
High-Impact AMPAkines Elevate Calcium Levels in Cortical Astrocytes by Mobilizing Endoplasmic Reticular Calcium Stores


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Solution Preparation
2.2. Astrocyte Cultures
2.3. Calcium Fluorescence Measurements
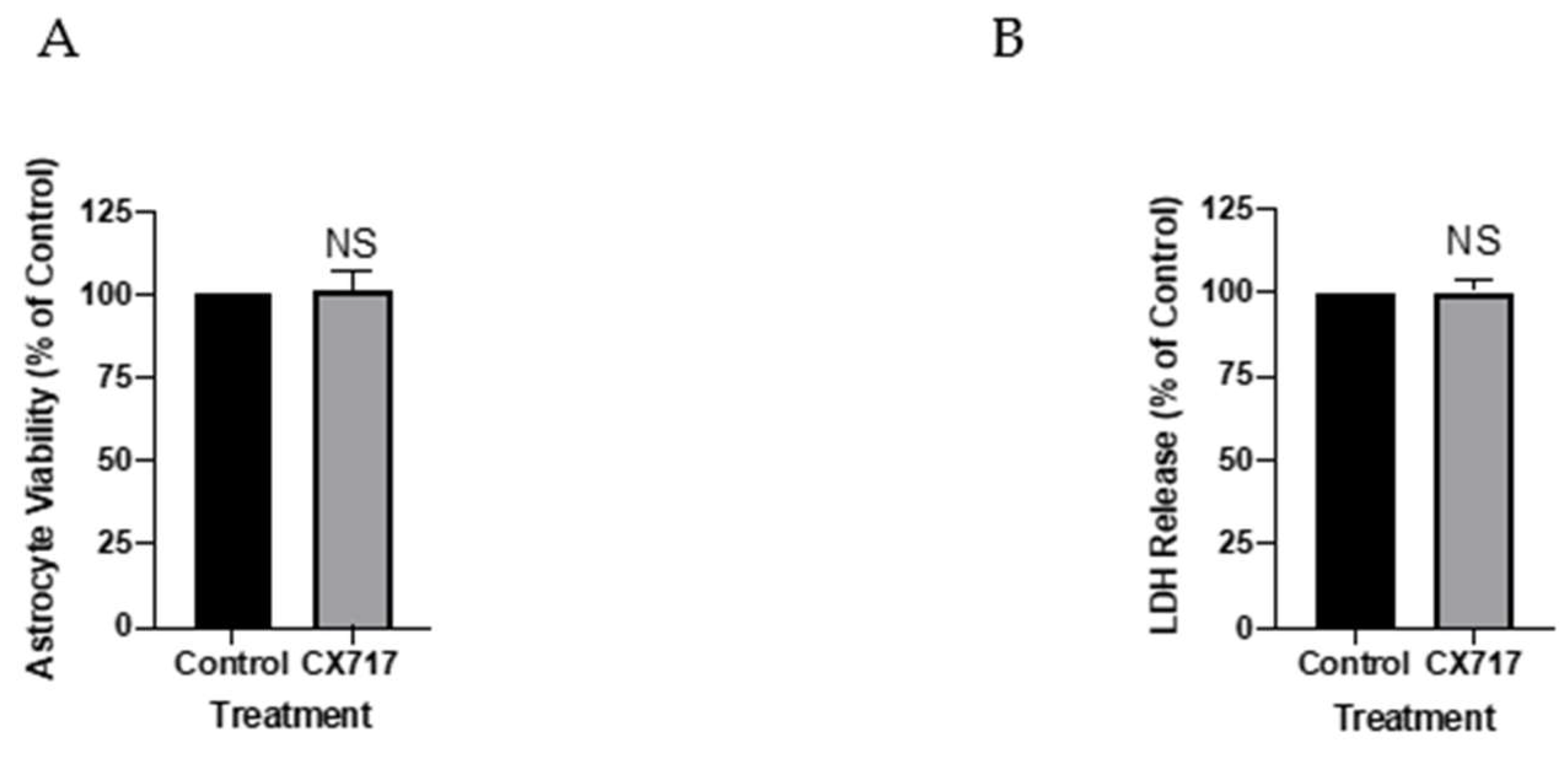
2.4. Assessment of CX717 Toxicity in Cortical Astrocytes
2.5. Statistical Analysis
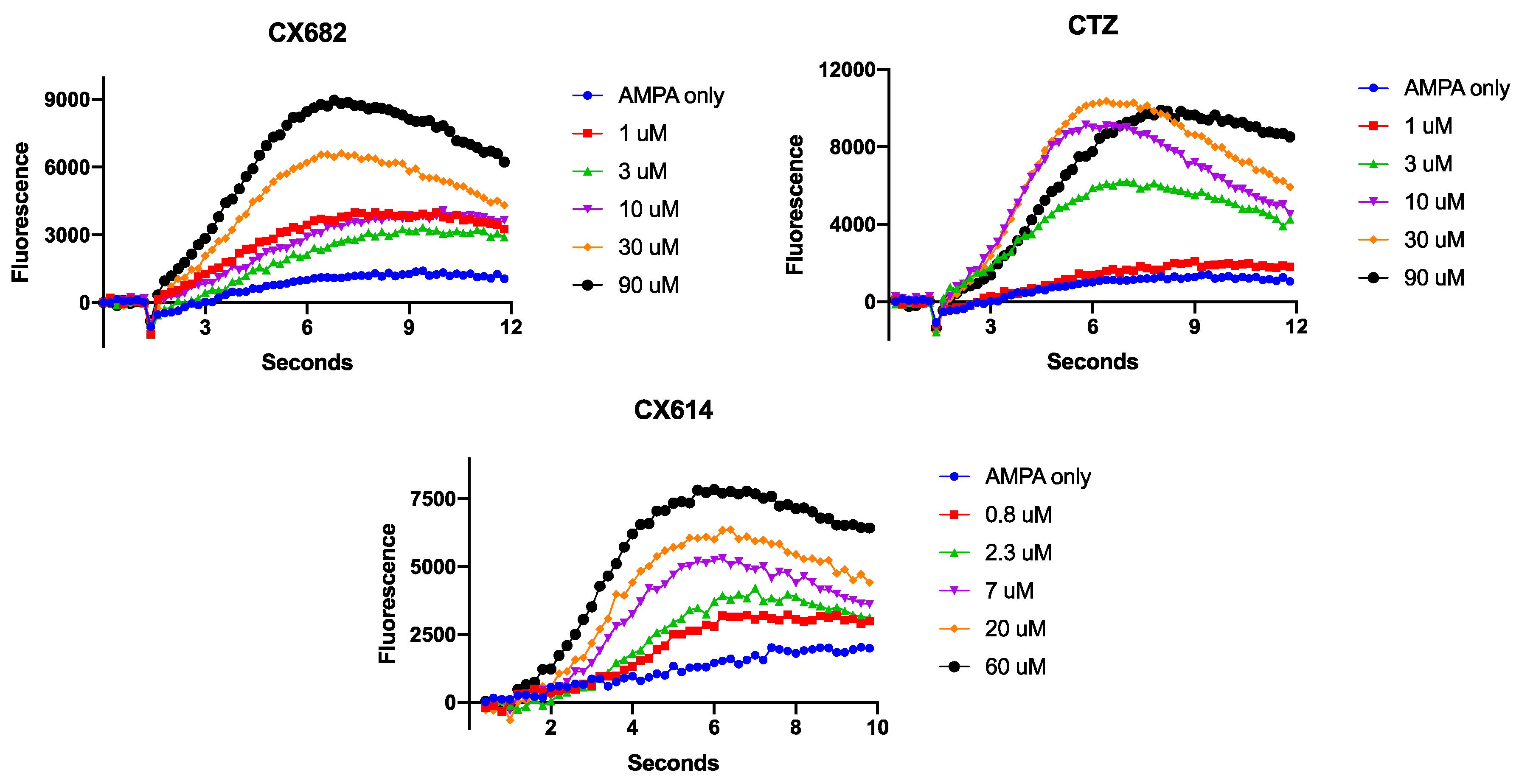
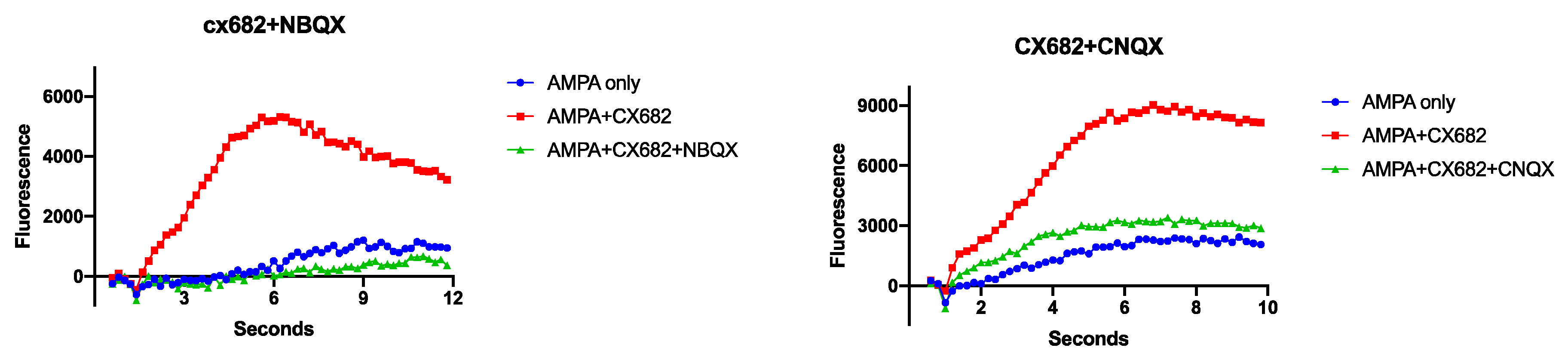
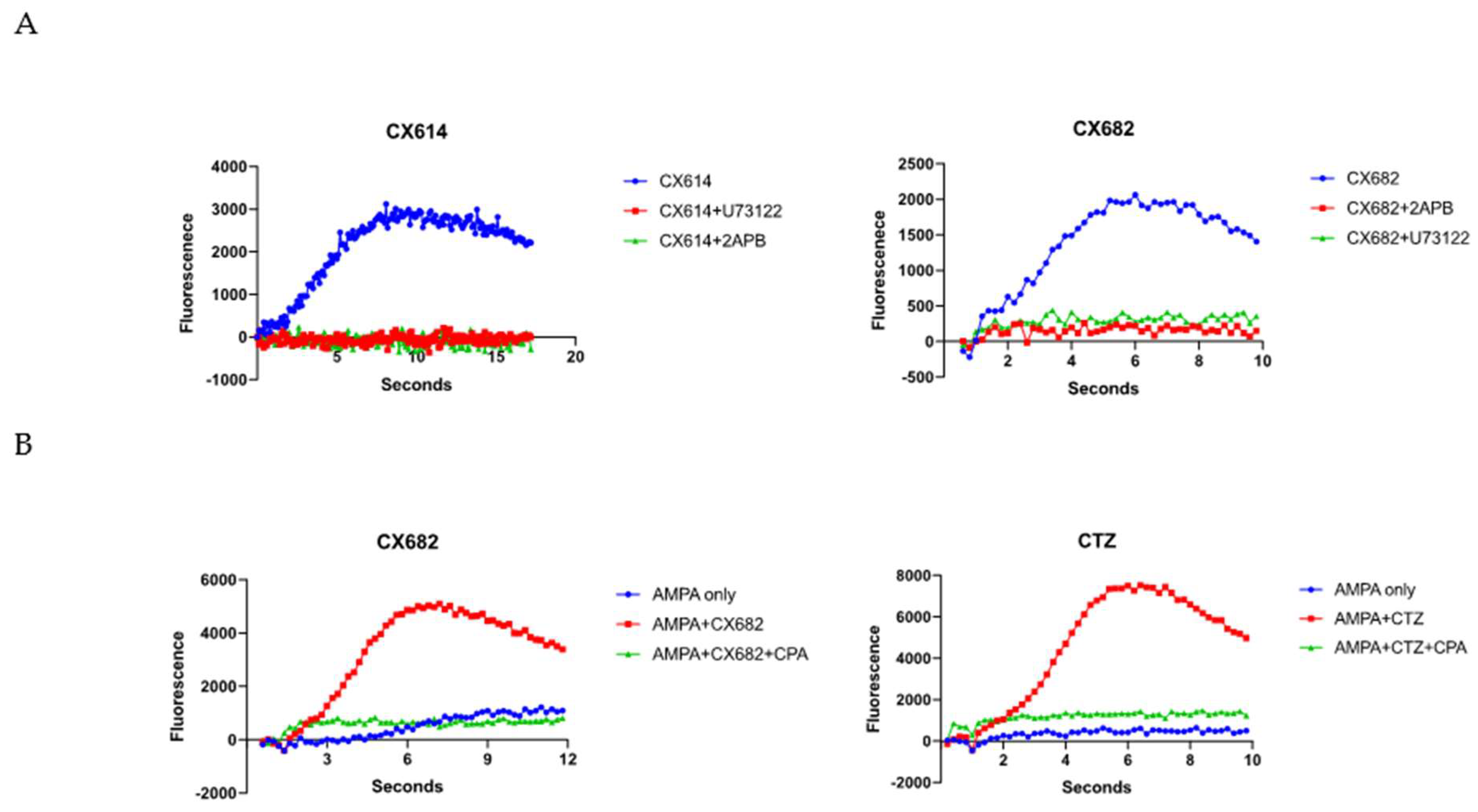
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Brandebura, A.N.; Paumier, A.; Onur, T.S.; Allen, N.J. Astrocyte contribution to dysfunction, risk and progression in neurodegenerative disorders. Nat. Rev. Neurosci. 2023, 24, 23–39. [Google Scholar] [CrossRef] [PubMed]
- Profaci, C.P.; Munji, R.N.; Pulido, R.S.; Daneman, R. The blood-brain barrier in health and disease: Important unanswered questions. J. Exp. Med. 2020, 217, e20190062. [Google Scholar] [CrossRef] [PubMed]
- Christopherson, K.S.; Ullian, E.M.; Stokes, C.C.; Mullowney, C.E.; Hell, J.W.; Agah, A.; Lawler, J.; Mosher, D.F.; Bornstein, P.; Barres, B.A. Thrombospondins are astrocyte-secreted proteins that promote CNS synaptogenesis. Cell 2005, 120, 421–433. [Google Scholar] [CrossRef]
- Allen, N.J.; Bennett, M.L.; Foo, L.C.; Wang, G.X.; Chakraborty, C.; Smith, S.J.; Barres, B.A. Astrocyte glypicans 4 and 6 promote formation of excitatory synapses via GluA1 AMPA receptors. Nature 2012, 486, 410–414. [Google Scholar] [CrossRef]
- Blanco-Suarez, E.; Liu, T.F.; Kopelevich, A.; Allen, N.J. Astrocyte-Secreted Chordin-like 1 Drives Synapse Maturation and Limits Plasticity by Increasing Synaptic GluA2 AMPA Receptors. Neuron 2018, 100, 1116–1132 e1113. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Feng, X.; Wang, Y.; Xia, X.; Zheng, J.C. Astrocytes: GABAceptive and GABAergic Cells in the Brain. Front. Cell. Neurosci. 2022, 16, 892497. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.P.; Cunningham, L.A.; Partridge, L.D. Coupling of AMPA receptors with the Na+/Ca2+ exchanger in cultured rat astrocytes. Brain Res. 2000, 887, 98–109. [Google Scholar] [CrossRef]
- Pellerin, L.; Magistretti, P.J. Ampakine CX546 bolsters energetic response of astrocytes: A novel target for cognitive-enhancing drugs acting as alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) receptor modulators. J. Neurochem. 2005, 92, 668–677. [Google Scholar] [CrossRef]
- Parpura, V.; Scemes, E.; Spray, D.C. Mechanisms of glutamate release from astrocytes: Gap junction “hemichannels”, purinergic receptors and exocytotic release. Neurochem. Int. 2004, 45, 259–264. [Google Scholar] [CrossRef]
