
Plasma CXCL4–DNA/RNA Complexes and Anti-CXCL4 Antibodies Modulation in an SSc Cohort under Iloprost Treatment

 ,
, 
 and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design, SSc Patients and Healthy Donors (HD)
2.2. Reagents
2.3. ELISA for CXCL4 Measurement Plasma
2.4. ELISA for CXCL4–DNA and CXCL4–RNA Complex Detection in Plasma
2.5. ELISA for IFN-α and TNF-α Measurement in Plasma
2.6. Detection of Anti-CXCL4 Antibodies in Plasma
2.7. PDC Isolation and Stimulation
2.8. Statistical Analysis
3. Results
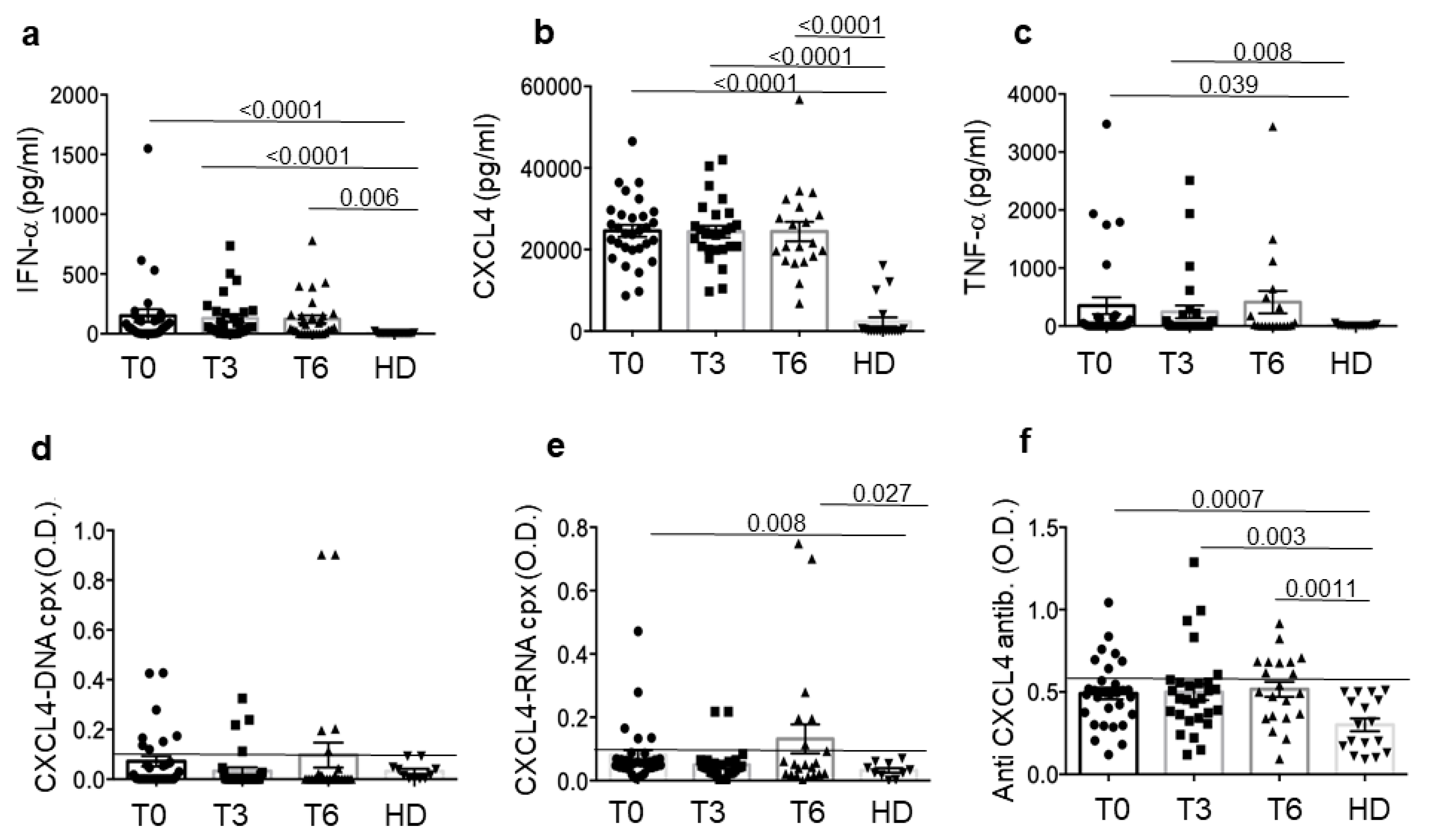
3.1. SSc Patients Show Up-Regulation of CXCL4 and Related Parameters Compared to Healthy Donors (HD)
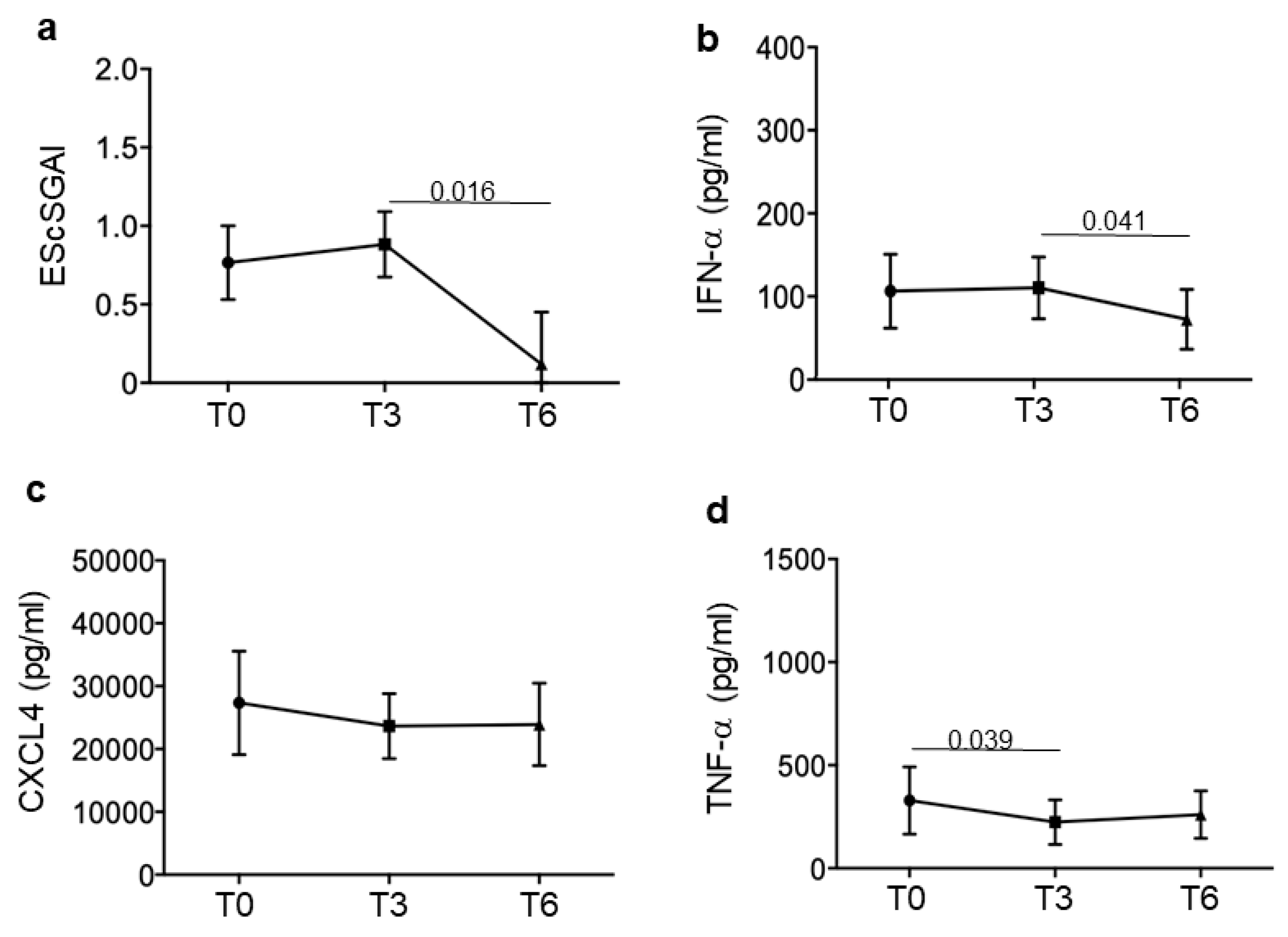
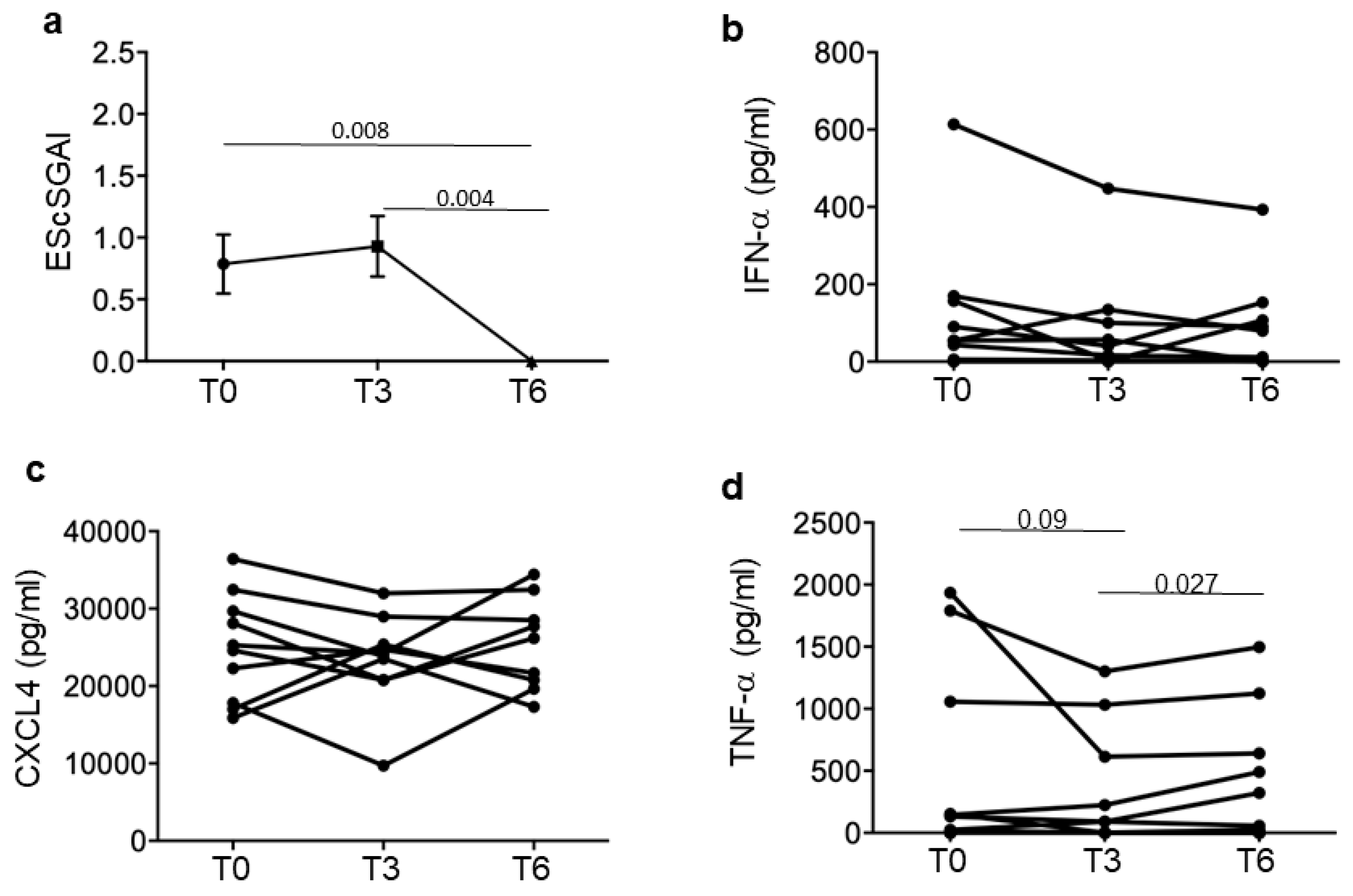
3.2. SSc Responders Reduce IFN-I Plasma Levels and TNF-α
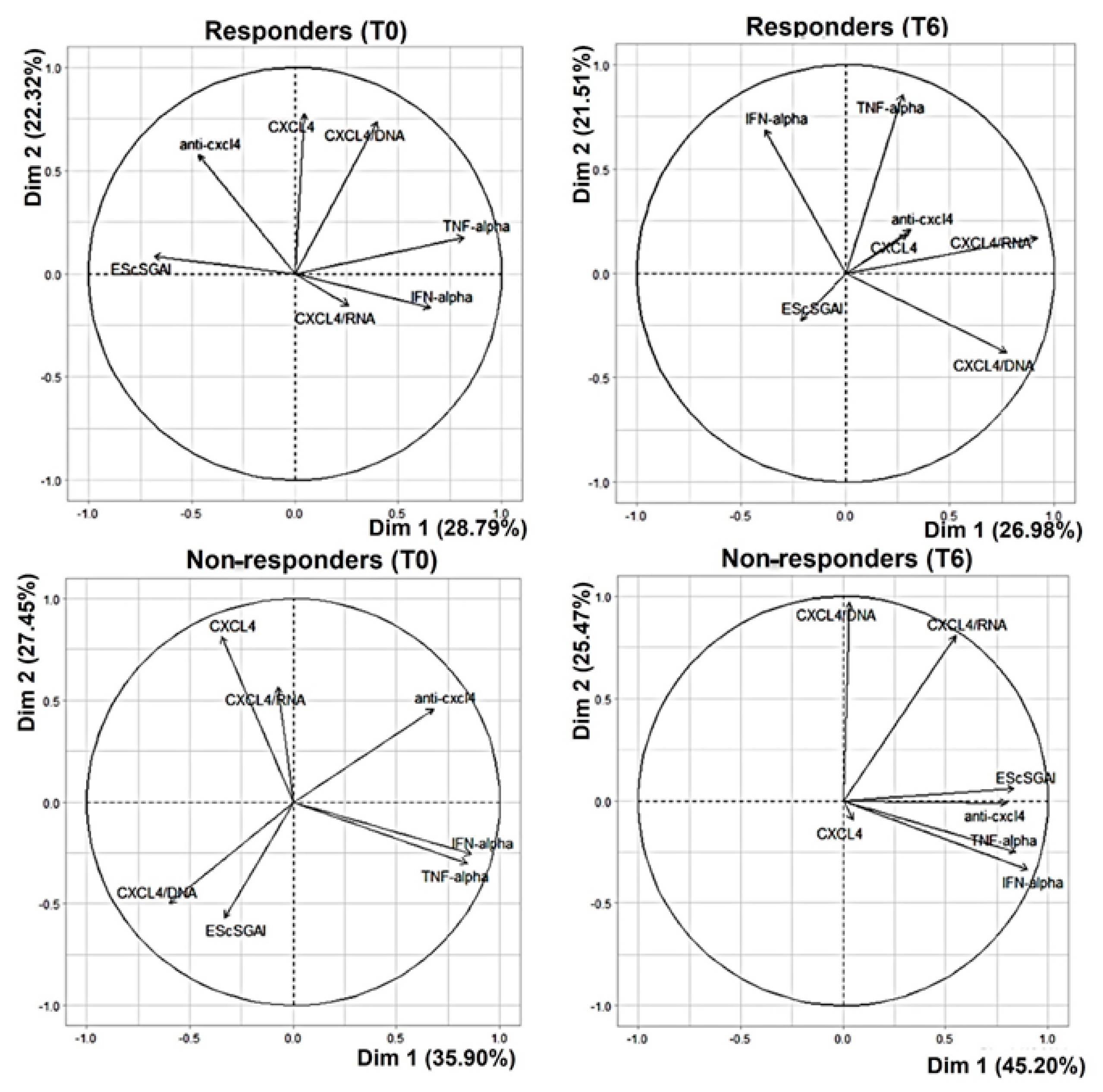
3.3. In the Non-Responders, CXCL4 and All Related Parameters Correlate with the EScSGAI
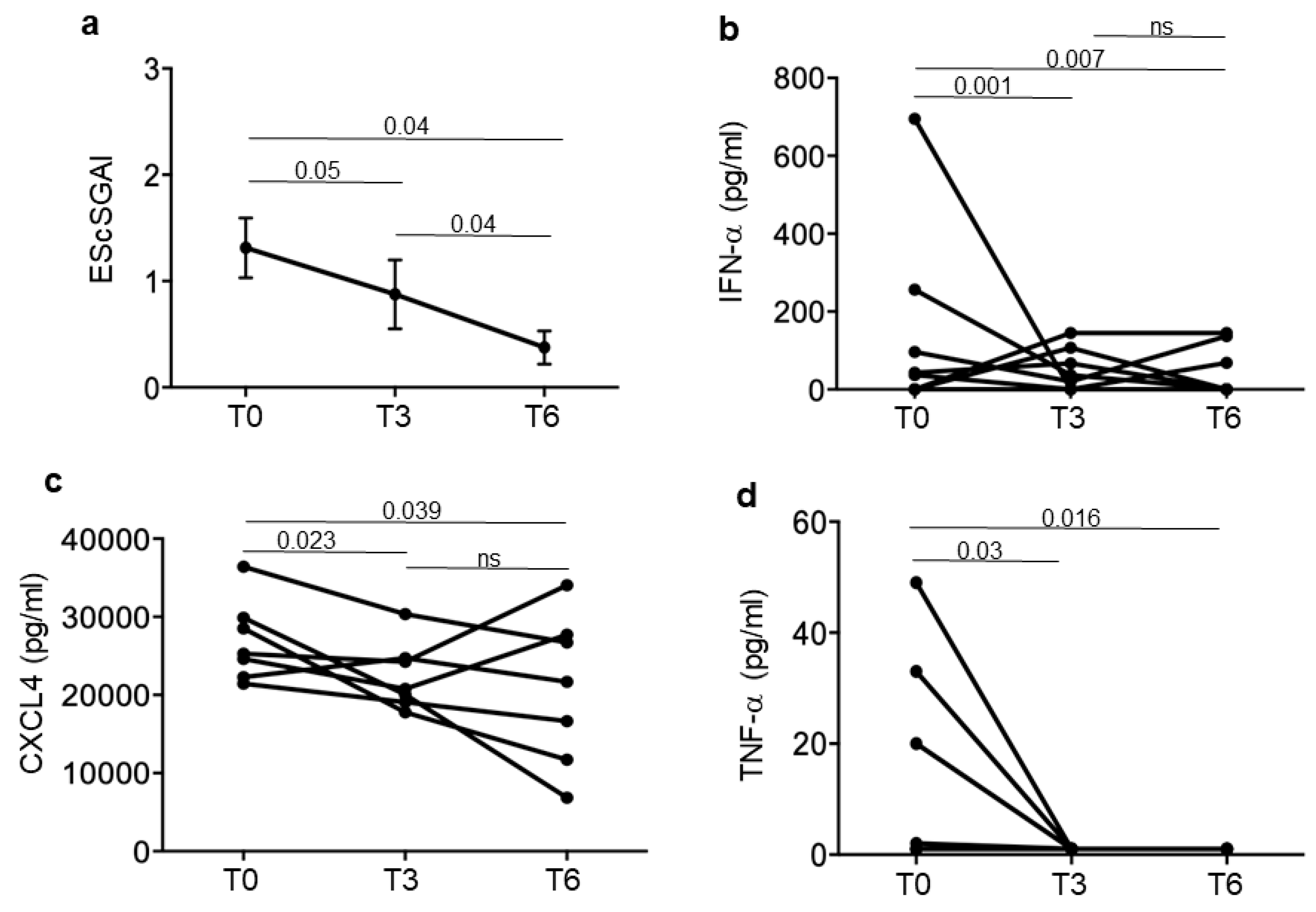
3.4. Iloprost Treatment Reduces Disease Activity in an SSc Subgroup with a Shorter Disease Duration
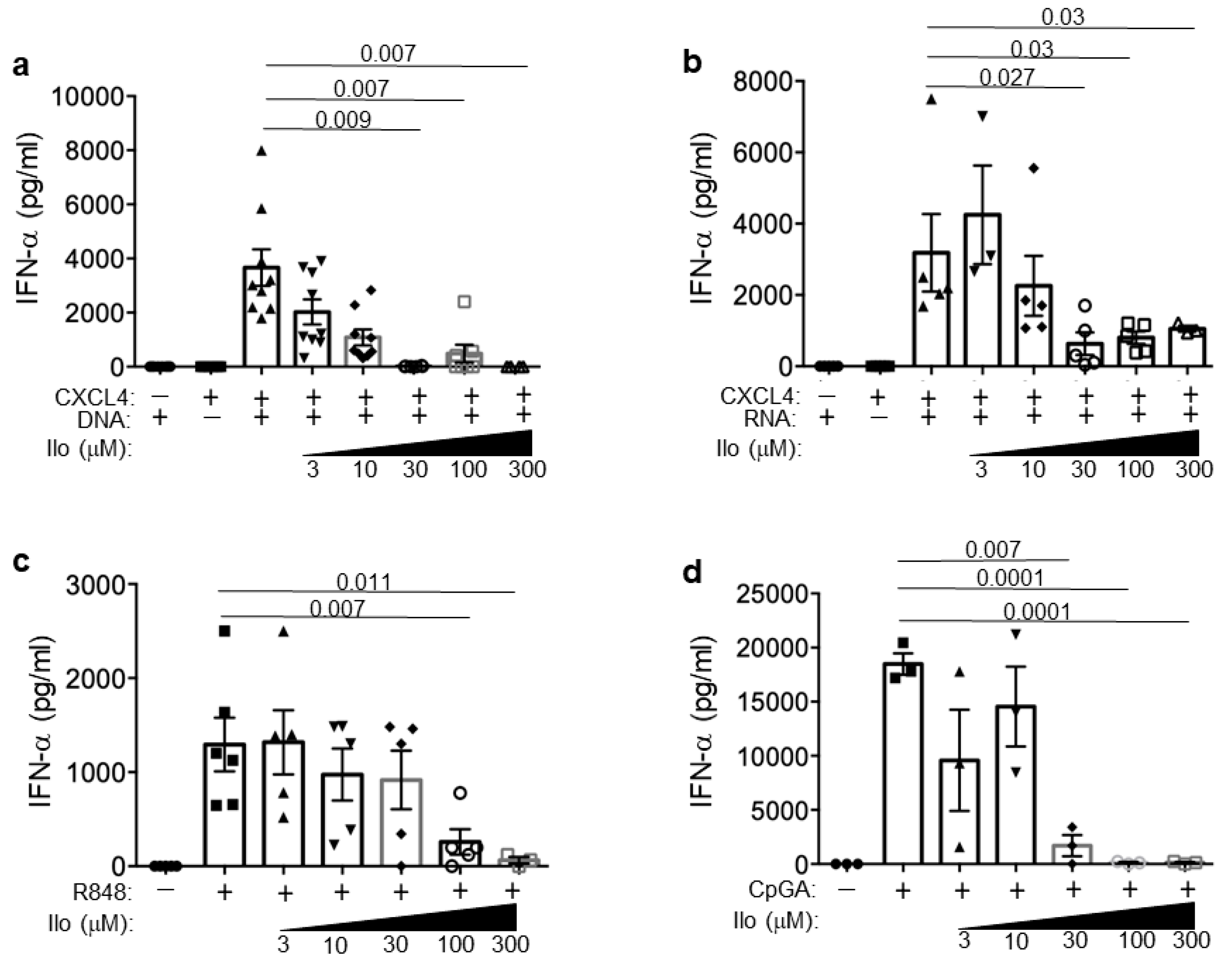
3.5. Iloprost Impairs pDCs Activation in the Presence of CXCL4–DNA/RNA Complexes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Abraham, D.J.; Krieg, T.; Distler, J.; Distler, O. Overview of Pathogenesis of Systemic Sclerosis. Rheumatology 2006, 48, iii3–iii7. [Google Scholar] [CrossRef] [PubMed]
- Cutolo, M.; Soldano, S.; Smith, V. Pathophysiology of Systemic Sclerosis: Current Understanding and New Insights. Expert Rev. Clin. Immunol. 2019, 15, 753–764. [Google Scholar] [CrossRef]
- LeROY, E.C.; Jr, T.A.M. Criteria for the Classification of Early Systemic Sclerosis. J. Rheumatol. 2001, 28, 1573–1576. [Google Scholar] [PubMed]
