Soil Properties Related to the Occurrence of Rock Phosphate-Solubilizing Bacteria in the Rhizosphere Soil of Faba Bean (Vicia faba L.) in Morocco


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Soil Sampling
2.2. Physicochemical Soil Characteristics
2.3. Bacterial Counting
2.4. Statistical Analysis
3. Results
3.1. Physicochemical Soil Properties
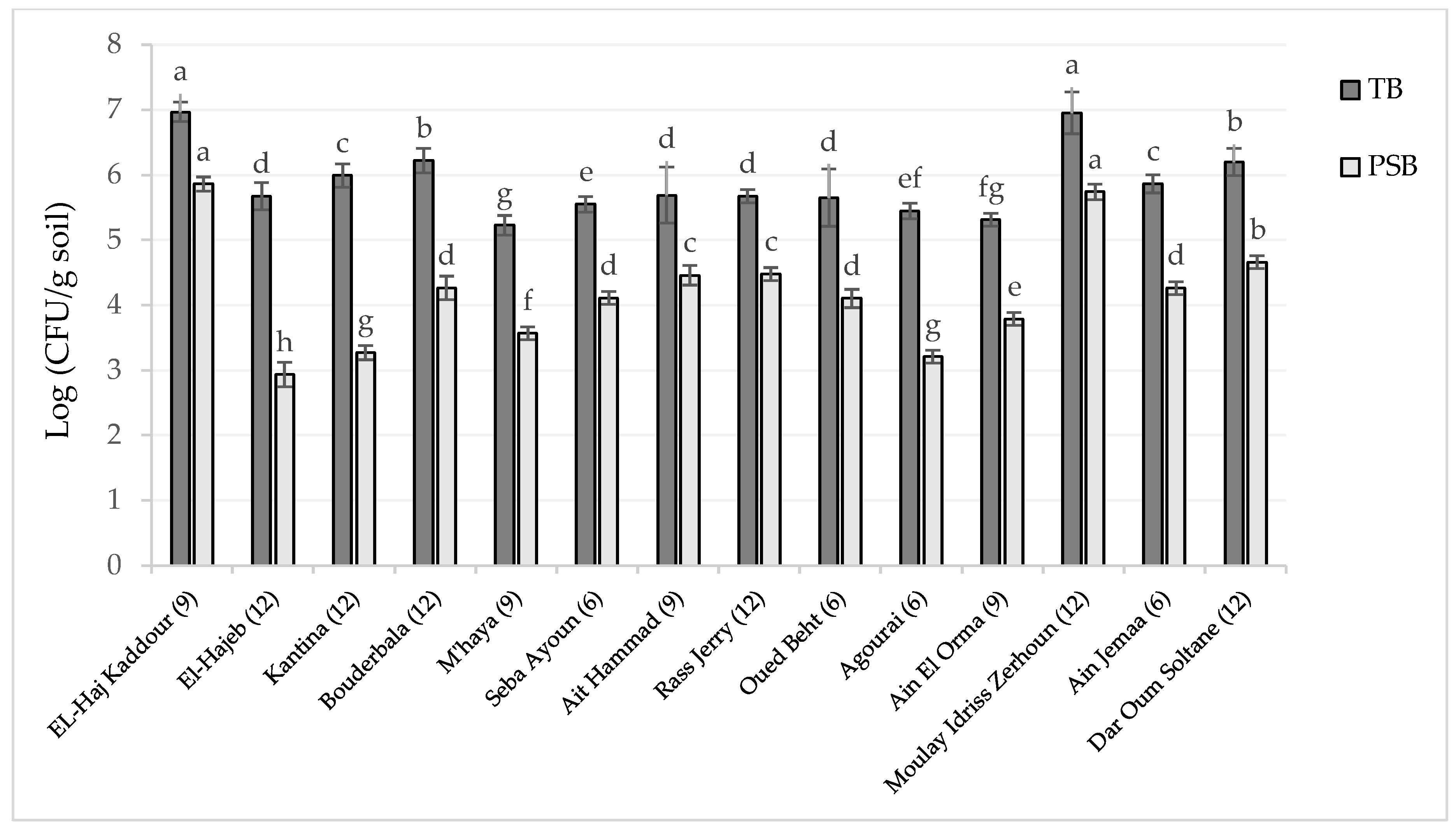
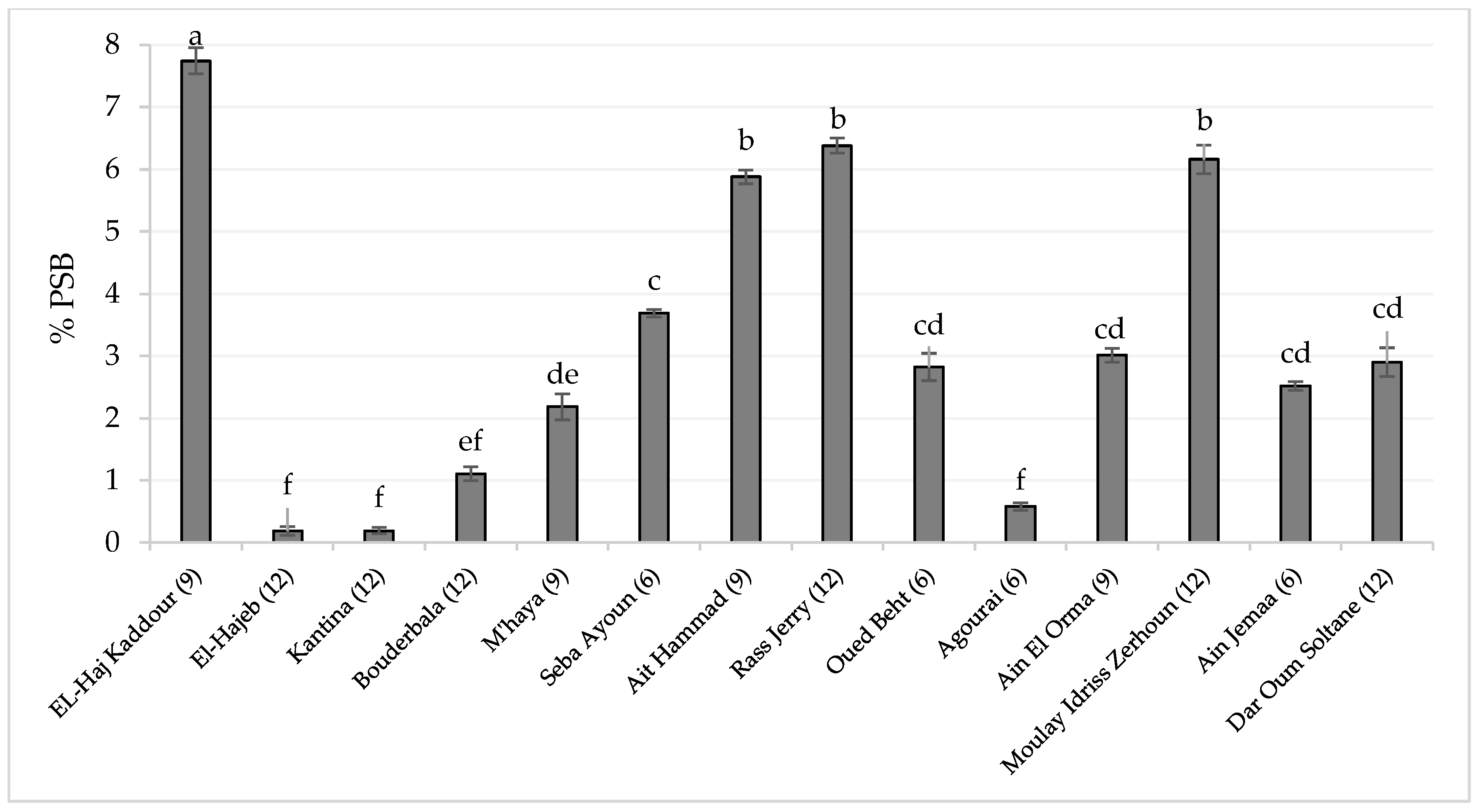
3.2. Total Soil Bacteria (TB) and PSB Counting
3.3. The Relationship between Soil Properties and PSB Population
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Richardson, A.E. Prospects for using soil microorganisms to improve the acquisition of phosphorus by plants. Aust. J. Plant Physiol. 2001, 28, 897–906. [Google Scholar] [CrossRef]
- Zaidi, A.; Khan, M.S.; Ahemad, M.; Oves, M. Plant growth promotion by phosphate solubilizing bacteria. Acta Microbiol. Immunol. Hung. 2009, 56, 263–284. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.S.; Zaidi, A.; Ahemad, M.; Oves, M.; Wani, P.A. Plant growth promotion by phosphate solubilizing fungi—Current perspective. Arch. Agron. Soil Sci. 2010, 56, 73–98. [Google Scholar] [CrossRef]
- Hamdali, H.; Smirnov, A.; Esnault, C.; Ouhdouch, Y.; Virolle, M.J. Physiological studies and comparative analysis of rock phosphate solubilization abilities of actinomycetales originating from Moroccan phosphate mines and of Streptomyces lividans. Appl. Soil Ecol. 2010, 44, 24–31. [Google Scholar] [CrossRef]
- Hiltner, L. Uber neuere erfahrungen und probleme auf dem gebiete der bodenbakteriologie unter besonderer berucksichtigung der grundungung und brache. Arb. Dtsc. Landwirtsch. Ges. 1904, 98, 59–78. [Google Scholar]
- Kamaludeen, S.P.B.; Ramasamy, K. Rhizoremediation of metals: Harnessing microbial communities. Indian J. Microbiol. 2008, 48, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, X.; Chen, J.; Shim, H.; Bai, Z. New advances in plant growth-promoting rhizobacteria for bioremediation. Environ. Int. 2007, 33, 406–413. [Google Scholar] [CrossRef] [PubMed]
- McNear, D., Jr. The rhizosphere-roots, soil and everything in between. Nat. Educ. Knowl. 2013, 4, 1. [Google Scholar]
