Abstract
Intensive cultivation of ‘uala (sweet potato) in the Leeward Kohala field system on Hawai’i Island supported substantial populations of Native Hawaiians prior to its abandonment in the 19th century. Productivity is influenced by the heterogeneity of the climate and biogeochemical soil characteristics across the substantial ecological gradient. Agricultural infrastructure and associated practices were developed to manage crop production eventuating from the variation in inter- and intra-annual rainfall. Mechanisms of sustaining soil nitrogen (N) are still unclear; however, a pronounced source of N is soil organic matter decomposition. This study investigated in situ the effects of indigenous Hawaiian mulching practices against two control treatments on soil moisture and temperature dynamics to facilitate N mineralization in soil mounds cultivated with ‘uala. Field experiments were set in two agricultural restoration plots with distinct climatic and soil characteristics. Data included soil moisture and temperature, soil and plant N, growth and development of ‘uala, and real-time weather data. Concurrently, N mineralization was also investigated under controlled conditions. All indigenous mulching treatments were found to significantly increase soil moisture, regulate temperature variation, and improve N availability compared to control. Differences in soil properties between treatments translated to significant differences in above-ground biomass. The data suggest that these differences would extend to tuber production, but the use of a long-gestation variety limited tuber production in the study. Increased temperatures in the controlled experiments were observed to increase inorganic N significantly, but less substantially than soil moisture. Indigenous practices in the region could have greatly mitigated plant stress due to moisture, temperature, and N availability, increasing productivity and reducing the variability of the Leeward Kohala Field System.
1. Introduction
For at least four centuries, commencing in the 1400s, a vast agricultural system occupied the central elevations of the Leeward Kohala landscape on Hawai’i island and produced food for populous communities [1]. The ~6500-hectare system, known as the Leeward Kohala Field System (LKFS) (Figure 1), was a dense network of infrastructure used for rain-fed cultivation [1,2,3,4]. The system spans a broad ecological range—extending from sea level to ~750 masl and encompassing ~750 to ~2200 mm/y in rainfall—with the productivity of the systems influenced by rainfall, temperature and soil nutrient levels [5,6].
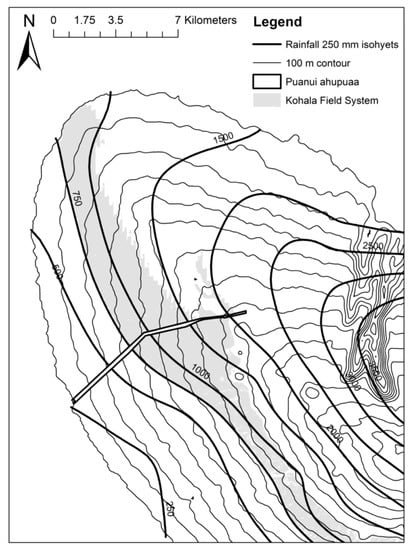
Figure 1.
Map of the study area, depicting the landscape, the modeled extent of the Leeward Kohala Field System, and the ahupua’a of Puanui.
The dominant crop cultivated in the LKFS was ‘uala (sweet potato; Ipomea batatas), likely due to its tolerance to limited moisture conditions [5,7,8] and, arguably, the relatively late introduction of the sweet potato to Hawai’i spurred the expansion of rainfed agriculture in these drier, leeward areas [9]. However, land productivity was, and is, governed by temporal and spatial climatic variability. A critical driver is the intermittent rainfall on both intra- and inter-annual timescales that would have limited tuber production both directly through water limitation and indirectly through water-mediated nutrient availability [5,6,10,11,12]. To mitigate water limitation, indigenous Hawaiians developed place-specific methods to manage soil moisture during cropping seasons [13]. These included the use of rock and vegetative mulch, mounds, construction of berms, and establishment of sugar cane windbreaks/mist traps [3,14,15].
Mechanisms of managing and sustaining soil N in Hawaiian dryland agriculture systems are unclear as weathering of basalt does not supply N [16], there is no confirmation of the incorporation of external N input or N-fixing crops [17], and atmospheric deposition of N is inadequate to sustain intensive cultivation [6]. N mineralization—the conversion of organic N in soil organic matter to inorganic N through microbial decomposition—is an important source for plant-available N [6]. Extracellular proteolytic enzymes of saprophytic bacteria and fungi degrade protein to small peptides and amino acids before finally releasing ammonium [18]. A predominant factor of the process is instantaneous soil moisture status [19,20,21]. Microbial activity in dry soils is limited by water availability, and the lack of oxygen in saturated environments results in the increased importance of anaerobic microorganisms [22]. Soil temperature is also an important driver. Exponential increases in mineralization rates are observed within a range of temperatures in agricultural soils [23]. Modeling of plant-available N within the Leeward Kohala field system demonstrates that N can be expected to be eminently variable both spatially and temporally within the system [6].
Established, long-term research sites within the ahupua’a (traditional land division) of Puanui have made substantial contributions to documenting and understanding the outcomes of traditional management practices. Following European arrival in the 19th century, cultivation of the LKFS was abandoned and largely converted to an open cattle range. In 2008, three permanent experimental gardens were established within Puanui spanning the ecological gradient of the LKFS [2,3]. Previous work [2,3,15,17,24] has used a range of methods to understand and revitalize the ancient cropping system, but affirm that the understanding of how they functioned as dynamic systems is inadequate.
The effects of indigenous Hawaiian soil moisture conservation practices, and especially their functions as driven by climatic factors and their outcomes in facilitating N mineralization, is yet to be explored. Concurrent to soil moisture, soil temperature regulated under mulching regimes is also a critical factor affecting N mineralization that has largely been overlooked. This study contributes to the growing knowledge of the functionality and biogeochemical impacts of traditional cropping methods at Puanui by investigating soil moisture and nitrogen dynamics under indigenous practices, and their benefaction to sweet potato production. It is predicted that indigenous practices increase soil moisture and regulate temperature, leading to an increase in N mineralization rates and sweet potato growth.