- Xu, J.; Peng, H.; Kang, N.; Zhao, Z.; Lin, J.H.; Stanton, P.K.; Kang, J. Glutamate-induced exocytosis of glutamate from astrocytes. J. Biol. Chem. 2007, 282, 24185–24197. [Google Scholar] [CrossRef]
- Collingridge, G.L.; Singer, W. Excitatory amino acid receptors and synaptic plasticity. Trends Pharmacol. Sci. 1990, 11, 290–296. [Google Scholar] [CrossRef]
- Wisden, W.; Seeburg, P.H. Mammalian ionotropic glutamate receptors. Curr. Opin. Neurobiol. 1993, 3, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Arai, A.C.; Kessler, M. Pharmacology of ampakine modulators: From AMPA receptors to synapses and behavior. Curr. Drug Targets 2007, 8, 583–602. [Google Scholar] [CrossRef] [PubMed]
- Arai, A.C.; Xia, Y.F.; Suzuki, E. Modulation of AMPA receptor kinetics differentially influences synaptic plasticity in the hippocampus. Neuroscience 2004, 123, 1011–1024. [Google Scholar] [CrossRef]
- Lynch, G.; Gall, C.M. Ampakines and the threefold path to cognitive enhancement. Trends Neurosci. 2006, 29, 554–562. [Google Scholar] [CrossRef]
- O’Neill, M.J.; Dix, S. AMPA receptor potentiators as cognitive enhancers. IDrugs Investig. Drugs J. 2007, 10, 185–192. [Google Scholar]
- Lynch, G.; Palmer, L.C.; Gall, C.M. The likelihood of cognitive enhancement. Pharmacol. Biochem. Behav. 2011, 99, 116–129. [Google Scholar] [CrossRef]
- Radin, D.P.; Zhong, S.; Purcell, R.; Lippa, A. Acute ampakine treatment ameliorates age-related deficits in long-term potentiation. Biomed. Pharmacother. 2016, 84, 806–809. [Google Scholar] [CrossRef]
- Ikonomovic, M.D.; Mizukami, K.; Davies, P.; Hamilton, R.; Sheffield, R.; Armstrong, D.M. The loss of GluR2(3) immunoreactivity precedes neurofibrillary tangle formation in the entorhinal cortex and hippocampus of Alzheimer brains. J. Neuropathol. Exp. Neurol. 1997, 56, 1018–1027. [Google Scholar] [CrossRef]
- Dirson, G.; Desjardins, P.; Tannenberg, T.; Dodd, P.; Butterworth, R.F. Selective loss of expression of glutamate GluR2/R3 receptor subunits in cerebellar tissue from a patient with olivopontocerebellar atrophy. Metab. Brain Dis. 2002, 17, 77–82. [Google Scholar] [CrossRef]
- Porrino, L.J.; Daunais, J.B.; Rogers, G.A.; Hampson, R.E.; Deadwyler, S.A. Facilitation of task performance and removal of the effects of sleep deprivation by an ampakine (CX717) in nonhuman primates. PLoS Biol. 2005, 3, e299. [Google Scholar] [CrossRef] [PubMed]
- Wright, A.; Vissel, B. The essential role of AMPA receptor GluR2 subunit RNA editing in the normal and diseased brain. Front. Mol. Neurosci. 2012, 5, 34. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.; Han, H.; Li, H.; Bai, Y.; Wang, W.; Tu, M.; Peng, Y.; Zhou, L.; He, W.; Wu, X.; et al. Long-term potentiation decay and memory loss are mediated by AMPAR endocytosis. J. Clin. Investig. 2015, 125, 234–247. [Google Scholar] [CrossRef] [PubMed]