- Koenig, M.; Joyal, F.; Fritzler, M.J.; Roussin, A.; Abrahamowicz, M.; Boire, G.; Goulet, J.-R.; Rich, É.; Grodzicky, T.; Raymond, Y.; et al. Autoantibodies and Microvascular Damage Are Independent Predictive Factors for the Progression of Raynaud’s Phenomenon to Systemic Sclerosis: A Twenty-Year Prospective Study of 586 Patients, with Validation of Proposed Criteria for Early Systemic Sclerosis. Arthritis Rheum. 2008, 58, 3902–3912. [Google Scholar] [CrossRef] [PubMed]
- van den Hoogen, F.; Khanna, D.; Fransen, J.; Johnson, S.R.; Baron, M.; Tyndall, A.; Matucci-Cerinic, M.; Naden, R.P.; Medsger, T.A.; Carreira, P.E.; et al. 2013 Classification Criteria for Systemic Sclerosis: An American College of Rheumatology/European League against Rheumatism Collaborative Initiative. Ann. Rheum. Dis. 2013, 72, 1747. [Google Scholar] [CrossRef]
- Van Bon, L.; Affandi, A.J.; Broen, J.; Christmann, R.B.; Marijnissen, R.J.; Stawski, L.; Farina, G.A.; Stifano, G.; Mathes, A.L.; Cossu, M.; et al. Proteome-Wide Analysis and CXCL4 as a Biomarker in Systemic Sclerosis. N. Engl. J. Med. 2014, 370, 433–443. [Google Scholar] [CrossRef]
- Vandercappellen, J.; Van Damme, J.; Struyf, S. The Role of the CXC Chemokines Platelet Factor-4 (CXCL4/PF-4) and Its Variant (CXCL4L1/PF-4var) in Inflammation, Angiogenesis and Cancer. Cytokine Growth Factor Rev. 2011, 22, 1–18. [Google Scholar] [CrossRef]
- Moreno, M.; Pauels, K.; Hundelshausen, P.; Berres, M.-L.; Schmitz, P.; Bornemann, J.; Kowalska, M.A.; Gassler, N.; Streetz, K.; Weiskirchen, R.; et al. CXC Chemokine Ligand 4 (CXCL4) Is a Platelet-Derived Mediator of Experimental Liver Fibrosis. Hepatology 2010, 51, 1345–1353. [Google Scholar] [CrossRef]
- Kim, D.; Peck, A.; Santer, D.; Patole, P.; Schwartz, S.M.; Molitor, J.A.; Arnett, F.C.; Elkon, K.B. Induction of Interferon-α by Scleroderma Sera Containing Autoantibodies to Topoisomerase I: Association of Higher Interferon-α Activity with Lung Fibrosis. Arthritis Rheum. 2008, 58, 2163–2173. [Google Scholar] [CrossRef]
