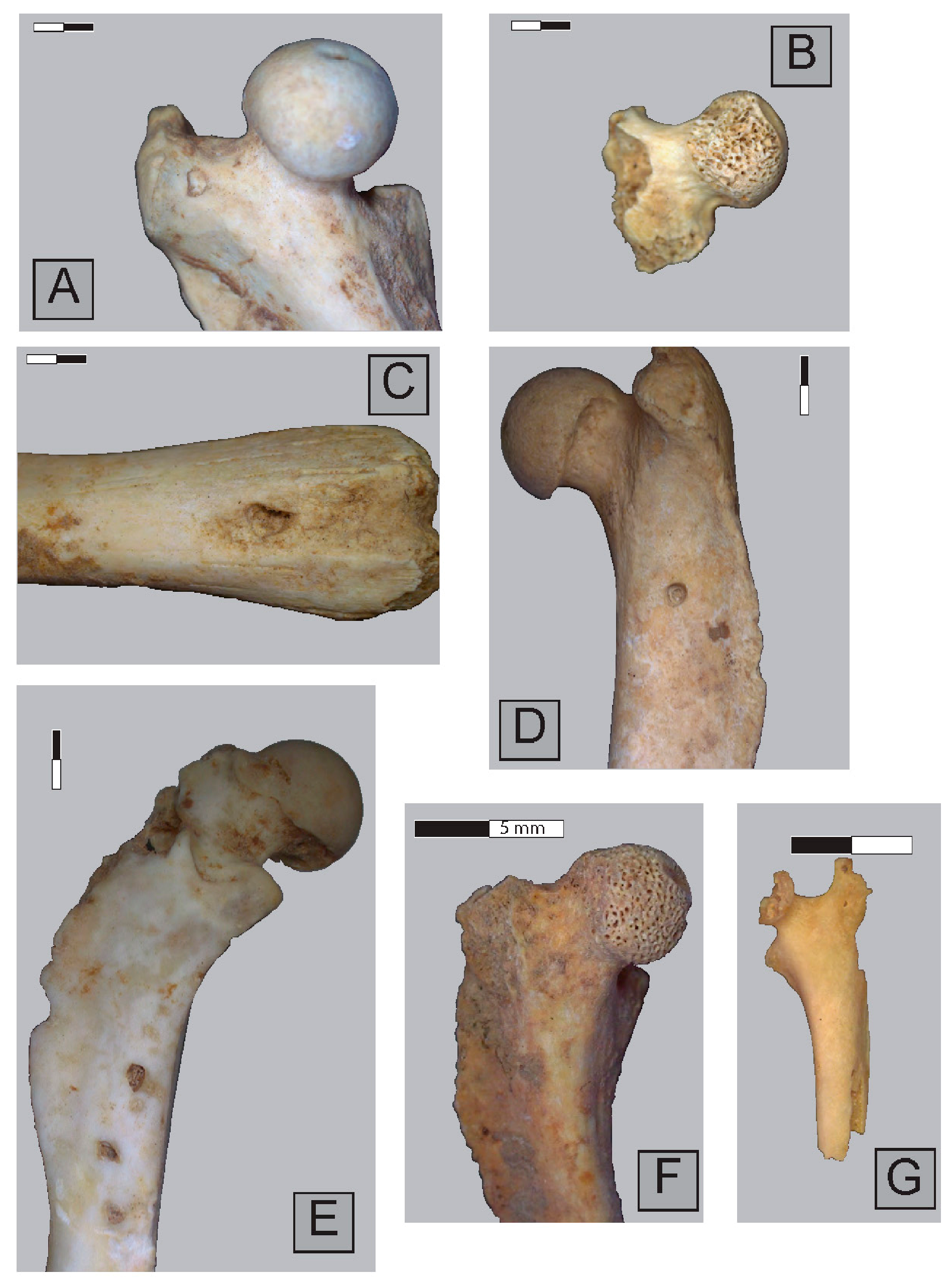
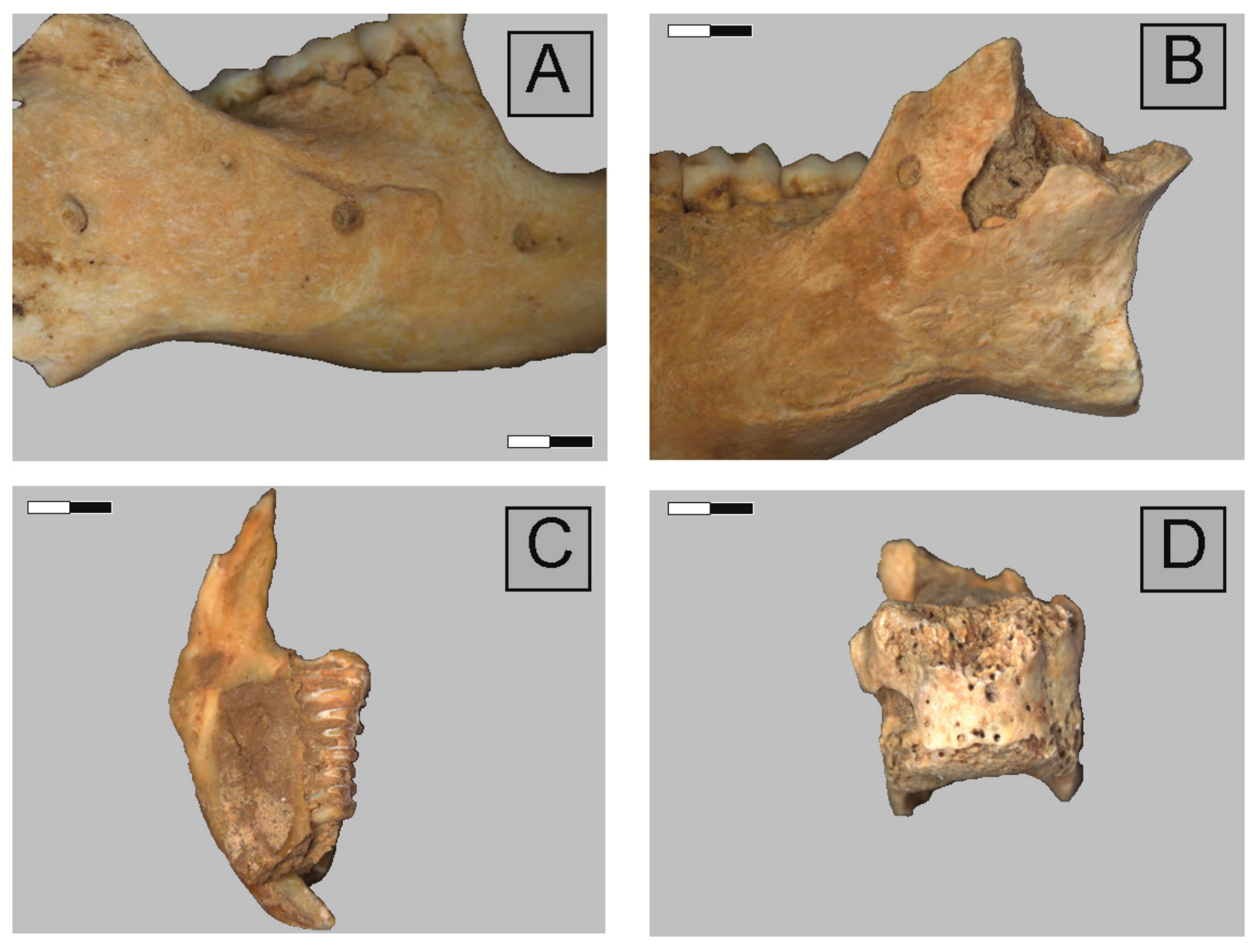
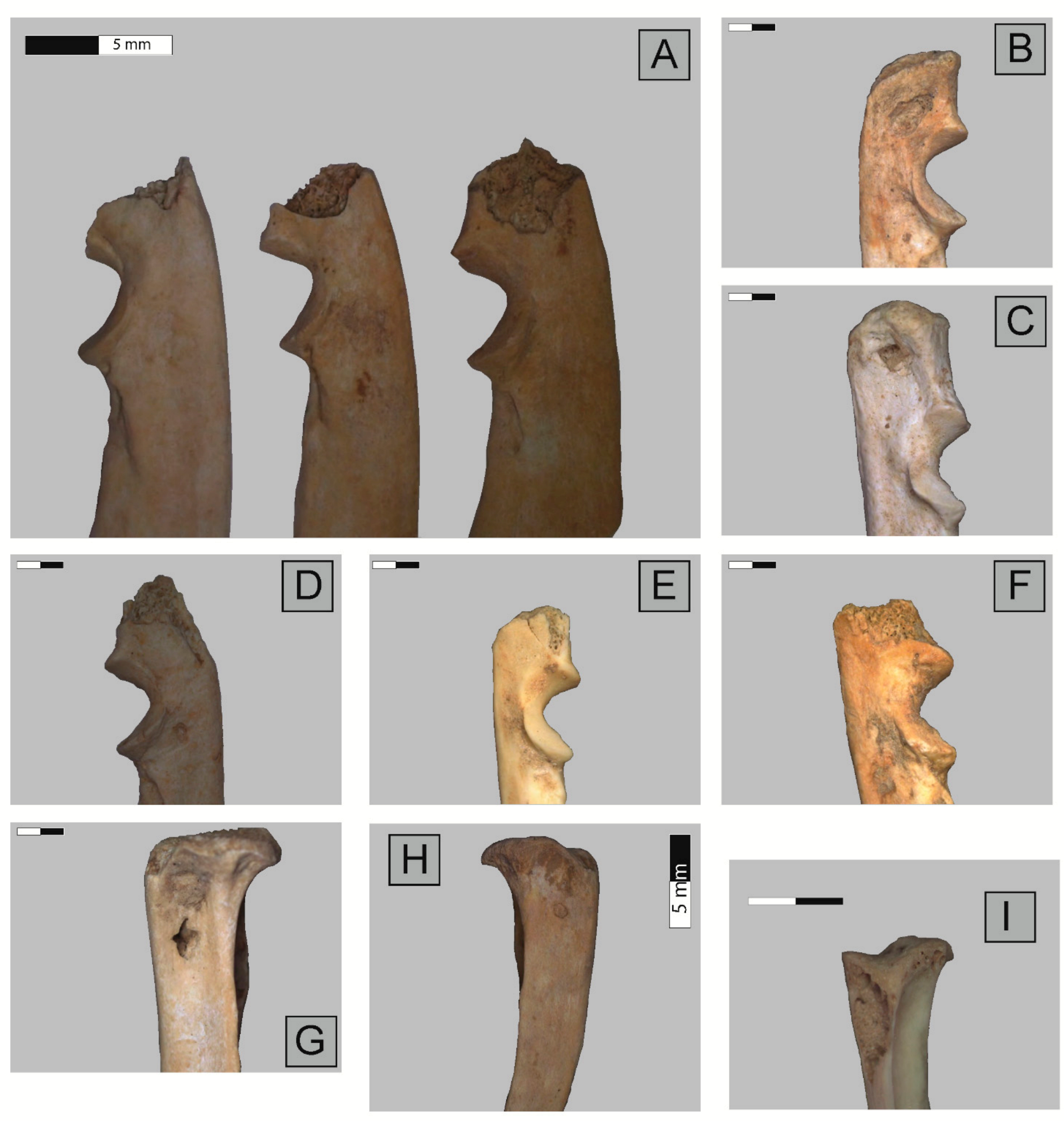
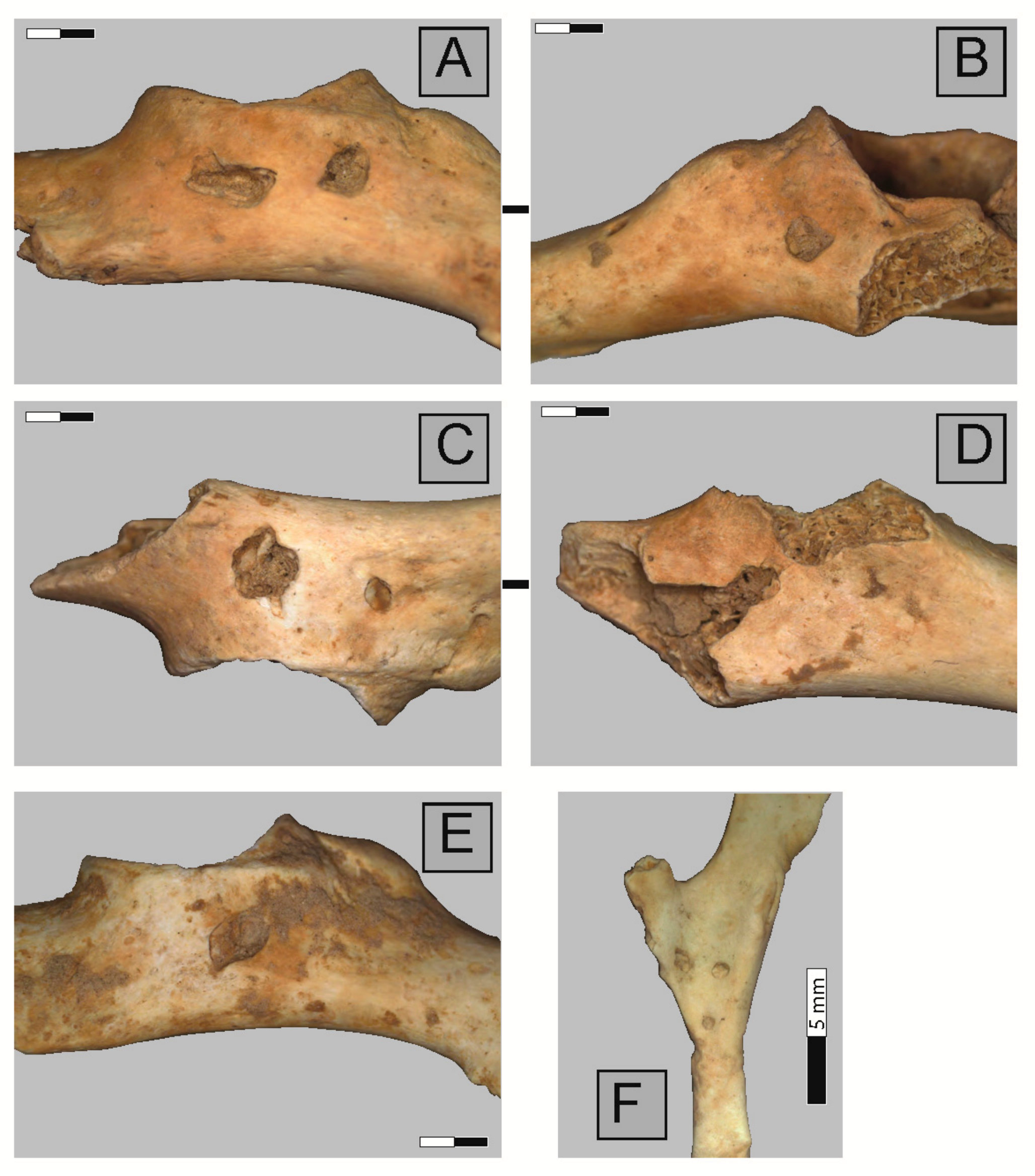
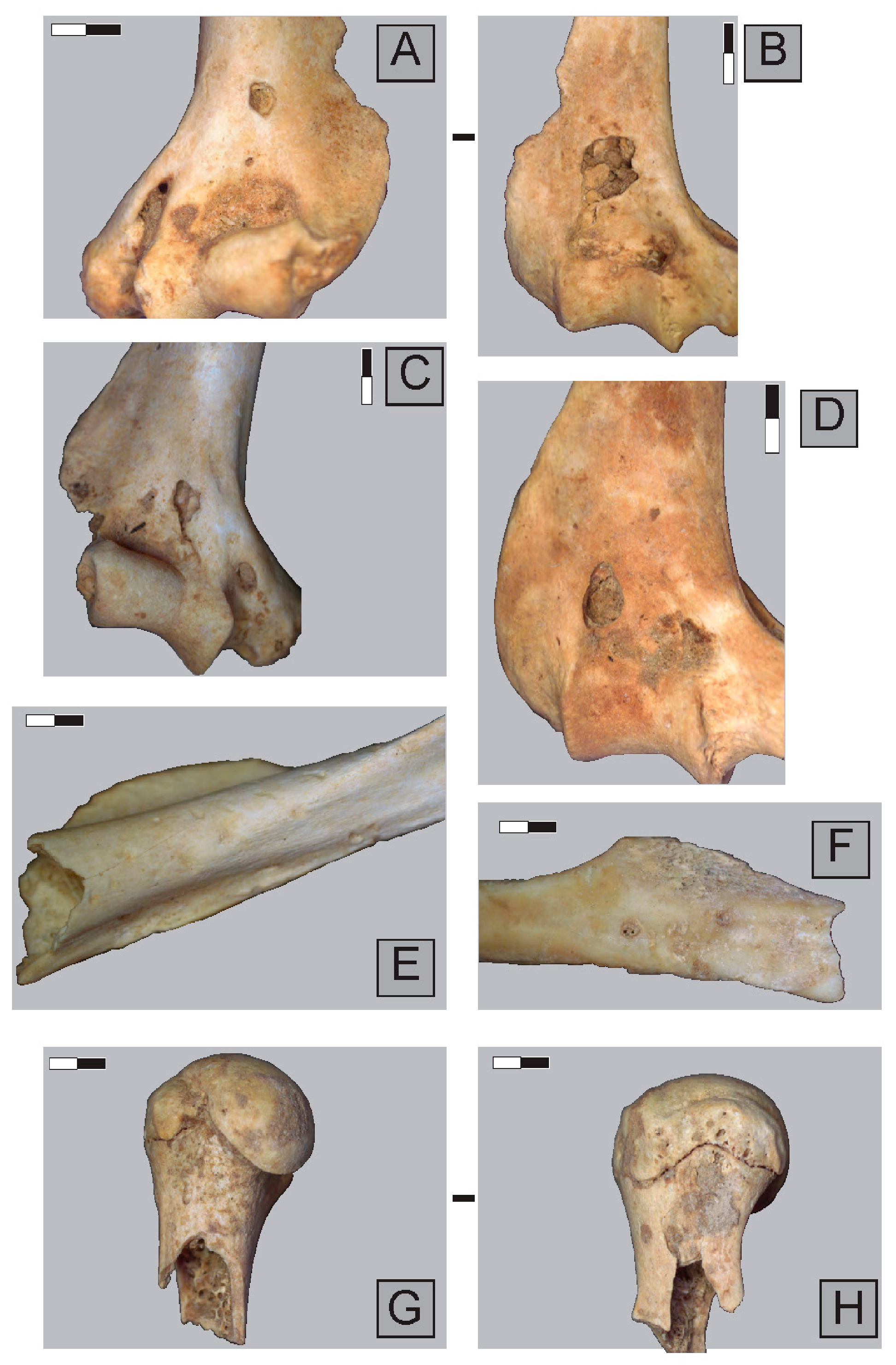
2.3. Anatomical Representation, Bone Breakage, and Surface Modifications
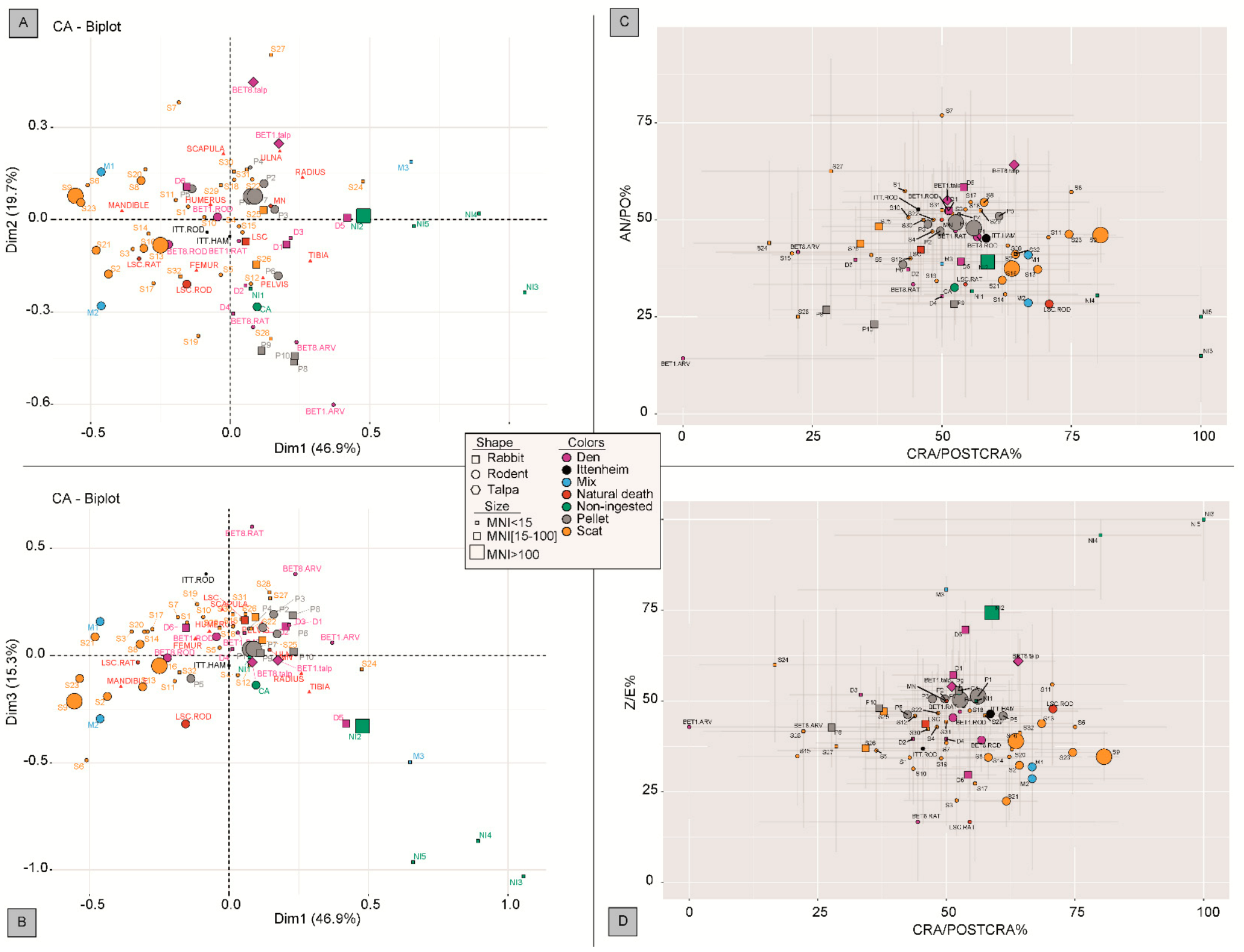
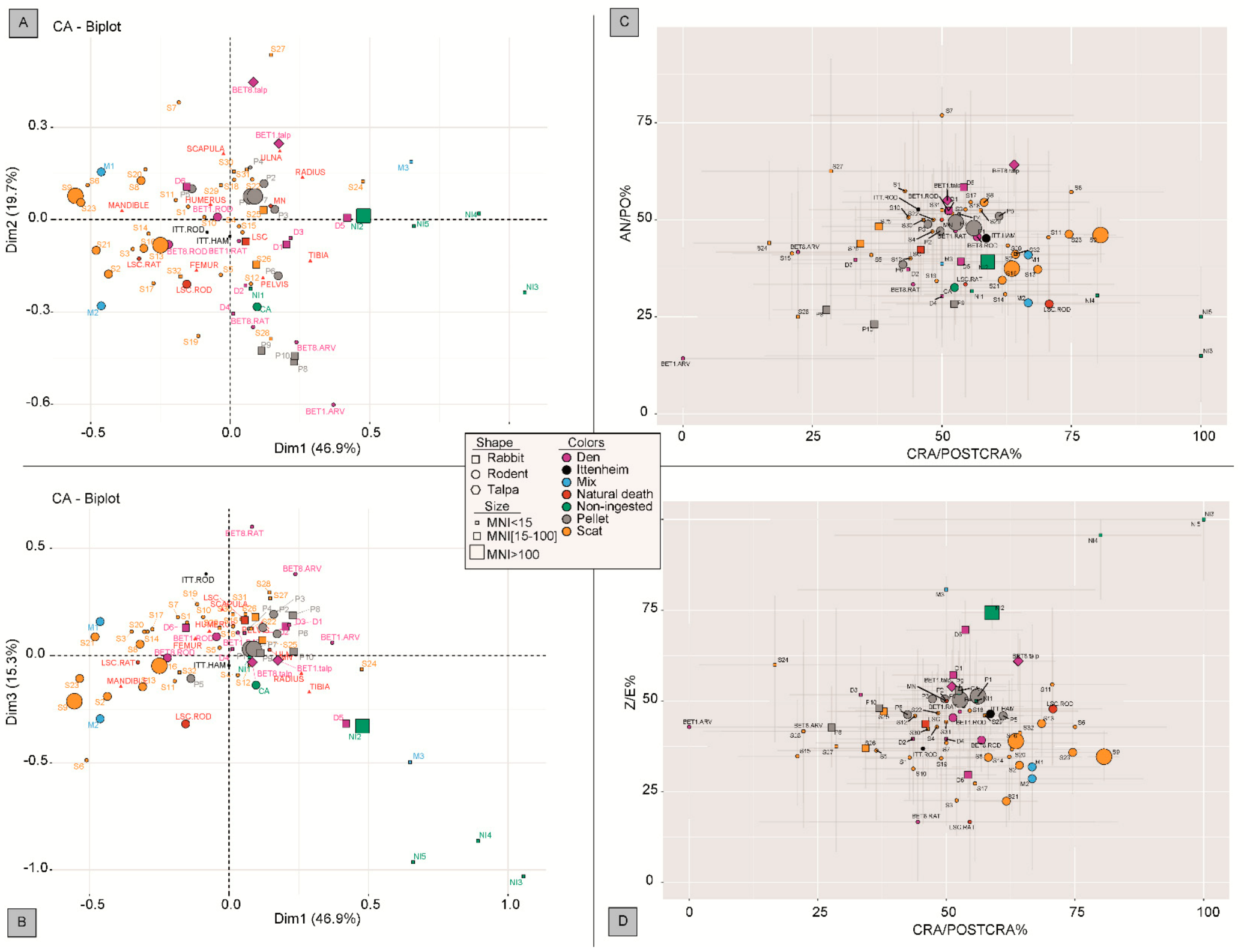
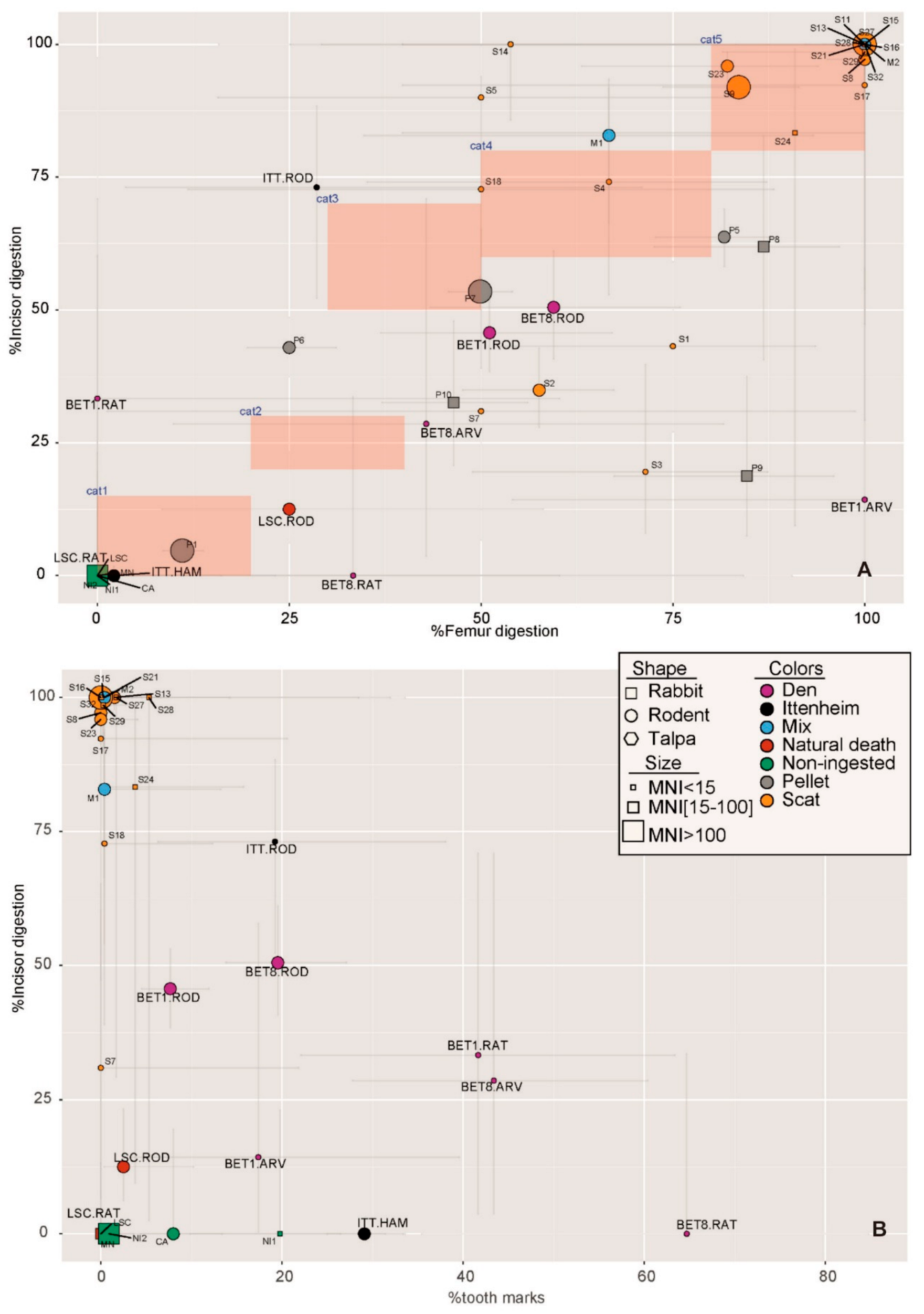
2.4. Statistical Analyses
3. Results
3.1. Ittenheim
3.2. Bettant I, a Meles/Vulpes den
3.3. Bettant VIII, a Meles/Vulpes den
3.4. Les Six Chemins, a Rabbit Warren
3.5. Comparison with Other Carnivore Reference Collections
4. Discussion
4.1. An Integrative Approach for Reconstructing the Taphonomic History of the Ittenheim Accumulation
4.2. Implications for New Reference Collections and Fossil Material
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mellet, J.S. Scatological origin of microvertebrate fossil accumulation. Science 1974, 185, 349–350. [Google Scholar] [CrossRef] [PubMed]
- Korth, W.W. Taphonomy of Microvertebrate fossil assemblages. Ann. Carnegie Mus. 1979, 48, 235–285. [Google Scholar]
- Andrews, P. Owls, Caves and Fossils; Natural History Museum Publication: London, UK, 1990. [Google Scholar]
- Denys, C. Introduction générale. Taphonomie des microvertébrés: Méthodologie et état de l’art: Micromammifères, oiseaux et problèmes taphonomiques. Artefacts 1994, 9, 195–206. [Google Scholar]
- Lyman, R.L. Vertebrate Taphonomy; Cambridge University Press: Cambridge, MA, USA, 1994. [Google Scholar]
- Stahl, P.W. The recovery and interpretation of microvertebrate bone assemblages from archaeological contexts. J. Archaeol. Method Theory 1996, 3, 31–75. [Google Scholar] [CrossRef]
- Lyman, R.L. Actualistic neotaphonomic research on bone modifying animal species: An analysis of the literature. Palaios 2018, 33, 542–554. [Google Scholar] [CrossRef]
- Shipman, P.; Walker, A. Bone-collecting by harvesting ants. Paleobiology 1980, 6, 496–502. [Google Scholar] [CrossRef]
- Brain, C. Porcupines as bone collectors in African caves. In The Hunters or the Hunted; University of Chicago Press: Chicago, IL, USA, 1981; pp. 109–117. [Google Scholar]
- Leoni, F.; Boudadi-Maligne, M.; Mallye, J.B.; Beauval, C.; Faivre, J.P. Renards chapardeurs et amas de boucherie en pleine nature. Quantification des modifications et incidence sur les analyses archéozoologiques. In Relations Hommes—Canidés de la Préhistoire aux Périodes Modernes; Boudadi-Maligne, M., Mallye, J.-B., Eds.; Ausonius éditions; Collection DAN@: Pessac, France, 2020; pp. 69–94. [Google Scholar]
- Stahl, P.W. On small mammal remains in archaeological context. Am. Antiq. 1982, 47, 822–829. [Google Scholar] [CrossRef]
- Fiedler, L.A. Rodents as a food source. In Proceedings of the Fourteenth Vertebrate Pest Conference, Sacramento, CA, USA, 6–8 March 1990; p. 30. [Google Scholar]
- Simonetti, J.A.; Cornejo, L.E. Archaeological evidence of rodent consumption in central Chile. Lat. Am. Antiq. 1991, 2, 92–96. [Google Scholar] [CrossRef]
- Henshilwood, C.S. Identifying the collector: Evidence for human processing of the Cape dune mole-rat, Bathyergus suillus, from Blombos Cave, southern Cape, South Africa. J. Archaeol. Sci. 1997, 24, 659–662. [Google Scholar] [CrossRef]
- Stiner, M.C.; Munro, N.D.; Surovell, T. The Tortoise and the Hare. Small-Game Use, the Broad Spectrum Revolution, and Paleolithic Demography. Curr. Anthropol. 2000, 41, 39–79. [Google Scholar] [CrossRef]
- Cochard, D.; Brugal, J.-P. Importance des fonctions de sites dans les accumulations paléolithiques de léporidés. In Petits Animaux et Sociétés Humaines. Du Complément Alimentaire aux Ressources Utilitaires, Actes Des XXIVème Rencontres Internationales D’archéologie et D’histoire d’Antibes; Brugal, J.-P., Desse, J., Eds.; APDCA: Antibes, France, 2004; pp. 217–230. [Google Scholar]
- Armstrong, A. Small mammal utilization by Middle Stone Age humans at Die Kelders Cave 1 and Pinnacle Point Site 5-6, Western Cape Province, South Africa. J. Hum. Evol. 2016, 101, 17–44. [Google Scholar] [CrossRef] [Green Version]
- Mallye, J.-B.; Laroulandie., V.; Avec la Collaboration de Bonnet-Jacquement, P.; Langlais, M.; Man-Estier, E.; Vercoutère, C.; Paillet, P. L’utilisation des petits gibiers au Tardiglaciaire: Le cas de Rochereil (collection Jude, Dordogne). In Proceedings of the Animal Symbolisé—Animal Exploité, Du Paléolithique à la Protohistoire, Actes du 141eme Congrès du CTHS, Rouen, Paris, France, 11–15 April 2016; Costamagno, S.C., Dupont, O., Dutour, L., Gourichon, D.V., Eds.; CTHS: Cherrybrook, Australia, 2018. [Google Scholar]
- Pelletier, M.; Desclaux, E.; Brugal, J.P.; Texier, P.J. The exploitation of rabbits for food and pelts by last interglacial Neandertals. Quat. Sci. Rev. 2019, 224, 105972. [Google Scholar] [CrossRef]