- Zaidi, A.; Khan, M.S.; Ahmad, E. Microphos: Principles, Production and Application Strategies. In Phosphate Solubilizing Microorganisms: Principles and Application of Microphos Technology; Khan, M.S., Zaidi, A., Musarrat, J., Eds.; Springer International Publishing: Cham, Switzerland, 2014; pp. 1–30, ISBN 978-3-319-08215-8, 978-3-319-08216-5. [Google Scholar]
- Vazquez, P.; Holguin, G.; Puente, M.; Cortes, A.E.; Bashan, Y. Phosphate solubilizing microorganisms associated with the rhizosphere of mangroves in a semi-arid coastal lagoon. Biol. Fertil. Soils 2000, 30, 460–468. [Google Scholar] [CrossRef]
- Hinsinger, P. Bioavailability of soil inorganic P in the rhizosphere as affected by root induced chemical changes: A review. Plant Soil 2001, 237, 173–195. [Google Scholar] [CrossRef]
- Tunesi, S.; Poggi, V.; Gessa, C. Phosphate adsorption and precipitation in calcareous soils: The role of calcium ions in solution and carbonate minerals. Nut. Cycl. Agroecosyst. 1999, 53, 219–227. [Google Scholar] [CrossRef]
- Tisdale, S.L.; Nelson, W.L.; Beaton, J.D.; Havlin, J.L. Soil Fertility and Fertilizers, 5th ed.; Macmillan Publishing Company: New York, NY, USA, 1993; p. 634. ISBN 0-02-420835-3. [Google Scholar]
- Baig, K.S.; Arshad, M.; Shaharoona, B.; Khalid, A.; Ahmed, I. Comparative effectiveness of Bacillus spp. possessing either dual or single growth-promoting traits for improving phosphorus uptake, growth and yield of wheat (Triticum aestivum L.). Ann. Microbiol. 2012, 62, 1109–1119. [Google Scholar] [CrossRef]
- Vassilev, N.; Vassileva, M. Biotechnological solubilization of rock phosphate on media containing agro-industrial wastes. Appl. Microbiol. Biotechnol. 2003, 61, 435–440. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Yuan, L.; Zhang, J.; Li, H.; Bai, Z.; Chen, X.; Zhang, W.; Zhang, F. Phosphorus dynamics: From soil to plant. Plant Physiol. 2011, 156, 997–1005. [Google Scholar] [CrossRef] [PubMed]
- Katiyar, V.; Goel, R. Solubilization of inorganic phosphate and plant growth promotion by cold tolerant mutants of Pseudomonas fluorescens. Microbiol. Res. 2003, 158, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Chabot, R.; Antoun, H.; Cescas, M.P. Stimulation de la croissance du maïs et de la laitue romaine par des microorganismes dissolvant le phosphore inorganique. Can. J. Micribiol. 1993, 39, 941–947. [Google Scholar] [CrossRef]
