
Long-Term Effects of Forest Plantation Species on Chemical Soil Properties in Southern Rwanda


Abstract
:1. Introduction
2. Materials and Methods
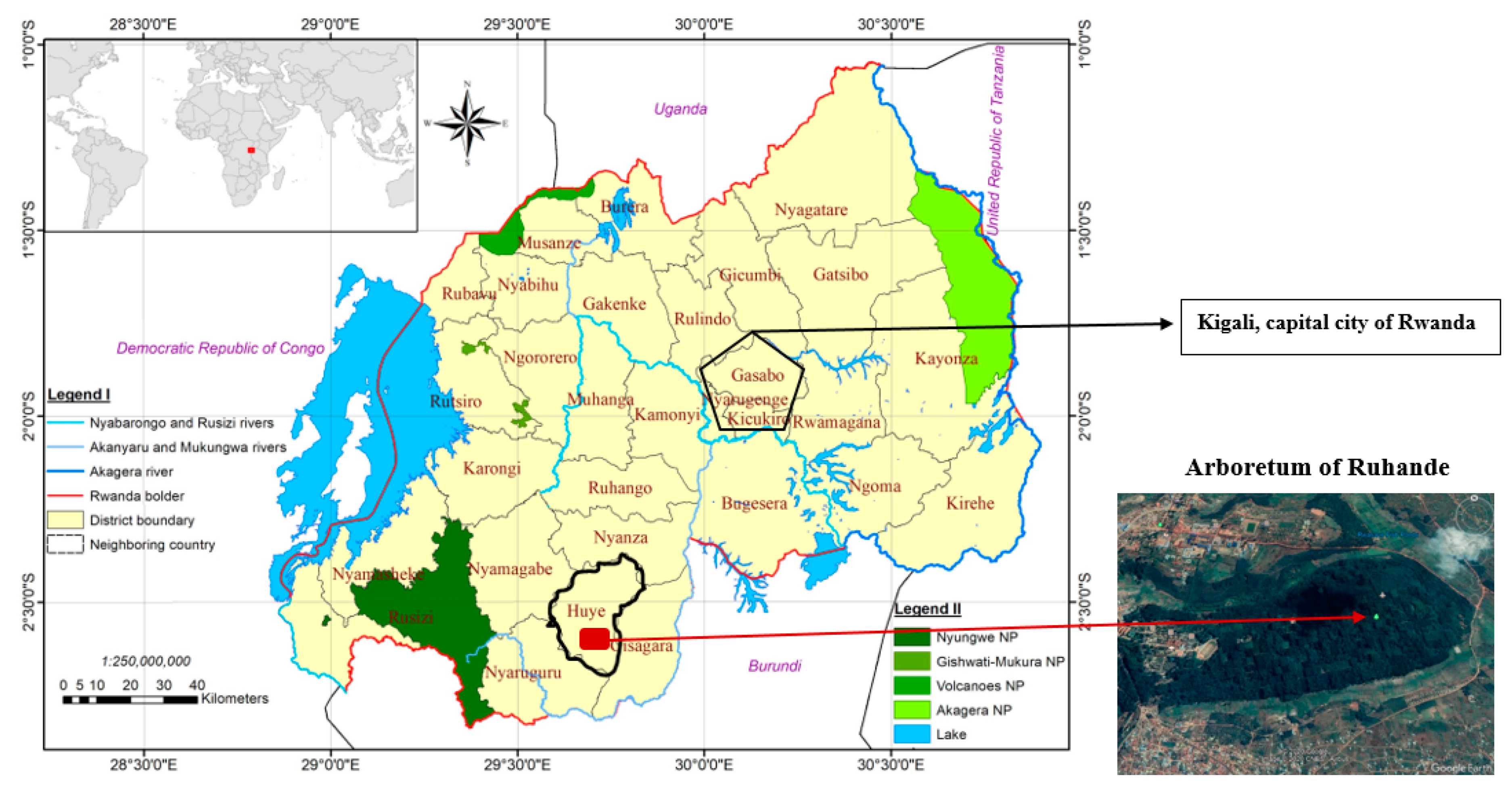
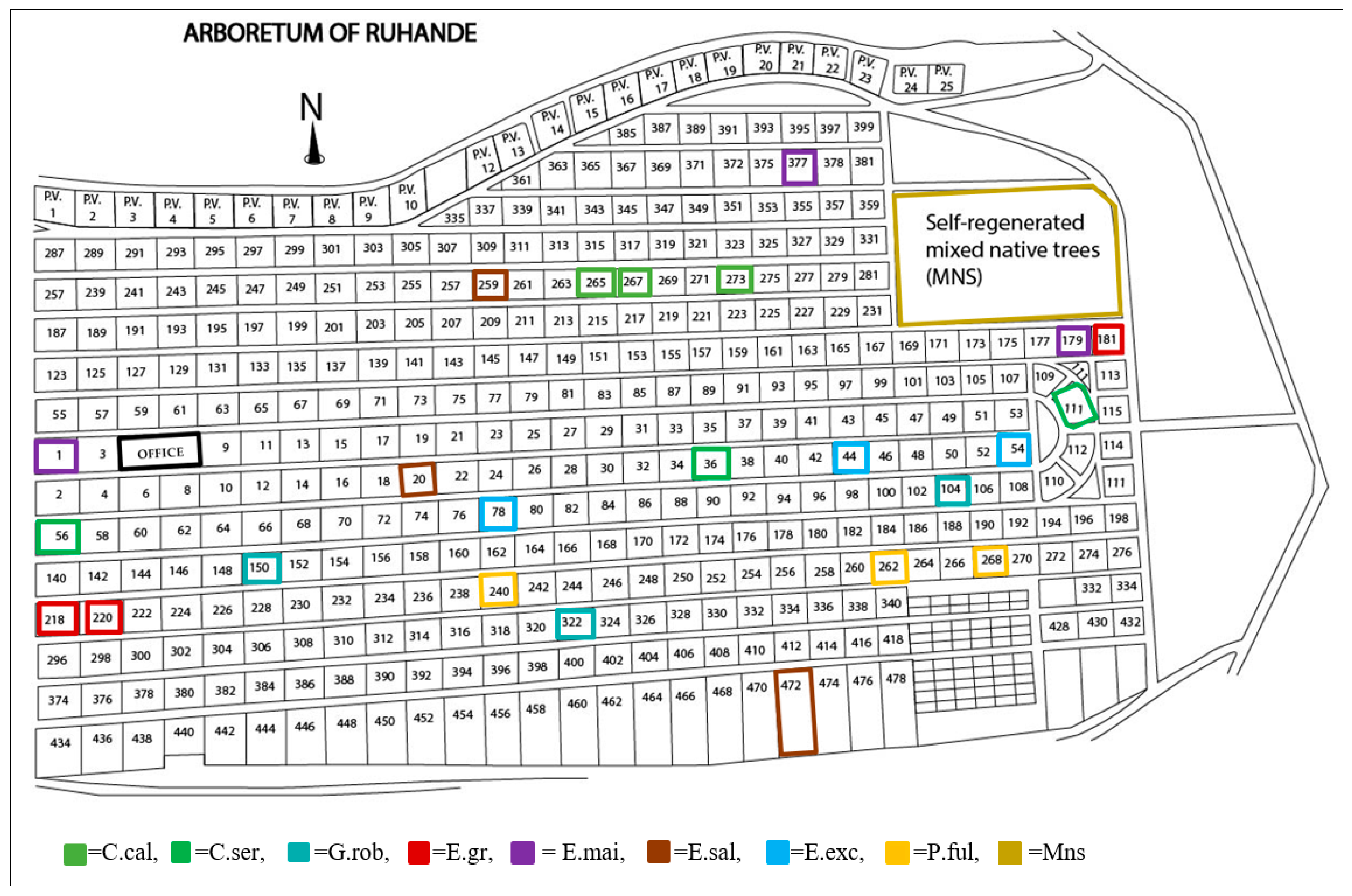
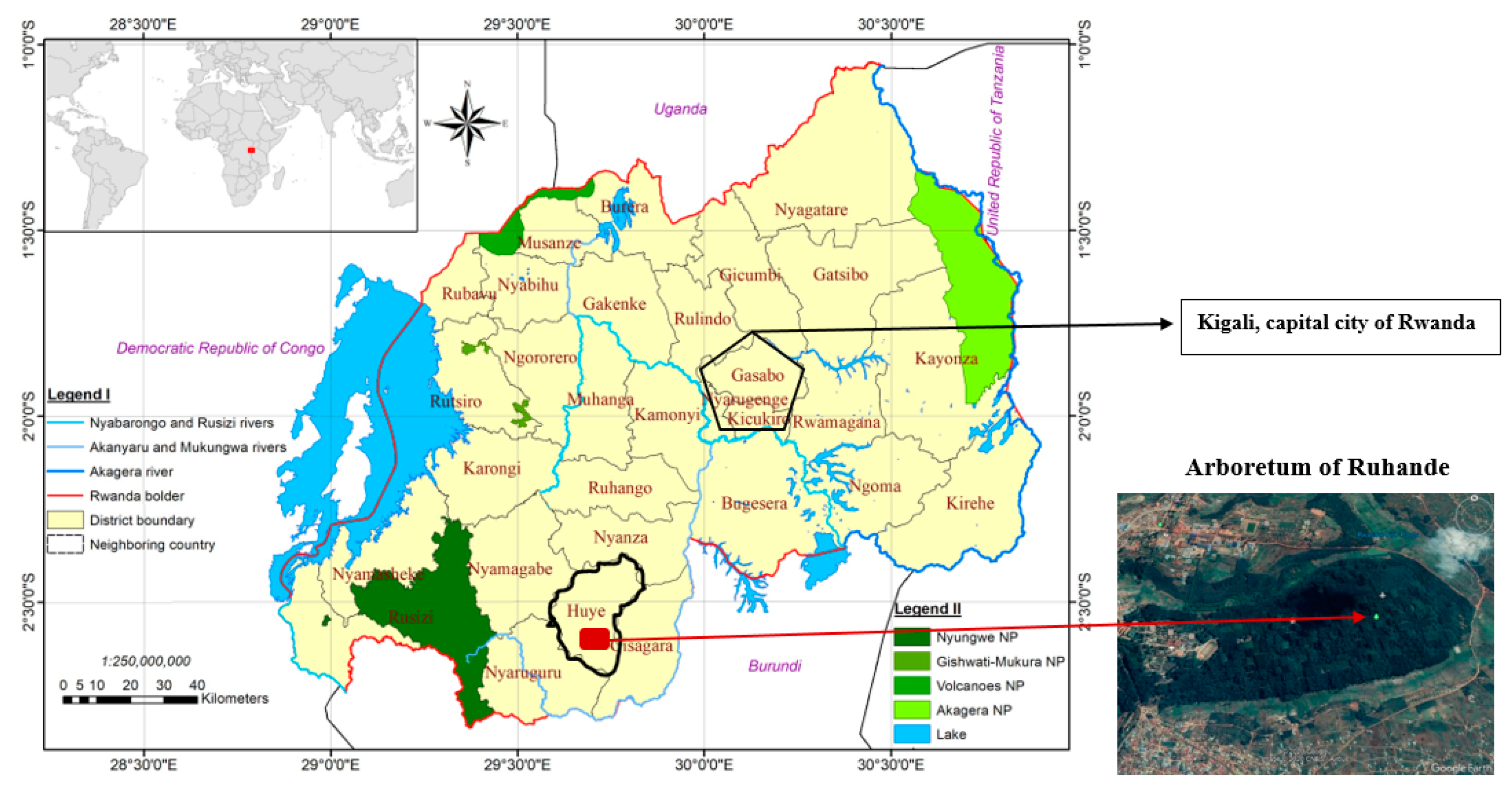
2.1. Study Site
2.2. Soil Sampling and Chemical Analyses
2.3. Statistical Analysis
3. Results
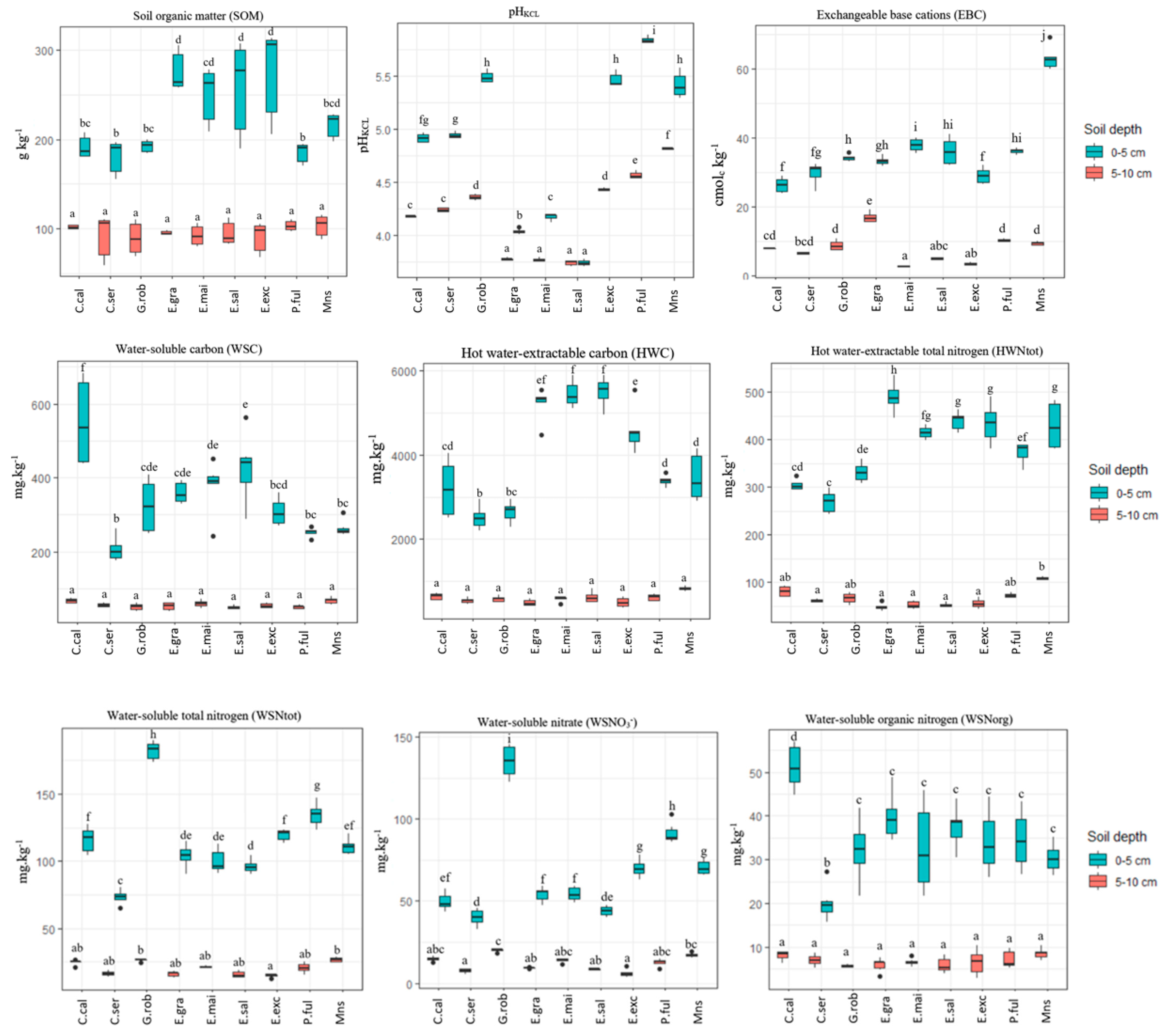
3.1. Chemical Soil Properties in Two Topsoil Layers
3.2. Effects of Tree Species on Water-Extractable C and N and Other Soil Properties
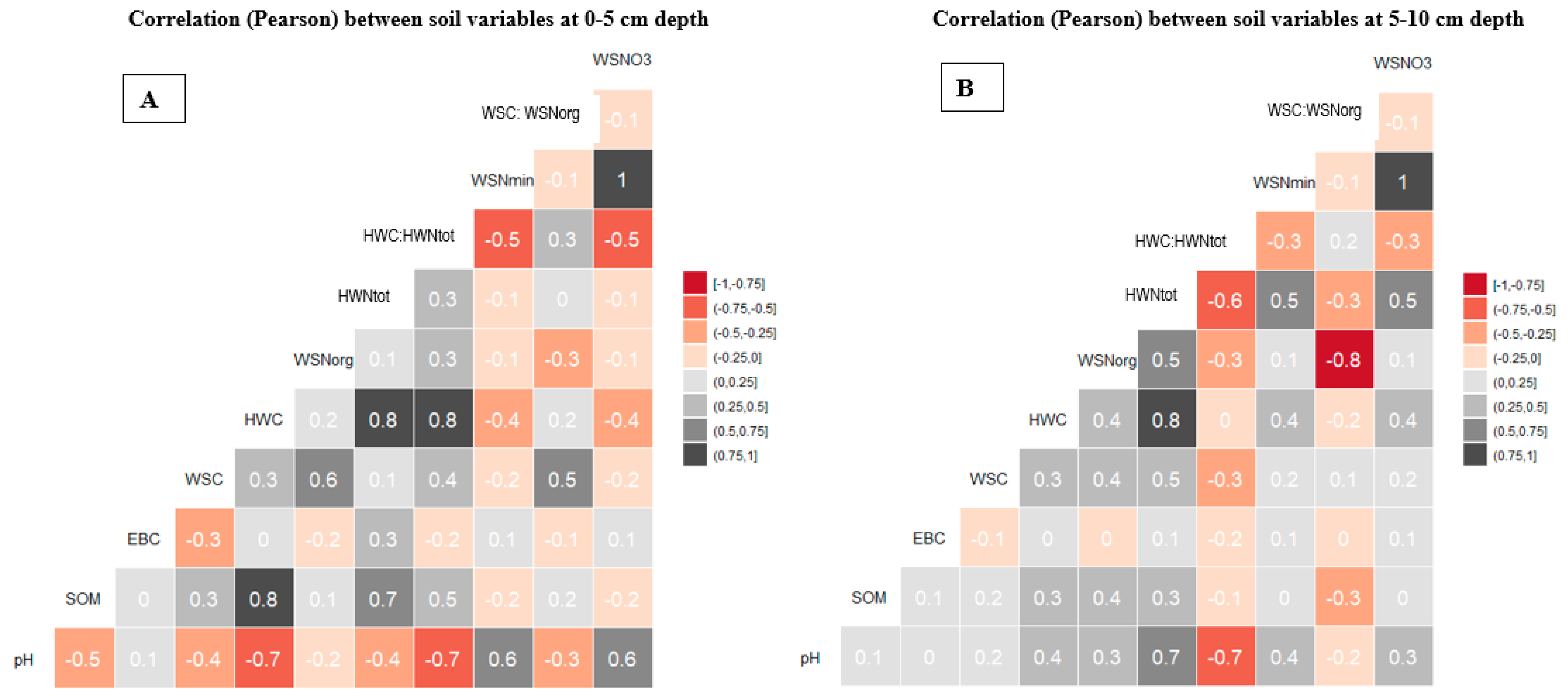
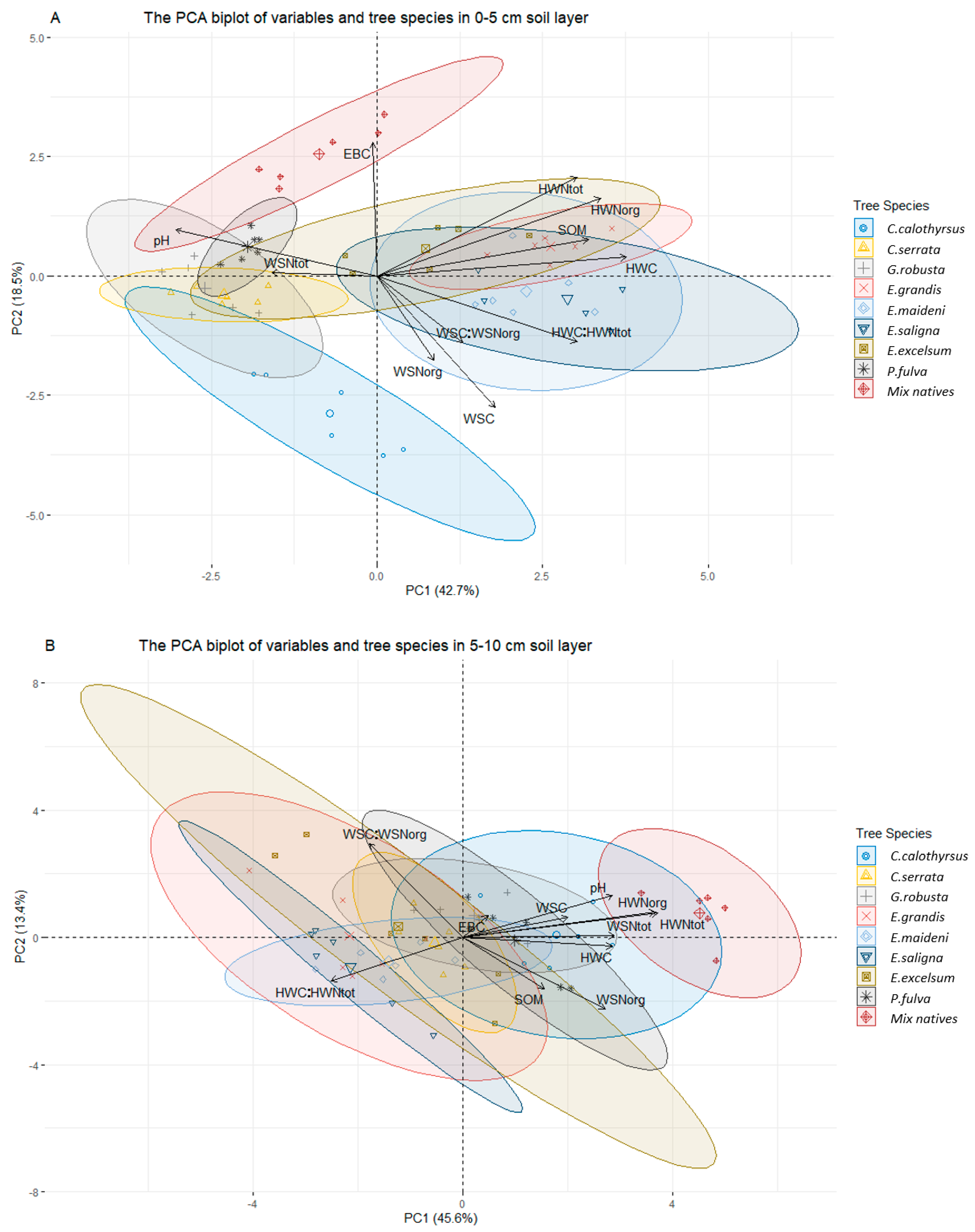
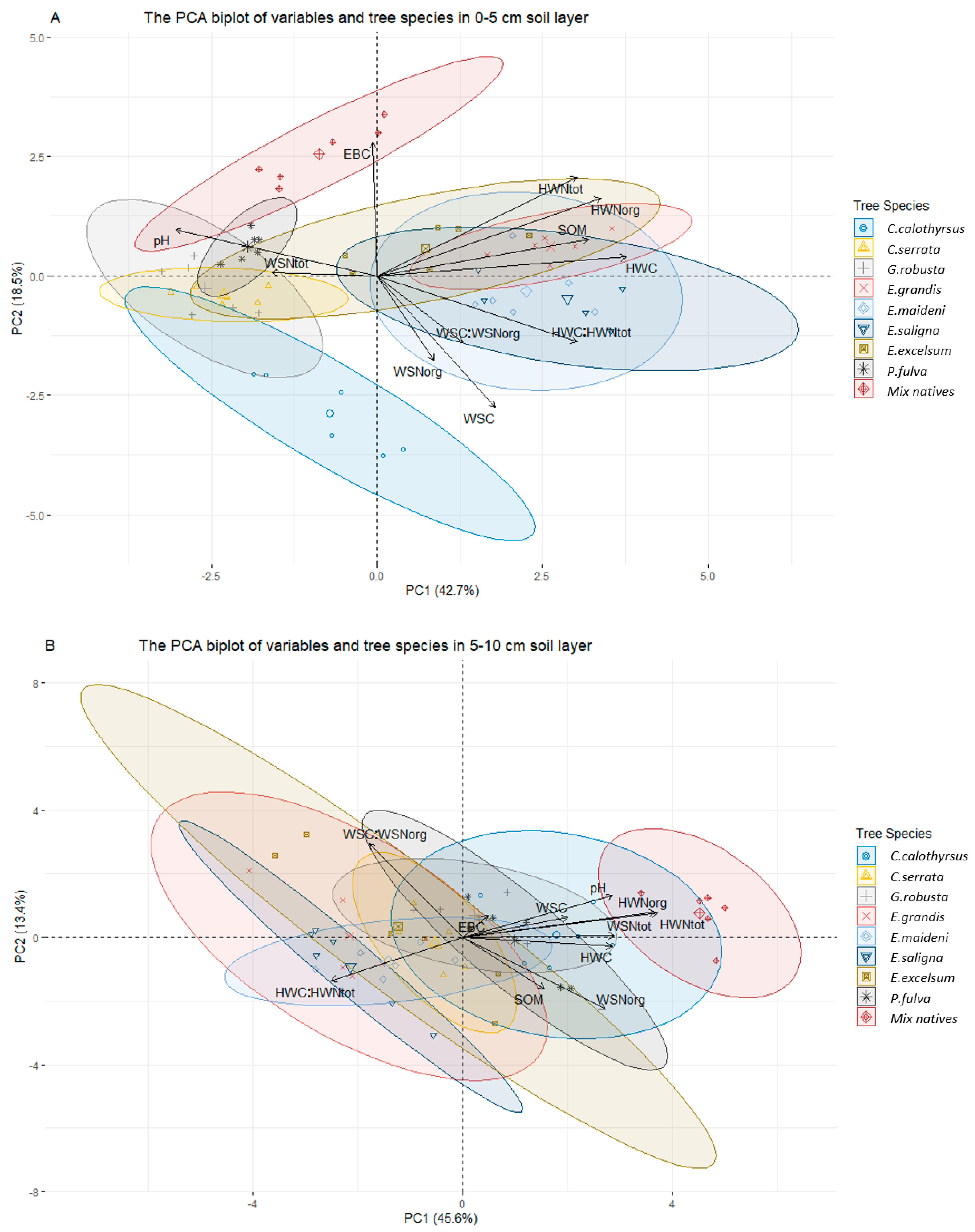
3.3. Relationships between Water-Extractable Elements (C, N) and Other Soil Properties
4. Discussion
4.1. Importance of the Thin Upper Soil Layer (0–5 cm Depth)
4.2. Effects of Tree Species on Chemical Soil Properties
4.3. Differences in Water-Extractable C and N between Tree Species
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plot ID | Species | Native/Exotic | Latitude | Longitude | Elevation | Age in 2016 (Years) |
|---|---|---|---|---|---|---|
| Plot273 | Calliandra calothyrsus (C. cal) | Exotic | 02°36′69″ S | 29°45′30″ E | 1722 m | 31 |
| Plot265 | Calliandra calothyrsus (C. cal) | Exotic | 02°36′71″ S | 29°45′18″ E | 1713 m | 31 |
| Plot267 | Calliandra calothyrsus (C. cal) | Exotic | 02°36′72″ S | 29°45′21″ E | 1714 m | 31 |
| Plot56 | Cedrela serrata (C. ser) | Exotic | 02°36′94″ S | 29°44′79″ E | 1713 m | 70 |
| Plot111 | Cedrela serrata (C. ser) | Exotic | 02°36′75″ S | 29°45′60″ E | 1709 m | 79 |
| Plot36 | Cedrela serrata (C. ser) | Exotic | 02°36′83″ S | 29°45′30″ E | 1730 m | 73 |
| Plot150 | Grevillea robusta (G. rob) | Exotic | 02°36′97″ S | 29°44′96″ E | 1713 m | 75 |
| Plot322 | Grevillea robusta (G. rob) | Exotic | 02°36′94″ S | 29°45′19″ E | 1709 m | 69 |
| Plot104 | Grevillea robusta (G. rob) | Exotic | 02º36.84 S | 29º45.51 E | 1720 m | 35 |
| Plot218 | Eucalyptus grandis (E. gr) | Exotic | 02°37′03″ S | 29°44′83″ E | 1707 m | 70 |
| Plot220 | Eucalyptus grandis (E. gr) | Exotic | 02°37′05″ S | 29°44′86″ E | 1706 m | 65 |
| Plot181 | Eucalyptus grandis (E. gr) | Exotic | 02°36′65″ S | 29°45′64″ E | 1680 m | 65 |
| Plot179 | Eucalyptus maideni (E. mai) | Exotic | 02°36′66″ S | 29°45′61″ E | 1685 m | 70 |
| Plot377 | Eucalyptus maideni (E. mai) | Exotic | 02°36′59″ S | 29°45′32″ E | 1695 m | 82 |
| Plot1 | Eucalyptus maideni (E. mai) | Exotic | 02°36′89″ S | 29°44′78″ E | 1732 m | 67 |
| Plot472 | Eucalyptus saligna (E. sal) | Exotic | 02°37′01″ S | 29°45′12″ E | 1710 m | 82 |
| Plot259 | Eucalyptus saligna (E. sal) | Exotic | 02°36′93″ S | 29°45′38″ E | 1709 m | 36 |
| Plot20 | Eucalyptus saligna (E. sal) | Exotic | 02°36′89″ S | 29°45′06″ E | 1729 m | 59 |
| Plot78 | Entandrophragma excelsum (E. exc) | Native | 02°36′90″ S | 29°45′12″ E | 1727 m | 67 |
| Plot44 | Entandrophragma excelsum (E. exc) | Native | 02°36′81″ S | 29°45′42″ E | 1727 m | 64 |
| Plot54 | Entandrophragma excelsum (E. exc) | Native | 02°36′78″ S | 29°45′57″ E | 1718 m | 45 |
| Plot240 | Polyscias fulva (P. ful) | Native | 02°36′96″ S | 29°45′15″ E | 1714 m | 80 |
| Plot262 | Polyscias fulva (P. ful) | Native | 02°36′91″ S | 29°45′46″ E | 1695 m | 80 |
| Plot268 | Polyscias fulva (P. ful) | Native | 02°36′88″ S | 29°45′54″ E | 1693 m | 80 |
| MNS1 | Mix natives species (Mns) | Native | 02°36′65″ S | 29°44′65″ E | 1700 m | 83 |
| MNS1 | Mix natives species (Mns) | Native | 02°36′68″ S | 29°45′51″ E | 1692 m | 83 |
| MNS3 | Mix natives species (Mns) | Native | 02°36′59″ S | 29°45′63″ E | 1680 m | 83 |
| Soil Parameters | Layer | N | Mean | SD | Median | Min | Max | Skew | Kurtosis | SE |
|---|---|---|---|---|---|---|---|---|---|---|
| pHKCL | 0–5 cm | 54 | 4.89 | 0.71 | 4.96 | 3.71 | 5.89 | −0.33 | −1.38 | 0.1 |
| 5–10 cm | 54 | 4.21 | 0.36 | 4.24 | 3.71 | 4.83 | 0.04 | −1.25 | 0.05 | |
| SOM (%) | 0–5 cm | 54 | 22.49 | 4.71 | 20.55 | 15.53 | 31.32 | 0.63 | −1.03 | 0.64 |
| 5–10 cm | 54 | 9.6 | 1.4 | 9.87 | 5.93 | 11.56 | −0.95 | 0.21 | 0.19 | |
| SOC (gkg−1) | 0–5 cm | 54 | 130.4 | 27.31 | 119.2 | 90.1 | 181.66 | 0.63 | −1.04 | 3.72 |
| 5–10 cm | 54 | 55.67 | 8.13 | 57.22 | 34.41 | 67.06 | −0.94 | 0.2 | 1.11 | |
| WSC (mgkg−1) | 0–5 cm | 54 | 340.8 | 115.1 | 323 | 175.2 | 683.5 | 1.1 | 1.05 | 15.6 |
| 5–10 cm | 54 | 56.31 | 9.41 | 55.9 | 39.77 | 78.74 | 0.36 | −0.61 | 1.28 | |
| WSNO3 (mgkg−1) | 0–5 cm | 54 | 67.7 | 28.82 | 58.13 | 32.77 | 145.64 | 1.34 | 0.93 | 3.92 |