- Brkic, Z.; Van Bon, L.; Cossu, M.; Van Helden-Meeuwsen, C.G.; Vonk, M.C.; Knaapen, H.; Van Den Berg, W.; Dalm, V.A.; Van Daele, P.L.; Severino, A.; et al. The Interferon Type I Signature Is Present in Systemic Sclerosis before Overt Fibrosis and Might Contribute to Its Pathogenesis through High BAFF Gene Expression and High Collagen Synthesis. Ann. Rheum. Dis. 2016, 75, 1567–1573. [Google Scholar] [CrossRef]
- Ah Kioon, M.D.; Tripodo, C.; Fernandez, D.; Kirou, K.A.; Spiera, R.F.; Crow, M.K.; Gordon, J.K.; Barrat, F.J. Plasmacytoid Dendritic Cells Promote Systemic Sclerosis with a Key Role for TLR8. Sci. Transl. Med. 2018, 10, eaam8458. [Google Scholar] [CrossRef]
- Lande, R.; Lee, E.Y.; Palazzo, R.; Marinari, B.; Pietraforte, I.; Santos, G.S.; Mattenberger, Y.; Spadaro, F.; Stefanantoni, K.; Iannace, N.; et al. CXCL4 Assembles DNA into Liquid Crystalline Complexes to Amplify TLR9-Mediated Interferon-α Production in Systemic Sclerosis. Nat. Commun. 2019, 10, 1731. [Google Scholar] [CrossRef]
- Skaug, B.; Assassi, S. Biomarkers in Systemic Sclerosis. Curr. Opin. Rheumatol. 2019, 31, 595–602. [Google Scholar] [CrossRef]
- Frasca, L.; Lande, R. Toll-like Receptors in Mediating Pathogenesis in Systemic Sclerosis. Clin. Exp. Immunol. 2020, 201, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Kafaja, S.; Valera, I.; Divekar, A.A.; Saggar, R.; Abtin, F.; Furst, D.E.; Khanna, D.; Singh, R.R. pDCs in Lung and Skin Fibrosis in a Bleomycin-Induced Model and Patients with Systemic Sclerosis. JCI Insight 2018, 3, e98380. [Google Scholar] [CrossRef] [PubMed]
- Pietraforte, I.; Butera, A.; Gaddini, L.; Mennella, A.; Palazzo, R.; Campanile, D.; Stefanantoni, K.; Riccieri, V.; Lande, R.; Frasca, L. CXCL4-RNA Complexes Circulate in Systemic Sclerosis and Amplify Inflammatory/Pro-Fibrotic Responses by Myeloid Dendritic Cells. Int. J. Mol. Sci. 2022, 24, 653. [Google Scholar] [CrossRef]