- Ndung’u-Magiroi, K.W.; Herrmann, L.; Okalebo, J.R.; Othieno, C.O.; Pypers, P.; Lesueur, D. Occurrence and genetic diversity of phosphate-solubilizing bacteria in soils of differing chemical characteristics in Kenya. Ann. Microbiol. 2012, 62, 897–904. [Google Scholar] [CrossRef]
- Azziz, G.; Bajsa, N.; Haghjou, T.; Taulé, C.; Valverde, Á.; Igual, J.M.; Arias, A. Abundance, diversity and prospecting of culturable phosphate solubilizing bacteria on soils under crop–pasture rotations in a no-tillage regime in Uruguay. Appl. Soil Ecol. 2012, 61, 320–326. [Google Scholar] [CrossRef] [Green Version]
- Fernández, L.A.; Agaras, B.; Wall, L.G.; Valverde, C. Abundance and ribotypes of phosphate-solubilizing bacteria in Argentinean agricultural soils under no-till management. Ann. Microbiol. 2015, 65, 1667–1678. [Google Scholar] [CrossRef]
- Olsen, S.R.; Cole, C.V.; Watanabe, F.S.; Dean, L.A. Estimation of available phosphorus in soils by extraction with sodium bicarbonate. In USDA Circular; Government Publication; Printing Office: Washington, DC, USA, 1954; pp. 1–19. [Google Scholar]
- Murphy, J.; Riley, J.P. A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Trivedy, R.K.; Goel, P.K.; Trisal, C.L. Practical Methods in Ecology and Environmental Media; Series in Methodology-2; Enviro Media Publication: Karad, India, 1998. [Google Scholar]
- Walkley, A.; Black, A. An examination of the Degtjareff method for determining soil organic matter and proposed modification of the chromic acid titration method. Soil Sci. 1934, 37, 29–37. [Google Scholar] [CrossRef]
- Metson, A.J. Methods of Chemical Analysis for Soil Survey Samples; Bulletin 12; Department of Scientific and Industrial Research: Wellington, New Zealand, 1956; p. 207. [Google Scholar]
- Araújo, W.L.; Lima, A.O.S.; Azevedo, J.L.; Marcon, J.; Sobral, J.K.; Lakava, P.L. Manual Isolation of Endophytic Microorganisms; Department of Genetics—College of Agriculture “Luiz de Queiroz”—Piracicaba ESALQ—USP: São Paulo, Brazil, 2002. (In Portuguese) [Google Scholar]
- Atlas, R.M. Handbook of Microbiological Media, 4th ed.; Taylor & Francis Group: Boca Raton, FL, USA, 2010; p. 2040. [Google Scholar]