| 5–10 cm | 54 | 12.31 | 4.68 | 12.3 | 3.93 | 20.82 | 0.18 | −1.07 | 0.64 | |
| WSNH4 (mgkg−1) | 0–5 cm | 54 | 12.85 | 2.97 | 12.97 | 7.62 | 18.34 | −0.12 | −1.28 | 0.4 |
| 5–10 cm | 54 | 1.23 | 0.67 | 0.95 | 0.47 | 2.9 | 0.96 | −0.22 | 0.09 | |
| WSNorg (mgkg−1) | 0–5 cm | 54 | 34.73 | 9.74 | 34.41 | 15.84 | 57.02 | 0.22 | −0.51 | 1.32 |
| 5–10 cm | 54 | 6.85 | 1.74 | 6.56 | 2.99 | 10.42 | 0.03 | −0.48 | 0.24 | |
| WSNmin (mgkg−1) | 0–5 cm | 54 | 80.54 | 29.07 | 70.65 | 45.01 | 162.93 | 1.47 | 1.38 | 3.96 |
| 5–10 cm | 54 | 13.54 | 4.34 | 13.4 | 5.53 | 21.33 | 0.22 | −1.2 | 0.59 | |
| WSNtot (mgkg−1) | 0–5 cm | 54 | 115.2 | 29.68 | 111.2 | 65.1 | 190.1 | 0.97 | 0.63 | 4.04 |
| 5–10 cm | 54 | 20.39 | 4.86 | 20.38 | 12.86 | 28.78 | 0.1 | −1.48 | 0.66 | |
| WSC/WSNorg | 0–5 cm | 54 | 10.07 | 2.86 | 9.28 | 5.89 | 18.79 | 1.13 | 1.33 | 0.39 |
| 5–10 cm | 54 | 8.66 | 2.21 | 8.43 | 4.61 | 15.5 | 0.96 | 1.55 | 0.3 | |
| WSC/WSNtot | 0–5 cm | 54 | 3.11 | 1.2 | 2.71 | 1.34 | 6.39 | 0.86 | 0.19 | 0.16 |
| 5–10 cm | 54 | 2.88 | 0.66 | 2.87 | 1.5 | 4.43 | 0.02 | −0.63 | 0.09 | |
| HWC (mgkg−1) | 0–5 cm | 54 | 3994.6 | 1201.2 | 3904.7 | 2203.4 | 5893.4 | 0.11 | −1.51 | 163.4 |
| 5–10 cm | 54 | 603.43 | 119.8 | 601.8 | 382.16 | 888.07 | 0.48 | −0.33 | 16.31 | |
| HWNO3 (mgkg−1) | 0–5 cm | 54 | 14.13 | 4.86 | 14.11 | 4.57 | 23.36 | -0.03 | −0.79 | 0.66 |
| 5–10 cm | 54 | 2.22 | 0.85 | 2.01 | 0.83 | 3.92 | 0.32 | −0.94 | 0.12 | |
| HWNH4 (mgkg−1) | 0–5 cm | 54 | 71.7 | 18.6 | 71.05 | 37.07 | 104.15 | -0.01 | −0.95 | 2.53 |
| 5–10 cm | 54 | 8.35 | 3.15 | 7.97 | 4.18 | 14.97 | 0.5 | −0.91 | 0.43 | |
| HWNorg (mgkg−1) | 0–5 cm | 54 | 301.64 | 62.12 | 294.93 | 194.96 | 440.04 | 0.18 | −1.2 | 8.45 |
| 5–10 cm | 54 | 55.44 | 16.45 | 53.03 | 31.94 | 95.43 | 0.9 | −0.04 | 2.24 | |
| HWNtot (mgkg−1) | 0–5 cm | 54 | 387.47 | 73.9 | 388.71 | 242.65 | 536.49 | −0.1 | −1 | 10.06 |
| 5–10 cm | 54 | 66.01 | 19.36 | 61.47 | 40.02 | 111.71 | 0.9 | −0.01 | 2.63 | |
| HWC/HWNorg | 0–5 cm | 54 | 13.08 | 2.12 | 12.64 | 10.08 | 17.6 | 0.37 | −1.14 | 0.29 |
| 5–10 cm | 54 | 11.3 | 2.15 | 10.93 | 8.52 | 17.22 | 0.86 | 0.03 | 0.29 | |
| HWC/HWNtot | 0–5 cm | 54 | 10.2 | 1.91 | 9.85 | 7.45 | 13.77 | 0.34 | −1.26 | 0.26 |
| 5–10 cm | 54 | 9.48 | 1.84 | 9.16 | 7.01 | 14.21 | 0.89 | 0.09 | 0.25 | |
| Al3+ (cmolc kg−1) | 0–5 cm | 54 | 0.06 | 0.05 | 0.04 | 0.00 | 0.16 | 0.93 | −0.31 | 0.01 |
| 5–10 cm | 54 | 1.22 | 0.9 | 0.94 | 0.01 | 2.82 | 0.38 | −1.37 | 0.12 | |
| Ca2+ (cmolc kg−1) | 0–5 cm | 54 | 28.5 | 7.89 | 27.27 | 17.39 | 52.9 | 1.42 | 1.74 | 1.07 |
| 5–10 cm | 54 | 5.97 | 3.55 | 5.97 | 1.54 | 16.6 | 1.02 | 0.81 | 0.48 | |
| Fe2+ (cmolc kg−1) | 0–5 cm | 54 | 0.00 | 0.00 | 0.00 | 0.00 | 0.01 | 1.22 | 0.89 | 0.00 |
| 5–10 cm | 54 | 0.01 | 0.03 | 0.00 | 0.00 | 0.25 | 6.93 | 46.94 | 0.00 | |
| K+ (cmolc kg−1) | 0–5 cm | 54 | 0.61 | 0.17 | 0.57 | 0.31 | 0.96 | 0.29 | −0.94 | 0.02 |
| 5–10 cm | 54 | 0.21 | 0.14 | 0.15 | 0.08 | 0.56 | 1.47 | 0.81 | 0.02 | |
| Mg2+ (cmolc kg−1) | 0–5 cm | 54 | 6.9 | 2.87 | 5.67 | 4.45 | 15.17 | 1.74 | 1.94 | 0.39 |
| 5–10 cm | 54 | 1.51 | 0.55 | 1.49 | 0.67 | 2.34 | 0.05 | −1.42 | 0.07 | |
| Mn2+ (cmolc kg−1) | 0–5 cm | 54 | 0.09 | 0.07 | 0.06 | 0.02 | 0.29 | 1.22 | 0.68 | 0.01 |
| 5–10 cm | 54 | 0.01 | 0.01 | 0.01 | 0.00 | 0.03 | 0.35 | −0.88 | 0.00 | |
| Na+ (cmolc kg−1) | 0–5 cm | 54 | 0.15 | 0.06 | 0.14 | 0.07 | 0.28 | 0.32 | −1.16 | 0.01 |
| 5–10 cm | 54 | 0.18 | 0.03 | 0.19 | 0.09 | 0.24 | -0.74 | 0.86 | 0.00 | |
| Zn2+ (cmolc kg−1) | 0–5 cm | 54 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 1.08 | 0.57 | 0.00 |
| 5–10 cm | 54 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.96 | −0.28 | 0.00 | |
| EBC (cmolc kg−1) | 0–5 cm | 54 | 36.16 | 10.46 | 33.96 | 23.98 | 69.18 | 1.75 | 2.39 | 1.42 |
| 5–10 cm | 54 | 7.87 | 4.1 | 7.75 | 2.71 | 19.27 | 0.88 | 0.39 | 0.56 | |
| ∑cations (cmolc kg−1) | 0–5 cm | 54 | 36.31 | 10.41 | 34.07 | 24.32 | 69.24 | 1.77 | 2.42 | 1.42 |
| 5–10 cm | 54 | 9.11 | 3.31 | 8.62 | 5.08 | 19.31 | 1.29 | 1.33 | 0.45 |
| Soil Parameters | Layer (cm) | Calliandra calothyrsus | Cedrela serrata | Grevillea robusta | Eucalyptus grandis | Eucalyptus maideni | Eucalyptus saligna | Entandrophragma excelsum | Polyscias fulva | Mixed natives |
|---|---|---|---|---|---|---|---|---|---|---|
| pHKCl | 0–5 | 4.9 ± 0.01 fg | 4.9 ± 0.01 fg | 5.5 ± 0.02 h | 4.0 ± 0.01 b | 4.2 ± 0.01 c | 3.7 ± 0.01 a | 5.5 ± 0.02 h | 5.8 ± 0.01 i | 5.4 ± 0.04 h |
| 5–10 | 4.2 ± 0.0 c | 4.2 ± 0.0 c | 4.4 ± 0.0 d | 3.8 ± 0.0 a | 3.8 ± 0.0 a | 3.7 ± 0.0 a | 4.4 ± 0.0 d | 4.6 ± 0.0 e | 4.8 ± 0.0 f | |