- Lande, R.; Mennella, A.; Palazzo, R.; Pietraforte, I.; Stefanantoni, K.; Iannace, N.; Butera, A.; Boirivant, M.; Pica, R.; Conrad, C.; et al. Anti-CXCL4 Antibody Reactivity Is Present in Systemic Sclerosis (SSc) and Correlates with the SSc Type I Interferon Signature. Int. J. Mol. Sci. 2020, 21, 5102. [Google Scholar] [CrossRef]
- Lande, R.; Palazzo, R.; Hammel, P.; Pietraforte, I.; Surbeck, I.; Gilliet, M.; Chizzolini, C.; Frasca, L. Generation of Monoclonal Antibodies Specific for Native LL37 and Citrullinated LL37 That Discriminate the Two LL37 Forms in the Skin and Circulation of Cutaneous/Systemic Lupus Erythematosus and Rheumatoid Arthritis Patients. Antibodies 2020, 9, 14. [Google Scholar] [CrossRef] [PubMed]
- Palazzo, R.; Stefanantoni, K.; Cadar, M.; Butera, A.; Riccieri, V.; Lande, R.; Frasca, L. Heparin-Independent and Heparin-Dependent Anti-CXCL4 Antibodies Have a Reciprocal Expression in a Systemic Sclerosis Patients’ Cohort. Antibodies 2022, 11, 77. [Google Scholar] [CrossRef]
- Kowal-Bielecka, O.; Fransen, J.; Avouac, J.; Becker, M.; Kulak, A.; Allanore, Y.; Distler, O.; Clements, P.; Cutolo, M.; Czirjak, L.; et al. Update of EULAR Recommendations for the Treatment of Systemic Sclerosis. Ann. Rheum. Dis. 2017, 76, 1327. [Google Scholar] [CrossRef]
- Kowal-Bielecka, O.; Landewé, R.; Avouac, J.; Chwiesko, S.; Miniati, I.; Czirjak, L.; Clements, P.; Denton, C.; Farge, D.; Fligelstone, K.; et al. EULAR Recommendations for the Treatment of Systemic Sclerosis: A Report from the EULAR Scleroderma Trials and Research Group (EUSTAR). Ann. Rheum. Dis. 2009, 68, 620. [Google Scholar] [CrossRef] [PubMed]