2. Materials and Methods
2.1. Field Experimentation
2.1.1. Site Description
In July 2019, we established replicated field experiments within experimental māla (gardens) within the ahupua’a of Puanui (Figure 1) managed by Ulu Mau Puanui—a non-profit organization focused on promoting research, awareness, and education of the cultural and historical prominence of the LKFS. The sites are located on Andisols from the Hawi volcanic series with surface flows aged ~150 ky. Three replicated plots were established. Two adjacent plots were installed in māla mawaena (the middle garden), located 20.150 N, 155.825 W at ~690 m elevation with a mean rainfall of 1476.2 mm/y [25]. The specific plot locations had been under kikuyu grass (Pennisetum clandestinum) fallow for the past 5 years. Site 1 had been slashed and remains overlaid with a weed mat 2 months prior to setting the trial. Site 2 was cleared with fallow vegetation removed preceding field set up. A third plot was established at māla maluna (the upper garden) located at 20.152 N, 155.819 W at ~730 m above sea level with a mean rainfall of 1660.5 mm/y [25]. The site had been fallowed for eight months prior to establishment, with Zoysia grass being the most abundant species cleared by uprooting and removing. Measurements for rainfall, solar radiation, wind speed, air temperature and relative humidity were obtained through Onset weather stations (Onset Computer Corporation; Bourne, MA, USA) situated within each garden.
2.1.2. Treatment Design
A randomized complete block design with three replicates was established at each of the three sites to evaluate three indigenous soil moisture conservation practices relative to two controls. A fourth block was integrated for destructive sampling in the mid-cropping season (60 days after planting). Each treatment was allocated to a soil mound 1 m in diameter, and separated from its nearest neighbor by 1 m. Treatments included (1) bare soil; (2) bare soil with irrigation; (3) rock mulch; (4) sugar cane leaf mulch; and (5) rock mulch over sugar cane mulch (Table 1, Figure 2). For the irrigation treatment, application rates represented the difference between the optimum and actual weekly rainfall levels using real-time rainfall data from weather stations in each plot [26]. Within each treatment mound, the heirloom Hawaiian variety “Lanikeha” was cultivated as a traditional cultivar preserved within the local community. The 40 cm terminal vine cuttings, collected onsite, were prepared by removing all leaves, excluding that from the terminal bud. Rooting was induced under sterile moisture for 3 days. Three slips were planted 10 cm apart and 20 cm deep in each mound.

Table 1.
Summary of treatments applied in the replicated block field experiment.

Figure 2.
Examples of rock mulch (a) and sugar cane mulch (b) treatments.
2.1.3. Soil Moisture and Temperature
At Sites 1 and 3, a single block of replicates was instrumented for continuous monitoring of soil moisture and temperature. The latter served as a representation of Site 2, as both locations were adjacent to each other within the same climatic environments. Onset 10HS Soil Moisture Smart Sensors and Onset 12-Bit Temperature Smart Sensor to measure volumetric soil moisture and soil temperature were installed to a depth of 15cm in the center of the windward side of each mound. Data were recorded in 30-min intervals on a Hobo USB Micro Station Data Logger. Bi-weekly evaluations for all blocks were carried out by inserting probes for one minute in the same location on each mound as outlined above.
2.1.4. Nitrogen Dynamics In Situ
Cumulative inorganic N was evaluated using ion-exchange (IX) resin beads (Rexyn* I-300 H-OH). In every mound, four nylon pouches of 5.0 g of IX beads were installed at a depth of 15 cm during mound construction; a single pouch was harvested every 30 days. Ion-exchange resins were thoroughly rinsed with DI water, shaken in 50 mL of 2M KCl for two hours, and the filtrate analyzed for inorganic N. Every 30 days, small soil cores were collected to a depth of 15 cm. Soils were immediately homogenized, sieved to 2 mm, and split into subsets. For one subset, 2.5 g of each sample was shaken in 50 mL of 2M KCl, then filtered and analyzed for inorganic N. A second subset was oven-dried at 45 °C, then pulverized and encapsulated for Total C and N analysis using a Costech EA Carbon Nitrogen Analyzer.
2.1.5. Sweet Potato Assessment
During plant growth, a SPAD meter was used to measure leaf chlorophyll concentration on a bi-weekly basis from the 7th, 8th and 9th leaf of the primary vine. Destructive sampling was conducted to assess total plant growth at 60 days (one replicate) and 120 days (three replicates) in categories of above ground, root mass, pencil tubers, and storage tuber. At both 60 and 120 days, three terminal vines were collected from each replicate. Tissue samples were dried at 45 °C to a constant weight, then ground and foil-balled for total N and C, and the stable isotope ratios for 13C and 15N, analyzed using a PDZ Europa ANCA-GSL elemental analyzer.
2.2. Incubation Experiment
A 10-week laboratory incubation study was performed using soils collected from the three sites to estimate N mineralization rates under specified soil moisture and temperature conditions. In incubation bags, 120 g of soil (dry equivalent) was maintained at specified soil moistures under different temperatures. A split-plot, randomized complete block design with 5 replicates was used, testing three temperature levels (20, 26 and 30 °C), three moisture levels (10, 30, and 40% gravimetric soil content), and soils from the three plots. From the incubation bags, 2.5 g of soil was sampled at 0, 4, and 10 weeks and analyzed for inorganic nitrogen as described above.
2.3. Data Analysis
RStudio Software (Rstudio, Public Benefit Corporation; Boston, MA, USA) libraries ggplot2 [27] and tseries [28] were used to analyze autocorrelations for continuous soil moisture and temperature data with a maximum lag of 24 h applied. Relationships between time-series data of weather variables to soil moisture and temperature were evaluated using cross-correlation. Linear regression, ANOVA, and fixed effect models were conducted in JMP Pro 15 (SAS Institute; Cary, NC, USA) to assess individual and interactive effects of Treatment, Site and Time for various data. Site was treated as a fixed effect as study interests were specific to the particular soil and climate characteristics. Significant differences in individual effects were further evaluated using the Tukey HSD Test. Significant interactions were evaluated using interaction plots. When appropriate, data transformation was used. Above-ground biomass measurements were initially transformed to near normal distribution using log 10 transformation, and biomass data were converted to standard z scores and used to test for differences between Treatments and Sites. The conversion was necessary for explicit comparison of treatment performance to address the high variation between sites. To assess the overall influence of N dynamics and water stress on sweet potato growth, a multivariate regression analysis was applied to all data collected.