- Nautiyal, C.S. An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS Microbiol. Lett. 1999, 170, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Hamdali, H.; Hafidi, M.; Virolle, M.J.; Ouhdouch, Y. Rock phosphate solubilizing Actinomycetes: Screening for plant growth promoting activities. World J. Microbiol. Biotechnol. 2008, 24, 2565–2575. [Google Scholar] [CrossRef]
- Mehlich, A. Mehlich-3 soil test extractant: A modification of Mehlich 2 extractant. Commun. Soil Sci. Plant Anal. 1984, 15, 1409–1416. [Google Scholar] [CrossRef]
- Tóth, G.; Guicharnaud, R.A.; Tóth, B.; Hermann, T. Phosphorus levels in croplands of the European Union with implications for P fertilizer use. Eur. J. Agron. 2014, 55, 42–52. [Google Scholar] [CrossRef]
- Tóth, G.; Jones, A.; Montanarella, L. LUCAS Topsoil Survey. Methodology, Data and Results. JRC Technical Reports; EUR26102–Scientific and Technical Research Series; Publications Office of the European Union: Luxembourg, 2013; p. 141. [Google Scholar] [CrossRef]
- Giroux, M.; Cantin, J.; Rivest, M.; Tremblay, G. L’évolution des teneurs en phosphore dans les sols selon leur fertilité, leur richesse en phosphore et les types de sol. In Colloque sur le Phosphore: Une Gestion Éclairée; Ordre des agronomes du Québec: Drummondville, QC, Canada, 2002. [Google Scholar]
- Kuhad, R.C.; Singh, S.; Lata, S.A. Phosphate solubilizing microorganisms. In Bioaugmentation, Biostimulation and Biocontrol; Singh, A., Parmar, N., Kuhad, R.C., Eds.; Soil Biology Series; Springer: Heidelberg, Germany, 2011; Volume 28, pp. 65–84. [Google Scholar] [CrossRef]
- Mutai, C.; Njuguna, J.; Ghimire, S. Brachiaria Grasses (Brachiaria spp.) harbor a diverse bacterial community with multiple attributes beneficial to plant growth and development. MicrobiologyOpen 2017, 6, e497. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Xu, G.; Ma, P.; Lin, Y.; Yang, X.; Cao, C. Isolation and Characterization of a Phosphorus-Solubilizing Bacterium from Rhizosphere Soils and Its Colonization of Chinese Cabbage (Brassica campestris ssp. chinensis). Front. Microbiol. 2017, 8, 1270. [Google Scholar] [CrossRef] [PubMed]
