4. Discussion
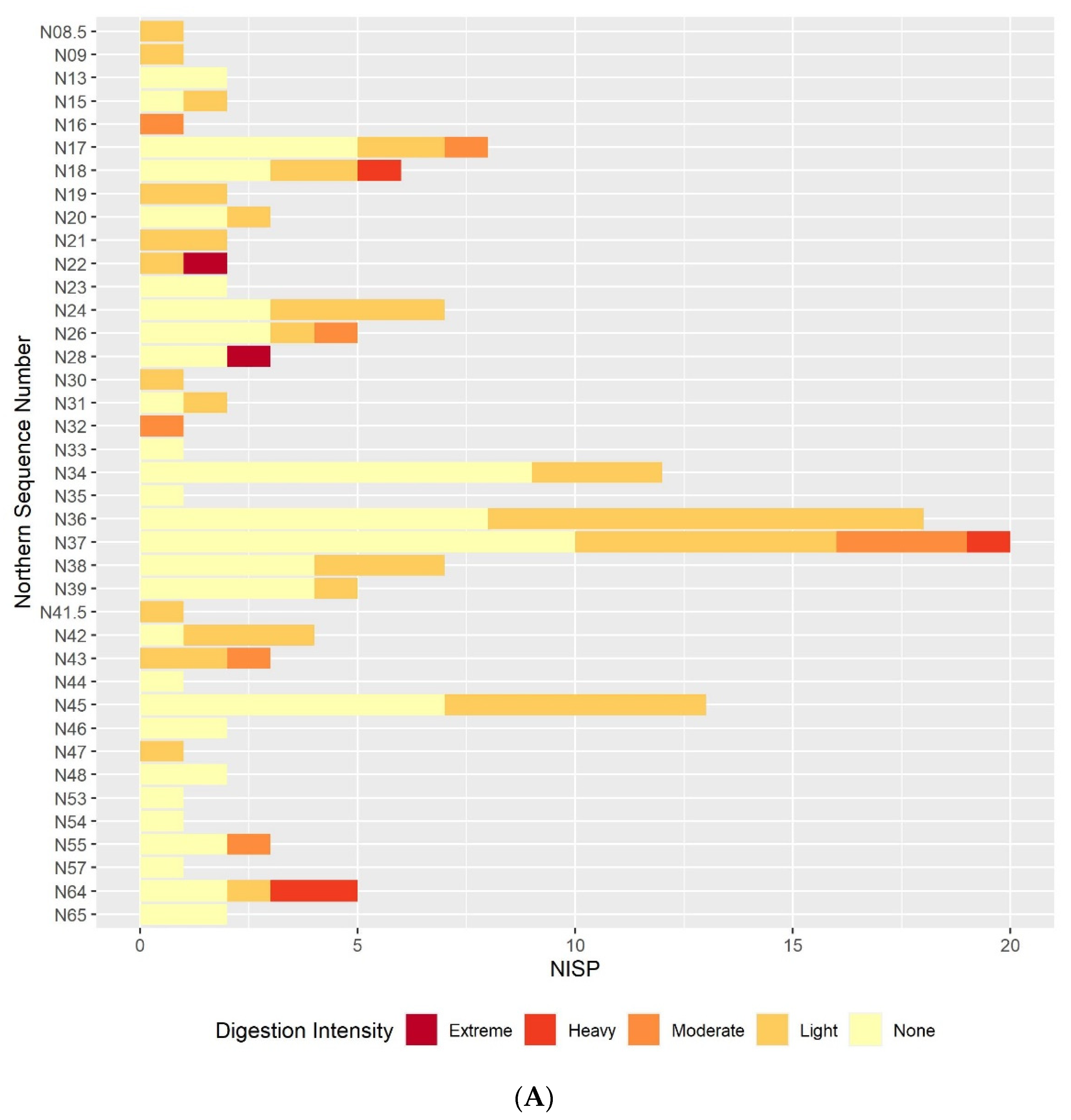
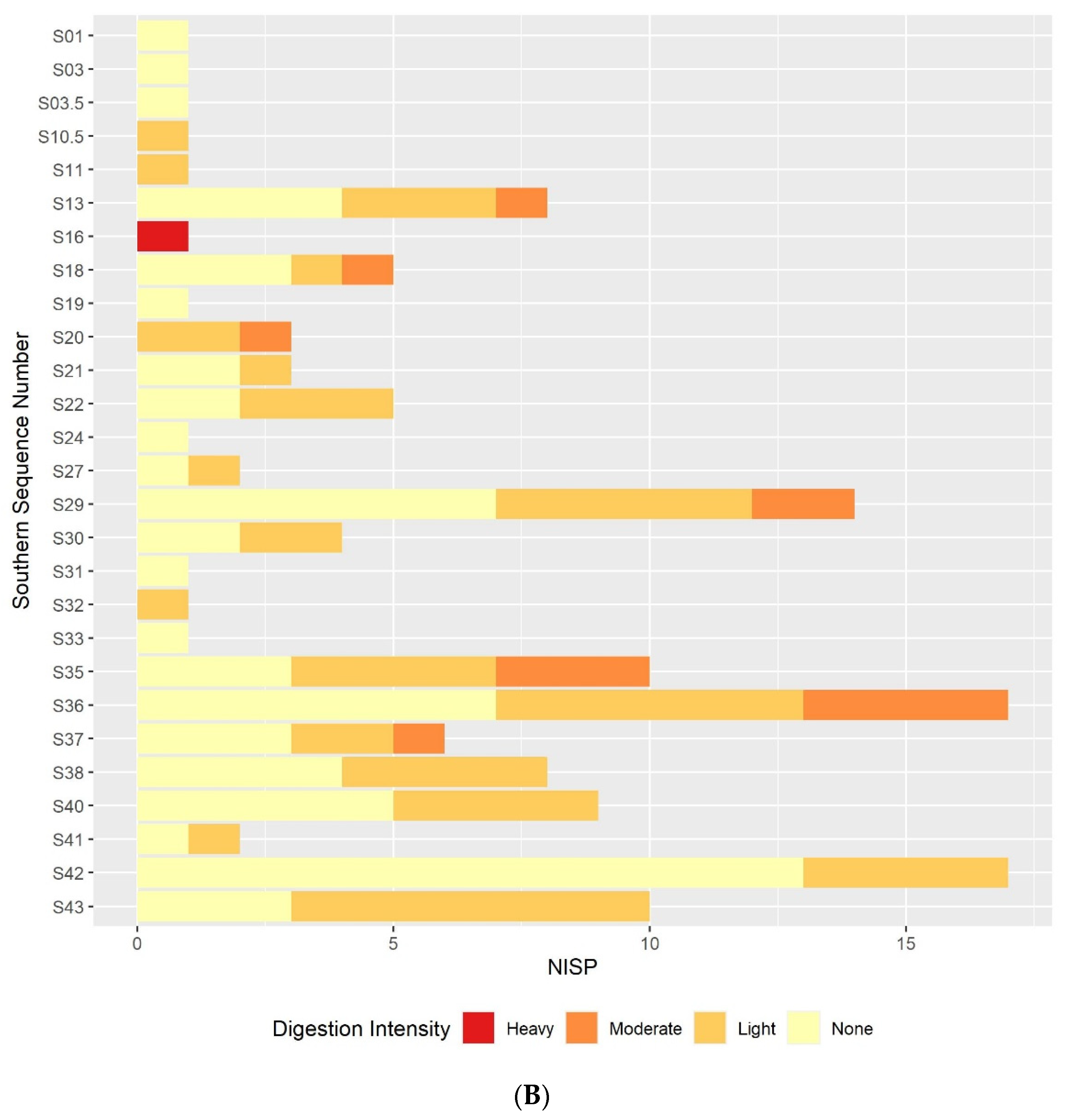
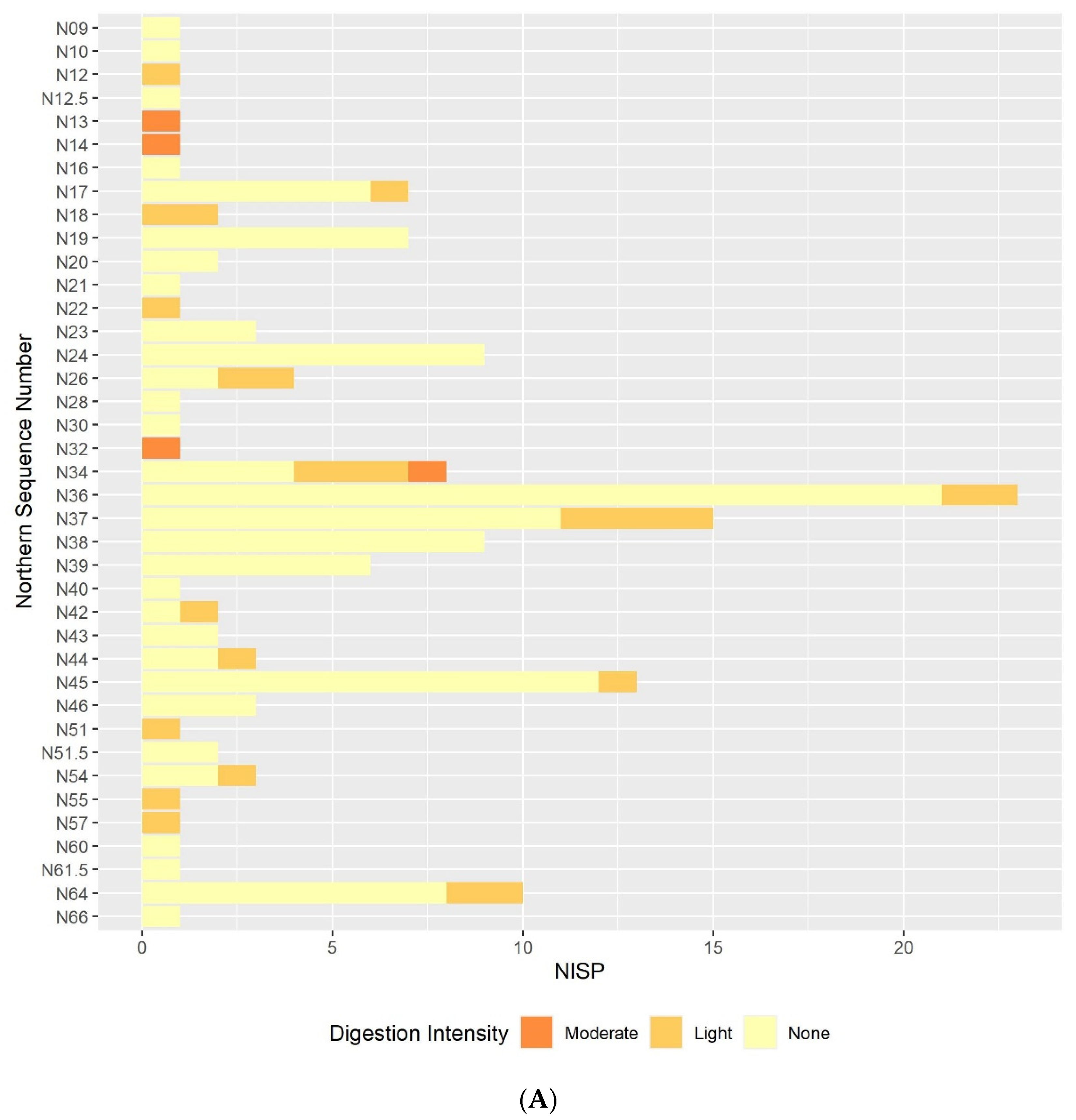
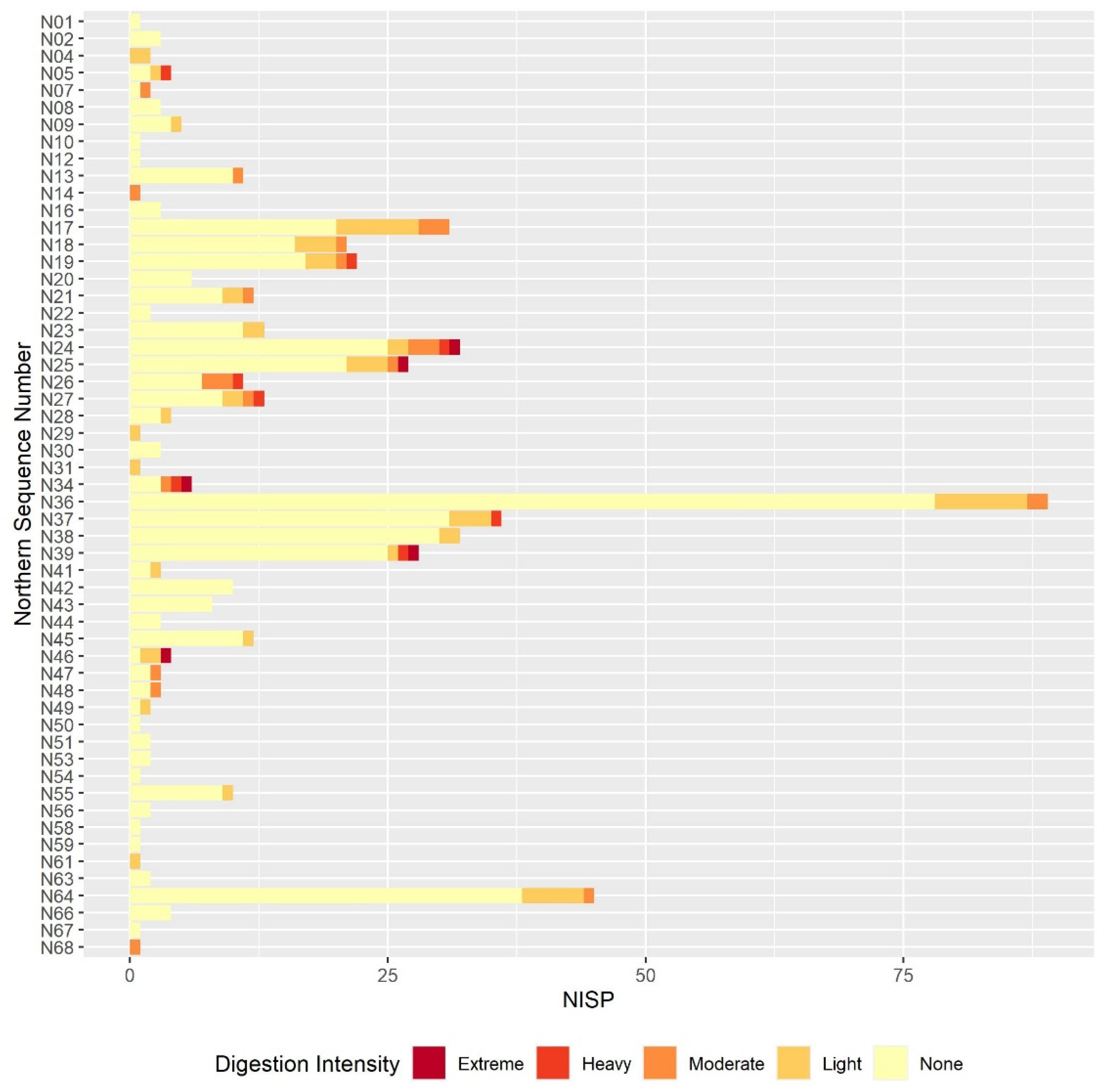
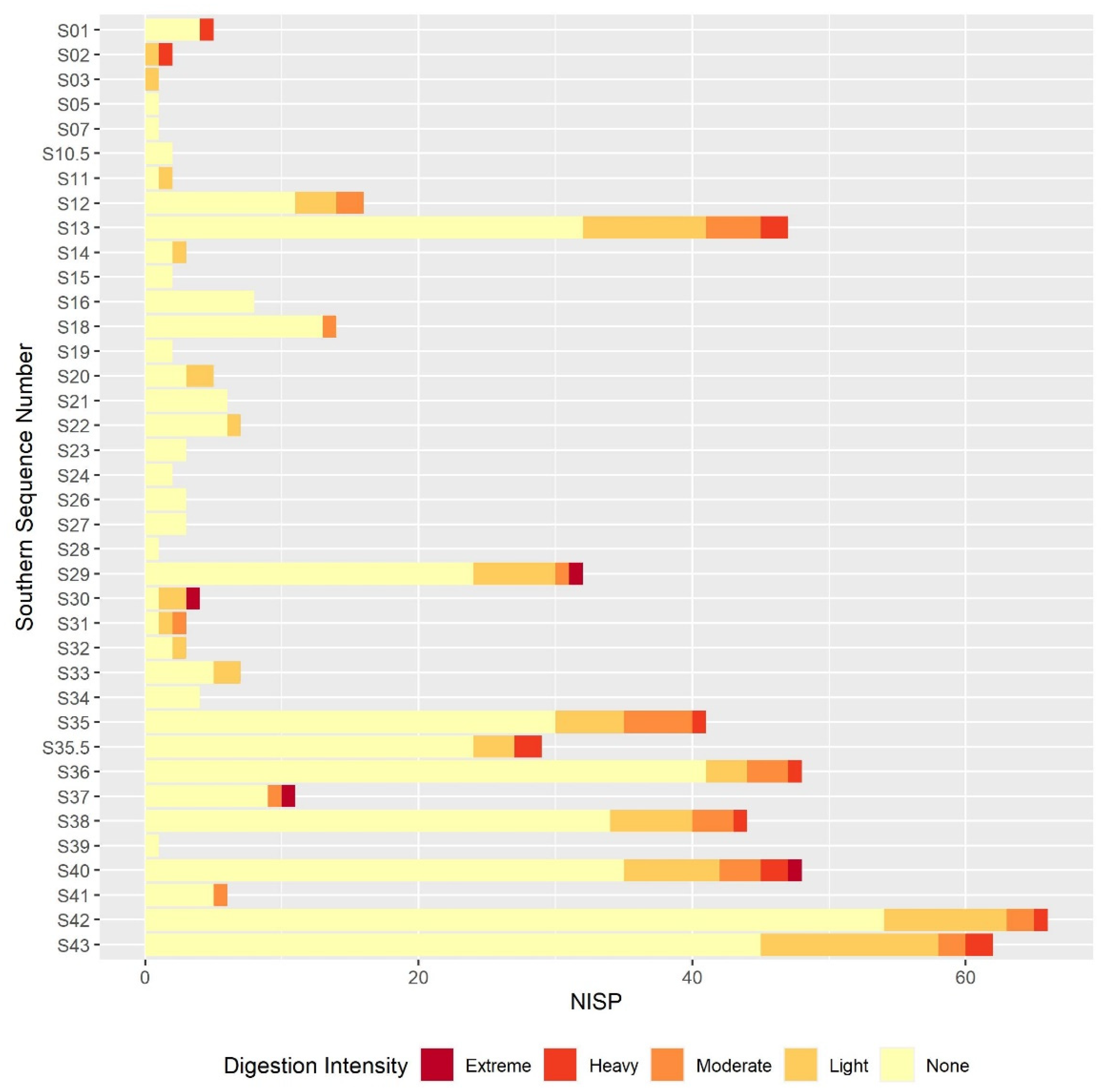
The results of this preliminary analysis suggest that in most respects the taphonomic processes affecting the microvertebrate assemblage remain constant throughout the sequence. Certain individual sequence units do appear to have particularly distinct digestion or breakage patterns, but these are generally extremely small units. If these extremely small sequence units are excluded, the overall picture of the assemblage is one of high fragmentation and low digestion intensity throughout the sequence. The only exception to this continuity is the slight decline in