- Pelletier, M.; Brugal, J.P.; Cochard, D.; Lenoble, A.; Mallye, J.B.; Royer, A. Identifying fossil rabbit warrens: Insights from a taphonomical analysis of a modern warren. J. Archaeol. Sci. Rep. 2016, 10, 331–344. [Google Scholar] [CrossRef]
- Mondini, M. Taphonomic action of Foxes in Puna Rockshelters: A case study in Antofagasta de la Sierra (Province of Catamarca, Argentina). In Ethnoarchaeology of Andean South America; International Monographs in Prehistory. Ethnoarchaeology Series; Kuznar, L.A., Ed.; Berghahn Books: New York, NY, USA, 2001; Volume 4, pp. 266–295. [Google Scholar]
- Mondini, M. Accumulation of small and large vertebrates by carnivores in Andean South America. In Petits Animaux et Sociétés Humaines. Du Complément Alimentaire aux Ressources Utilitaires. XXIVè Rencontres Internationales D’archéologie et D’histoire d’Antibes; Brugal, J.-P., Desse, J., Eds.; APDCA: Antibes, France, 2004; pp. 483–487. [Google Scholar]
- Mondini, M. Use of Rockshelters by carnivores in the Puna. Implications for hunter-gatherer archaeology. Before Farming 2005, 2, 1–25. [Google Scholar] [CrossRef]
- Denys, C.; Cochard, D. Taphonomie des restes ingérés par les petits carnivores. In TaphonomieS; Brugal, J.P., Ed.; EAC, Editions des Archives Contemporaines: Paris, France, 2017; pp. 369–388. [Google Scholar]
- Lloveras, L. Taphonomy of carnivores: Understanding archaeological small prey accumulations. In Themes in Old World Zooarchaeology, From the Mediterranean to the Atlantic; Albraella, U., Detry, C., Gabriel, S., Ginja, C., Pires, A.E., Tereso, J.P., Eds.; Oxbow Books: Oxford, UK, 2021; pp. 13–27. [Google Scholar]
- Vigne, J.D.; Méniel, P.; Lambot, B.; Bonani, G. Un assemblage d’ossements de micromammifères dans une cache de petit mustélidé de l’âge du fer (Acy-Romance, Ardennes, France): Micromammifères, oiseaux et problèmes taphonomiques. Artefacts 1994, 9, 207–215. [Google Scholar]
- Williams, J.P. Small Mammal Deposits in Archaeology: A Taphonomic Investigation of Tyto alba (Barn Owl) Nesting and Roosting Sites. Ph.D. Thesis, University of Sheffield, Sheffield, UK, 2001. [Google Scholar]
- Denys, C. Taphonomy and experimentation. Archaeometry 2002, 44, 469–484. [Google Scholar] [CrossRef]
- Denys, C. Des référentiels en taphonomie des petits vertébrés: Bilan et perspectives. In Taphonomie des Petits Vertébrés: Référentiels et Transferts aux Fossiles; Laroulandie, V., Mallye, J.-B., Denys, C., Eds.; BAR Publishing: Oxford, UK, 2011; Volume 2269, pp. 20–21. [Google Scholar]
- Royer, A.; Montuire, S.; Gilg, O.; Laroulandie, V. A taphonomic investigation of small vertebrate accumulations produced by the snowy owl (Bubo scandiacus) and its implications for fossil studies. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2019, 514, 189–205. [Google Scholar] [CrossRef]
- Andrews, P.; Nesbit Evans, E.M. Small mammal bone accumulations produced by mammalian carnivores. Paleobiology 1983, 9, 289–307. [Google Scholar] [CrossRef]
- Sanchís, A.S. Los restos de Oryctolagus cuniculus en las tafocenosis de Bubo bubo y Vulpes vulpes y su aplicación a la caracterización del registro faunístico arqueológico. Saguntum 2000, 32, 31–49. [Google Scholar]
- Cochard, D. Etude taphonomique des léporidés d’une tanière de renard actuelle: Apport d’un référentiel à la reconnaissance des accumulations anthropiques. Rev. Paléobiol. 2004, 23, 659–673. [Google Scholar]
- Sanchís, A.S.; Pascual, J.B. Análisis de las acumulaciones óseas de una guarida de pequeños mamíferos carnívoros (Sitjar Baix, Onda, Castellón): Implicaciones arqueológicas. Archaeofauna 2011, 20, 47–71. [Google Scholar]
- Lloveras, L.; Moreno-García, M.; Nadal, J. Feeding the foxes: An experimental study to assess their taphonomic signature on leporid remains. Int. J. Osteoarchaeol. 2012, 22, 577–590. [Google Scholar] [CrossRef]
- Cohen, B.F.; Kibii, J.M. Taphonomy of a novel small carnivore: Experimental analysis of honey badger (Mellivora capensis) modifications on leporid prey. Archaeol. Anthropol. Sci. 2019, 11, 2309–2327. [Google Scholar] [CrossRef]
- Mallye, J.-B.; Cochard, D.; Laroulandie, V. Accumulations osseuses en périphérie de terriers de petits carnivores: Les stigmates de prédation et de fréquentation. Ann. Paléontol. 2008, 94, 187–208. [Google Scholar] [CrossRef]
- Campmas, E.; Stoetzel, E.; Denys, C. African carnivores as taphonomic agents: Contribution of modern coprogenic sample analysis to their identification. Int. J. Osteoarchaeol. 2018, 28, 237–263. [Google Scholar] [CrossRef]
- Rufà, A.; Laroulandie, V. Prey size as a critical factor for bird bone taphonomy in Eagle Owl (Bubo bubo) pellets. Sci. Rep. 2019, 9, 19200. [Google Scholar] [CrossRef]
- Matthews, T. Taphonomic characteristics of micromammals predated by small mammalian carnivores in South Africa: Application to fossil accumulations. J. Taphon. 2006, 4, 143–161. [Google Scholar]