- Buscot, F. What are soils? In Microorganisms in Soils: Roles in Genesis and Functions; Buscot, F., Varma, S., Eds.; Springer: Heidelberg, Germany, 2005; Volume 3, pp. 3–18. [Google Scholar] [CrossRef]
- Nahas, E. Phosphate solubilizing microorganisms: Effects of carbon, nitrogen and phosphorus. In First International Meeting on Microbial Phosphate Solubilization: Developments in Plant and Soil Sciences; Velazquez, E., Rodriguez-Barrueco, C., Eds.; Springer SBM: Dordrecht, The Netherlands, 2007; Volume 102, pp. 111–115. [Google Scholar] [CrossRef]
- Brady, N.C.; Weil, R.R. The Nature and Properties of Soils, 13th ed.; Prentice Hall: Upper Saddle River, NJ, USA, 2002; p. 960, ISBN 0130167630, 9780130167637. [Google Scholar]
- Rowell, D.L. Soil Science: Methods and Application; Longman Scientific & Technical Group: New York, NY, USA; London, UK, 1994; p. 350, ISBN 0582087848, 9780582087842. [Google Scholar]
- Mbonigaba Muhinda, J.J.; Nzeyimana, I.; Bucagu, C.; Culot, M. Caractérisation physique, chimique et microbiologique de trois sols acides tropicaux du Rwanda sous jachères naturelles et contraintes à leur productivité. Biotechnol. Agron. Soc. Environ. 2009, 13, 545–558. [Google Scholar]
- He, Z.L.; Yang, X.E.; Baligar, B.C.; Calvert, D.V. Microbiological and biochemical indexing systems for assessing quality of acid soils. Adv. Agron. 2003, 78, 89–138. [Google Scholar]
- Chu, H.; Fujii, T.; Morimoto, S.; Lin, X.; Yagi, K.; Hu, J.; Zhang, J. Soil microbial biomass, dehydrogenase activity, bacterial community structure in response to long-term fertilizer management. Soil Biol. Biochem. 2007, 39, 2971–2976. [Google Scholar] [CrossRef]
- Kennedy, A.C. Bacterial diversity in agroecosystems. Agric. Ecosyst. Environ. 1999, 74, 65–76. [Google Scholar] [CrossRef]
- Hu, J.; Lin, X.; Wang, J.; Chu, H.; Yin, R.; Zhang, J. Population size and specific potential of P-mineralizing and—Solubilizing bacteria under long-term P-deficiency fertilization in a sandy loam soil. Pedobiologia 2009, 53, 49–58. [Google Scholar] [CrossRef]
- Kundu, B.; Nehra, K.; Yadav, R.; Tomar, M. Biodiversity of phosphate solubilizing bacteria in rhizosphere of chickpea, mustard and wheat grown in different regions of Haryana, Indian. J. Microbiol. 2009, 49, 120–127. [Google Scholar] [CrossRef]