| SOM (%) | 0–5 | 19.1 ± 0.52 bc | 18.1 ± 0.81 b | 19.2 ± 0.26 bc | 21.6 ± 0.59 bcd | 27.6 ± 0.92 d | 25 ± 1.33 cd | 25.8 ± 2.24 d | 18.5 ± 0.46 b | 27.5 ± 2.2 d |
| 5–10 | 10.2 ± 0.1 a | 9.19 ± 1.0 a | 8.93 0.7 a | 10.3 ± 0.5 a | 9.58 ± 0.08 a | 9.27 ± 0.4 a | 9.5 ± 0.5 a | 10.3 ± 0.2 a | 9.06 ± 0.7 a | |
| SOC (g kg−1) | 0–5 | 110 ± 3 bc | 100 ± 4.7 b | 110 ± 1.6 bc | 160 ± 5.4 d | 140 ± 7.7 cd | 150 ± 13 d | 160 ± 13 d | 110 ± 2.7 b | 130 ± 3.4 bcd |
| 5–10 | 59 ± 0.57 a | 53 ± 6 a | 52 ± 4.3 a | 56 ± 0.52 a | 54 ± 2.8 a | 55 ± 3.2 a | 53 ± 4.1 a | 60 ± 1.3 a | 60 ± 3 a | |
| Al3+ (cmolc kg−1) | 0–5 | 0.079 ± 0.00 a | 0.085 ± 0.01 a | 0.026 ± 0.00 a | 0.006 ± 0.00 a | 0.067 ± 0.01 a | 0.04 ± 0.00 a | 0.15 ± 0.00 a | 0.01 ± 0.00 a | 0.03 ± 0.00 a |
| 5–10 | 0.93 ± 0.035 d | 1.2 ± 0.036 e | 0.77 ± 0.04 d | 0.027 ± 0.00 a | 2.6 ± 0.083 h | 2.2 ± 0.05 f | 2.4 ± 0.059 g | 0.34 ± 0.01 b | 0.53 ± 0.01 c | |
| Ca2+ (cmolckg−1) | 0–5 | 19 ± 0.77 e | 24 ± 1.3 fg | 28 ± 0.33 hi | 27 ± 0.45 gh | 29 ± 0.59 hi | 31 ± 1.6 i | 23 ± 0.85 f | 28 ± 0.25 hi | 48 ± 1.2 j |
| 5–10 | 6 ± 0.06 bc | 4.8 ± 0.067 abc | 7 ± 0.47 bc | 14 ± 0.64 d | 1.6 ± 0.026 a | 3.8 ± 0.097 ab | 2.2 ± 0.081 a | 7.8 ± 0.15 c | 6.7 ± 0.18 bc | |
| Fe2+ (cmolckg−1) | 0–5 | 0.004 ± 0.00 ef | 0.001 ± 0.00 ab | 0.001 ± 0.00 a | 0.0008 ± 0.00 a | 0.002 ± 0.00 abcd | 0.002 ± 0.00 abc | 0.002 ± 0.00 abcd | 0.001 ± 0.00 ab | 0.001 ± 0.00 a |
| 5–10 | 0.003 ± 0.0 bcdef | 0.003 ± 0.00 abcdef | 0.004 ± 0.00 def | 0.0028 ± 0.00 abcdef | 0.005 ± 0.00 f | 0.045 ± 0.04 cdef | 0.004 ± 0.00 def | 0.003 ± 0.00 abcde | 0.003 ± 0.00 abcde | |
| K+ (cmolckg−1) | 0–5 | 0.63 ± 0.023 efg | 0.52 ± 0.016 cde | 0.7 ± 0.045 gh | 0.81 ± 0.018 hi | 0.41 ± 0.014 bc | 0.47 ± 0.012 cd | 0.43 ± 0.042 bcd | 0.64 ± 0.01 fg | 0.85 ± 0.04 i |
| 5–10 | 0.12 ± 0.01 a | 0.12 ± 0.0091 a | 0.13 ± 0.018 a | 0.53 ± 0.009 def | 0.14 ± 0.0044 a | 0.15 ± 0.0083 a | 0.17 ± 0.016 a | 0.17 ± 0.00 a | 0.35 ± 0.01 b | |
| Mg2+ (cmolckg−1) | 0–5 | 6.3 ± 0.14 h | 4.9 ± 0.08 ef | 5.6 ± 0.036 g | 5.1 ± 0.047 fg | 8.2 ± 0.18 i | 4.6 ± 0.053 e | 5.5 ± 0.099 g | 7.7 ± 0.09 i | 14 ± 0.22 j |
| 5–10 | 1.5 ± 0.042 c | 1.5 ± 0.035 c | 1.4 ± 0.089 bc | 2.2 ± 0.054 d | 0.82 ± 0.015 a | 0.91 ± 0.098 a | 0.92 ± 0.048 ab | 2.1 ± 0.04 d | 2.2 ± 0.03 d | |
| Mn2+ (cmolckg−1) | 0–5 | 0.26 ± 0.01 g | 0.061 ± 0.00 d | 0.056 ± 0.00 cd | 0.019 ± 0.00 ab | 0.1 ± 0.00 e | 0.13 ± 0.00 f | 0.12 ± 0.01 ef | 0.02 ± 0.00 ab | 0.033 ± 0.00 bc |
| 5–10 | 0.024 ± 0.00 ab | 0.02 ± 0.00 ab | 0.009 ± 0.00 ab | 0.004 ± 0.00 a | 0.017 ± 0.00 ab | 0.013 ± 0.00 ab | 0.022 ± 0.00 ab | 0.006 ± 0.00 a | 0.013 ± 0.00 ab | |
| Na+ (cmolckg−1) | 0–5 | 0.091 ± 0.01 a | 0.1 ± 0.00 a | 0.11 ± 0.00 a | 0.12 ± 0.017 ab | 0.19 ± 0.01 cde | 0.21 ± 0.0045 e | 0.21 ± 0.01 e | 0.13 ± 0.02 abc | 0.18 ± 0.02 bcde |
| 5–10 | 0.19 ± 0.01 cde | 0.18 ± 0.00 bcde | 0.2 ± 0.00 de | 0.17 ± 0.0077 bcde | 0.2 ± 0.00 de | 0.18 ± 0.015 bcde | 0.19 ± 0.00 cde | 0.17 ± 0.01 bcde | 0.14 ± 0.013 abcd | |
| Zn2+ (cmolckg−1) | 0–5 | 0.002 ± 0.00 e | 0.001 ± 0.00 abc | 0.001 ± 0.00 bc | 0.0009 ± 0.00 abc | 0.0007 ± 0.00 abc | 0.0008 ± 0.00 abc | 0.0012 ± 0.00 cd | 0.0006 ± 0.00 abc | 0.0017 ± 0.00 de |
| 5–10 | 0.001 ± 0.00 bcd | 0.0006 ± 0.00 abc | 0.0005 ± 0.00 a | 0.0007 ± 0.00 abc | 0.0006 ± 0.00 abc | 0.0006 ± 0.00 abc | 0.0006 ± 0.00 ab | 0.0006 ± 0.00 abc | 0.0011 ± 0.00 bc | |
| EBC (cmolckg−1) | 0–5 | 26 ± 0.89 f | 30 ± 1.2 fg | 34 ± 0.36 h | 33 ± 0.48 gh | 38 ± 0.75 i | 36 ± 1.6 hi | 29 ± 0.9 f | 36 ± 0.31 hi | 63 ± 1.4 j |
| 5–10 | 7.9 ± 0.05 cd | 6.6 ±0.08 bcd | 8.7 ± 0.57 d | 17 ± 0.6 e | 2.8 ± 0.02 a | 5 ± ±0.12 abc | 3.4 ± 0.1 ab | 10 ± 0.19 d | 9.3 ± 0.2 d | |
| Cations (cmolckg−1) | 0–5 | 27 ± 0.9 e | 30 ± 1.2 ef | 34 ± 0.36 g | 33 ± 0.47 fg | 38 ± 0.74 h | 36 ± 1.6 gh | 29 ± 0.91 e | 36 ± 0.31 gh | 63 ± 1.4 i |
| 5–10 | 8.8 ± 0.06 abc | 7.8 ± 0.11 abc | 9.5 ± 0.55 bc | 17 ± 0.6 d | 5.4 ± 0.07 a | 7.2 ± 0.09 abc | 5.9 ± 0.07 ab | 11 ± 0.19 c | 9.9 ± 0.2 c |
| Soil Layer | Water Extract. N Fractions | Calliandra Calothyrsus | Cedrela Serrata | Grevillea Robusta | Eucalyptus Grandis | Eucalyptus Maideni | Eucalyptus Saligna | Entandrophragma Excelsum | Polyscias Fulva | Mixed natives |
|---|---|---|---|---|---|---|---|---|---|---|
| Water-soluble fractions | ||||||||||
| 0–5 cm | WSNO3 (%) | 43 | 55 | 73 | 52 | 54 | 46 | 12 | 68 | 64 |
| WSNH4 (%) | 13 | 18 | 9 | 10 | 13 | 16 | 27 | 6 | 9 | |
| WSNorg (%) | 44 | 27 | 18 | 38 | 33 | 38 | 61 | 26 | 27 | |