- Clapp, L.H.; Gurung, R. The Mechanistic Basis of Prostacyclin and Its Stable Analogues in Pulmonary Arterial Hypertension: Role of Membrane versus Nuclear Receptors. Prostaglandins Other Lipid Mediat. 2015, 120, 56–71. [Google Scholar] [CrossRef] [PubMed]
- Dorris, S.L.; Peebles, R.S. PGI2 as a Regulator of Inflammatory Diseases. Mediat. Inflamm. 2012, 2012, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Pola, R.; Gaetani, E.; Flex, A.; Aprahamian, T.R.; Bosch-Marcé, M.; Losordo, D.W.; Smith, R.C.; Pola, P. Comparative Analysis of the in Vivo Angiogenic Properties of Stable Prostacyclin Analogs: A Possible Role for Peroxisome Proliferator-Activated Receptors. J. Mol. Cell. Cardiol. 2004, 36, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Biscetti, F.; Straface, G.; Pitocco, D.; Zaccardi, F.; Ghirlanda, G.; Flex, A. Peroxisome Proliferator-Activated Receptors and Angiogenesis. Nutr. Metab. Cardiovasc. Dis. 2009, 19, 751–759. [Google Scholar] [CrossRef]
- Truchetet, M.-E.; Allanore, Y.; Montanari, E.; Chizzolini, C.; Brembilla, N.C. Prostaglandin I2 Analogues Enhance Already Exuberant Th17 Cell Responses in Systemic Sclerosis. Ann. Rheum. Dis. 2012, 71, 2044. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, T.; Jinnin, M.; Yamane, K.; Honda, N.; Kajihara, I.; Makino, T.; Masuguchi, S.; Fukushima, S.; Okamoto, Y.; Hasegawa, M.; et al. Impaired IL-17 Signaling Pathway Contributes to the Increased Collagen Expression in Scleroderma Fibroblasts. J. Immunol. 2012, 188, 3573–3583. [Google Scholar] [CrossRef]
- Truchetet, M.-E.; Mossalayi, M.D.; Boniface, K. IL-17 in the Rheumatologist’s Line of Sight. BioMed Res. Int. 2013, 2013, 295132. [Google Scholar] [CrossRef]
- Brembilla, N.; Montanari, E.; Truchetet, M.-E.; Raschi, E.; Meroni, P.; Chizzolini, C. Th17 Cells Favor Inflammatory Responses While Inhibiting Type I Collagen Deposition by Dermal Fibroblasts: Differential Effects in Healthy and Systemic Sclerosis Fibroblasts. Arthritis Res. Ther. 2013, 15, R151. [Google Scholar] [CrossRef]