- Hampson, R.E.; Rogers, G.; Lynch, G.; Deadwyler, S.A. Facilitative effects of the ampakine CX516 on short-term memory in rats: Correlations with hippocampal neuronal activity. J. Neurosci. Off. J. Soc. Neurosci. 1998, 18, 2748–2763. [Google Scholar] [CrossRef] [PubMed]
- Hampson, R.E.; Rogers, G.; Lynch, G.; Deadwyler, S.A. Facilitative effects of the ampakine CX516 on short-term memory in rats: Enhancement of delayed-nonmatch-to-sample performance. J. Neurosci. Off. J. Soc. Neurosci. 1998, 18, 2740–2747. [Google Scholar] [CrossRef]
- Radin, D.P.; Zhong, S.; Cerne, R.; Shoaib, M.; Witkin, J.M.; Lippa, A. Low-Impact Ampakine CX1739 Exerts Pro-Cognitive Effects and Reverses Opiate-Induced Respiratory Depression in Rodents. Future Pharmacol. 2024, 4, 173–187. [Google Scholar] [CrossRef]
- Ingvar, M.; Ambros-Ingerson, J.; Davis, M.; Granger, R.; Kessler, M.; Rogers, G.A.; Schehr, R.S.; Lynch, G. Enhancement by an ampakine of memory encoding in humans. Exp. Neurol. 1997, 146, 553–559. [Google Scholar] [CrossRef]
- Ogier, M.; Wang, H.; Hong, E.; Wang, Q.; Greenberg, M.E.; Katz, D.M. Brain-derived neurotrophic factor expression and respiratory function improve after ampakine treatment in a mouse model of Rett syndrome. J. Neurosci. Off. J. Soc. Neurosci. 2007, 27, 10912–10917. [Google Scholar] [CrossRef]
- Simmons, D.A.; Rex, C.S.; Palmer, L.; Pandyarajan, V.; Fedulov, V.; Gall, C.M.; Lynch, G. Up-regulating BDNF with an ampakine rescues synaptic plasticity and memory in Huntington’s disease knockin mice. Proc. Natl. Acad. Sci. USA 2009, 106, 4906–4911. [Google Scholar] [CrossRef]
- Jourdi, H.; Hamo, L.; Oka, T.; Seegan, A.; Baudry, M. BDNF mediates the neuroprotective effects of positive AMPA receptor modulators against MPP+-induced toxicity in cultured hippocampal and mesencephalic slices. Neuropharmacology 2009, 56, 876–885. [Google Scholar] [CrossRef]
- Simmons, D.A.; Mehta, R.A.; Lauterborn, J.C.; Gall, C.M.; Lynch, G. Brief ampakine treatments slow the progression of Huntington’s disease phenotypes in R6/2 mice. Neurobiol. Dis. 2011, 41, 436–444. [Google Scholar] [CrossRef]
- Xu, X.J.; Hao, J.X.; Seiger, A.; Wiesenfeld-Hallin, Z. Systemic excitatory amino acid receptor antagonists of the alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) receptor and of the N-methyl-D-aspartate (NMDA) receptor relieve mechanical hypersensitivity after transient spinal cord ischemia in rats. J. Pharmacol. Exp. Ther. 1993, 267, 140–144. [Google Scholar]