| 5–10 cm | WSNO3 (%) | 60 | 45 | 76 | 58 | 65 | 55 | 41 | 62 | 64 |
| WSNH4 (%) | 7 | 12 | 2 | 5 | 4 | 8 | 15 | 4 | 3 | |
| WSNorg (%) | 33 | 43 | 22 | 37 | 31 | 37 | 44 | 34 | 33 | |
| Hot water-extractable fractions | ||||||||||
| 0–5 cm | HWNO3 (%) | 4 | 3 | 5 | 3 | 5 | 3 | 3 | 3 | 3 |
| HWNH4 (%) | 17 | 16 | 22 | 16 | 15 | 15 | 19 | 25 | 23 | |
| HWNorg (%) | 79 | 81 | 73 | 81 | 80 | 82 | 78 | 72 | 74 | |
| 5–10 cm | HWNO3 (%) | 4 | 3 | 4 | 4 | 4 | 4 | 2 | 3 | 3 |
| HWNH4 (%) | 15 | 15 | 10 | 11 | 10 | 11 | 17 | 11 | 13 | |
| HWNorg (%) | 81 | 82 | 86 | 85 | 86 | 85 | 81 | 86 | 84 | |
References
- Binkley, D.; Menyailo, O. Gaining insights on the effects of tree spiecies on soils. In Tree Species Effects on Soils: Implications for Global Change; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2005; pp. 1–16. [Google Scholar]
- Carnol, M.; Bazgir, M. Nutrient return to the forest floor through litter and throughfall under 7 forest species after conversion from Norway spruce. For. Ecol. Manag. 2013, 309, 66–75. [Google Scholar] [CrossRef]
- Bauters, M.; Ampoorter, E.; Huygens, D.; Kearsley, E.; De Haulleville, T.; Sellan, G.; Verbeeck, H.; Boeckx, P.; Verheyen, K. Functional identity explains carbon sequestration in a 77-year-old experimental tropical plantation. Ecosphere 2015, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Lathwell, D.J.; Grove, T.L. Soil-Plant Relationships in the Tropics. Ecology 2011, 17, 1–16. [Google Scholar] [CrossRef]
- Hobbie, S.E.; Reich, P.; Oleksyn, J.; Ogdahl, M.; Zytkowiak, R.; Hale, C.; Karolewski, P. Tree Species Effects on decomposition and forest floor dynamics in a common garden. Ecology 2006, 87, 2288–2297. [Google Scholar] [CrossRef]
- Prescott, C.E.; Vesterdal, L. Tree species effects on soils in temperate and boreal forests: Emerging themes and research needs. For. Ecol. Manag. 2013, 309, 1–3. [Google Scholar] [CrossRef]
- Veldkamp, A. Pedogenesis and Soil Forming Factors. In Land Use and Land Cover; Encyclopedia of Life Support Systems (EOLSS): Wageningen, The Netherlands, 2000. [Google Scholar]
- Sayer, E.J.; Banin, L.F. Tree Nutrient Status and Nutrient Cycling in Tropical Forest—Lessons from Fertilization Experiments. Tree Physiol. 2016, 6, 275–297. [Google Scholar] [CrossRef]
- Schulte, A.; Ruhiyat, D. Soils of Tropical Forest Ecosystems; Springer: Berlin/Heidelberg, Germany, 1998. [Google Scholar]
- Karlen, D.L.; Mausbach, M.J.; Doran, J.W.; Cline, R.G.; Harris, R.F.; Schuman, G.E. Soil Quality: A Concept, Definition, and Framework for Evaluation (A Guest Editorial). Soil Sci. Soc. Am. J. 1997, 61, 4–10. [Google Scholar] [CrossRef] [Green Version]
- Salomé, C.; Coll, P.; Lardo, E.; Metay, A.; Villenave, C.; Marsden, C.; Blanchart, E.; Hinsinger, P.; Le Cadre, E. The soil quality concept as a framework to assess management practices in vulnerable agroecosystems: A case study in Mediterranean vineyards. Ecol. Indic. 2016, 61, 456–465. [Google Scholar] [CrossRef]
- Baveye, P.C.; Ebaveye, J.; Egowdy, J. Soil “Ecosystem” Services and Natural Capital: Critical Appraisal of Research on Uncertain Ground. Front. Environ. Sci. 2016, 4, 41. [Google Scholar] [CrossRef]
- Bünemann, E.K.; Bongiorno, G.; Bai, Z.; Creamer, R.E.; De Deyn, G.; de Goede, R.; Fleskens, L.; Geissen, V.; Kuyper, T.W.; Mäder, P.; et al. Soil quality—A critical review. Soil Biol. Biochem. 2018, 120, 105–125. [Google Scholar] [CrossRef]
- Greiner, L.; Keller, A.; Grêt-Regamey, A.; Papritz, A. Soil function assessment: Review of methods for quantifying the contributions of soils to ecosystem services. Land Use Policy 2017, 69, 224–237. [Google Scholar] [CrossRef]
- Haines-Young, R.; Potschin, M. England’s Terrestrial Ecosystem Services and Rationale for an Ecosystem Approach; Ecosystem Services: London, UK, 2008. [Google Scholar]
- Kennedy, A.C. Biological Indicators of Soil Health. J. Environ. Qual. 1998, 27, 1275–1276. [Google Scholar] [CrossRef]
- Vogel, H.-J.; Eberhardt, E.; Franko, U.; Lang, B.; Ließ, M.; Weller, U.; Wiesmeier, M.; Wollschläger, U. Quantitative Evaluation of Soil Functions: Potential and State. Front. Environ. Sci. 2019, 7, 164. [Google Scholar] [CrossRef]
- Augusto, L.; Ranger, J.; Binkley, D.; Rothe, A. Impact of several common tree species of European temperate forests on soil fertility. Ann. For. Sci. 2002, 59, 233–253. [Google Scholar] [CrossRef] [Green Version]
- Franzluebbers, A. Soil organic matter stratification ratio as an indicator of soil quality. Soil Tillage Res. 2002, 66, 95–106. [Google Scholar] [CrossRef]