3. Results
3.1. Soil Temperature
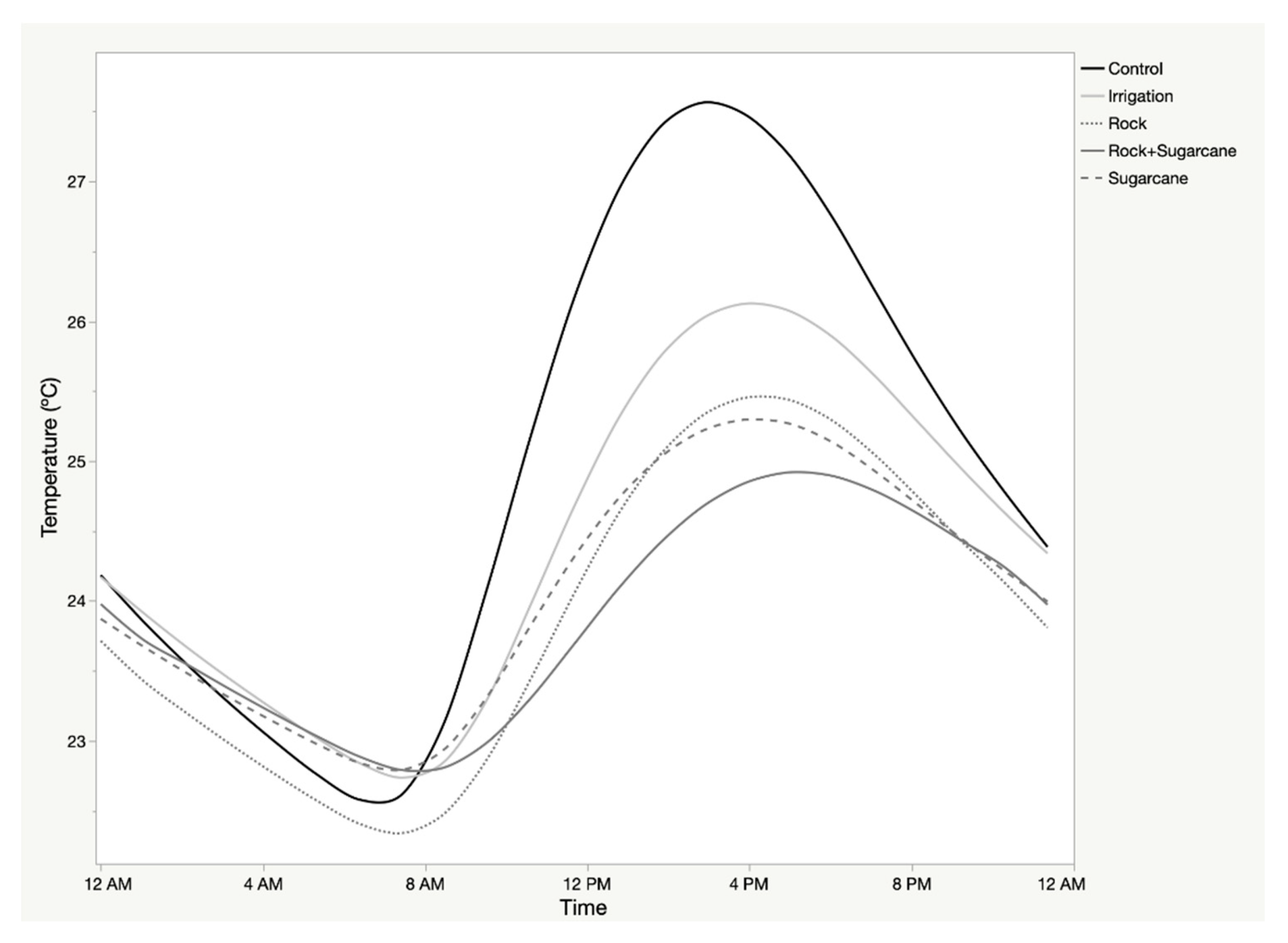
Diurnal soil temperature patterns demonstrated a high degree of autocorrelation on a daily cycle, and a high cross-correlation with air temperature. The 24-h sequences of mean soil temperature demonstrated differentiation in (1) mean soil temperature, (2) the total range of temperature, and (3) the timing of inflection points (Figure 3; Table 2). Rock mulch recorded the lowest mean temperatures and the most moderate range, while the control exhibited the highest mean and largest range. Cross-correlation of air and soil temperature indicated different temporal lags for the different treatments (Table 2). No other weather parameter demonstrated a significant cross-correlation with soil temperature.
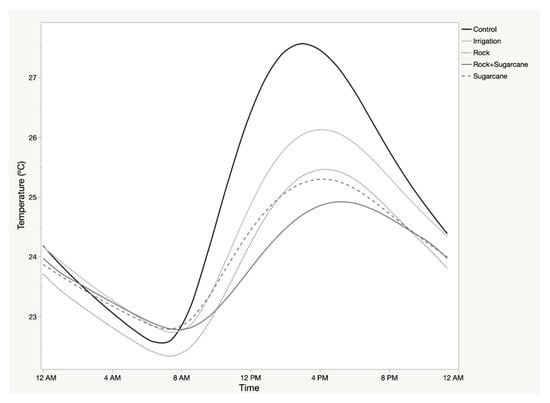
Figure 3.
Mean soil temperature of treatments across Sites during growing season of sweet potato.
Table 2.
Mean parameters of core data by treatments across study sites. Where relevant, connecting letters are used to indicate significant differences as determined by Tukey HSD.
3.2. Soil Moisture
Soil moisture patterns were sporadic, with no “seasonal” patterns in the autocorrelations. The temporal analysis illustrated exponential decay in soil moisture following wetting events. Treatment and sites demonstrated significant differences in soil moisture (Table 2). Soil moisture under the irrigation treatments was observed to be the highest (0.262 m3 m−3) but followed closely by rock-sugar cane mulch (0.250 m3 m−3). The control treatment yielded the lowest mean soil moisture measurement (0.190 m3 m−3). Cross-correlation coefficients of weather parameters were significant but very weak relationships (r < 0.15); association of climatic variables to soil moisture were stronger (r > 0.3) after aggregating time as 24-h increments.