- Montalvo, C.I.; Pessino, M.E.M.; Bagatto, F.C. Taphonomy of the bones of rodents consumed by Andean hog-nosed skunks (Conepatus chinga, Carnivora, Mephitidae) in central Argentina. J. Archaeol. Sci. 2008, 35, 1481–1488. [Google Scholar] [CrossRef]
- Montalvo, C.I.; Bisceglia, S.; Kin, M.S.; Sosa, R.A. Taphonomic analysis of rodent bone accumulations produced by Geoffroy’s cat (Leopardus geoffroyi, Carnivora, Felidae) in Central Argentina. J. Archaeol. Sci. 2012, 39, 1933–1941. [Google Scholar] [CrossRef]
- Montalvo, C.I.; Vezzosi, R.I.; Kin, M.S. Taphonomic analysis of rodent bones from Lontra longicaudis (Mustelidae, Carnivora) scats in fluvial environments. Mastozool. Neotrop. 2015, 22, 319–333. [Google Scholar]
- López, J.M.; Rosi, M.I.; Tabeni, S.; Bender, B.; Chiavazza, H. Taphonomic analysis of small mammal bone remains preyed upon by wildcats (Carnivora: Felidae) from the central Monte Desert (Mendoza, Argentina). Boreas 2017, 46, 282–293. [Google Scholar] [CrossRef]
- Scheifler, N.A.; Álvarez, M.C.; Rafuse, D.J.; Kaufmann, C.A.; Massigoge, A.; González, M.E.; Gutiérrez, M.A. Taphonomic signature of Geoffroy’s cat (Leopardus geoffroyi) on small sized preys: A comparative study of ingested and non-ingested leporid bones. J. Archaeol. Sci. Rep. 2020, 31, 102340. [Google Scholar] [CrossRef]
- Marin-Monfort, M.D.; García-Morato, S.; Olucha, R.; Yravedra, J.; Piñeiro, A.; Barja, I.; Andrews, P.; Fernández-Jalvo, Y. Wildcat scats: Taphonomy of the predator and its micromamal prey. Quat. Sci. Rev. 2019, 225, 106024. [Google Scholar] [CrossRef]
- Denys, C.; Stoetzel, E.; Andrews, P.; Bailon, S.; Rihane, A.; Huchet, J.B.; Fernández-Jalvo, Y.; Laroulandie, V. Taphonomy of Small Predators multi-taxa accumulations: Palaeoecological implications. Hist. Biol. 2018, 30, 868–881. [Google Scholar] [CrossRef]
- Lefranc, P.; Griselin, S.; Schneider, N.; Arbogast, R.-M.; Chenal, F.; Gebhardt, A.; Jodry, F.; Mallye, J.-B.; Moine, O.; Royer, A.; et al. Ittenheim Bas-Rhin. A355-Contournement Ouest de Strasbourg—Tronçon 3—Site 3.4. In der Flies et Musau; La Formation d’un Versant Puis son Occupation en Rive Droite de la Vallée du Musaubach, une Histoire Dynamique du Paléolithique, au Néolithique et Jusqu’à L’époque Moderne; Rapport de Fouille INR AP; SRA Grand Est: Vandœuvre-lès-Nancy, France, 2021. [Google Scholar]
- Mallye, J.-B. Les Restes de Blaireau en Contexte Archéologique: Taphonomie, Archéozoologie et Eléments de Discussion des Séquences Préhistoriques. Ph.D. Thesis, The University of Bordeaux 1, Bordeaux, France, 2007. [Google Scholar]
- Pelletier, M.; Royer, A.; Holliday, T.W.; Discamps, E.; Madelaine, S.; Maureille, B. Rabbits in the grave! Consequences of bioturbation on the Neandertal “burial” at Regourdou (Montignac-sur-Vézère, Dordogne). J. Hum. Evol. 2017, 110, 1–17. [Google Scholar] [CrossRef]
- Biadi, F.; Le Gall, A. Le Lapin de Garenne. Vie Gestion et Chasse d’un Gibier Authentique; Office National de la Chasse Hatier: Paris, France, 1993. [Google Scholar]
- Mulder, J.L.; Wallage-Drees, J.M. Red fox predation on young rabbits in breeding burrows. Neth. J. Zool. 1979, 29, 144–149. [Google Scholar] [CrossRef]
- Harris, S. Age determination in the Red fox (Vulpes vulpes)—An evaluation of technique efficiency as applied to a sample of suburban foxes. J. Zool. 1978, 184, 91–117. [Google Scholar] [CrossRef]
- Pelletier, M.; Desclaux, E.; Mallye, J.B.; Crégut-Bonnoure, E. Identifying the accidental-natural mortality of leporids in the archaeological record: Insights from a taphonomical analysis of a pitfall without evidence of human presence. J. Quat. Sci. 2020, 35, 677–694. [Google Scholar] [CrossRef] [Green Version]
- Dodson, P.; Wexlar, D. Taphonomic investigations of owl pellets. Paleobiology 1979, 5, 275–284. [Google Scholar] [CrossRef]
- Royer, A.; Lécuyer, C.; Montuire, S.; Escarguel, G.; Fourel, F.; Mann, A.; Maureille, B. Late Pleistocene (MIS 3–4) climate inferred from micromammal communities and δ18O of rodents from Les Pradelles, France. Quat. Res. 2013, 80, 113–124. [Google Scholar] [CrossRef]
- Fernández-Jalvo, Y.; Andrews, P.; Denys, C.; Sesé, C.; Stoetzel, E.; Marin-Monfort, D.; Pesquero, D. Taphonomy for taxonomists: Implications of predation in small mammal studies. Quat. Sci. Rev. 2016, 139, 138–157. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. v3.6.3.; R Core Team: Vienna, Austria, 2020. [Google Scholar]
- Josse, J.; Husson, F. missMDA: A package for handling missing values in multivariate data analysis. J. Stat. Softw. 2016, 70, 1–31. [Google Scholar] [CrossRef]
- Husson, F.; Josse, J.; Le, S.; Mazet, J.; Husson, M.F. Package ‘FactoMineR’. R Package 2016, 96, 698. [Google Scholar]