- Wellock, M.L.; LaPerle, C.M.; Kiely, G. What is the impact of afforestation on the carbon stocks of Irish mineral soils? For. Ecol. Manag. 2011, 262, 1589–1596. [Google Scholar] [CrossRef]
- DeGryze, S.; Six, J.; Paustian, K.; Morris, S.J.; Paul, E.A.; Merckx, R. Soil organic carbon pool changes following land-use conversions. Glob. Chang. Biol. 2004, 10, 1120–1132. [Google Scholar] [CrossRef] [Green Version]
- Peri, P.L.; Gargaglione, V.; Pastur, G.J.M.; Lencinas, M.V. Carbon accumulation along a stand development sequence of Nothofagus antarctica forests across a gradient in site quality in Southern Patagonia. For. Ecol. Manag. 2010, 260, 229–237. [Google Scholar] [CrossRef]
- Del Galdo, I.; Six, J.; Peressotti, A.; Cotrufo, M.F. Assessing the impact of land-use change on soil C sequestration in agricultural soils by means of organic matter fractionation and stable C isotopes. Glob. Chang. Biol. 2003, 9, 1204–1213. [Google Scholar] [CrossRef]
- Mao, R.; Zeng, D.-H.; Hu, Y.-L.; Li, L.-J.; Yang, D. Soil organic carbon and nitrogen stocks in an age-sequence of poplar stands planted on marginal agricultural land in Northeast China. Plant Soil 2010, 332, 277–287. [Google Scholar] [CrossRef]
- Ross, D.; Tate, K.; Scott, N.; Feltham, C. Land-use change: Effects on soil carbon, nitrogen and phosphorus pools and fluxes in three adjacent ecosystems. Soil Biol. Biochem. 1999, 31, 803–813. [Google Scholar] [CrossRef]
- Paul, K.; Polglase, P.; Nyakuengama, J.; Khanna, P. Change in soil carbon following afforestation. For. Ecol. Manag. 2002, 168, 241–257. [Google Scholar] [CrossRef]
- Nyberg, G.; Ekblad, A.; Buresh, R.; Högberg, P. Short-term patterns of carbon and nitrogen mineralisation in a fallow field amended with green manures from agroforestry trees. Biol. Fertil. Soils 2002, 36, 18–25. [Google Scholar] [CrossRef]
- Curtin, D.; Beare, M.H.; Qiu, W. Distinguishing functional pools of soil organic matter based on solubility in hot water. Soil Res. 2021, 59, 319. [Google Scholar] [CrossRef]
- Paul, E.A. The nature and dynamics of soil organic matter: Plant inputs, microbial transformations, and organic matter stabilization. Soil Biol. Biochem. 2016, 98, 109–126. [Google Scholar] [CrossRef] [Green Version]
- Choudhary, O.; Gill, J.; Singh, B. Water-Extractable Carbon Pools and Microbial Biomass Carbon in Sodic Water-Irrigated Soils Amended with Gypsum and Organic Manures. Pedosphere 2013, 23, 88–97. [Google Scholar] [CrossRef]
- Haynes, R. Labile Organic Matter Fractions as Central Components of the Quality of Agricultural Soils: An Overview. Adv. Agron. 2005, 85, 221–268. [Google Scholar] [CrossRef]
- Strosser, E. Methods for determination of labile soil organic matter: An overview. J. Agrobiol. 2010, 27, 49–60. [Google Scholar] [CrossRef]
- Zhang, M.; He, Z.; Zhao, A.; Zhang, H.; Endale, D.M.; Schomberg, H.H. Water-Extractable Soil Organic Carbon and Nitrogen Affected by Tillage and Manure Application. Soil Sci. 2011, 176, 307–312. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, S.; Huang, Y. Comparisons of litterfall, litter decomposition and nutrient return in a monoculture Cunninghamia lanceolata and a mixed stand in southern China. For. Ecol. Manag. 2008, 255, 1210–1218. [Google Scholar] [CrossRef]
- Habiyaremye, G.; Jiwen, G.; de la Paix Mupenzi, J.; Balogun, W.O. Demographic pressure impacts on forests in Rwanda. Afr. J. Agric. Res. 2011, 6, 4533–4538. [Google Scholar] [CrossRef]
- Clay, D.C.; Lewis, L.A. Land use, soil loss, and sustainable agriculture in Rwanda. Hum. Ecol. 1990, 18, 147–161. [Google Scholar] [CrossRef] [Green Version]
- MINILAF. Forest Sector Strategic Plan 2018–2024; GoR: Kigali, Rwanda, 2018. [Google Scholar]
- Warnest, M.; Sagashya, D.; Nkurunziza, E. Emerging in a Changing Climate-Sustainable Land Use Management in Rwanda; International Federation of Surveyors (FIG): Rome, Italy, 2012. [Google Scholar]
- RFA. Rwanda Forestry Status; GoR: Kigali, Rwanda, 2020; Available online: https://www.rfa.rw (accessed on 6 May 2020).
- MoE. Rwanda Forest cover Mapping; GoR: Kigali, Rwanda, 2019. [Google Scholar]
- IUCN. A Thousand Hills Turn Green: How Rwanda Became a Leader in Forest Landscape restoration. Int. Union Conserv. Nat. 2020. Available online: https://iucn.maps.arcgis.com/apps/MapJournal/index.html?appid=0c40be7f83c64bb9b7c8f3372358c6f9 (accessed on 4 April 2020).
- Bauters, M.; Verbeeck, H.; Doetterl, S.; Ampoorter, E.; Baert, G.; Vermeir, P.; Verheyen, K.; Boeckx, P. Functional Composition of Tree Communities Changed Topsoil Properties in an Old Experimental Tropical Plantation. Ecosystems 2016, 20, 861–871. [Google Scholar] [CrossRef]