3.3. Soil Nitrogen Dynamics
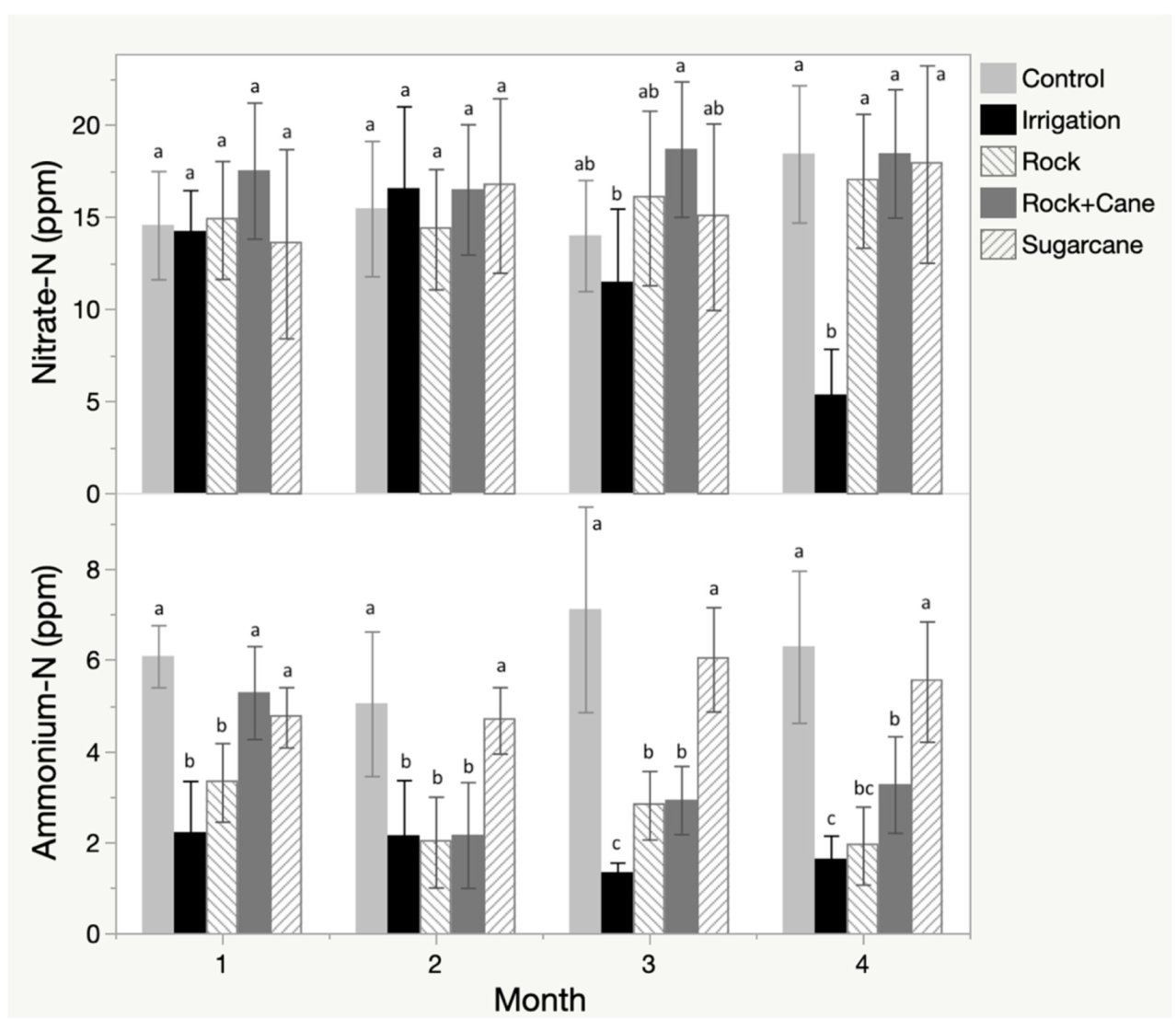
In the field study, nitrate in the soil remained relatively constant over the growing season and statistically indistinguishable between treatments, with the exception of the irrigation treatment that demonstrated a significant decline in nitrate concentrations at 90 and 120 days (Figure 4). Exchange resin demonstrated increased capture of nitrate over time, with the rock mulch, rock-sugar cane mulch, and irrigation treatments accumulating significantly more nitrate than the sugar cane mulch or control treatments (Table 2). Ammonium showed accumulation in the soil over time in the control and sugar cane treatments, but depletion in the remaining three treatments, resulting in significant differences between those two groups (Figure 4). The exchange resins demonstrated increased capture of ammonium over time, with less pronounced and non-significant differences between treatments (Table 2).
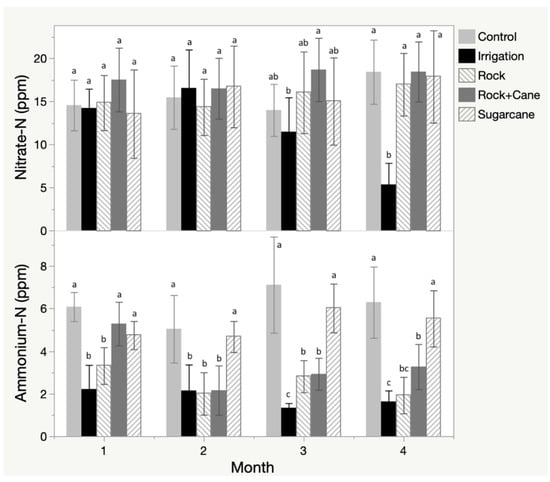
Figure 4.
Nitrate and ammonium accumulation in the field experiment as measured by monthly soil samples, with letters representing significance using a Tukey HSD test.
The evaluation of N mineralization under controlled conditions indicated significant differences between temperature, moisture, and site (Table 3). Rates of nitrate accumulation significantly increased with increased moisture (r2 0.265, p < 0.001), while nitrate increased much more moderately with increased temperature (r2 0.065, p < 0.005). Changes in ammonium were generally small and insignificant, with the only exception being the 10% soil moisture treatment that demonstrated a significant accumulation of ammonium during the incubation period (Table 3).
Table 3.
Results of controlled N mineralization incubation. Vales are in ppm N per week, with connecting letters resulting from Tukey HSD test.
3.4. Sweet Potato Growth and Development
Due to lower statistical power, minimal statistical differences in sweet potato production were detected (Table 2). The irrigation treatment produced the greatest biomass, though it was not significantly different than biomass under rock mulch. There were no differences observed between treatments for storage tuber yields (p > 0.05), and minimal differences were detected in pencil tubers. A significant linear relationship between the above-ground biomass and pencil tubers (p < 0.0001, r = 0.55), and between storage tuber production and pencil tuber production (r = 0.30) suggests that the significant differences in above ground biomass may have eventually translated to differences in tuber production. Regression analysis of plant N concentrations and production of above-ground biomass indicated a moderate and positive relationship (r = 0.61, p = 0.02). The linear relationship between 13C stable isotope and above-ground biomass showed a significant positive association (p < 0.001, r2 = 0.216). A fixed model effects analysis showed no significant effects of experimental treatments on plant N concentrations nor foliar 13C.