fragmentation in the lower contexts that are dated to the earlier phases of the sequence. The majority of humeri and femora are still broken, and most teeth are still loose from the jaw but there is a small reduction in the overall proportion in each case. This is best illustrated in the teeth breakage data, with the proportion of teeth broken in the lowest sample column and area half that of the proportion in the highest sample columns. There are multiple possible causes of the high fragmentation rates, including rockfall, trampling, predation, and sediment movement. Although some predators, such as diurnal raptors and mammalian carnivores, can produce very high levels of fragmentation [
45], they are unlikely to be the cause in this case. Predators that produce high levels of fragmentation generally leave more intensive digestive marks on the teeth, and this does not seem to be the case in the Shanidar Cave assemblage. Indeed, many of the incomplete specimens examined display no traces of digestion.
Extensive rockfall is visible throughout most of the assemblage, particularly in the upper layers, and these high energy rockfall events are likely to be responsible for most of the observed fragmentation. Geomorphological analyses to determine the exact nature and causes of these repeated rockfall events are ongoing. One current explanation is that they are associated with colder, arid periods, as they are generally absent from the lowest layers of the sequence [
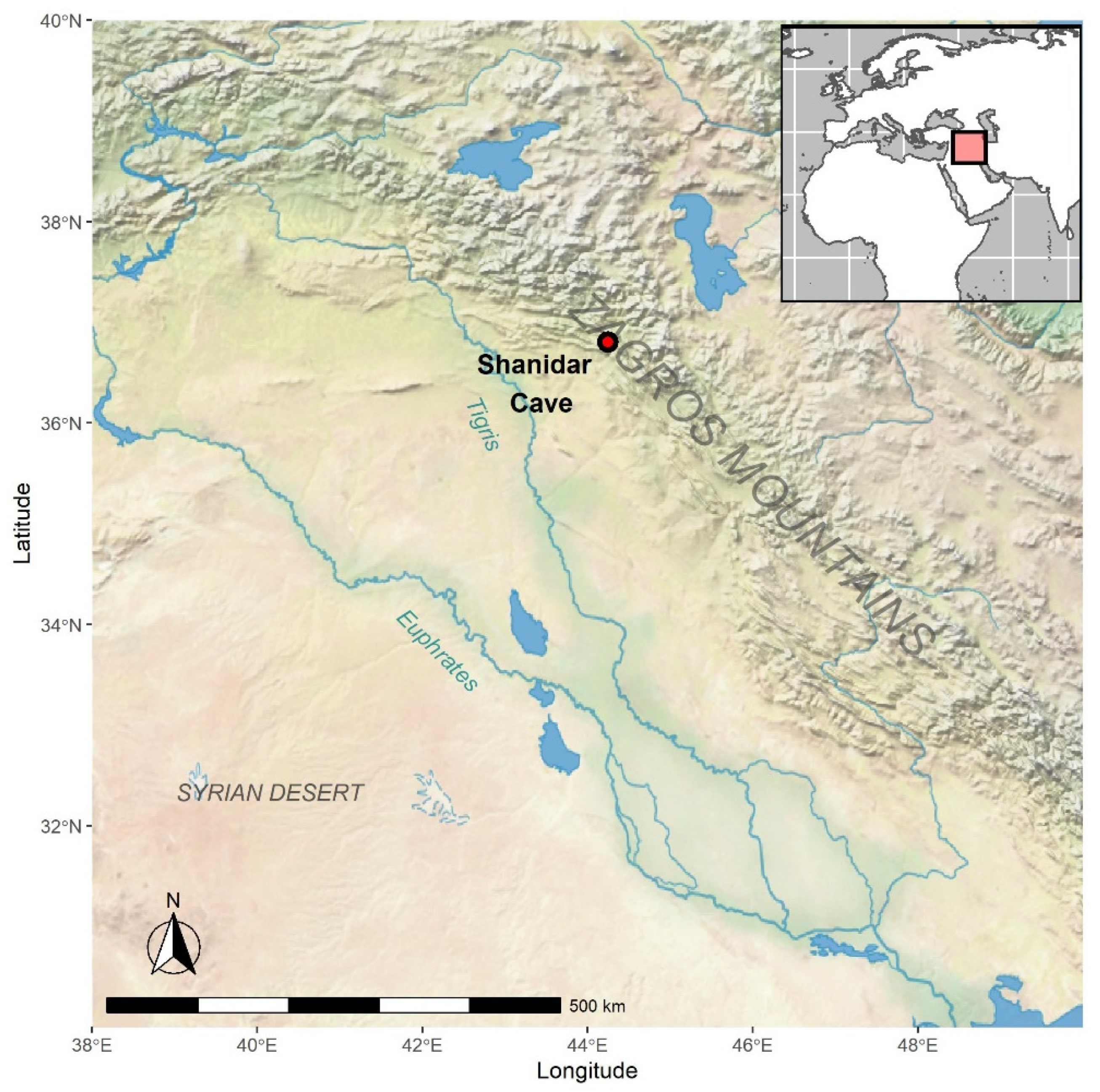
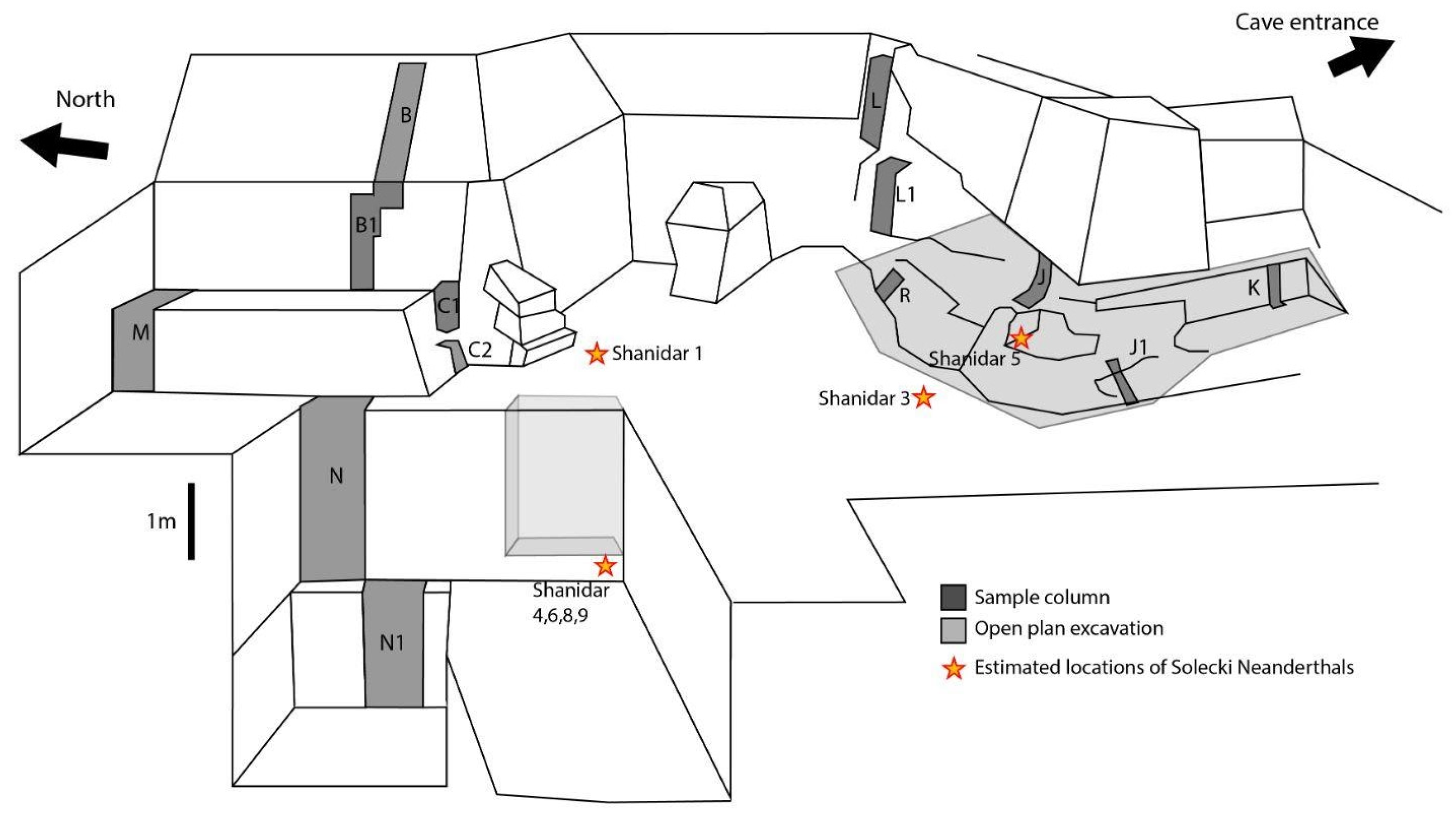