- Müller, T.; Dürk, T.; Blumenthal, B.; Herouy, Y.; Sorichter, S.; Grimm, M.; Panther, E.; Cicko, S.; Norgauer, J.; Idzko, M. Iloprost Has Potent Anti-Inflammatory Properties on Human Monocyte-Derived Dendritic Cells. Clin. Exp. Allergy 2010, 40, 1214–1221. [Google Scholar] [CrossRef]
- Kuo, C.-H.; Lin, C.-H.; Yang, S.-N.; Huang, M.-Y.; Chen, H.-L.; Kuo, P.-L.; Hsu, Y.-L.; Huang, S.-K.; Jong, Y.-J.; Wei, W.-J.; et al. Effect of Prostaglandin I2 Analogs on Cytokine Expression in Human Myeloid Dendritic Cells via Epigenetic Regulation. Mol. Med. 2012, 18, 433–444. [Google Scholar] [CrossRef] [PubMed]
- Fabricius, D.; Neubauer, M.; Mandel, B.; Schütz, C.; Viardot, A.; Vollmer, A.; Jahrsdörfer, B.; Debatin, K.-M. Prostaglandin E2 Inhibits IFN-α Secretion and Th1 Costimulation by Human Plasmacytoid Dendritic Cells via E-Prostanoid 2 and E-Prostanoid 4 Receptor Engagement. J. Immunol. 2009, 184, 677–684. [Google Scholar] [CrossRef] [PubMed]
- Ah Kioon, M.D.; Cakan, E.; Garcia-Carmona, Y.; Cunningham-Rundles, C.; Meffre, E.; Barrat, F.J. CXCL4, a Chemokine Upregulated in Systemic Sclerosis Patients, Abrogates TLR9 Signaling and Central Tolerance in B Cells. J. Immunol. 2023, 210, 247.10. [Google Scholar] [CrossRef]
- Zhu, Y.; Liu, Y.; Zhou, W.; Xiang, R.; Jiang, L.; Huang, K.; Xiao, Y.; Guo, Z.; Gao, J. A Prostacyclin Analogue, Iloprost, Protects from Bleomycin-Induced Pulmonary Fibrosis in Mice. Respir. Res. 2010, 11, 34. [Google Scholar] [CrossRef] [PubMed]
- Silva, I.S.; Ferreira, B.H.; Almeida, C.R. Molecular Mechanisms Behind the Role of Plasmacytoid Dendritic Cells in Systemic Sclerosis. Biology 2023, 12, 285. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Ah Kioon, M.D.; Laurent, P.; Chaudhary, V.; Pierides, M.; Yang, C.; Oliver, D.; Ivashkiv, L.B.; Barrat, F.J. Chemokines Form Nanoparticles with DNA and Can Superinduce TLR-Driven Immune Inflammation. J. Exp. Med. 2022, 219, e20212142. [Google Scholar] [CrossRef] [PubMed]