- Wollman, L.B.; Streeter, K.A.; Fusco, A.F.; Gonzalez-Rothi, E.J.; Sandhu, M.S.; Greer, J.J.; Fuller, D.D. Ampakines stimulate phrenic motor output after cervical spinal cord injury. Exp. Neurol. 2020, 334, 113465. [Google Scholar] [CrossRef] [PubMed]
- Rana, S.; Sunshine, M.D.; Greer, J.J.; Fuller, D.D. Ampakines Stimulate Diaphragm Activity after Spinal Cord Injury. J. Neurotrauma 2021, 38, 3467–3482. [Google Scholar] [CrossRef]
- Witkin, J.M.; Radin, D.P.; Rana, S.; Fuller, D.D.; Fusco, A.F.; Demers, J.C.; Pradeep Thakre, P.; Smith, J.L.; Lippa, A.; Cerne, R. AMPA receptors play an important role in the biological consequences of spinal cord injury: Implications for AMPA receptor modulators for therapeutic benefit. Biochem. Pharmacol. 2024, 116302. [Google Scholar] [CrossRef] [PubMed]
- Rana, S.; Alom, F.; Martinez, R.C.; Fuller, D.D.; Mickle, A.D. Acute ampakines increase voiding function and coordination in a rat model of SCI. Elife 2024, 12, RP89767. [Google Scholar] [CrossRef] [PubMed]
- Rana, S.; Thakre, P.P.; Fuller, D.D. Ampakines increase diaphragm activation following mid-cervical contusion injury in rats. Exp. Neurol. 2024, 376, 114769. [Google Scholar] [CrossRef]
- Arai, A.C.; Xia, Y.F.; Rogers, G.; Lynch, G.; Kessler, M. Benzamide-type AMPA receptor modulators form two subfamilies with distinct modes of action. J. Pharmacol. Exp. Ther. 2002, 303, 1075–1085. [Google Scholar] [CrossRef]
- Arai, A.C.; Kessler, M.; Rogers, G.; Lynch, G. Effects of the potent ampakine CX614 on hippocampal and recombinant AMPA receptors: Interactions with cyclothiazide and GYKI 52466. Mol. Pharmacol. 2000, 58, 802–813. [Google Scholar] [CrossRef]
- Kessler, M.; Arai, A.C. Use of [3H]fluorowillardiine to study properties of AMPA receptor allosteric modulators. Brain Res. 2006, 1076, 25–41. [Google Scholar] [CrossRef]
- Montgomery, K.E.; Kessler, M.; Arai, A.C. Modulation of agonist binding to AMPA receptors by 1-(1,4-benzodioxan-6-ylcarbonyl)piperidine (CX546): Differential effects across brain regions and GluA1-4/transmembrane AMPA receptor regulatory protein combinations. J. Pharmacol. Exp. Ther. 2009, 331, 965–974. [Google Scholar] [CrossRef] [PubMed]
- Purcell, R.; Lynch, G.; Gall, C.; Johnson, S.; Sheng, Z.; Stephen, M.R.; Cook, J.; Garman, R.H.; Jortner, B.; Bolon, B.; et al. Brain Vacuolation Resulting From Administration of the Type II Ampakine CX717 Is An Artifact Related to Molecular Structure and Chemical Reaction With Tissue Fixative Agents. Toxicol. Sci. 2018, 162, 383–395. [Google Scholar] [CrossRef]