- Bauters, M.; Verbeeck, H.; Demol, M.; Bruneel, S.; Taveirne, C.; Van der Heyden, D.; Cizungu, L.; Boeckx, P. Parallel functional and stoichiometric trait shifts in South American and African forest communities with elevation. Biogeosciences 2017, 14, 5313–5321. [Google Scholar] [CrossRef] [Green Version]
- Nsabimana, L.; Klemedtson, B.; Kaplin, G. Wallin, Soil carbon and nutrient accumulation under forest plantations in southern Rwanda. African J. Environ. Sci. Technol. 2008, 2, 142–149. [Google Scholar]
- Meteorwanda, Climatology of Rwanda. 2021. Available online: https://www.meteorwanda.gov.rw/index.php?id=30 (accessed on 30 April 2021).
- Nsabimana, D.; Klemedtson, L.; Kaplin, B.; Wallin, G. Soil CO2 flux in six monospecific forest plantations in Southern Rwanda. Soil Biol. Biochem. 2009, 41, 396–402. [Google Scholar] [CrossRef] [Green Version]
- Steiner, K. Using farmers’ knowledge of soils in making research results more relevant to field practice: Experiences from Rwanda. Agric. Ecosyst. Environ. 1998, 69, 191–200. [Google Scholar] [CrossRef]
- Moeyersons, J. The topographic thresholds of hillslope incisions in southwestern Rwanda. Catena 2003, 50, 381–400. [Google Scholar] [CrossRef]
- Kalinganire, A. Performance of Grevillea robusta in plantations and on farms under varying environmental conditions in Rwanda. For. Ecol. Manage. 1995, 80, 279–285. [Google Scholar] [CrossRef]
- Burren, C. Les Eucalyptus au Rwanda: Analyse de 60 ans D’expérience avec Référence Particulière à l’Arboretum de Ruhande; Switzerland-ISAR/Intercooperation: Bern, Switzerland, 1995. [Google Scholar]
- Commonwealth. Arboretum of Ruhande: A 500 Acre Stretch of Botanical Garden, QCC Proj. 2018. Available online: https://queenscommonwealthcanopy.org/projects/arboretum-of-ruhande (accessed on 30 August 2020).
- ISAR. Arboretum de Ruhande 1987: Note Explicative à L’intention des Visiteurs Élaborée par les Collaborateurs du Département de Foresterie, 3rd ed.; ISAR: Butare, Rwanda, 1987. [Google Scholar]
- Ndayambaje, J.D. State of Rwanda’s Biodiversity for Food and Agriculture; FAO-Rwanda: Kigali, Rwanda, 2016. [Google Scholar]
- Iiyama, M.; Mukuralinda, A.; Ndayambaje, J.D.; Musana, B.; Ndoli, A.; Mowo, J.G.; Garrity, D.; Ling, S.; Ruganzu, V. Tree-Based Ecosystem Approaches (TBEAs) as Multi-Functional Land Management Strategies—Evidence from Rwanda. Sustainability 2018, 10, 1360. [Google Scholar] [CrossRef] [Green Version]
- Bini, D.; dos Santos, C.A.; Bouillet, J.-P.; Gonçalves, J.L.M.; Cardoso, E.J.B.N. Eucalyptus grandis and Acacia mangium in monoculture and intercropped plantations: Evolution of soil and litter microbial and chemical attributes during early stages of plant development. Appl. Soil Ecol. 2013, 63, 57–66. [Google Scholar] [CrossRef]
- Zalamea, M.; González, G.; Lodge, D. Physical, Chemical, and Biological Properties of Soil under Decaying Wood in a Tropical Wet Forest in Puerto Rico. Forests 2016, 7, 168. [Google Scholar] [CrossRef] [Green Version]
- Rorison, I.H.; Allen, S.E.; Grimshaw, H.M.; Parkinson, J.A.; Quarmby, C. Chemical Analysis of Ecological Materials. J. Appl. Ecol. 1976, 13, 650. [Google Scholar] [CrossRef]
- Périé, C.; Ouimet, R. Organic carbon, organic matter and bulk density relationships in boreal forest soils. Can. J. Soil Sci. 2008, 88, 315–325. [Google Scholar] [CrossRef]
- Jenkinson, D.S.; Powlson, D.S. The effects of biocidal treatments on metabolism in soil—V: A method for measuring soil biomass. Soil Biol. Biochem. 1976, 9, 209–213. [Google Scholar] [CrossRef]
- Hendershot, W.H.; Duquette, M. A simple barium chloride for determining cation exchange capacity and exchangeable cations. Soil Sci. Soc. Am. J. 1986, 50, 605–608. [Google Scholar] [CrossRef]
- Ghani, A.; Dexter, M.; Perrott, K. Hot-water extractable carbon in soils: A sensitive measurement for determining impacts of fertilisation, grazing and cultivation. Soil Biol. Biochem. 2003, 35, 1231–1243. [Google Scholar] [CrossRef]
- Gregorich, E.; Beare, M.; Stoklas, U.; St-Georges, P. Biodegradability of soluble organic matter in maize-cropped soils. Geoderma 2003, 113, 237–252. [Google Scholar] [CrossRef]
- Bates, D.; Maechler, M.; Bolker, B.M.; Walker, S.C. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 76, 1–48. [Google Scholar]
- Venables, W.N.; Ripley, B.D. Package MASS. Available online: http://www.r-project.org (accessed on 17 October 2012).