- Oksanen, J.; Blanchet, F.; Friendly, M.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; et al. Vegan: Community Ecology Package, R package, v. 2.4-6. 2016.
- Martinez Arbizu, P. pairwiseAdonis: Pairwise multilevel comparison using adonis. R package version 0.0.1. 2017. [Google Scholar]
- Donard, E. Recherche sur le Léporinés Quaternaires (Pléistocène Moyen et Supérieur, Holocène; Université de Bordeaux: Bordeaux, France, 1982; Volume 1, 161p. [Google Scholar]
- Koby, F.-E. Contribution au diagnostic ostéologique différentiel de Lepus timidus Linné et L. europaeus Pallas. Verh. Der Nat. Ges. 1959, 70, 19–44. [Google Scholar]
- Koby, F.-E. Contribution à la connaissance des lièvres fossiles, principalement de ceux de la dernière glaciation. Verh. Der Nat. Ges. 1960, 71, 149–173. [Google Scholar]
- Morel, P.; Müller, W. Hauterive-Champréveyres: Un Campement Magdalénien au Bord du lac de Neuchâtel. Étude Archéozoologique; Archéologie Neuchâteloise: Neuchâtel, France, 1997; 149p. [Google Scholar]
- Müller, W. Le Site Magdalénien de Monruz, 3 Acquisition, Traitement et Consommation des Ressources Animales; Archéologie Neuchâteloise: Neuchâtel, France, 2013. [Google Scholar]
- Flux, J.E.C. Life history of the Mountain hare (Lepus timidis scoticus) in north-east Scotland. J. Zool. 1970, 161, 75–123. [Google Scholar] [CrossRef]
- Matthews, T. South African micromammals and predators: Some comparative results. Archaeometry 2002, 44, 363–370. [Google Scholar] [CrossRef]
- Lebreton, L.; Bailon, S.; Guillaud, E.; Testu, A.; Perrenoud, C. Multi-taxa referential of a modern Eurasian Eagle-Owl (Bubo bubo) aerie. J. Archaeol. Sci. Rep. 2020, 32, 102417. [Google Scholar] [CrossRef]
- Gómez, G.N.; Kaufmann, C.A. Taphonomic analysis of Pseudalopex griseus (Gray, 1837) scat assemblages and their archaeological implications. J. Taphon. 2007, 5, 59–70. [Google Scholar]
- Denys, C.; Kowalski, K.; Dauphin, Y. Mechanical and chemical alterations of skeletal tissues in a recent Saharian accumulation of faeces from Vulpes rueppelli (Carnivora, Mammalia). Acta Zool. Cracovensia 1992, 35, 265–283. [Google Scholar]
- Schmitt, D.N.; Juell, K.E. Toward the identification of coyote scatological faunal accumulations in archaeological contexts. J. Archaeol. Sci. 1994, 21, 249–262. [Google Scholar] [CrossRef]
- Montalvo, C.I.; Pessino, M.E.; González, V.H. Taphonomic analysis of remains of mammals eaten by pumas (Puma concolor Carnivora, Felidae) in central Argentina. J. Archaeol. Sci. 2007, 34, 2151–2160. [Google Scholar] [CrossRef]
- Coll, D.G.; Montalvo, C.I.; Fernández, F.J.; Pia, M.V.; Mondini, M. Actualistic taphonomic study of the rodents digested by the Achala culpeo fox (Lycalopex culpaeus smithersi) in the highlands of central Argentina. Boreas 2021, 50, 1146–1160. [Google Scholar] [CrossRef]
- Lloveras, L.; Moreno-García, M.; Nadal, J. Taphonomic analysis of leporid remains obtained from modern Iberian lynx (Lynx pardinus) scats. J. Archaeol. Sci. 2008, 35, 1–13. [Google Scholar] [CrossRef]
- Royer, A.; Sécher, A.; Langlais, M. A Brief Note on the Presence of the Common Hamster during the Late Glacial Period in Southwestern France. Quaternary 2018, 1, 8. [Google Scholar] [CrossRef] [Green Version]
- Arilla, M.; Rufà, A.; Rosell, J.; Blasco, R. Small carnivores’ cave-dwelling: Neo-taphonomic study of a badger (Meles meles) sett and its archaeological implications. Hist. Biol. 2020, 32, 951–965. [Google Scholar] [CrossRef]
- Rodríguez-Hidalgo, A.; Saladié, P.; Marín, J.; Canals, A. Expansion of the referential framework for the rabbit fossil accumulations generated by Iberian lynx. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2015, 418, 1–11. [Google Scholar] [CrossRef]
- Rodríguez-Hidalgo, A.; Sanz, M.; Daura, J.; Sánchez-Marco, A. Taphonomic criteria for identifying Iberian lynx dens in quaternary deposits. Sci. Rep. 2020, 10, 7225. [Google Scholar] [CrossRef] [PubMed]
- Álvarez, M.C.; Kaufmann, C.A.; Massigoge, A.; Gutiérrez, M.A.; Rafuse, D.J.; Scheifler, N.A.; González, M.E. Bone modification and destruction patterns of leporid carcasses by Geoffroy’s cat (Leopardus geoffroyi): An experimental study. Quat. Int. 2012, 278, 71–80. [Google Scholar] [CrossRef]
- Rodríguez-Hidalgo, A.; Lloveras, L.; Moreno-García, M.; Saladié, P.; Canals, A.; Nadal, J. Feeding behaviour and taphonomic characterization of non-ingested rabbit remains produced by the Iberian lynx (Lynx pardinus). J. Archaeol. Sci. 2013, 40, 3031–3045. [Google Scholar] [CrossRef]
- Lloveras, L.; Nadal, J.; Fullola, J.M. Distinguishing the taphonomic signature of wolves from humans and other predators on small prey assemblages. Sci. Rep. 2020, 10, 8030. [Google Scholar] [CrossRef]
- Lloveras, L.; Moreno-García, M.; Nadal, J. The eagle owl (Bubo bubo) as a leporid remains accumulator: Taphonomic analysis of modern rabbit remains recovered from nests of this predator. Int. J. Osteoarchaeol. 2009, 19, 573–592. [Google Scholar] [CrossRef]