- Laurent, P.; Sisirak, V.; Lazaro, E.; Richez, C.; Duffau, P.; Blanco, P.; Truchetet, M.-E.; Contin-Bordes, C. Innate Immunity in Systemic Sclerosis Fibrosis: Recent Advances. Front. Immunol. 2018, 9, 1702. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Hashimoto, K.; Goleniewska, K.; O’Neal, J.F.O.; Ji, S.; Blackwell, T.S.; FitzGerald, G.A.; Egan, K.M.; Geraci, M.W.; Peebles, R.S., Jr. Prostaglandin I 2 Analogs Inhibit Proinammatory Cytokine Pro duction and T Cell Stimulatory Function of Dendritic Cells. J. Immunol. 2007, 178, 702–710. [Google Scholar] [CrossRef] [PubMed]
- Wong, T.H.; Gau, R.J.; Chen, Y.F.; Shen, H.H.; Lin, C.T.; Chen, S.L.; Suen, J.L. Dendritic cells treated with a prostaglandin I2 analog, iloprost, promote antigen-specific regulatory T cell differentiation in mice. Int. Immunopharmacol. 2020, 79, 106106. [Google Scholar] [CrossRef]
- Maugeri, N.; Capobianco, A.; Rovere-Querini, P.; Ramirez, G.A.; Tombetti, E.; Valle, P.D.; Monno, A.; D’Alberti, V.; Gasparri, A.M.; Franchini, S.; et al. Platelet Microparticles Sustain Autophagy-Associated Activation of Neutrophils in Systemic Sclerosis. Sci. Transl. Med. 2018, 10, eaao3089. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Demographic Data: | Patients N = 30 |
| Sex (M/F) | 1/29 |
| Mean age in years (range) | 58.2 (37–79) |
| Mean disease duration in months (range) | 153.6 (12–468) |
| SSc subtype (lSSc/dSSc) | 18/12 |
| Clinical and Laboratory Features N/% | |
| Raynaud’s phenomenon | 30/100 |
| Digital ulcers | 20/66.6 |
| Telangiectasia | 20/66.6 |
| Calcinosis | 6/20 |
| Lung fibrosis | 14/46.6 |
| Pulmonary arterial hypertension | 0/0 |