- Barton, K. MuMIn: Multi-Model Inference. 2018. Available online: https://cran.rproject.org/package=MuMIn (accessed on 15 September 2021).
- Gelsomino, A.; Azzellino, A. Multivariate analysis of soils: Microbial biomass, metabolic activity, and bacterial-community structure and their relationships with soil depth and type. J. Plant Nutr. Soil Sci. 2011, 174, 381–394. [Google Scholar] [CrossRef]
- FAO. Topsoil characterization for sustainable land management. In Land and Water Development Division Soil Resources, Management and Conservation Service; FAO: Rome, Italy, 1998; p. 71. [Google Scholar]
- Adugna, A.; Abegaz, A. Effects of soil depth on the dynamics of selected soil properties among the highlands resources of Northeast Wollega, Ethiopia: Are these sign of degradation? Solid Earth Discuss. 2015, 7, 2011–2035. [Google Scholar] [CrossRef]
- Cleveland, C.C.; Townsend, A.R.; Schmidt, S.K.; Constance, B.C. Soil microbial dynamics and biogeochemistry in tropical forests and pastures, southwestern costa rica. Ecol. Appl. 2003, 13, 314–326. [Google Scholar] [CrossRef]
- Yamoah, C.F.; Burleigh, J.R.; Malcolm, M.R. Application of expert systems to study of acid soils in Rwanda. Agric. Ecosyst. Environ. 1990, 30, 203–218. [Google Scholar] [CrossRef]
- Mbonigaba, J.J.M.; Nzeyimana, I.; Bucagu, C.; Culot, M. Caractérisation physique, chimique et microbiologique de trois sols acides tropicaux du Rwanda sous jachères naturelles et contraintes à leur productivité. Biotechnol. Agron. Soc. Environ. 2009, 13, 545–558. [Google Scholar]
- Fujii, K. Soil acidification and adaptations of plants and microorganisms in Bornean tropical forests. Ecol. Res. 2014, 29, 371–381. [Google Scholar] [CrossRef] [Green Version]
- Duchaufour, P. Pédologie: Sol, Végétation, Environnement; Elsevier Masson: Paris, France, 1994. [Google Scholar]
- Dudek, T.; Cuadros, J.; Fiore, S. Interstratified kaolinite-smectite: Nature of the layers and mechanism of smectite kaolinization. Am. Miner. 2006, 91, 159–170. [Google Scholar] [CrossRef]
- Ryan, P.C.; Huertas, F.J. Reaction Pathways of Clay Minerals in Tropical Soils: Insights from Kaolinite-smectite Synthesis Experiments. Clays Clay Miner. 2013, 61, 303–318. [Google Scholar] [CrossRef]
- Celentano, D.; Zahawi, R.; Finegan, B.; Ostertag, R.; Cole, R.J.; Holl, K.D. Litterfall Dynamics Under Different Tropical Forest Restoration Strategies in Costa Rica. Biotropica 2010, 43, 279–287. [Google Scholar] [CrossRef]
- Mugunga, C.P.; Kool, D.; Van Wijk, M.T.; Mohren, G.M.J.; Giller, K.E. Water use by short rotation Eucalyptus woodlots in southern Rwanda. Agrofor. Syst. 2015, 89, 1119–1139. [Google Scholar] [CrossRef] [Green Version]
- Behera, N.; Sahani, U. Soil microbial biomass and activity in response to Eucalyptus plantation and natural regeneration on tropical soil. For. Ecol. Manag. 2003, 174, 1–11. [Google Scholar] [CrossRef]
- Laclau, J.-P.; Ranger, J.; Gonçalves, J.L.M.; Maquère, V.; Krusche, A.; M’Bou, A.T.; Nouvellon, Y.; Saint-André, L.; Bouillet, J.-P.; Piccolo, M.D.C.; et al. Biogeochemical cycles of nutrients in tropical Eucalyptus plantations: Main features shown by intensive monitoring in Congo and Brazil. For. Ecol. Manag. 2010, 259, 1771–1785. [Google Scholar] [CrossRef]
- Rhoades, C.; Binkley, D. Factors influencing decline in soil pH in Hawaiian Eucalyptus and Albizia plantations. For. Ecol. Manag. 1996, 80, 47–56. [Google Scholar] [CrossRef]
- Jobbágy, E.G.; Jackson, R.B. Patterns and mechanisms of soil acidification in the conversion of grasslands to forests. Biogeochemistry 2003, 64, 205–229. [Google Scholar] [CrossRef]
- de Medeiros, E.V.; Duda, G.P.; dos Santos, L.A.R.; Lima, J.R.D.S.; de Almeida-Cortêz, J.S.; Hammecker, C.; Lardy, L.; Cournac, L. Soil organic carbon, microbial biomass and enzyme activities responses to natural regeneration in a tropical dry region in Northeast Brazil. Catena 2017, 151, 137–146. [Google Scholar] [CrossRef]
- Ahmed, A.G.; Murungi, L.K.; Babin, R.; Peng, Y.; Schmidt, I.K.; Zheng, H.; Heděnec, P.; Bachega, L.R.; Yue, K.; Wu, F.; et al. Six, Tree species effects on soil organic matter dynamics: The role of soil cation composition. For. Ecol. Manage. 2015, 36, 119–127. [Google Scholar] [CrossRef]
- Koutika, L.-S.; Epron, D.; Bouillet, J.-P.; Mareschal, L. Changes in N and C concentrations, soil acidity and P availability in tropical mixed acacia and eucalypt plantations on a nutrient-poor sandy soil. Plant Soil 2014, 379, 205–216. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, S.; Yu, X. Decline of soil fertility during forest conversion of secondary forest to Chinese fir plantations in subtropical China. Land Degrad. Dev. 2011, 22, 444–452. [Google Scholar] [CrossRef]
- Bankó, L.; Tóth, G.; Marton, C.L.; Hoffmann, S. Hot-water extractable C and N as indicators for 4p1000 goals in a temperate-climate long-term field experiment: A case study from Hungary. Ecol. Indic. 2021, 126, 107364. [Google Scholar] [CrossRef]
- Gregorich, E.G.; Carter, M.R.; Angers, D.A.; Monreal, C.M.; Ellert, B.H. Towards a minimum data set to assess soil organic matter quality in agricultural soils. Can. J. Soil Sci. 1994, 74, 367–385. [Google Scholar] [CrossRef] [Green Version]
- Ostrowska, A.; Porȩbska, G.; Kanafa, M. Carbon accumulation and distribution in profiles of forest soils. Polish J. Environ. Stud. 2010, 19, 1307–1315. [Google Scholar]
- Russell, A.E.; Raich, J.W.; Valverde-Barrantes, O.J.; Fisher, R.F. Tree Species Effects on Soil Properties in Experimental Plantations in Tropical Moist Forest. Soil Sci. Soc. Am. J. 2007, 71, 1389–1397. [Google Scholar] [CrossRef] [Green Version]
- Landgraf, D.; Leinweber, P.; Makeschin, F. Cold and hot water–extractable organic matter as indicators of litter decomposition in forest soils. J. Plant Nutr. Soil Sci. 2006, 169, 76–82. [Google Scholar] [CrossRef]
- Binkley, D.; Menyailo, O. Tree Species Effects on Soils: Implications for Global Change; Springer: Krasnoyarsk, Russia, 2004; p. 361. [Google Scholar] [CrossRef] [Green Version]
- Ćirić, V.; Belić, M.; Nešić, L.; Šeremešić, S.; Pejić, B.; Bezdan, A.; Manojlović, M. The sensitivity of water extractable soil organic carbon fractions to land use in three soil types. Arch. Agron. Soil Sci. 2016, 62, 1654–1664. [Google Scholar] [CrossRef]



| Labile C and N Fractions | Soil Layer (cm) | Calliandra calothyrsus | Cedrela serrata | Grevillea robusta | Eucalyptus grandis | Eucalyptus maideni | Eucalyptus saligna | Entandrophragma excelsum | Polyscias fulva | Mixed natives |
|---|---|---|---|---|---|---|---|---|---|---|
| Water-soluble C and N fractions | ||||||||||
| WSC (mgkg−1) | 0–5 | 550 ± 49 f | 210 ± 13 b | 320 ± 30 cde | 360 ± 12 cde | 380 ± 29 de | 430 ± 38 e | 310 ± 15 bcd | 250 ± 5 bc | 260 ± 8.9 bc |
| 5–10 | 67 ± 2.5 a | 56± 2.3 a | 50 ± 3.4 a | 52 ± 3.9 a | 60 ± 3.4 a | 50 ± 1.7 a | 53 ± 2.6 a | 50 ± 1.9 a | 68± 3.3 a | |
| WSNtot (mgkg−1) | 0–5 | 120 ± 3.8 f | 73 ± 2.2 c | 180 ± 2.8 h | 100 ± 3.4 de | 100 ± 3.8 de | 96 ± 2 d | 120 ± 1.7 f | 130 ± 3.5 g | 110 ± 2.3 ef |