- Radin, D.P.; Zhong, S.; Cerne, R.; Witkin, J.; Lippa, A. High impact AMPAkines induce a Gq-protein coupled endoplasmic calcium release in cortical neurons: A possible mechanism for explaining the toxicity of high impact AMPAkines. bioRxiv 2024. [Google Scholar] [CrossRef]
- Schildge, S.; Bohrer, C.; Beck, K.; Schachtrup, C. Isolation and culture of mouse cortical astrocytes. J. Vis. Exp. 2013. [Google Scholar] [CrossRef]
- David, J.C.; Yamada, K.A.; Bagwe, M.R.; Goldberg, M.P. AMPA receptor activation is rapidly toxic to cortical astrocytes when desensitization is blocked. J. Neurosci. Off. J. Soc. Neurosci. 1996, 16, 200–209. [Google Scholar] [CrossRef]
- Oertel, B.G.; Felden, L.; Tran, P.V.; Bradshaw, M.H.; Angst, M.S.; Schmidt, H.; Johnson, S.; Greer, J.J.; Geisslinger, G.; Varney, M.A.; et al. Selective antagonism of opioid-induced ventilatory depression by an ampakine molecule in humans without loss of opioid analgesia. Clin. Pharmacol. Ther. 2010, 87, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Ding, X.; Funk, G.D.; Greer, J.J. Ampakine CX717 protects against fentanyl-induced respiratory depression and lethal apnea in rats. Anesthesiology 2009, 110, 1364–1370. [Google Scholar] [CrossRef]
- Ren, J.; Ding, X.; Greer, J.J. Respiratory depression in rats induced by alcohol and barbiturate and rescue by ampakine CX717. J. Appl. Physiol. 2012, 113, 1004–1011. [Google Scholar] [CrossRef]
- Partin, K.M.; Fleck, M.W.; Mayer, M.L. AMPA receptor flip/flop mutants affecting deactivation, desensitization, and modulation by cyclothiazide, aniracetam, and thiocyanate. J. Neurosci. Off. J. Soc. Neurosci. 1996, 16, 6634–6647. [Google Scholar] [CrossRef]
- Fleck, M.W.; Bahring, R.; Patneau, D.K.; Mayer, M.L. AMPA receptor heterogeneity in rat hippocampal neurons revealed by differential sensitivity to cyclothiazide. J. Neurophysiol. 1996, 75, 2322–2333. [Google Scholar] [CrossRef]
- Rammes, G.; Swandulla, D.; Spielmanns, P.; Parsons, C.G. Interactions of GYKI 52466 and NBQX with cyclothiazide at AMPA receptors: Experiments with outside-out patches and EPSCs in hippocampal neurones. Neuropharmacology 1998, 37, 1299–1320. [Google Scholar] [CrossRef] [PubMed]
- Lauterborn, J.C.; Lynch, G.; Vanderklish, P.; Arai, A.; Gall, C.M. Positive modulation of AMPA receptors increases neurotrophin expression by hippocampal and cortical neurons. J. Neurosci. Off. J. Soc. Neurosci. 2000, 20, 8–21. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Lenal, F.; Yang, M.; Ding, X.; Greer, J.J. Coadministration of the AMPAKINE CX717 with propofol reduces respiratory depression and fatal apneas. Anesthesiology 2013, 118, 1437–1445. [Google Scholar] [CrossRef]