- Peix, A.; Rivas-Boyero, A.A.; Mateos, P.F.; Rodríguez-Barrueco, C.; Martínez-Molina, E.; Velazquez, E. Growth promotion of chickpea and barley by a phosphate solubilizing strain of Mesorhizobium mediterraneum under growth chamber conditions. Soil Biol. Biochem. 2001, 33, 103–110. [Google Scholar] [CrossRef]
- Vikram, A.; Alagawadi, A.R.; Hamzehzarghani, H.; Krishnaraj, P.U. Factors related to the occurrence of phosphate solubilizing bacteria and their isolation in Vertisols. Int. J. Agric. Res. 2007, 2, 571–580. [Google Scholar] [CrossRef]
- Costa, R.; Götz, M.; Mrotzek, N.; Lottmann, J.; Berg, G.; Smalla, K. Effects of site and plant species on rhizosphere community structure as revealed by molecular analysis of microbial guilds. FEMS Microbiol. Ecol. 2006, 56, 236–249. [Google Scholar] [CrossRef] [PubMed]
- Belimov, A.A.; Puhalsky, I.V.; Safronova, V.I.; Shaposhnikov, A.I.; Vishnyakova, M.A.; Semenova, E.V.; Nadezda, Y.Z.; Natalya, M.M.; Walter, W.; Igor, A.T. Role of plant genotype and soil conditions in symbiotic plant-microbe interactions for adaptation of plants to cadmium-polluted soils. Water Air Soil Pollut. 2015, 226, 1–15. [Google Scholar] [CrossRef]
- Schreiter, S.; Sandmann, M.; Smalla, K.; Grosch, R. Soil type dependent rhizosphere competence and biocontrol of two bacterial inoculant strains and their effects on the rhizosphere microbial community of field-grown lettuce. PLoS ONE 2014, 9, e103726. [Google Scholar] [CrossRef] [PubMed]
- Yahya, A.I.; Al-Azawi, S.K. Occurrence of phosphate-solubilizing bacteria in some Iraqi soils. Plant Soil 1989, 117, 135–141. [Google Scholar] [CrossRef]
- Kucey, R.M.N. Phosphate solubilizing bacteria and fungi in various cultivated and virgin Alberta soils. Can. J. Soil Sci. 1983, 63, 671–678. [Google Scholar] [CrossRef]
- Moat, A.G.; Foster, J.W. Microbial Physiology, 2nd ed.; Wiley-Interscience: New York, NY, USA, 1988; p. 597. [Google Scholar]
- John, L.; Herms, D.; Stinner, B.; Hostink, H. Mulch Effect on Soil Microbial Activity, Nutrient Cycling, and Plant Growth in Ornamental Landscape; Ornamental Plant Annual Report and Research Reviews; The Ohio State University-Columbus: Columbus, OH, USA, 2001. [Google Scholar]