| 5–10 | 25 ± 0.81 ab | 17 ± 0.73 ab | 26 ± 0.36 b | 16 ± 0.73 ab | 22 ± 0.3 ab | 16 ± 0.99 ab | 15 ± 0.46 a | 21 ± 1.4 ab | 27 ± 0.69 b | |
| WSNorg (mgkg−1) | 0–5 | 51 ± 2 d | 20 ± 1.6 b | 32 ± 2.8 c | 40 ± 2.2 c | 33 ± 4.1 c | 37 ± 1.9 c | 34 ± 2.9 c | 35 ± ±2.7 c | 30 ± 1.3 c |
| 5–10 | 8.1 ± 0.44 a | 7.1 ± 0.5 a | 5.8 ± 0.12 a | 5.9 ± 0.64 a | 6.6 ± 0.37 a | 5.9 ± 0.69 a | 6.6 ± 1.2 a | 7.2 ± 0.83 a | 8.6 ± 0.49 a | |
| WSNH4 (mgkg−1) | 0–5 | 15 ± 0.66 d | 13 ± 0.45 cd | 15 ± 0.75 d | 9.9 ± 0.92 b | 13 ± 1.1 cd | 15 ± 0.53 d | 15 ± 1.2 d | 8.7 ± 0.3 b | 10 ± 0.24 bc |
| 5–10 | 1.8 ± 0.14 a | 2 ± 0.27 a | 0.52 ± 0.01 a | 0.76 ± 0.06 a | 0.9 ±0.07 a | 1.2 ± 0.2 a | 2.2 ± 0.2 a | 0.82 ± 0.03 a | 0.8 ± 0.05 a | |
| WSNO3 (mgkg−1) | 0–5 | 50 ± 2.1 ef | 40 ± 2 d | 130 ± 4.1 i | 54 ± 1.8 f | 54 ± 1.7 f | 44 ± 1.2 de | 70 ± 2.2 g | 91 ± 2.6 h | 70 ± 1.8 g |
| 5–10 | 15 ± 0.49 abc | 7.6 ± 0.55 a | 20 ± 0.39 c | 9.3 ± 0.17 ab | 14 ± 0.54 abc | 8.7 ± 0.21 ab | 6.1 ± 0.93 a | 13 ± 0.89 abc | 17 ± 0.53 bc | |
| WSC/WSNtot | 0–5 | 4.8 ± 0.5 f | 2.9 ± 0.2 abcd | 1.9 ± 0.2 a | 3.5 ± 0.1 bcde | 3.8 ± 0.3 def | 4.5 ± 0.4 ef | 2.9 ± 0.1 abc | 1.9 ± 0.1 a | 2.4 ± 0.0 ab |
| 5–10 | 2.7 ± 0.2 abcd | 3.4 ± 0.1 bcde | 1.9 ± 0.1 a | 3.3 ± 0.3 bcde | 2.8 ± 0.1 abc | 3.2 ± 0.1 bcd | 3.6 ± 0.2 cdef | 2.5 ± 0.2 abc | 2.6 ± 0.1 abc | |
| WSC/WSNorg | 0–5 | 11 ± 1 a | 11 ± 0.9 a | 11 ± 1.8 a | 9.2 ± 0.5 a | 12 ± 1.5 a | 12 ± 1.3 a | 9.3 ± 0.9 a | 7.5 ± 0.6 a | 8.7 ± 0.2 a |
| 5–10 | 8.6 ± 0.83 a | 8 ± 0.34 a | 8.8 ± 0.58 a | 9.6 ± 1.5 a | 9 ± 0.2 a | 8.9 ± 0.79 a | 9.6 ± 1.7 a | 7.4 ± 0.81 a | 8 ± 0.55 a | |
| Hot water-extractable C and N fractions | ||||||||||
| HWC (mgkg−1) | 0–5 | 3200 ± 280 cd | 2500 ± 110 b | 2700 ± 97 bc | 5200 ± 150 ef | 5400 ± 120 f | 5500 ± 140 f | 4600 ± 210 e | 3400 ± 51 d | 3500 ± 230 d |
| 5–10 | 640 ±32 a | 540 ± 25 a | 590 ± 25 a | 500 ± 29 a | 580 ± 25 a | 630 ± 51 a | 500 ± 46 a | 620 ± 31 a | 830 ± 16 a | |
| HWNtot (mgkg−1) | 0–5 | 300 ± 4.5 cd | 270 ± 9.4 c | 330 ± 8 de | 490 ± 13 h | 420 ± 5.2 fg | 440 ± 8 g | 430 ± 17 g | 370 ± 8.9 ef | 430 ± 20 g |
| 5–10 | 80 ± 4.6 ab | 61 ± 1.2 a | 66 ± 4.4 ab | 48± 3.1 a | 52 ± 3.2 a | 51 ± 1.7 a | 55 ± 3.8 a | 72 ± 1.5 ab | 110± 1.3 b | |
| HWNorg (mgkg−1) | 0–5 | 240 ± 1.9 cd | 220 ± 7.5 c | 240 ± 5.5 cd | 400 ± 11 g | 330 ± 4.8 ef | 360 ± 6.2 f | 340 ± 13 ef | 270 ± 6 d | 320 ± 18 e |
| 5–10 | 65 ± 4.3 ab | 50 ± 1.7 a | 57 ± 3.8 ab | 41 ± 2.9 a | 44 ± 2.9 a | 43 ± 1.6 a | 45 ± 3.9 a | 62 ± 1.7 ab | 91 ± 1.2 b | |
| WSNH4 (mgkg−1) | 0–5 | 50 ± 2.4 b | 43 ± 2.1 b | 73 ± 3.1 cde | 77 ± 2.8 de | 63 ± 1.8 c | 68 ± 2 cd | 82 ± 3.7 e | 94 ± 4.9 f | 97 ± 2.1 f |
| 5–10 | 12 ± 0.35 a | 9.1 ± 0.75 a | 6.4 ± 0.42 a | 5.2 ± 0.15 a | 5.1 ± 0.35 a | 5.9 ± 0.81 a | 9.3 ± 0.59 a | 8.2 ± 0.43 a | 14 ± 0.42 a | |
| WSNO3 (mgkg−1) | 0–5 | 12 ± 0.98 cd | 8 ± 0.19 bc | 17 ± 0.7 de | 17 ± 1.4 de | 21 ± 0.97 e | 14 ± 1.3 d | 14 ± 1.2 d | 12 ± 0.92 cd | 12 ± 3.2 cd |
| 5–10 | 3 ± 0.35 ab | 1.6 ± 0.11 a | 2.8 ± 0.25 ab | 2 ± 0.26 a | 2.2 ± 0.29 a | 1.9 ± 0.03 a | 0.99 ± 0.03 a | 2.4 ± 0.29 a | 3.1 ± 0.21 ab | |
| HWC/HWNtot | 0–5 | 11 ± 0.9 bcd | 9.4 ± 0.3 abc | 8 ± 0.1 a | 11 ± 0.1 bcd | 13.9 ± 0.1 e | 13 ± 0.2 de | 11 ± 0.3 bcd | 9.1 ± 0.2 ab | 8 ± 0.1 a |
| 5–10 | 8 ± 0.1 a | 8.9 ± 0.5 ab | 9 ± 0.4 ab | 10 ± 0.3 bcd | 11 ± 0.8 cde | 12 ± 0.6 de | 9.1 ± 0.5 ab | 8.5 ± 0.4 ab | 7.7 ± 0.1 a | |
| HWC/HWNorg | 0–5 | 13 ± 1.2 defg | 12 ± 0.4 abcdef | 11 ± 0.2 abcde | 13 ± 0.2 defg | 16 ± 0.2 h | 15 ± 0.3 gh | 14 ± 0.4 efgh | 13 ± 0.3 cdefg | 11 ± 0.2 abcde |
| 5–10 | 9.8 ± 0.17 abc | 11 ± 0.85 abcde | 10 ± 0.54 abcd | 12 ± 0.38 bcdefg | 13 ± 0.95 defg | 14 ± 0.77 fgh | 11 ± 0.6 abcde | 10 ±0.5 ab | 9.1 ± 0.16 a | |
| Upper Soil Layer (0–5 cm) | Lower Soil Layer (5–10 cm) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Principal Components | PC1 | PC2 | PC3 | PC4 | PC5 | PC1 | PC2 | PC3 | PC4 | PC5 |
| Eigenvalues | 4.69 | 2.03 | 1.57 | 1.03 | 0.71 | 5.02 | 1.472 | 1.19 | 1.02 | 0.89 |
| % variance | 42.66 | 18.52 | 14.27 | 9.43 | 6.48 | 45.64 | 13.38 | 10.85 | 9.34 | 8.16 |
| Cumulative % of the total variance | 42.66 | 61.19 | 75.46 | 84.89 | 91.38 | 45.64 | 59.03 | 69.88 | 79.22 | 87.39 |
| Loadings (weight) of variables on PCs (%) | ||||||||||
| pHKCL | −0.77 | 0.24 | 0.22 | 0.25 | −0.15 | 0.73 | 0.33 | −0.21 | −0.22 | −0.09 |
| SOM | 0.81 | 0.19 | 0.07 | 0.17 | −0.25 | 0.40 | −0.41 | 0.04 | 0.50 | 0.38 |
| WSC | 0.45 | −0.70 | 0.24 | 0.34 | 0.29 | 0.51 | 0.16 | 0.50 | −0.01 | 0.59 |
| WSNorg | 0.22 | −0.45 | 0.80 | −0.20 | 0.21 | 0.70 | −0.58 | −0.04 | −0.19 | 0.18 |
| WSNtot | −0.40 | 0.017 | 0.63 | 0.48 | −0.23 | 0.74 | 0.01 | 0.08 | −0.07 | −0.20 |
| WSC:WSNorg | 0.33 | −0.35 | −0.55 | 0.66 | 0.06 | −0.46 | 0.76 | 0.35 | 0.19 | 0.13 |
| HWC | 0.96 | 0.10 | 0.05 | −0.05 | −0.08 | 0.73 | −0.07 | 0.41 | 0.30 | −0.39 |
| HWNorg | 0.86 | 0.41 | 0.14 | 0.04 | −0.05 | 0.94 | 0.20 | 0.04 | 0.08 | −0.14 |
| HWNtot | 0.77 | 0.53 | 0.23 | 0.14 | −0.05 | 0.95 | 0.19 | 0.04 | 0.05 | −0.09 |
| HWC:HWNtot | 0.77 | −0.35 | −0.11 | −0.24 | −0.03 | −0.64 | −0.35 | 0.46 | 0.29 | −0.32 |
| EBC | −0.01 | 0.71 | 0.02 | 0.14 | 0.64 | 0.12 | 0.18 | −0.59 | 0.67 | 0.04 |
| Contribution of variables to PCs (%) | ||||||||||
| pHKCL | 12.96 | 2.98 | 3.34 | 6.25 | 3.21 | 10.82 | 7.78 | 3.90 | 4.79 | 0.94 |
| SOM | 14.30 | 1.85 | 0.35 | 3.09 | 9.37 | 3.20 | 11.63 | 0.18 | 24.70 | 16.20 |
| WSC | 4.43 | 24.50 | 3.79 | 11.33 | 12.24 | 5.33 | 1.81 | 21.35 | 0.03 | 38.75 |
| WSNorg | 1.03 | 10.07 | 40.84 | 4.15 | 6.21 | 9.79 | 22.89 | 0.15 | 3.78 | 3.92 |
| WSNtot | 3.54 | 0.01 | 26.03 | 22.46 | 7.58 | 11.08 | 0.01 | 0.55 | 0.50 | 4.67 |
| WSC:WSNorg | 2.35 | 6.33 | 19.55 | 42.61 | 0.58 | 4.25 | 39.38 | 10.45 | 3.68 | 1.98 |
| HWC | 19.85 | 0.49 | 0.19 | 0.24 | 0.89 | 10.72 | 0.34 | 14.47 | 9.28 | 17.71 |
| HWNorg | 15.96 | 8.55 | 1.34 | 0.22 | 0.42 | 17.77 | 2.88 | 0.15 | 0.64 | 2.44 |
| HWNtot | 12.76 | 13.79 | 3.63 | 2.01 | 0.37 | 18.31 | 2.62 | 0.16 | 0.32 | 1.06 |
| HWC:HWNtot | 12.77 | 6.21 | 0.84 | 5.56 | 0.19 | 8.36 | 8.37 | 18.14 | 8.36 | 12.02 |
| EBC | 0.01 | 25.16 | 0.05 | 2.02 | 58.90 | 0.30 | 2.23 | 30.45 | 43.87 | 0.25 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rwibasira, P.; Naramabuye, F.X.; Nsabimana, D.; Carnol, M. Long-Term Effects of Forest Plantation Species on Chemical Soil Properties in Southern Rwanda. Soil Syst. 2021, 5, 59. https://doi.org/10.3390/soilsystems5040059
Rwibasira P, Naramabuye FX, Nsabimana D, Carnol M. Long-Term Effects of Forest Plantation Species on Chemical Soil Properties in Southern Rwanda. Soil Systems. 2021; 5(4):59. https://doi.org/10.3390/soilsystems5040059
Chicago/Turabian StyleRwibasira, Peter, Francois Xavier Naramabuye, Donat Nsabimana, and Monique Carnol. 2021. "Long-Term Effects of Forest Plantation Species on Chemical Soil Properties in Southern Rwanda" Soil Systems 5, no. 4: 59. https://doi.org/10.3390/soilsystems5040059
APA StyleRwibasira, P., Naramabuye, F. X., Nsabimana, D., & Carnol, M. (2021). Long-Term Effects of Forest Plantation Species on Chemical Soil Properties in Southern Rwanda. Soil Systems, 5(4), 59. https://doi.org/10.3390/soilsystems5040059