27]. Relatively warmer, humid conditions are also indicated by the presence of increased manganese dioxide staining in these layers. These lower layers are present at the bottom of the northern sequence only, as the southern sequence does not extend as deep. This is significant because the northern sequence is located towards the back of the cave, further away from the entrance than the southern sequence. The presence of manganese staining and root etching indicates that the warm and humid conditions extended quite far into the cave, enough to support some plant growth. Although dating for the lowest layers is still ongoing, preliminary dates suggest that these sediments below the lower Neanderthal cluster are older than 70,000 years BP. This would place these sediments in the range of Marine Isotope Stage 5a, a period of relatively warmer conditions. Further palaeoecological analyses of the microvertebrate assemblage and other environmental samples are needed to confirm these warmer conditions, but these results present an intriguing starting point.
Results from the digestion analyses also show no large-scale changes in digestion intensity, or at least none that can be distinguished from others due to the small sample size. In the molars and, to a slightly lesser degree, the humeri and femora, the majority of specimens show no signs of digestion, with the next most commonly occurring digestion category of light. The presence of occasional more heavily digested specimens indicates that at least one predator is responsible for at least part of the assemblage accumulation, and that this predator is likely to be a category 1 or 2 predator following Andrews’ classification of different predators’ modification of microvertebrate teeth, humeri and femora [
40], though further analyses of the incisors and other microvertebrate taxa will be needed to confirm this. The proportion of digested molars does not closely match the proportions seen in modern predators, except for the Eurasian eagle owl (
Bubo bubo), the tawny owl (
Strix aluco), and possibly the barn owl (
Tyto alba), long-eared owl (
Asio otus), and short-eared owl (
Asio flammeus) [
44]. These species can be found in present-day Iraq;
B. Bubo and
T. alba are resident year-round, whereas
S. alucois,
A. otus, and
A. flammeus are much rarer and present only in the winter [
48]. None of these species are reported in Hesse’s preliminary list of microvertebrates reported in Appendix B of Evins’ 1981 report [
22]. It should be noted that absence of physical evidence of the predator responsible is not necessarily evidence of absence, and that we would not generally expect the remains of the predator responsible to be found among the prey fossils. The only bird of prey found was a kestrel,
Falco cf.
tinnunculus [
22]. If
Falco was responsible for assemblage accumulation, a much higher proportion of digested molars would be expected (~53%), and those that were digested would be categorized as heavily digested.
S. aluco also produces teeth that would be categorized as heavily digested, and so is unlikely to be the primary contributor to the assemblage.