- Bashan, Y.; Puente, M.E.; Rodriquea, M.N.; Toledo, G.; Holguin, G.; Ferrera-Cerrato, R.; Pedrin, S. Survival of Azorhizobium brasilense in the bulk soil and rhizosphere of 23 soil types. Appl. Environ. Microbiol. 1995, 61, 1938–1945. [Google Scholar] [PubMed]



{kind=link}
{kind=link}
| No. | Sampling Site | Location | Soil pH | Organic Matter (g·kg−1) | Available P (mg·kg−1) | Total N (g·kg−1) | CEC (cmol (+)/kg) |
|---|---|---|---|---|---|---|---|
| 1 | EL-Haj Kaddour (9) | 33°49′18 N; 005°25′31 W | 6.15 ± 0.08 de | 45.49 ± 0.91 a | 40.59 ± 0.62 a | 1.71 ± 0.05 a | 18.26 ± 1.21 def |
| 2 | El-Hajeb (12) | 33°39′45 N; 005°21′21 W | 7.22 ± 0.03 abc | 11.70 ± 1.06 ef | 32.84 ± 1.18 c | 0.11 ± 0.01 f | 29.33 ± 0.91 cde |
| 3 | Kantina (12) | 33°41′23 N; 005°31′37 W | 7.94 ± 0.05 b | 16.48 ± 0.94 de | 40.47 ± 0.65 a | 0.12 ± 0.01 f | 29.45 ± 0.62 cde |
| 4 | Bouderbala (12) | 33°49′55 N; 005°16′09 W | 8.07 ± 0.24 b | 31.67 ± 0.88 b | 40.37 ± 0.65 a | 0.93 ± 0.05 cd | 11.57 ± 0.94 f |
| 5 | M’haya (9) | 33°57′44 N; 005°13′42 W | 8.12 ± 0.04 a | 9.73 ± 0.37 f | 14.05 ± 0.11 f | 0.93 ± 0.04 cd | 13.72 ± 0.93 ef |
| 6 | Seba Ayoun (6) | 33°54′27 N; 005°26′35 W | 7.11 ± 0.09 abcd | 25.43 ± 1.18 c | 41.58 ± 0.47 a | 0.78 ± 0.03 de | 8.87 ± 0.11 f |
| 7 | Ait Hammad (9) | 33°52′46 N; 005°09′14 W | 6.03 ± 0.18 e | 22.43 ± 0.91 c | 27.40 ± 0.58 de | 0.72 ± 0.04 e | 10.31 ± 0.78 f |
| 8 | Rass Jerry (12) | 33°46′06 N; 005°45′17 W | 7.45 ± 0.02 abc | 35.85 ± 1.25 b | 30.48 ± 0.57 cd | 1.18 ± 0.02 b | 12.71 ± 0.16 f |
| 9 | Oued Beht (6) | 33°52′15 N; 005°53′46 W | 7.62 ± 0.02 b | 24.52 ± 0.77 c | 39.96 ± 1.01 a | 0.78 ± 0.03 de | 42.58 ± 0.95 abc |
| 10 | Agourai (6) | 33°37′32 N; 005°38′41 W | 7.58 ± 0.04 abc | 13.88 ± 1.29 ef | 29.64 ± 0.93 d | 0.16 ± 0.03 f | 44.51 ± 0.50 abc |
| 11 | Ain El Orma (9) | 33°54′02 N; 005°46′11 W | 7.06 ± 0.06 bcde | 20.58 ± 0.49 cd | 36.62 ± 0.51 b | 1.17 ± 0.04 b | 52.62 ± 0.65 a |
| 12 | Moulay Idriss Zerhoun (12) | 34°01′48 N; 005°34′33 W | 7.97 ± 0.06 b | 43.39 ± 0.54 a | 25.69 ± 0.36 e | 1.68 ± 0.02 a | 30.52 ± 0.66 bcd |
| 13 | Ain Jemaa (6) | 33°59′03 N; 005°41′39 W | 7.53 ± 0.06 abc | 31.06 ± 0.15 b | 29.90 ± 0.25 cd | 1.09 ± 0.01 bc | 46.50 ± 1.01 ab |