- Akasaka, T.; Balasas, T.; Russell, L.J.; Sugimoto, K.J.; Majid, A.; Walewska, R.; Karran, E.L.; Brown, D.G.; Cain, K.; Harder, L.; et al. Five members of the CEBP transcription factor family are targeted by recurrent IGH translocations in B-cell precursor acute lymphoblastic leukemia (BCP-ALL). Blood 2007, 109, 3451–3461. [Google Scholar] [CrossRef] [PubMed]
- Molders, A.; Koch, A.; Menke, R.; Klocker, N. Heterogeneity of the astrocytic AMPA-receptor transcriptome. Glia 2018, 66, 2604–2616. [Google Scholar] [CrossRef]
- Priel, A.; Kolleker, A.; Ayalon, G.; Gillor, M.; Osten, P.; Stern-Bach, Y. Stargazin reduces desensitization and slows deactivation of the AMPA-type glutamate receptors. J. Neurosci. Off. J. Soc. Neurosci. 2005, 25, 2682–2686. [Google Scholar] [CrossRef]
- Rouach, N.; Byrd, K.; Petralia, R.S.; Elias, G.M.; Adesnik, H.; Tomita, S.; Karimzadegan, S.; Kealey, C.; Bredt, D.S.; Nicoll, R.A. TARP gamma-8 controls hippocampal AMPA receptor number, distribution and synaptic plasticity. Nat. Neurosci. 2005, 8, 1525–1533. [Google Scholar] [CrossRef]
- Nakagawa, T.; Cheng, Y.; Sheng, M.; Walz, T. Three-dimensional structure of an AMPA receptor without associated stargazin/TARP proteins. Biol. Chem. 2006, 387, 179–187. [Google Scholar] [CrossRef]
- Tomita, S.; Byrd, R.K.; Rouach, N.; Bellone, C.; Venegas, A.; O’Brien, J.L.; Kim, K.S.; Olsen, O.; Nicoll, R.A.; Bredt, D.S. AMPA receptors and stargazin-like transmembrane AMPA receptor-regulatory proteins mediate hippocampal kainate neurotoxicity. Proc. Natl. Acad. Sci. USA 2007, 104, 18784–18788. [Google Scholar] [CrossRef]
- Cho, C.H.; St-Gelais, F.; Zhang, W.; Tomita, S.; Howe, J.R. Two families of TARP isoforms that have distinct effects on the kinetic properties of AMPA receptors and synaptic currents. Neuron 2007, 55, 890–904. [Google Scholar] [CrossRef]
- Kato, A.S.; Zhou, W.; Milstein, A.D.; Knierman, M.D.; Siuda, E.R.; Dotzlaf, J.E.; Yu, H.; Hale, J.E.; Nisenbaum, E.S.; Nicoll, R.A.; et al. New transmembrane AMPA receptor regulatory protein isoform, gamma-7, differentially regulates AMPA receptors. J. Neurosci. Off. J. Soc. Neurosci. 2007, 27, 4969–4977. [Google Scholar] [CrossRef] [PubMed]
- Milstein, A.D.; Zhou, W.; Karimzadegan, S.; Bredt, D.S.; Nicoll, R.A. TARP subtypes differentially and dose-dependently control synaptic AMPA receptor gating. Neuron 2007, 55, 905–918. [Google Scholar] [CrossRef]
- Milstein, A.D.; Nicoll, R.A. Regulation of AMPA receptor gating and pharmacology by TARP auxiliary subunits. Trends Pharmacol. Sci. 2008, 29, 333–339. [Google Scholar] [CrossRef]