4. Discussion
Indigenous practices, such as rock mulch, have a substantial effect on soil moisture and temperature that further affect nitrogen dynamics and production in agricultural systems. Such practices were employed broadly across various ecosystems to increase production and resilience in ancient agriculture. The specific primary and secondary effects of these treatments have rarely been investigated and are dependent on the specific limitations of each habitat. We examined the direct effects of mulching practices on soil temperature and moisture, along with the secondary effects of nitrogen availability.
The productivity of the Puanui dryland field system is highly dependent on the variable rainfall. Measured rainfall during the trial was 40% less than expected for an average year [25]. Irrigation was observed to enhance the production of above-ground biomass, but comparable increases were observed from mulching with rock. Although sweet potato is generally drought tolerant, water stress still results in growth reduction [26,29,30,31]. Low production in the remaining treatments indicated inadequacy to conserve soil moisture. Within our experiment, administering scheduled irrigation allowed for increased soil moisture in the absence of rainfall, however, temporal patterns demonstrated that the effects of irrigation on soil moisture declined rapidly after application, and between weekly applications of irrigation, soil moisture would drop below levels of other treatments that preserved soil moisture more effectively. Although rainfall was perceived as the primary driver of soil moisture through longer time scales; alternate weather variables, such as wind speed and humidity, were influential on the dynamics [32], hence comparable correlations relative to rainfall were distinct.
The Indigenous practice of rock mulching was able to conserve more soil moisture than under sugar cane mulch. We believe that, under the sugar cane mulch, the percolation of moisture from precipitation is impeded from reaching the soil surface, in particular the light-misty precipitation that is characteristic of the study location [15]. Conversely, the non-porous rock mulch both captured the light mist by providing an accretion surface and allowed that deposited moisture to efficiently reach the soil.
Though increases in soil moisture generally enhance N mineralization, anaerobic conditions can limit soil carbon mineralization through energetic and enzymatic constraints on microbial activity, even under short durations (hours-days) [33,34,35]. Volcanic soils generally have a high water-holding capacity that can contribute to reduced C respiration [34,36,37]. Reduced decomposition of organic matter consequently hinders N mineralization and available plant N. Low measures of inorganic N under the irrigation treatment may be a result of suppressed C mineralization due to excessive moisture supplement applications [34]. Alternatively, the low measures may have been caused by increased transport and leeching of inorganic N.
An increase in temperatures tends to enhance net mineralized N due to increased microbial activity [38]—a trend observed in our ex situ incubation experiment conducted with local soils. Microbial activity for N mineralization is potentially reduced under the mulch treatments, which generally reduced temperature below the optimum temperatures 35 °C [39,40]. However, the diurnal patterns must also be considered as treatments with lower daytime temperatures typically exhibited increased nighttime temperatures, potentially increasing nocturnal microbial activity.
Different mulch types alter soil temperature in different ways and can be associated with the thermophysical properties of these materials [41]. Low thermal conductivity and high specific heat capacity of basalt (0.84 kJ kg−1 °C) [42] function as a more efficient thermal insulator compared to dry leaves, with a higher specific heat capacity (1.17–5.17 kJ kg−1 °C) [43]. Interactive effects between soil moisture and temperature can be seen in that the irrigation treatment results show a slightly lower average temperature compared to the control due to water absorbing and regulating heat (specific heat at 15 °C–4.187 kJ kg−1 °C), suggesting feedback mechanisms of mulch regulating soil temperature directly through heat absorption and indirectly by altering soil moisture.
Treatments exhibiting the highest measures of cumulative inorganic N were appreciated to be the same treatments measuring high soil moisture content. Movement of NO3− through soil is controlled by convection flow with moving soil solution, as well as diffusion within the soil solution; suggesting improved soil moisture facilitated infiltration of the soil solution through the resin pouches [44]. Limiting soil moisture conditions under the control and sugar cane mulch likely inhibited both mineralization of organic N and restricted mass flow of inorganic N, consequently hindering the potential amassing of inorganic N ions. In contrast, measures of inorganic N evaluated from sampling soil are governed by the N cycle and include processes of ammonification, immobilization, nitrification, adsorption, volatilization, denitrification and leaching loss, other than mineralization [45].
Asserting that soil productivity to be high in regions of intermediate rainfall [16,46,47,48], measures of instantaneous inorganic soil N in the middle māla were higher than the upper māla. However, it is possible enhanced convectional flow in upper māla, resulting from improved rainfall conditions, resulted in similar measures of cumulative inorganic soil N to the middle māla. Despite distinct variation of measures of inorganic N, total plant N was comparable between treatments and sites, potentially due to the low statistical power of the analysis. However, appreciating the low input farming system and environmental factors, the crop’s efficiency to use N can be enhanced through strategized nutrient management. A strong, positive correlation between cumulative inorganic N to SPAD measurements indicates that plant productivity can be improved through increasing N availability [49]. Understanding crop nutrient needs, N rates and sources, placement methods, the timing of application, irrigation management, and residue management can affect N uptake and its efficiency of use.
Mean tuber production across all mulching treatments was observed to be ~250 g m2. Previous experimental harvests using similar mulching treatments in consistent locations and planting dates had produced ~62.9% more [3]. Though high aboveground biomass production under the irrigation and rock mulch treatment should have translated to elevated tuber production, there was a lack of yield difference in the storage tubers compared to other treatments. Timing and frequency of moisture application are critical to sweet potato tuberization. Tuber development is encouraged by two mm of water per day and the maintenance of available soil moisture at 25% during the early parts of the growing season; available soil moisture exceeding 50% though does not encourage tuber production [50,51]. Increased pencil root development is a result of deficit soil moisture during the first 2 weeks of planting, during which time the development of the number of storage roots is highly influenced by soil moisture [52,53]. Low storage tuber yields and excessive pencil tubers across sites were also indicative of water stress across all sites and treatments. While unsuccessful in substantial tuberization, the results suggest that the differences in moisture and nitrogen would have translated to production if given more time for gestation.