- Cucchi, T.; Vigne, J.D.; Auffray, J.C. First occurrence of the house mouse (Mus musculus domesticus Schwarz & Schwarz, 1943 in the Western Mediterranean: A zooarchaeological revision of subfossil occurrences. Biol. J. Linn. Soc. 2005, 84, 429–445. [Google Scholar]
- Quéré, J.P.; Le Louarn, H. Les rongeurs de France: Faunistique et biologie. In 3e Edition Revue et Augmentée; Editions Quae: Versailles, France, 2011. [Google Scholar]
- Górecki, A. Energy flow through the common hamster population. Acta Theriol. 1977, 22, 25–66. [Google Scholar] [CrossRef] [Green Version]
- Nechay, G. Status of Hamsters Cricetus Cricetus, Cricetus Migratorius, Mesocricetus Newtoni, and Other Hamster Species in Europe Nature and Environment; Council of Europe: Strasburg, France, 2000; Volume 106. [Google Scholar]
- Delattre, P. La Belette (Mustela Nivalis, Linnaeus, 1766) et L’hermine (Mustela Erminea, Linnaeus, 1758); Société Française pour l’étude et la Protection des Mammifères: Paris, France, 1987. [Google Scholar]
- Labrid, M. Labrid, M. La Martre (Martes Martes, Linnaeus, 1758. In Encyclopédie des Carnivores de France; Société française pour l’Étude et la Protection des Mammifères: Bourges, France, 1986; pp. 1–22. [Google Scholar]
- Artois, M. Le Renard Roux: Vulpes Vulpes Linnaeus, 1758; Société Française Pour l’Étude et la Protection des Mammifères: Bourges, France, 1989; 90p. [Google Scholar]
- Meia, J.-S. Le Renard: Description, Comportement, vie Sociale, Mythologie, Observation; Delachaux et Niestlé: Paris, France, 2011; 183p. [Google Scholar]
- Lloveras, L.; Thomas, R.; Cosso, A.; Pinyol, C.; Nadal, J. When wildcats feed on rabbits: An experimental study to understand the taphonomic signature of European wildcats (Felis silvestris silvestris). Archaeol. Anthropol. Sci. 2018, 10, 449–464. [Google Scholar] [CrossRef] [Green Version]
- Mondini, M. Carnivore taphonomy in South America: A review of actualistic studies and their implications in the southern Neotropics. Hist. Biol. 2018, 30, 774–785. [Google Scholar] [CrossRef]
- Montalvo, C.I.; Fernández, F.J. Review of the actualistic taphonomy of small mammals ingested by South American predators. Its importance in the interpretation of the fossil record. Publicación Electrónica De La Asoc. Paleontológica Argent. 2019, 19, 18–46. [Google Scholar] [CrossRef] [Green Version]
- Terry, R.C. Inferring predator identity from skeletal damage of small-mammal prey remains. Evol. Ecol. Res. 2007, 9, 199–219. [Google Scholar]
- Comay, O.; Dayan, T. Taphonomic signatures of owls: New insights into micromammal assemblages. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2018, 492, 81–91. [Google Scholar] [CrossRef]
- Laroulandie, V.; Royer, A.; Mallye, J.B.; Bailon, S.; Danger, M. Les petits vertébrés de Peyrazet et les chasseurs-collecteurs du Magdalénien supérieur. In La Grotte-Abri de Peyrazet (Creysse, Lot, France) au Magdalénien; Originalité Fonctionnelle d’un Habitat des Derniers Chasseurs de Rennes du Quercy, CNRS, Supplément à Gallia Préhistoire 43; Langlais, M., Laroulandie, V., Eds.; CNRS: Paris, France, 2021; pp. 141–152. Available online: https://hal.archives-ouvertes.fr/hal-03202072 (accessed on 1 November 2021).
- Kowalski, K. Some problems of the taphonomy of small mammals. In International Symposium Evolution, Phylogeny and Biostratigraphy of Arvicolids Rohanov, May 1987; Fejfar, O., Heinrich, W.-D., Eds.; Czech Geological Survey: Prague, Czech Republic, 1990; pp. 285–296. [Google Scholar]
- Butler, D.R. Zoogeomorphology: Animals as Geomorphic Agents; Cambridge University Press: Cambridge, MA, USA, 1995. [Google Scholar]
- Denys, C. Nouveaux critères de reconnaissance des concentrations de microvertébrés d’après l’étude des pelotes de chouettes du Botswana (Afrique australe). Bulletin du Muséum national d’histoire naturelle. Sect. A Zool. Biol. Écologie Anim. 1985, 7, 879–933. [Google Scholar]
- Hoffman, R. The contribution of raptorial birds to patterning in small mammal assemblages. Paleobiology 1988, 14, 81–90. [Google Scholar] [CrossRef]
- Denys, C.; Dauphin, Y.; Rzebik-Kowalska, B.; Kowalski, K. Taphonomic study of Algerian owl pellet assemblages and differential preservation of some rodents: Palaeontological implications. Acta Zool. Crac. 1996, 39, 103–116. [Google Scholar]
- Denys, C.; Dauphin, Y.; Fernández-Jalvo, Y. Apports biostratigraphiques et paléoécologiques de l’étude taphonomique des assemblages de micromammifères. Bilan et perspectives. Geobios 1997, 30, 197–206. [Google Scholar] [CrossRef]
- Saavedra, B.; Simonetti, J.A. Small mammal taphonomy: Intraspecific bone assemblage comparison between South and North American barn owl, Tyto alba, populations. J. Archaeol. Sci. 1998, 25, 165–170. [Google Scholar] [CrossRef]