- Payne, H.L. The role of transmembrane AMPA receptor regulatory proteins (TARPs) in neurotransmission and receptor trafficking (Review). Mol. Membr. Biol. 2008, 25, 353–362. [Google Scholar] [CrossRef]
- Menuz, K.; O’Brien, J.L.; Karmizadegan, S.; Bredt, D.S.; Nicoll, R.A. TARP redundancy is critical for maintaining AMPA receptor function. J. Neurosci. Off. J. Soc. Neurosci. 2008, 28, 8740–8746. [Google Scholar] [CrossRef]
- Suzuki, E.; Kessler, M.; Arai, A.C. The fast kinetics of AMPA GluR3 receptors is selectively modulated by the TARPs gamma 4 and gamma 8. Mol. Cell. Neurosci. 2008, 38, 117–123. [Google Scholar] [CrossRef]
- Morimoto-Tomita, M.; Zhang, W.; Straub, C.; Cho, C.H.; Kim, K.S.; Howe, J.R.; Tomita, S. Autoinactivation of neuronal AMPA receptors via glutamate-regulated TARP interaction. Neuron 2009, 61, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Soto, D.; Coombs, I.D.; Renzi, M.; Zonouzi, M.; Farrant, M.; Cull-Candy, S.G. Selective regulation of long-form calcium-permeable AMPA receptors by an atypical TARP, gamma-5. Nat. Neurosci. 2009, 12, 277–285. [Google Scholar] [CrossRef]
- Shi, Y.; Lu, W.; Milstein, A.D.; Nicoll, R.A. The stoichiometry of AMPA receptors and TARPs varies by neuronal cell type. Neuron 2009, 62, 633–640. [Google Scholar] [CrossRef] [PubMed]
- Menuz, K.; Kerchner, G.A.; O’Brien, J.L.; Nicoll, R.A. Critical role for TARPs in early development despite broad functional redundancy. Neuropharmacology 2009, 56, 22–29. [Google Scholar] [CrossRef]
- Kato, A.S.; Gill, M.B.; Ho, M.T.; Yu, H.; Tu, Y.; Siuda, E.R.; Wang, H.; Qian, Y.W.; Nisenbaum, E.S.; Tomita, S.; et al. Hippocampal AMPA receptor gating controlled by both TARP and cornichon proteins. Neuron 2010, 68, 1082–1096. [Google Scholar] [CrossRef]
- Jackson, A.C.; Nicoll, R.A. Stargazin (TARP gamma-2) is required for compartment-specific AMPA receptor trafficking and synaptic plasticity in cerebellar stellate cells. J. Neurosci. Off. J. Soc. Neurosci. 2011, 31, 3939–3952. [Google Scholar] [CrossRef] [PubMed]
- Boyle, J.; Stanley, N.; James, L.M.; Wright, N.; Johnsen, S.; Arbon, E.L.; Dijk, D.J. Acute sleep deprivation: The effects of the AMPAKINE compound CX717 on human cognitive performance, alertness and recovery sleep. J. Psychopharmacol. 2012, 26, 1047–1057. [Google Scholar] [CrossRef] [PubMed]
- Haw, A.J.; Meyer, L.C.; Greer, J.J.; Fuller, A. Ampakine CX1942 attenuates opioid-induced respiratory depression and corrects the hypoxaemic effects of etorphine in immobilized goats (Capra hircus). Vet. Anaesth. Analg. 2016, 43, 528–538. [Google Scholar] [CrossRef] [PubMed]
- Radin, D.P.; Johnson, S.; Purcell, R.; Lippa, A.S. Effects of chronic systemic low-impact ampakine treatment on neurotrophin expression in rat brain. Biomed. Pharmacother. 2018, 105, 540–544. [Google Scholar] [CrossRef]