Plant response to environmental stress, particularly heat and drought, can be reflected in carbon composition [54]. Variations in 13C stable isotope can concede response of plants to local environmental conditions [55,56], with moisture stress tending to decrease δ13C abundance [57,58]. We saw decreased biomass with decreasing δ13C, suggesting that moisture was directly limiting the growth within the study system. Despite total rainfall measuring 21.0 mm and 61.8 mm in the first 30 days of the middle and upper māla respectively, ~88.0% of these days had less than 2 mm of precipitation.
Agronomic characteristics of the lanikeha variety may have contributed to the exceptionally low yield. It is a late-maturity cultivar, harvestable at 8 months. Tuber bulking (>21 weeks) increases during the middle or towards the end of the growing cycle; though favorable agroclimatic conditions produce high yields in the early periods [51]. Hence, tuber bulking patterns can be highly influenced by genotype by environment interaction [59]. The crop was however harvested at 5 months, as this was admissible time to conduct the field research and avoid crop loss to rats, pheasants and feral pigs. Additionally, above-ground biomass production of the lanikeha variety is characterized to be extensive and rampant (K. Marshall, personal communication, 1 July 2019), and produces profusely pencil and fibrous roots. Though the source-sink relationships of the Hawaiian heirloom sweet potato varieties have not been researched, high above-ground biomass production in sweet potato can occur at the expense of storage root development, and can even be a desirable trait in cultures that eat the young foliage as cooked greens.
5. Conclusions
Traditional moisture conservation techniques are shown to have the potential to enhance soil moisture, regulate temperature and increase soil N availability. Regardless of soil management practices, interaction responses between sites and treatments affirm the productivity of Puanui to be immensely influenced by temporal and spatial climatic variability with intermittent rainfall, which supports previous research. The variable effect of different practices as they interact with the weather (which varies in both space and time) suggests that any maximization process would involve adaptive practices rather than a prescriptive approach. The differences across sites are substantial, and any consideration of the potential productivity of these ancient Hawaiian rainfed systems would do well to take a spatially explicit approach, as blanket assumptions about yields, labor, and other agricultural inputs are highly dependent on the specific site. The traditional systematic approach of sustaining crop production presents an avenue to reshaping Hawaiian conventional farming practices to accommodate environmental variability. An economic evaluation under the settings of an enterprise can determine the complexity of the cropping system as well as the commercial sustainability of indigenous practices in a changing environment.
Author Contributions
Conceptualization, W.C.L.S., N.K.L., T.R. and M.B.K.; methodology, W.C.L.S. and N.K.L.; software, W.C.L.S., M.B.K. and N.K.L.; formal analysis, W.C.L.S., M.B.K. and N.K.L.; investigation, W.C.L.S.; resources, N.K.L.; data curation, W.C.L.S. and N.K.L.; writing—original draft preparation, W.C.L.S.; writing—review and editing, W.C.L.S., N.K.L., T.R. and M.B.K.; visualization, W.C.L.S. and N.K.L.; supervision, N.K.L.; project administration, N.K.L.; funding acquisition, W.C.L.S. and N.K.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work is supported by the USDA National Institute of Food and Agriculture HATCH (project number 8035-H) managed by the College of Tropical Agriculture and Human Resources.
Data Availability Statement
Daily weather data for the experimental gardens can be accessed here: http://www.ulumaupuanui.org/weather.html, accessed on 7 November 2021. Soil and plant data are held in multiple backups at the Indigenous Cropping Systems Laboratory at the University of Hawai’i at Mānoa and are available upon request.
Acknowledgments
We would like to acknowledge Ulu Mau Puanui and their executive director, Kehaulani Marshall, for the access and physical and logistic support on this project, and the East-West Center for their support of the lead author’s graduate education.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ladefoged, T.N.; Kirch, P.V.; Gon, S.M., III; Chadwick, O.A.; Hartshorn, A.S.; Vitousek, P.M. Opportunities and constraints for intensive agriculture in the Hawaiian archipelago prior to European contact. J. Archaeol. Sci. 2009, 36, 2374–2383. [Google Scholar] [CrossRef]
- Kagawa, A.K.; Vitousek, P.M. The Ahupua ‘a of Puanui: A Resource for Understanding Hawaiian Rain-Fed Agriculture1. Pac. Sci. 2012, 66, 161–172. [Google Scholar] [CrossRef]
- Marshall, K.; Koseff, C.; Roberts, A.L.; Lindsey, A.; Kagawa-Viviani, A.K.; Lincoln, N.K.; Vitousek, P.M. Restoring people and productivity to Puanui: Challenges and opportunities in the restoration of an intensive rain-fed Hawaiian field system. Ecol. Soc. 2017, 22, 23. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Ladefoged, T.N.; Kirch, P.V.; Hartshorn, A.S.; Graves, M.W.; Hotchkiss, S.C.; Tuljapurkar, S.; Chadwick, O.A. Soils, agriculture, and society in precontact Hawaii. Science 2004, 304, 1665–1669. [Google Scholar] [CrossRef] [PubMed]