| 14 | Dar Oum Soltane (12) | 33°53′56 N; 005°38′50 W | 6.03 ± 0.18 cde | 22.43 ± 0.84 c | 27.06 ± 1.04 de | 0.76 ± 0.01 de | 11. 31 ± 0.74 def |
| Sum of Squares | ddl | Average Square | F | Signification | ||
|---|---|---|---|---|---|---|
| TB | Intergroup | 11.348 | 13 | 0.873 | 380.847 | 0.000 |
| Intragroup | 0.064 | 28 | 0.002 | |||
| Total | 11.412 | 41 | ||||
| PSB | Intergroup | 28.570 | 13 | 2.198 | 617.152 | 0.000 |
| Intragroup | 0.100 | 28 | 0.004 | |||
| Total | 28.669 | 41 | ||||
| %PSB | Intergroup | 234.418 | 13 | 18.032 | 93.522 | 0.000 |
| Intragroup | 5.399 | 28 | 0.193 | |||
| Total | 239.817 | 41 | ||||
| PSB | TB | pH | OM | P | N | CEC | |
|---|---|---|---|---|---|---|---|
| PSB | 1 | ||||||
| TB | 0.800 ** | 1 | |||||
| pH | −0.346 | −0.170 | 1 | ||||
| OM | 0.902 ** | 0.777 ** | −0.148 | 1 | |||
| P | 0.031 | 0.200 | −0.080 | 0.268 | 1 | ||
| N | 0.866 ** | 0.551 ** | −0.119 | 0.822 ** | −0.068 | 1 | |
| CEC | −0.283 | −0.195 | 0.274 | −0.149 | 0.129 | −0.101 | 1 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rfaki, A.; Zennouhi, O.; Nassiri, L.; Ibijbijen, J. Soil Properties Related to the Occurrence of Rock Phosphate-Solubilizing Bacteria in the Rhizosphere Soil of Faba Bean (Vicia faba L.) in Morocco. Soil Syst. 2018, 2, 31. https://doi.org/10.3390/soilsystems2020031
Rfaki A, Zennouhi O, Nassiri L, Ibijbijen J. Soil Properties Related to the Occurrence of Rock Phosphate-Solubilizing Bacteria in the Rhizosphere Soil of Faba Bean (Vicia faba L.) in Morocco. Soil Systems. 2018; 2(2):31. https://doi.org/10.3390/soilsystems2020031
Chicago/Turabian StyleRfaki, Abderrazak, Omar Zennouhi, Laila Nassiri, and Jamal Ibijbijen. 2018. "Soil Properties Related to the Occurrence of Rock Phosphate-Solubilizing Bacteria in the Rhizosphere Soil of Faba Bean (Vicia faba L.) in Morocco" Soil Systems 2, no. 2: 31. https://doi.org/10.3390/soilsystems2020031
APA StyleRfaki, A., Zennouhi, O., Nassiri, L., & Ibijbijen, J. (2018). Soil Properties Related to the Occurrence of Rock Phosphate-Solubilizing Bacteria in the Rhizosphere Soil of Faba Bean (Vicia faba L.) in Morocco. Soil Systems, 2(2), 31. https://doi.org/10.3390/soilsystems2020031