B. bubo produces a lower proportion of teeth displaying any digestion, though those teeth still tend to be more heavily digested than those examined in the Shanidar Cave assemblage [
44].
B. bubo nests in rocky habitats, cliffs, and small caves, and hunts over open habitats, very similar to the karstic landscape around Shanidar Cave. Attempts were made during the excavation seasons to collect owl pellets in order to understand the local avian predator community and diet. Unfortunately, no pellets have been recovered to date, though it is possible that some may be recovered in future excavation seasons. A motion-activated trail camera was placed in the cave during the 2018 excavation season. It recorded images of a larger owl species visiting the cave, but the image quality was too low to allow specific identification. Preliminary taphonomic analysis of the small mammal assemblage of Kaldar Cave (approximately 500 km northwest of Shanidar Cave) found a high proportion of digested elements in the assemblage, and also considered
S. aluco and
B. bubo to be possible contributors to the assemblage [
49,
50]. The high proportion of arvicoline teeth in the assemblage may be indicative of certain predators with a preference for this taxon. Several avian predators have been reported as preferring arvicolids, in particular
Asio otus,
A. flammeus, and
Strix nebulosa [
40]. The latter is not currently found in the region, but both
A. otus and
A. flammeus are known to visit the region in winter [
48].
Although no owl pellets were collected from the local area of Shanidar Cave, a recent taphonomic study of
T. alba was carried out in Birecik, Turkey, approximately 550 km west of Shanidar Cave [
51]. The study found that the
T. alba prey assemblage was dominated by
Meriones tristrami, followed by
Mus musculus. Species from both genera were also recovered from the Shanidar Cave assemblage (see
Table 1), and a similarly low level of digestion intensity was observed in both assemblages.
Microtus dominates the Shanidar Cave assemblage, and was not recovered from the Birecik
T. alba prey assemblage.
Microtus is known to be taken by
T. alba in the wider region; a 2006 study of
T. alba pellets in the Beit She’an Valley (approximately 890 km southwest of Shanidar Cave) found that
Microtus accounted for 19.9% of the total prey MNI, with
Meriones accounting for 32.3% and
Mus 37.3% [
52]. The difference between the two studies is unlikely to be due to season of capture, as the Birecik pellets were collected in June and the Beit She’an in July. The Beit She’an study noted a large difference in recovery of
Microtus between different study sites that was not observed in
Mus or
Meriones, and so it is possible that the lack of
Microtus in the Birecik study is due to a lack of vole populations in the study area.
T. alba is a category 1 predator that produces absent or minimal digestion of the microfaunal remains. This broadly matches the pattern seen in the Shanidar Cave assemblage. Occasional specimens with more intense digestion have been recorded in the Shanidar Cave assemblage, though as demonstrated by Williams [
53], there is variation in digestion intensity between different
T. alba individuals depending on age and other factors.
The results of the taphonomic analysis with respect to predation are not conclusive. The digestion patterns, landscape, and predators present in the region today suggest that the eagle owl is the most likely predator responsible for assemblage accumulation out of the avian species listed above, though the digestion intensities reported in the literature are slightly higher than those observed here. It should be recognised that the assemblage is being analysed over very large temporal ranges, and it is possible that other predators occasionally contributed to the assemblage at different points, which may explain the occasional teeth and bones displaying more intense digestion. As the agent of accumulation is uncertain, it is difficult to be confident that the community composition observed in any palaeoecological analyses is a genuine ecological rather than taphonomic signal. The taxonomic identification of microvertebrate specimens is outside the scope of this paper, but generally speaking, arvicolines dominate the assemblage. Arvicolines are known to dominate the diet of several avian predators [
44], but can also reach very high population densities in the landscape, and so it is difficult to know whether their presence is due to predator bias. Further studies of predator distribution, hunting behaviour, and preferences in the region are needed to resolve this problem.
Occasional animal burrows have been observed during excavation. Some of these are likely to have occurred during and following the 1950s excavations, when the trench walls were more accessible. Burrowing taxa are present in the preliminary faunal list, for example, Microtus, Meriones, and Ellobius, and may be responsible for some of these burrows, though many of these taxa more commonly construct burrows in open areas rather than within caves. Repeated rockfall throughout the sequence has resulted in very rocky sediments that may be unsuitable for some burrowing rodent taxa. Further data on the shape, depth and, where possible, age, of these burrows, and the palynological data from their sediments, will help to determine which are modern and which taxa may be responsible.















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}