| Gastro-intestinal involvement | 22/73.3 |
| Articular involvement | 15/50 |
| Cardiac involvement | 10/33.3 |
| mRSS ≥ 14 | 9/30 |
| ANA+ | 30/100 |
| Anti-Scl70+ | 17/56.6 |
| ACA+ | 6/20 |
| EaSSc/long-lasting SSc | 7/23 |
| Mean EScSGAI T0/T6 | 1.18/1.05 |
| Concomitant Drugs N/% | |
| DMARDs | 15/50 |
| 8/26.6 |
| 4/13.3 |
| 1/3.3 |
| 2/6.6 |
| Vasoactive drugs | 29/96.6 |
| 9/30 |
| 4/13.3 |
| 24/80 |
| 5/16.6 |
| Glucocorticoids | 17/56.6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mennella, A.; Stefanantoni, K.; Palazzo, R.; Ocone, G.; Pietraforte, I.; Truglia, S.; Bisconti, I.; Pisacreta, A.; Riccieri, V.; Lande, R.; et al. Plasma CXCL4–DNA/RNA Complexes and Anti-CXCL4 Antibodies Modulation in an SSc Cohort under Iloprost Treatment. Reports 2024, 7, 66. https://doi.org/10.3390/reports7030066
Mennella A, Stefanantoni K, Palazzo R, Ocone G, Pietraforte I, Truglia S, Bisconti I, Pisacreta A, Riccieri V, Lande R, et al. Plasma CXCL4–DNA/RNA Complexes and Anti-CXCL4 Antibodies Modulation in an SSc Cohort under Iloprost Treatment. Reports. 2024; 7(3):66. https://doi.org/10.3390/reports7030066
Chicago/Turabian StyleMennella, Anna, Katia Stefanantoni, Raffaella Palazzo, Giuseppe Ocone, Immacolata Pietraforte, Simona Truglia, Ilaria Bisconti, Alba Pisacreta, Valeria Riccieri, Roberto Lande, and et al. 2024. "Plasma CXCL4–DNA/RNA Complexes and Anti-CXCL4 Antibodies Modulation in an SSc Cohort under Iloprost Treatment" Reports 7, no. 3: 66. https://doi.org/10.3390/reports7030066
APA StyleMennella, A., Stefanantoni, K., Palazzo, R., Ocone, G., Pietraforte, I., Truglia, S., Bisconti, I., Pisacreta, A., Riccieri, V., Lande, R., & Frasca, L. (2024). Plasma CXCL4–DNA/RNA Complexes and Anti-CXCL4 Antibodies Modulation in an SSc Cohort under Iloprost Treatment. Reports, 7(3), 66. https://doi.org/10.3390/reports7030066