- Kagawa-Viviani, A.K.; Lincoln, N.K.; Quintus, S.; Lucas, M.P.; Giambelluca, T.W. Spatial patterns of seasonal crop production suggest coordination within and across dryland agricultural systems of Hawaii Island. Ecol. Soc. 2018, 23, 20. [Google Scholar] [CrossRef]
- Lee, C.T.; Tuljapurkar, S.; Vitousek, P.M. Risky business: Temporal and spatial variation in preindustrial dryland agriculture. Hum. Ecol. 2006, 34, 739–763. [Google Scholar] [CrossRef] [Green Version]
- Coil, J.; Kirch, P.V. An Ipomoean landscape: Archaeology and the sweet potato in Kahikinui, Maui, Hawaiian Islands. Sweet Potato Pac. Reappraisal Ocean. Monogr. 2005, 56, 71–84. [Google Scholar]
- Yen, D.E. The Sweet Potato and Oceania; An Essay in Ethnobotany; Bishop Museum Press: Honolulu, HI, USA, 1974. [Google Scholar]
- Lincoln, N.K.; Vitousek, P. Indigenous Polynesian agriculture in Hawaii. In Oxford Research Encyclopedia of Environmental Science; Oxford University Press: Oxford, UK, 2017. [Google Scholar]
- DiNapoli, R.J.; Morrison, A.E. A spatiotemporal model of risk and uncertainty for Hawaiian dryland agriculture and its implications for ahupua’a community formation. J. Archaeol. Sci. Rep. 2017, 15, 109–119. [Google Scholar] [CrossRef]
- Kamakau, S.M. The Works of the People of Old: Na Hana a ka Po’e Kahiko; Barrère, D.B., Ed.; Bishop Museum Press: Honolulu, HI, USA, 1976. [Google Scholar]
- Malo, D. Hawaiian Antiquities (Moolelo Hawaii); Hawaiian Gazette Company: Honolulu, HI, USA, 1903; Volume 2. [Google Scholar]
- Winter, K.; Lincoln, N.; Berkes, F.; Alegado, R.; Kurashima, N.; Frank, K.; Pascua, P.; Rii, Y.; Reppun, F.; Knapp, I. Ecomimicry in Indigenous resource management: Optimizing ecosystem services to achieve resource abundance, with examples from Hawaii. Ecol. Soc. 2020, 25, 26. [Google Scholar] [CrossRef]
- Handy, E.S.C.; Handy, E.G.; Pukui, M.K. Native Planters in Old Hawaii: Their Life, Lore, and Environment; Bishop Museum Press: Honolulu, HI, USA, 1972; Volume 233. [Google Scholar]
- Lincoln, N.K.; Kagawa-Viviani, A.; Marshall, K.; Vitousek, P.M. Observations of Sugarcane and Knowledge Specificity in Traditional Hawaiian Cropping Systems; Nova Science Publishers: Hauppauge, NY, USA, 2017. [Google Scholar]
- Chadwick, O.A.; Gavenda, R.T.; Kelly, E.F.; Ziegler, K.; Olson, C.G.; Elliott, W.C.; Hendricks, D.M. The impact of climate on the biogeochemical functioning of volcanic soils. Chem. Geol. 2003, 202, 195–223. [Google Scholar] [CrossRef]
- Lincoln, N.K.; Vitousek, P. Nitrogen fixation during decomposition of sugarcane (Saccharum officinarum) is an important contribution to nutrient supply in traditional dryland agricultural systems of Hawai’i. Int. J. Agric. Sustain. 2016, 14, 214–230. [Google Scholar] [CrossRef]
- Moreno-Vivián, C.; Cabello, P.; Matínez-Luque, M.; Blasco, R.; Castillo, F. Prokaryotic nitrate reduction: Molecular properties and functional distinction among bacterial nitrate reductases. J. Bacteriol. 1999, 181, 6573–6584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cook, F.J.; Orchard, V.A. Relationships between soil respiration and soil moisture. Soil Biol. Biochem. 2008, 40, 1013–1018. [Google Scholar] [CrossRef]
- Dessureault-Rompré, J.; Zebarth, B.J.; Georgallas, A.; Burton, D.L.; Grant, C.A. A biophysical water function to predict the response of soil nitrogen mineralization to soil water content. Geoderma 2011, 167, 214–227. [Google Scholar] [CrossRef]
- Liu, W.; Zhang, Z.H.E.; Wan, S. Predominant role of water in regulating soil and microbial respiration and their responses to climate change in a semiarid grassland. Glob. Change Biol. 2009, 15, 184–195. [Google Scholar] [CrossRef]
- Deenik, J. Nitrogen Mineralization Potential in Important Agricultural Soils of Hawai’i; University of Hawaii: Honolulu, HI, USA, 2006. [Google Scholar]
- Dessureault-Rompré, J.; Zebarth, B.J.; Georgallas, A.; Burton, D.L.; Grant, C.A.; Drury, C.F. Temperature dependence of soil nitrogen mineralization rate: Comparison of mathematical models, reference temperatures and origin of the soils. Geoderma 2010, 157, 97–108. [Google Scholar] [CrossRef]
- Lincoln, N.K.; Rossen, J.; Vitousek, P.; Kahoonei, J.; Shapiro, D.; Kalawe, K.; Pai, M.; Marshall, K.; Meheula, K. Restoration of ‘āina malo ‘o on Hawai‘i Island: Expanding Biocultural Relationships. Sustainability 2018, 10, 3985. [Google Scholar] [CrossRef] [Green Version]
- Giambelluca, T.W.; Chen, Q.; Frazier, A.G.; Price, J.P.; Chen, Y.-L.; Chu, P.-S.; Eischeid, J.K.; Delparte, D.M. Online rainfall atlas of Hawai‘i. Bull. Am. Meteorol. Soc. 2013, 94, 313–316. [Google Scholar] [CrossRef]
- Valenzuela, H.; Fukuda, S.; Arakaki, A. Sweetpotato Production Guides for Hawai’i; University of Hawaii: Honolulu, HI, USA, 1994. [Google Scholar]
- Wickham, H. Elegant graphics for data analysis. Media 2009, 35, 10–1007. [Google Scholar]
- Trapletti, A.; Hornik, K.; LeBaron, B. Tseries: Time Series Analysis and Computational Finance. R Package Version 0.10-47. 2019. Available online: https://cran.r-project.org/web/packages/tseries/index.html (accessed on 8 November 2021).