- Cochard, D. Discussion sur la variabilité intraréférentiel d’accumulations osseuses de petits prédateurs. Ann. Paléontologie 2008, 94, 89–101. [Google Scholar] [CrossRef]
- Rudzik, S.M.; Fernández, F.J.; Carrera, J.D. Taphonomic analysis of micromammal remains from striped owl (Pseudoscops clamator) pellets in northeastern Buenos Aires province, Argentina: Implications for archaeological sites formation. Int. J. Osteoarchaeol. 2015, 25, 550–563. [Google Scholar] [CrossRef]
- Raczyński, J.; Ruprecht, A.L. The effect of digestion on the osteological composition of owl pellets. Acta Ornithol. 1974, 14, 25–36. [Google Scholar]
- Andrews, P.; Fernández-Jalvo, Y. Seasonal variation in prey composition and digestion in small mammal predator assemblages. Int. J. Osteoarchaeol. 2018, 28, 318–331. [Google Scholar] [CrossRef]
- Cohen, B.F.; Kibii, J.M. Experimental study of bone modification by captive caracal (Caracal caracal); a model for fossil assemblage analysis. J. Archaeol. Sci. 2015, 62, 92–104. [Google Scholar] [CrossRef]
- Gidna, A.; Yravedra, J.; Domínguez-Rodrigo, M. A cautionary note on the use of captive carnivores to model wild predator behavior: A comparison of bone modification patterns on long bones by captive and wild lions. J. Archaeol. Sci. 2013, 40, 1903–1910. [Google Scholar] [CrossRef]
- Rafuse, D.J.; Kaufmann, C.A.; Gutiérrez, M.A.; González, M.E.; Scheifler, N.A.; Álvarez, M.C.; Massigoge, A. Taphonomy of modern communal burrow systems of the Plains vizcacha (Lagostomus maximus, Chinchillidae) in the Pampas region of Argentina: Implications for the fossil record. Hist. Biol. 2019, 31, 517–534. [Google Scholar] [CrossRef]
- Scheifler, N.A.; Massigoge, A.; Kaufmann, C.A.; Rafuse, D.J.; González, M.E.; Gutiérrez, M.A. Modern Bone Distribution in the Pampas of Argentina: Taphonomic Implications for the Regional Archaeological Record. In Actualistic Taphonomy in South America. Topics in Geobiology; Martínez, S., Rojas, A., Cabrera, F., Eds.; Springer: Cham, Switzerland, 2020; Volume 48, pp. 193–220. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | NISP | MNE | MNI |
|---|---|---|---|
| Lepus timidus | 108 | 68 | 3 |
| Vulpes vulpes | 53 | 45 | 2 |
| Lagomorpha/Vulpes | 10 | - | - |
| Cricetus cricetus | 2358 | 2279 | 64 |
| Apodemus cf. sylvaticus | 4 | 4 | 2 |
| Apodemus/mus | 2 | 2 | 1 |
| Muridae | 25 | 25 | 1 |
| Arvicola cf. amphibius | 8 | 8 | 1 |
| Microtus agrestis | 4 | 4 | 3 |
| Microtus arvalis/agrestis | 5 | 5 | 3 |
| Arvicolidae | 87 | 85 | - |
| Talpa europaea | 7 | 7 | 1 |
| Suinae | 1 | 1 | 1 |
| Bird | 2 | 2 | 1 |
| Indéterminés | 12 | - | |
| Total | 2686 | 2535 | 83 |
| Leporid (MNI = 3) | Fox (MNI = 2) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Anatomy | NISP | MNE | NISP CO | % CB | % RA | NISP Digested | NISP Bones with Tooth Marks | NISP | MNE | NISP CO | % CB | % RA | NISP Digested | NISP Bones with Tooth Marks |
| Mandible | - | - | - | - | - | - | - | 3 | 3 | 2 | 66.7 | 75.0 | 0 | 1 |
| Maxillary | 2 | 2 | 0 | 0.0 | 33.3 | - | 0 | 0 | 0 | - | - | 0.0 | - | - |
| Cranial | 5 | - | 0 | 0.0 | - | - | 0 | 7 | 1 | 1 | 14.3 | - | 0 | 0 |
| Incisors | 1 | 1 | 1 | 100.0 | 5.6 | - | 0 | 2 | 2 | 1 | 50.0 | 8.3 | 0 | 0 |
| Molars | 9 | 8 | 4 | 44.4 | 12.1 | 1 | 0 | 1 | 1 | 0 | 0.0 | 5.0 | 0 | 0 |
| Other teeth | - | - | - | - | - | - | - | 2 | 2 | 2 | 100.0 | 50.0 | 0 | 0 |
| Scapula | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Humerus | 1 | 1 | 0 | 0.0 | 16.7 | - | 0 | 1 | 1 | 1 | 100.0 | 25.0 | 0 | 0 |
| Radius | - | - | - | - | - | - | 0 | - | - | - | - | - | - | - |
| Ulna | 1 | 1 | 0 | 0.0 | 16.7 | - | 1 | 3 | 2 | 2 | 66.7 | 50.0 | 0 | 0 |
| Femur | 8 | 3 | 2 | 25.0 | 50.0 | - | 4 | 1 | 1 | 0 | 0.0 | 25.0 | 0 | 0 |
| Tibia | 9 | 3 | 5 | 55.6 | 50.0 | - | 3 | - | - | - | - | - | - | - |
| Pelvis | 1 | 1 | 0 | 0.0 | 16.7 | - | 1 | - | - | - | - | - | - | - |
| Calcaneum | 5 | 5 | 5 | 100.0 | 83.3 | - | 0 | - | - | - | - | - | - | - |
| Talus | 3 | 3 | 3 | 100.0 | 50.0 | - | 0 | 1 | 1 | 1 | 100.0 | 25.0 | - | 0 |
| Carpals/tarsals | 7 | 7 | 7 | 100.0 | 9.7 | - | 0 | 1 | 1 | 1 | 100.0 | 25.0 | 0 | 0 |
| Patella | 1 | 1 | 1 | 100.0 | 16.7 | - | - | - | - | - | - | - | - | - |
| Sesamoïd | - | - | - | - | - | - | 0 | 1 | 1 | 1 | 100.0 | - | - | 0 |
| Ribs | 2 | 2 | 2 | 100.0 | 2.6 | - | 0 | 3 | 3 | 3 | 100.0 | 5.8 | 0 | 0 |
| Vertebra | 1 | 1 | 1 | 100.0 | 0.9 | - | 0 | 16 | 8 | 0 | 0.0 | 9.8 | 0 | 0 |
| Sternum | - | - | - | - | - | - | 0 | 1 | 1 | 1 | 100.0 | 50.0 | - | 0 |
| Metapodial | 34 | 22 | 23 | 67.6 | 40.7 | - | 0 | 7 | 7 | 3 | 42.9 | 19.4 | 0 | 0 |
| Phalanx | 21 | 20 | 20 | 95.2 | 19.6 | - | 0 | 10 | 10 | 6 | 60.0 | 9.6 | 0 | 0 |
| Baculum | - | - | - | - | - | - | 0 | 1 | 1 | 1 | 100.0 | 50.0 | 0 | 0 |
| Total | 111 | 81 | 1 | 9 | 61 | 46 | 0 | 1 | ||||||