- Wollman, L.B.; Streeter, K.A.; Fuller, D.D. Ampakine pretreatment enables a single brief hypoxic episode to evoke phrenic motor facilitation. J. Neurophysiol. 2020, 123, 993–1003. [Google Scholar] [CrossRef]
- Thakre, P.P.; Sunshine, M.D.; Fuller, D.D. Ampakine pretreatment enables a single hypoxic episode to produce phrenic motor facilitation with no added benefit of additional episodes. J. Neurophysiol. 2021, 126, 1420–1429. [Google Scholar] [CrossRef]
- Lorier, A.R.; Funk, G.D.; Greer, J.J. Opiate-induced suppression of rat hypoglossal motoneuron activity and its reversal by ampakine therapy. PLoS ONE 2010, 5, e8766. [Google Scholar] [CrossRef]
- Liu, H.N.; Molina-Holgado, E.; Almazan, G. Glutamate-stimulated production of inositol phosphates is mediated by Ca2+ influx in oligodendrocyte progenitors. Eur. J. Pharmacol. 1997, 338, 277–287. [Google Scholar] [CrossRef]
- Wang, Y.; Small, D.L.; Stanimirovic, D.B.; Morley, P.; Durkin, J.P. AMPA receptor-mediated regulation of a Gi-protein in cortical neurons. Nature 1997, 389, 502–504. [Google Scholar] [CrossRef]
- Kawai, F.; Sterling, P. AMPA receptor activates a G-protein that suppresses a cGMP-gated current. J. Neurosci. Off. J. Soc. Neurosci. 1999, 19, 2954–2959. [Google Scholar] [CrossRef]
- Marin, P.; Fagni, L.; Torrens, Y.; Alcaraz, G.; Couraud, F.; Bockaert, J.; Glowinski, J.; Premont, J. AMPA receptor activation induces association of G-beta protein with the alpha subunit of the sodium channel in neurons. Eur. J. Neurosci. 2001, 14, 1953–1960. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Radin, D.P.; Cerne, R.; Witkin, J.; Lippa, A. High-Impact AMPAkines Elevate Calcium Levels in Cortical Astrocytes by Mobilizing Endoplasmic Reticular Calcium Stores. Neuroglia 2024, 5, 344-355. https://doi.org/10.3390/neuroglia5030023
Radin DP, Cerne R, Witkin J, Lippa A. High-Impact AMPAkines Elevate Calcium Levels in Cortical Astrocytes by Mobilizing Endoplasmic Reticular Calcium Stores. Neuroglia. 2024; 5(3):344-355. https://doi.org/10.3390/neuroglia5030023
Chicago/Turabian StyleRadin, Daniel P., Rok Cerne, Jeffrey Witkin, and Arnold Lippa. 2024. "High-Impact AMPAkines Elevate Calcium Levels in Cortical Astrocytes by Mobilizing Endoplasmic Reticular Calcium Stores" Neuroglia 5, no. 3: 344-355. https://doi.org/10.3390/neuroglia5030023
APA StyleRadin, D. P., Cerne, R., Witkin, J., & Lippa, A. (2024). High-Impact AMPAkines Elevate Calcium Levels in Cortical Astrocytes by Mobilizing Endoplasmic Reticular Calcium Stores. Neuroglia, 5(3), 344-355. https://doi.org/10.3390/neuroglia5030023