- Constantin, R.J.; Hernandez, T.P.; Jones, L.G. Effects of Irrigation and Nitrogen Fertilization on Quality of Sweet Potatoes; World Vegetable Center: Tainan City, Taiwan, 1974. [Google Scholar]
- Ghuman, B.S.; Lal, R. Growth and plant-water relations of sweet potato (Ipomea batata) as affected by soil moisture regimes. Plant Soil 1983, 70, 95–106. [Google Scholar] [CrossRef]
- Saraswati, P. Physiological and Growth Responses of Selected Sweet Potato (Ipomoea batatas (L.) Lam.) Cultivars to Water Stress. Ph.D. Thesis, James Cook University, Townsville, Australia, 2007. [Google Scholar]
- Ravi, S.; D’Odorico, P.; Over, T.M.; Zobeck, T.M. On the effect of air humidity on soil susceptibility to wind erosion: The case of air-dry soils. Geophys. Res. Lett. 2004, 31, 1–4. [Google Scholar] [CrossRef]
- Freeman, C.; Ostle, N.; Kang, H. An enzymic’latch’on a global carbon store. Nature 2001, 409, 149. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Hall, S.J. Elevated moisture stimulates carbon loss from mineral soils by releasing protected organic matter. Nat. Commun. 2017, 8, 1774. [Google Scholar] [CrossRef] [PubMed]
- Linn, D.M.; Doran, J.W. Effect of water-filled pore space on carbon dioxide and nitrous oxide production in tilled and nontilled soils. Soil Sci. Soc. Am. J. 1984, 48, 1267–1272. [Google Scholar] [CrossRef] [Green Version]
- Burke, I.C.; Lauenroth, W.K.; Parton, W.J. Regional and temporal variation in net primary production and nitrogen mineralization in grasslands. Ecology 1997, 78, 1330–1340. [Google Scholar] [CrossRef]
- Parton, W.J.; Schimel, D.S.; Cole, C.V.; Ojima, D.S. Analysis of factors controlling soil organic matter levels in Great Plains grasslands. Soil Sci. Soc. Am. J. 1987, 51, 1173–1179. [Google Scholar] [CrossRef]
- Lei, L.; McDonald, L.M. Soil moisture and temperature effects on nitrogen mineralization in a high tunnel farming system. Commun. Soil Sci. Plant Anal. 2019, 50, 2140–2150. [Google Scholar] [CrossRef]
- Hu, W.; Tian, S.; Di, Q.; Liu, J.; Zhang, S. Nitrogen mineralization simulation dynamic in tobacco soil. J. Soil Sci. Plant Nutr. 2018, 18, 448–465. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.J.; Smith, C.J.; Chen, D. Predicting soil nitrogen mineralization dynamics with a modified double exponential model. Soil Sci. Soc. Am. J. 2004, 68, 1256–1265. [Google Scholar] [CrossRef]
- Van Donk, S.J.; Tollner, E.W. Apparent thermal conductivity of mulch materials exposed to forced convection. Trans. ASAE 2000, 43, 1117. [Google Scholar] [CrossRef]
- Zimbelman, J.R. The role of porosity in thermal inertia variations on basaltic lavas. Icarus 1986, 68, 366–369. [Google Scholar] [CrossRef]
- Jayalakshmy, M.S.; Philip, J. Thermophysical properties of plant leaves and their influence on the environment temperature. Int. J. Thermophys. 2010, 31, 2295–2304. [Google Scholar] [CrossRef]
- Jury, W.A.; Nielsen, D.R. Nitrate transport and leaching mechanisms. In Developments in Agricultural and Managed Forest Ecology; Elsevier: Amsterdam, The Netherlands, 1989; Volume 21, pp. 139–157. [Google Scholar]
- Brady, N.C.; Weil, R.R.; Weil, R.R. The Nature and Properties of Soils; Prentice Hall: Upper Saddle River, NJ, USA, 2008; Volume 13. [Google Scholar]
- Vitousek, P.M.; Chadwick, O.A. Pedogenic thresholds and soil process domains in basalt-derived soils. Ecosystems 2013, 16, 1379–1395. [Google Scholar] [CrossRef] [Green Version]
- Vitousek, P.M.; Chadwick, O.A.; Hotchkiss, S.C.; Ladefoged, T.N.; Stevenson, C.M. Farming the rock: A biogeochemical perspective on intensive agriculture in Polynesia. J. Pac. Archaeol. 2014, 5, 51–61. [Google Scholar]
- Von Sperber, C.; Chadwick, O.A.; Casciotti, K.L.; Peay, K.G.; Francis, C.A.; Kim, A.E.; Vitousek, P.M. Controls of nitrogen cycling evaluated along a well-characterized climate gradient. Ecology 2017, 98, 1117–1129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, A.M.; Assunção, N.S.; Ribeiro, N.P.; Gazola, B.; da Silva, R.M. Nutrient uptake and removal by sweet potato fertilized with green manure and nitrogen on sandy soil. Rev. Bras. Ciênc. Solo 2020, 44, 1–25. [Google Scholar] [CrossRef]
- Nedunchezhiyan, M.; Gangadharan, B.; Susantha, K. Sweet Potato Agronomy. Fruit Veg. Cereal Sci. Biotechnol. 2012, 6, 1–10. [Google Scholar]
- Ravi, V.; Saravanan, R. Physiology of Sweetpotato. Fruit Veg. Cereal Sci. Biotechnol. 2012, 6, 17–29. [Google Scholar]
- Meyers, S.L.; Arancibia, R.A.; Shankle, M.W.; Main, J.; Gajanayake, B.; Reddy, K.R. Sweet Potato Storage Root Initiation; Extension Service; Mississippi State University: Starkville, MS, USA, 2014. [Google Scholar]
- Smith, T.P.; Villordon, A.O. Nitrogen Management in Louisiana Sweet Potatoes; Lousiana State Univeristy Agricultural Center: Baton Rouge, LA, USA, 2009. [Google Scholar]
- Spangenberg, J.E.; Schweizer, M.; Zufferey, V. Shifts in carbon and nitrogen stable isotope composition and epicuticular lipids in leaves reflect early water-stress in vineyards. Sci. Total Environ. 2020, 739, 140343. [Google Scholar] [CrossRef] [PubMed]
- Brugnoli, E.; Hubick, K.T.; von Caemmerer, S.; Wong, S.C.; Farquhar, G.D. Correlation between the carbon isotope discrimination in leaf starch and sugars of C3 plants and the ratio of intercellular and atmospheric partial pressures of carbon dioxide. Plant Physiol. 1988, 88, 1418–1424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cernusak, L.A.; Ubierna, N.; Winter, K.; Holtum, J.A.; Marshall, J.D.; Farquhar, G.D. Environmental and physiological determinants of carbon isotope discrimination in terrestrial plants. New Phytol. 2013, 200, 950–965. [Google Scholar] [CrossRef] [PubMed]
- Farquhar, G.D.; Ehleringer, J.R.; Hubick, K.T. Carbon isotope discrimination and photosynthesis. Annu. Rev. Plant Biol. 1989, 40, 503–537. [Google Scholar] [CrossRef]
- Robinson, D.; Handley, L.L.; Scrimgeour, C.M.; Gordon, D.C.; Forster, B.P.; Ellis, R.P. Using stable isotope natural abundances (δ 15 N and δ 13 C) to integrate the stress responses of wild barley (Hordeum spontaneum C. Koch.) genotypes. J. Exp. Bot. 2000, 51, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Ngailo, S.; Shimelis, H.; Sibiya, J.; Mtunda, K.; Mashilo, J. Genotype-by-environment interaction of newly-developed sweet potato genotypes for storage root yield, yield-related traits and resistance to sweet potato virus disease. Heliyon 2019, 5, e01448. [Google Scholar] [CrossRef] [PubMed] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).