| Hamster (MNI = 64) | Small Voles and Mice (MNI = 7) | Talpa europaea (NMI = 1) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Anatomy | MNE | % CB | % RA | % Digestion | % Bones with Tooth Marks | MNE | % CB | % RA | % Digestion | % Bones with Tooth Marks | MNE | % CB | % RA | NISP Digested | NISP Bones with Tooth Marks |
| Mandible | 119 | 47 | 93.0 | - | 13.1 | 5 | 20 | 35.7 | - | 0 | 0 | - | - | - | - |
| Maxillary | 125 | - | 97.7 | - | - | 2 | 0 | 14.3 | - | - | 0 | - | - | - | - |
| Incisors | 219 | - | 85.5 | 0 | - | 26 | - | 92.9 | 72.7 | - | - | - | - | - | - |
| Molars | 544 | - | 70.8 | - | - | 64 | - | 76.2 | 85.7 * | - | - | - | - | - | - |
| Scapula | 28 | 0 | 21.9 | - | - | - | - | - | - | - | 1 | 100 | 50 | 0 | 0 |
| Humerus | 74 | 32 | 57.8 | 5.7 | 40.2 | 5 | 0 | 35.7 | 60 | 0 | 1 | 100 | 50 | 0 | 0 |
| Radius | 72 | 52 | 56.3 | 1.2 | 10.7 | 1 | 0 | 7.1 | 100 | 0 | - | - | - | - | - |
| Ulna | 67 | 40 | 52.3 | 0 | 17.0 | 5 | 20 | 35.7 | 60 | 0 | 1 | 100 | 50 | 0 | 0 |
| Femur | 93 | 56 | 72.7 | 2.9 | 23.1 | 7 | 43 | 50.0 | 28.6 | 14.3 | 2 | 100 | 100 | 0 | 0 |
| Tibia | 78 | 53 | 60.9 | 0 | 21.6 | 2 | 0 | 14.3 | 100 | 0 | 1 | 0 | 50 | 0 | 0 |
| Pelvis | 85 | 8 | 66.4 | 0 | 39.8 | 7 | 0 | 50.0 | 57.1 | 57.1 | 1 | 100 | 50 | 0 | 0 |
| Calcaneum | 62 | - | 48.4 | x | - | - | - | - | - | - | - | - | - | - | - |
| Talus | 56 | - | 43.8 | - | - | - | - | - | - | - | - | - | - | - | - |
| Carpals/tarsals | - | - | - | - | - | - | - | - | - | - | 0 | - | - | - | - |
| Vertebra | 397 | - | 20.7 | x | x | 22 | - | - | x | - | - | - | - | - | - |
| Metapodial | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Phalanx | - | - | - | x | - | - | - | - | - | - | - | - | - | - | - |
| Total | 2019 | 146 | 7 | ||||||||||||
| Digestion | ||||||||
|---|---|---|---|---|---|---|---|---|
| Site | Size | Element | 0 | 1 | 2 | 3 | 4 | Total |
| Bettant I | Ondatra, Rattus and Erinaceus | I inf | 4 | 1 | 0 | 0 | 0 | 5 |
| I sup | 2 | 2 | 0 | 0 | 0 | 4 | ||
| m1 inf | 3 | 1 | 1 | 0 | 0 | 5 | ||
| FEM | 4 | - | - | - | - | 4 | ||
| Arvicola sp. | I inf | 4 | 1 | 0 | 0 | 0 | 5 | |
| I sup | 2 | 0 | 0 | 0 | 0 | 2 | ||
| m1 inf | 4 | 3 | 1 | 0 | 0 | 8 | ||
| FEM | 0 | 2 | 0 | 0 | 0 | 2 | ||
| Small voles and mice | I inf | 58 | 34 | 8 | 7 | 0 | 107 | |
| I sup | 43 | 26 | 6 | 3 | 1 | 79 | ||
| m1 inf | 64 | 37 | 23 | 5 | 4 | 133 | ||
| FEM | 10 | 3 | 0 | 0 | 0 | 13 | ||
| Bettant VIII | Rattus and Erinaceus | I inf | 3 | - | - | - | - | 3 |
| I sup | 6 | - | - | - | - | 6 | ||
| m1 inf | 1 | - | - | - | - | 1 | ||
| FEM | 1 | - | - | - | - | 1 | ||
| Arvicola sp. | I inf | 2 | 0 | 0 | 1 | 0 | 3 | |
| I sup | 3 | 1 | 0 | 0 | 0 | 4 | ||
| m1 inf | 2 | 2 | 4 | 3 | 1 | 12 | ||
| FEM | 1 | - | - | - | - | 1 | ||
| Small voles and mice | I inf | 29 | 16 | 6 | 3 | 3 | 57 | |
| I sup | 20 | 17 | 2 | 3 | 0 | 42 | ||
| m1 inf | 36 | 20 | 18 | 12 | 3 | 89 | ||
| FEM | 3 | 2 | 2 | 1 | 0 | 8 | ||
| Les Six chemins | Rattus rattus | I inf | 1 | - | - | - | - | 1 |
| I sup | 0 | - | - | - | - | 0 | ||
| m1 inf | 2 | - | - | - | - | 2 | ||
| FEM | 2 | - | - | - | - | 2 | ||
| Small voles and mice | I inf | 32 | 3 | 1 | 2 | 1 | 39 | |
| I sup | 28 | 0 | 0 | 1 | 0 | 29 | ||
| m1 inf | 18 | 6 | 4 | 0 | 0 | 28 | ||
| FEM | 6 | 1 | 0 | 0 | 0 | 7 | ||
| Ondatra, Rattus, and Erinaceus (MNI = 5) | Arvicola sp. (MNI = 5) | Small voles and Mice (MNI = 73) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Anatomy | MNE | %CB | %RA | %Digestion | %Bones with Tooth Marks | MNE | %CB | %RA | %Digestion | %Bones with Tooth Marks | MNE | %CB | %RA | %Digestion | %Bones with Tooth Marks |
| Mandible | 5 | 50.0 | 50.0 | - | 16.7 | 0 | - | 0.0 | - | - | 50 | 21.2 | 54.3 | - | 8.0 |
| Maxillary | 6 | - | 60.0 | - | - | 1 | - | 6.3 | - | - | 18 | - | 19.6 | - | - |
| Incisors | 7 | - | 35.0 | 33.3 | - | 7 | - | 21.9 | 11.1 | - | 186 | - | 101.1 | 45.7 | - |
| Molars | 16 | - | 26.7 | 33.3 ** | - | 29 | - | 30.2 | 50.0 * | - | 225 | - | 40.8 | 52.2* | - |
| Scapula | 3 | 0.0 | 30.0 | - | x | 1 | - | 6.3 | - | - | 22 | - | 23.9 | - | - |
| Humerus | 5 | 100.0 | 50.0 | 0.0 | 40.0 | 2 | 0.0 | 12.5 | 0.0 | 50 | 49 | 24.3 | 53.3 | 65.3 | 1.4 |
| Radius | 2 | 50.0 | 20.0 | 0.0 | 100.0 | 2 | 0.0 | 12.5 | 100.0 | 0.0 | 14 | 64.3 | 15.2 | 7.1 | 14.3 |
| Ulna | 3 | 66.7 | 30.0 | - | 33.3 | 0 | - | 0.0 | - | - | 42 | 44.2 | 45.7 | 42.9 | 2.4 |
| Femur | 4 | 100.0 | 40.0 | 0.0 | 25.0 | 6 | 25.0 | 37.5 | 100.0 | 12.5 | 46 | 23.2 | 50.0 | 51.1 | 8.9 |
| Tibia | 5 | 80.0 | 50.0 | 0.0 | 25.0 | 6 | 28.6 | 37.5 | 50.0 | 25.6 | 37 | 30.4 | 40.2 | 52.9 | 14.8 |
| Pelvis | 5 | 16.7 | 50.0 | 0.0 | 50.0 | 3 | 0.0 | 18.8 | 50.0 | 0.0 | 35 | 2.6 | 38.0 | 53.8 | 10.3 |
| Calcaneum | 1 | - | 10.0 | - | - | - | - | - | - | - | - | - | - | - | - |
| Talus | 0 | - | 0.0 | - | - | - | - | - | - | - | - | - | - | - | - |
| Carpals/tarsals | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Vertebra | 21 | - | 14.0 | - | - | 56 | - | 37.3 | - | - | 171 | - | 12.4 | - | - |
| Metapodial | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Phalanx | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Total | 83 | 113 | 895 | ||||||||||||
| Rattus and Erinaceus (MNI = 5) | Arvicola sp. (MNI = 8) | Small Voles and Mice (MNI = 46) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Anatomy | MNE | %CB | %RA | %Digestion | %Bones with Tooth Marks | MNE | %CB | %RA | %Digestion | %Bones with Tooth Marks | MNE | %CB | %RA | %Digestion | %Bones with Tooth Marks |
| Mandible | 2 | 50.0 | 20.0 | - | 0.0 | 2 | 50 | 12.5 | - | 0.0 | 50 | 14.8 | 54.3 | - | 13.0 |
| Maxillary | 1 | - | 10.0 | - | - | 0 | - | 0.0 | - | - | 13 | - | 14.1 | - | - |
| Incisors | 9 | - | 45.0 | 0.0 | - | 7 | - | 21.9 | 33.3 | - | 100 | - | 54.3 | 50.5 | - |
| Molars | 5 | - | 8.3 | - | - | 45 | - | 46.9 | 83.3 * | - | 170 | - | 30.8 | 59.6 * | - |
| Scapula | 3 | 0.0 | 30.0 | - | - | 3 | - | 18.8 | - | - | 14 | - | 15.2 | - | - |
| Humerus | 2 | 0.0 | 20.0 | 50.0 | 0.0 | 7 | 9.1 | 43.8 | 57.1 | 45.5 | 38 | 19.2 | 41.3 | 60.5 | 5.7 |
| Radius | 2 | 100.0 | 20.0 | 0.0 | 50.0 | 2 | 100.0 | 12.5 | 0.0 | 0.0 | 7 | 57.1 | 7.6 | 28.6 | 0.0 |
| Ulna | 0 | - | 0.0 | - | - | 3 | 33.3 | 18.8 | 33.3 | 0.0 | 19 | 21.1 | 20.7 | 50.0 | 10.5 |
| Femur | 3 | 20.0 | 30.0 | 33.3 | 40.0 | 7 | 8.3 | 43.8 | 42.9 | 75.0 | 38 | 18.2 | 41.3 | 59.5 | 31.8 |
| Tibia | 1 | 0.0 | 10.0 | 0.0 | 100.0 | 7 | 8.3 | 43.8 | 100.0 | 41.7 | 30 | 13.9 | 32.6 | 70.0 | 22.2 |
| Pelvis | 8 | 12.5 | 80.0 | 0.0 | 87.5 | 13 | 0.0 | 81.3 | 28.6 | 26.7 | 19 | 0.0 | 20.7 | 43.5 | 30.4 |
| Calcaneum | 0 | - | 0.0 | - | - | - | - | - | - | - | - | - | - | - | - |
| Talus | 0 | - | 0.0 | - | - | - | - | - | - | - | - | - | - | - | - |
| Carpals/tarsals | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Vertebra | 7 | - | 4.7 | - | x | 33 | - | 13.8 | - | - | 236 | - | 17.1 | - | - |
| Metapodial | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Phalanx | - | - | - | - | x | - | - | - | - | - | - | - | - | - | - |
| Total | 43 | 129 | 734 | ||||||||||||
| Rattus rattus (MNI = 3) | Small Voles and Mice (MNI = 21) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Anatomy | MNE | %CB | %RA | NISP Digested | NISP Bones with Tooth Marks | MNE | %CB | %RA | %Digestion | %Bones with Tooth Marks |
| Mandible | 3 | 33.3 | 50.0 | - | 0 | 29 | 60.0 | 69.0 | - | 0.0 |
| Maxillary | 1 | - | 16.7 | - | - | 13 | - | 31.0 | - | - |
| Incisors | 1 | - | 8.3 | 0 | - | 69 | - | 82.1 | 12.5 | - |
| Molars | 7 | - | 19.4 | - | - | 65 | - | 25.8 | 35.7 * | - |
| Scapula | 0 | - | 0.0 | - | - | 2 | - | - | - | - |
| Humerus | 1 | 0.0 | 16.7 | - | 0 | 10 | 28.6 | 23.8 | 16.7 | 0.0 |
| Radius | 2 | 50.0 | 33.3 | 0 | 0 | 11 | 72.7 | 26.2 | 0.0 | 0.0 |
| Ulna | 1 | 100.0 | 16.7 | 0 | 0 | 3 | 100.0 | 7.1 | 0.0 | 0.0 |
| Femur | 4 | 50.0 | 66.7 | 0 | 0 | 14 | 36.8 | 33.3 | 20.0 | 0.0 |
| Tibia | 0 | - | 0.0 | - | - | 19 | 54.2 | 45.2 | 0.0 | 0.0 |
| Pelvis | 1 | 0.0 | 16.7 | 0 | 0 | 11 | 9.1 | 26.2 | 44.4 | 18.2 |
| Calcaneum | 0 | - | 0.0 | - | - | - | - | - | - | - |
| Talus | 0 | - | 0.0 | - | - | - | - | - | - | - |
| Carpals/tarsals | - | - | - | - | - | - | - | - | - | - |
| Vertebra | 5 | - | 5.6 | - | - | 16 | - | - | - | - |
| Metapodial | - | - | - | - | - | - | - | - | - | - |
| Phalanx | - | - | - | - | - | - | - | - | - | - |
| Total | 26 | 275 | ||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Royer, A.; Mallye, J.-B.; Pelletier, M.; Griselin, S. Who Killed the Small Mammals of Ittenheim (Northeastern France)? An Integrative Approach and New Taphonomic Data for Investigating Bone Assemblages Accumulated by Small Carnivores. Quaternary 2021, 4, 41. https://doi.org/10.3390/quat4040041
Royer A, Mallye J-B, Pelletier M, Griselin S. Who Killed the Small Mammals of Ittenheim (Northeastern France)? An Integrative Approach and New Taphonomic Data for Investigating Bone Assemblages Accumulated by Small Carnivores. Quaternary. 2021; 4(4):41. https://doi.org/10.3390/quat4040041
Chicago/Turabian StyleRoyer, Aurélien, Jean-Baptiste Mallye, Maxime Pelletier, and Sylvain Griselin. 2021. "Who Killed the Small Mammals of Ittenheim (Northeastern France)? An Integrative Approach and New Taphonomic Data for Investigating Bone Assemblages Accumulated by Small Carnivores" Quaternary 4, no. 4: 41. https://doi.org/10.3390/quat4040041
APA StyleRoyer, A., Mallye, J.-B., Pelletier, M., & Griselin, S. (2021). Who Killed the Small Mammals of Ittenheim (Northeastern France)? An Integrative Approach and New Taphonomic Data for Investigating Bone Assemblages Accumulated by Small Carnivores. Quaternary, 4(4), 41. https://doi.org/10.3390/quat4040041